Introduction
Evodiamine (EVO) is a type of quinazoline carboline
alkaloid that is a traditional Chinese medicine isolated from Wu
Zhu Yu (Evodia rutaecarpa). EVO has been shown to improve
cognitive function, have anti-inflammatory properties and tackle
circulatory failure (1). In
addition, EVO may have vasodilatory and cardiotonic effects
(2). Accumulating evidence has
demonstrated the antitumor effects of EVO, including on
gastrointestinal (3),
genitourinary tract (4), breast
(5), prostate (6) and colon (7) cancer. Although the precise mechanisms
remain to be elucidated, induction of apoptosis is believed to be
one of the major mechanisms of action for evodiamine against cancer
cells.
Lung cancer is one of the leading causes of
cancer-associated deaths worldwide (8). When lung cancer is diagnosed, it is
frequently in the late stage and the 5-year survival rate is
discouraging. Although incremental improvements in the survival
rate and quality of life have been achieved in other common
malignancies, no efficient therapeutic strategies for lung cancer
have been developed (9). In recent
years, the existence of cancer stem-like cells (CSCs) in different
types of cancer has been recognized and accepted. The CSC
hypothesis demonstrates the existence of a population of rare,
stem-like tumor cells maintaining stemness, exerting self-renewal
capacity and undergoing asymmetric division (10-12).
CSCs share molecular features with embryonic stem cells, including
CD133(13), Nanog (14) and Oct4(15), which are considered CSC hallmarks.
CSCs have been isolated from a number of forms of human cancer,
including lung cancer (13,16).
Considering the central role of CSCs in tumorigenesis, inducing
malignant behavior and chemoresistance, CSCs might be considered a
therapeutic target to achieve effective cancer treatment (17). Although the effects of EVO on tumor
cells have been well studied, the role of EVO in regulating the
malignant behaviors of CSCs remains to be elucidated.
The epithelial-mesenchymal transition (EMT) is a
complex series of morphological changes, including the loss of
epithelial characteristics and the acquisition of a mesenchymal
phenotype. In solid cancers, the EMT tightly regulates the
processes of metastasis; it is responsible for survival in the
circulation and seeding at secondary sites (18). It has been reported that activation
of the EMT via either overexpression of the EMT hallmarks or
treatment with TGF-β confers a number of the properties of CSCs on
otherwise epithelial carcinoma cells (19,20),
indicating that activation of the EMT process is closely related to
entrance into the CSC state in several different types of cancer
cells.
The aims of the current study were to evaluate the
effects of EVO on the physiological processes of the non-small-cell
lung cancer cell line A549 and CSCs derived from A549. It then
focused on the effects of EVO on the maintenance of stemness and
the EMT process in CSCs to evaluate the potential role of CSCs as a
therapeutic target of non-small-cell lung cancer cells and
demonstrates that EVO may a promising natural compound targeting to
CSCs.
Materials and methods
Cell culture and CSCs enrichment
The human non-small cell lung cancer cell line A549
was purchased from ATCC and was cultured in Dulbecco's Modified
Eagle Medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (FBS; Thermo Fisher
Scientific, Inc.) at 37˚C in 5% CO2. In order to enrich
CSCs from A549 cells, 1x106 A549 cells were cultured in
DMEM/F12 (Gibco; Thermo Fisher Scientific, Inc.) supplemented with
20 ng/ml of epidermal growth factor (EGF; PeproTech, Inc), 10 ng/ml
of basic fibroblast growth factor (bFGF; PeproTech, Inc) and 2%
B-27 (Thermo Fisher Scientific, Inc.). Every four days, medium was
half-refreshed. At day 10 and 20, cells were imaged under a X71
(U-RFL-T) fluorescence microscope (Olympus Corporation) and stored
in liquid nitrogen.
EVO treatment
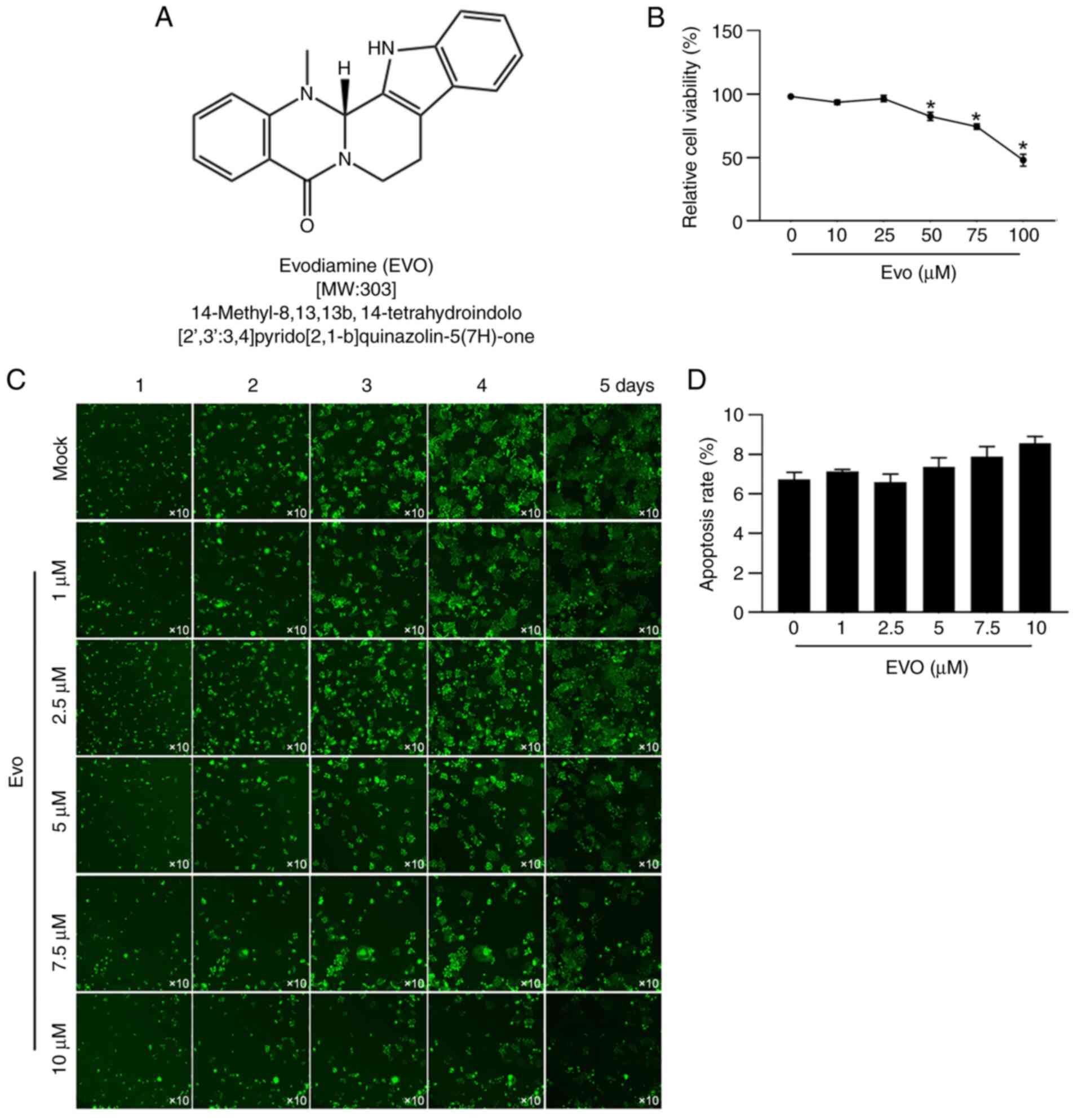
EVO (Fig. 1A,
MilliporeSigma) was with DMSO to a final concentration of 1 mmol/l.
A549 cells or CSCs were incubated in DMEM supplemented with 10% FBS
and without or without 1, 2.5, 5, 7.5 or 10 µmol/l of EVO at 37˚C.
For cell proliferation assay, 1, 2.5, 5, 7.5 or 10 µmol/l of EVO
exposure lasted for 24 h at 37˚C. For CCK-8 assay, cells were
co-cultured with EVO for 24, 48 and 72 h at 37˚C. The intralipid
treated cells were considered as negative control.
Cell cycle assay
The cell cycle distribution of 1x106
cells was checked using propidium iodide (PI; MilliporeSigma)
staining on a flow cytometer. The cells were collected by
centrifugation at 400 x g for 5 min at 37˚C. The cell pellet was
collected and fixed with 4% paraformaldehyde and stained with 5
µg/ml PI for 10 min in darkness at room temperature. Then cells
were washed three times with PBS and PI absorbance was determined
by FACS on a flow cytometry (FACSCalibur, Becton, Dickinson and
Company). Data was analyzed using FlowJo software (version 8.7.1;
Tree Star Inc.).
EdU staining
For EdU staining, EdU-labeling reagent (Thermo
Fisher Scientific, Inc.) was added and incubated with cells for 4
h. Then the medium was removed and cells were washed with PBS for
three times at 37˚C. Cells were fixed with 4% paraformaldehyde for
15 min at room temperature. Clik-iT kit (Thermo Fisher Scientific,
Inc.) was employed for EdU detection following the manufacturer's
instructions. Then the cells were counterstained with 5 µg/ml DAPI
at room temperature for 5 min. Images from five fields of view were
randomly chosen and captured using a X71 (U-RFL-T) fluorescence
microscope (Olympus Corporation) at x40 magnification and analyzed
using ImageJ software (version 1.2; National Institutes of
Health).
Transwell assay
Cell migration and invasion was evaluated using an
8-mm pore size Transwell system (Costar; Corning, Inc.) without or
with Matrigel (BD Biosciences), which was pre-coated at 37˚C for 2
h. Briefly, cells were dissociated into single cells and
resuspended in DMEM medium at a density of 1x105
cells/ml. The top chamber of the Transwell was loaded with 200 µl
cell suspension and 800 µl DMEM medium supplemented with 10% FBS
was added to each lower chamber. Following incubation in the
incubator for 24 h at 37˚C, the cells remain on the upper surface
of upper chamber were removed and the cells on the lower surface of
upper chamber were fixed in 4% paraformaldehyde at room temperature
for 10 min and subsequently stained with 0.25% crystal violet
(MilliporeSigma) at room temperature for 10 min followed by three
washes with PBS. Images of the stained cells from five random views
were captured under a X71 (U-RFL-T) fluorescence microscope
(Olympus Corporation) at x20 magnification.
Western blotting
Cells were collected by centrifugation at 800 x g,
4˚C for 10 min and washed with PBS for three times. Total protein
was extracted using NP-40 lysis buffer (Beyotime Institute of
Biotechnology), according to the manufacturer's instructions. The
concentration of total protein was measured using a BCA detection
kit (MilliporeSigma). Total protein (20 µg) was fractionated via
10% sodium dodecyl sulfate polyacrylamide gel (SDS-PAGE, Beyotime
Institute of Biotechnology) electrophoresis, followed by blot
transfer onto a polyvinylidene fluoride membrane (PVDF, Thermo
Fisher Scientific, Inc.). Following transfer, the PVDF membrane was
blocked in 5% skimmed milk at room temperature for 30 min. Then the
PVDF membrane was incubated with the following primary antibodies
at 4˚C overnight: Rabbit anti-human polyclonal antibody against
CD24 (1:1,000; cat. no. ab179821); rabbit anti-human polyclonal
antibody against CD44 (1:1,000; cat. no. ab157107); rabbit
anti-human polyclonal antibody against CD133 (1:1,000; cat. no.
ab19898); rabbit anti-human polyclonal antibody against Oct4
(1:1,000; cat. no. ab181557); rabbit anti-human polyclonal antibody
against Nanog (1:1,000; cat. no. ab21624); rabbit anti-human
polyclonal antibody against β-actin (1:5,000; cat. no. ab8227);
rabbit anti-human polyclonal antibody against E-cadherin (1:1,000;
cat. no. ab40772); rabbit anti-human polyclonal antibody against
Vimentin (1:1,000; cat. no. ab92547); rabbit anti-human polyclonal
antibody against Slug (1:1,000; cat. no. ab27568); and rabbit
anti-human polyclonal antibody against Snail (1:1,000; cat. no.
ab82846). All antibodies were purchased from Abcam. Following three
washes with PBS supplemented with 0.1% Tween-20, the membranes were
subsequently incubated with horseradish peroxidase-linked goat
anti-rabbit IgG (1:5,000, cat. no. ab7090) at room temperature for
2 h. Enhanced chemiluminescence solution (Thermo Fisher Scientific,
Inc.) was added for luminescent image development. The amount of
β-actin was considered as a reference. Blots were quantitatively
analyzed using Image J software (version 1.8.0.172).
RNA extraction and reverse
transcription-quantitative (RT-q) PCR
TRIzol® reagent (Thermo Fisher
Scientific, Inc.) was purchased for RNA extraction. In brief, cDNA
was synthesized by using the Reverse Transcriptional kit (Guangzhou
RiboBio Co., Ltd.) from 1 µg of total RNA. The RT reaction was
carried under the following conditions: 10 min at 25˚C, 60 min at
42˚C and 10 min at 85˚C in a total 20 µl of reaction mixture.
RT-qPCR was performed in an ABI7500 (Applied Biosystems; Thermo
Fisher Scientific, Inc.). The reaction mixture consists of 2 µl of
forward and reverse primers in 10 µl of SYBR Green Master Mix
(Thermo Fisher Scientific, Inc.) to a total volume of 20 µl. The
RT-qPCR cycle conditions were 2 min at 50˚C, 10 min at 95˚C,
followed by 40 cycles of 15 sec at 95˚C and 1 min at 60˚C. The
primer sequences were identified through PrimerBank (https://pga.mgh.harvard.edu/primerbank/)
and were: E-cadherin Forward: 5'-CGAGAGCTACACGTTCACGG-3',
E-cadherin Reverse: 5'-GGGTGTCGAGGGAAAAATAGG-3'; Vimentin Forward:
5'-GACGCCATCAACACCGAGTT-3', Vimentin Reverse:
5'-CTTTGTCGTTGGTTAGCTGGT-3'; Slug Forward:
5'-CGAACTGGACACACATACAGTG-3', Slug Reverse:
5'-CTGAGGATCTCTGGTTGTGGT-3'; Snail Forward:
5'-TCGGAAGCCTAACTACAGCGA-3', Snail Reverse:
5'-AGATGAGCATTGGCAGCGAG-3'; β-actin Forward:
5-CATGTACGTTGCTATCCAGGC-3; and β-actin Reverse:
5-CTCCTTAATGTCACGCACGAT-3. The relative quantification of LIN28B
gene was determined by using the comparative Cq
(ΔΔCq) method as recommended by the
manufacturer (21).
Serial replating assay
For evaluating self-renewal capacity, serial
replating assay was assessed. Briefly, 1,000 cells were plated in
DMEM/F12 medium supplemented with 20 ng/ml of epidermal growth
factor, 10 ng/ml of basic fibroblast growth factor and 2% B-27 and
incubated at 37˚C for 10 days. Then the same number of dissociated
cells were replated and four passages were replated.
CCK-8 assay
To evaluate the effects of EVO exposure on cell
viability, CCK-8 assay was performed. Cells were seeded in 96-well
plates (5x104 cells/well) and 10 µl CCK-8 solution
(MilliporeSigma) was added to each well and incubated at 37˚C for 2
h in a CO2 incubator. The absorbance value was measured
at 620 nm wavelength on the MultiSkan spectrum microplate reader
(Thermo Fisher Scientific, Inc.). The experiments were repeated
three times.
CFSE/PI double staining assay
The cytotoxicity of EVO was evaluated by
CFSE/propidium iodide (PI; MilliporeSigma) double staining.
Briefly, CFSE-pre-stained cells were incubated with EVO at 37˚C for
24 h, then cells were stained with 5 µg/ml PI at room temperature
for 10 min in darkness. Then cells were washed three times with PBS
and PI absorbance was determined by FACS on a flow cytometer
(FACSCalibur, Becton, Dickinson and Company).
Statistical analysis
All data in the present study were presented as mean
± SD. Unpaired t-test was applied to compare between two groups.
One-way analysis of variance (ANOVA) followed by Tukey's post-hoc
test was used to analyze the statistical significance of >2
groups. P<0.05 was considered to indicate a statistically
significant difference. All experiments were carried out in
triplicate.
Results
Physiological processes of A549 were
inhibited by EVO treatment
A549 cells were treated with 0, 1, 2.5, 5, 7.5 or 10
µmol/l EVO over 24 h and cell viability was determined following
EVO exposure. The results were that, with 5, 7.5 or 10 µmol/l EVO
exposure for 24 h, cell viability was significantly decreased
compared to mock group, respectively (Fig. 1B). Then high content imaging was
employed to observe cell growth from 24-72 h following addition of
EVO (Fig. 1C). Similar results
were found that with exposure to 5, 7.5 or 10 µmol/l EVO, cell
confluence was obviously decreased (Fig. 1C). Notably, 24-h treatment of EVO
presented no effect on apoptosis (Fig.
1D), which indicated that EVO affects proliferation but not
apoptosis after 24-hour treatment.
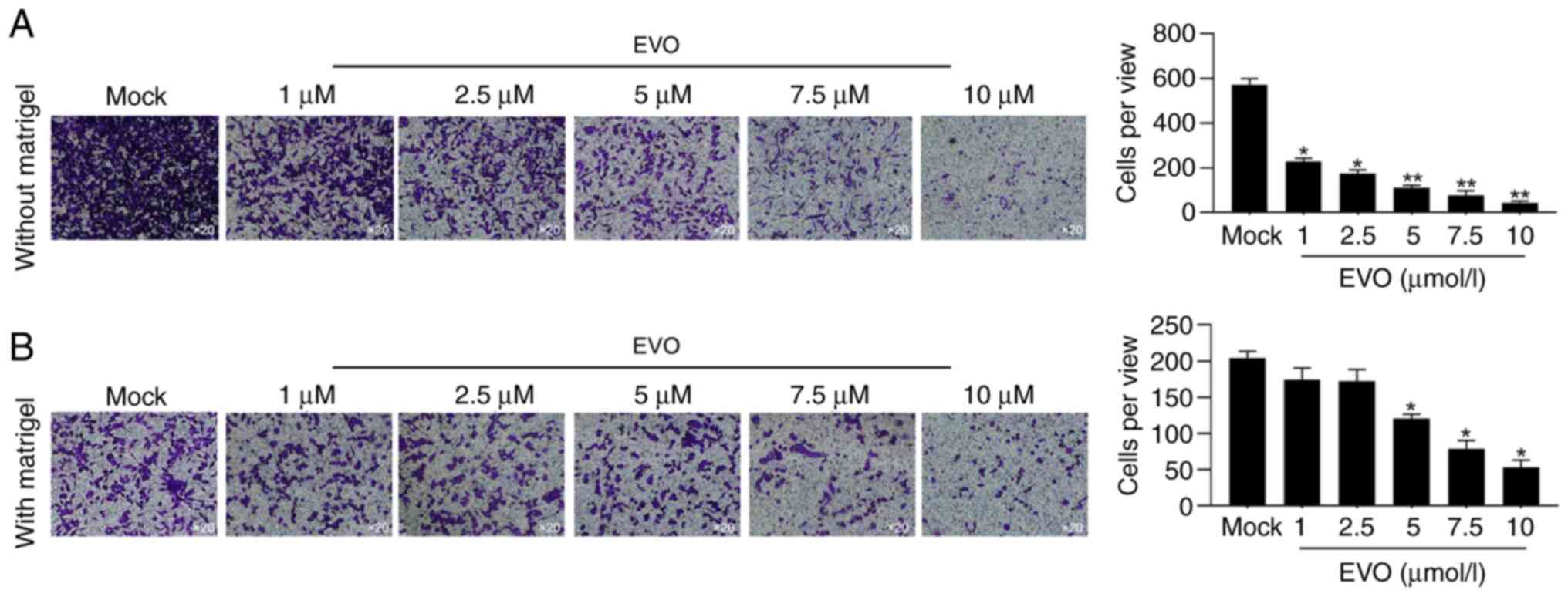
For evaluating the effects of EVO exposure on
malignant behaviors including cell migration and invasion,
Transwell assay without or with Matrigel was performed after
24-hour pretreatment of different concentration of EVO. As
expected, 1-10 µmol/l of EVO decreased both migration and invasion
activity in A549 (Fig. 2A and
B), which is consistent with a
previous finding (6).
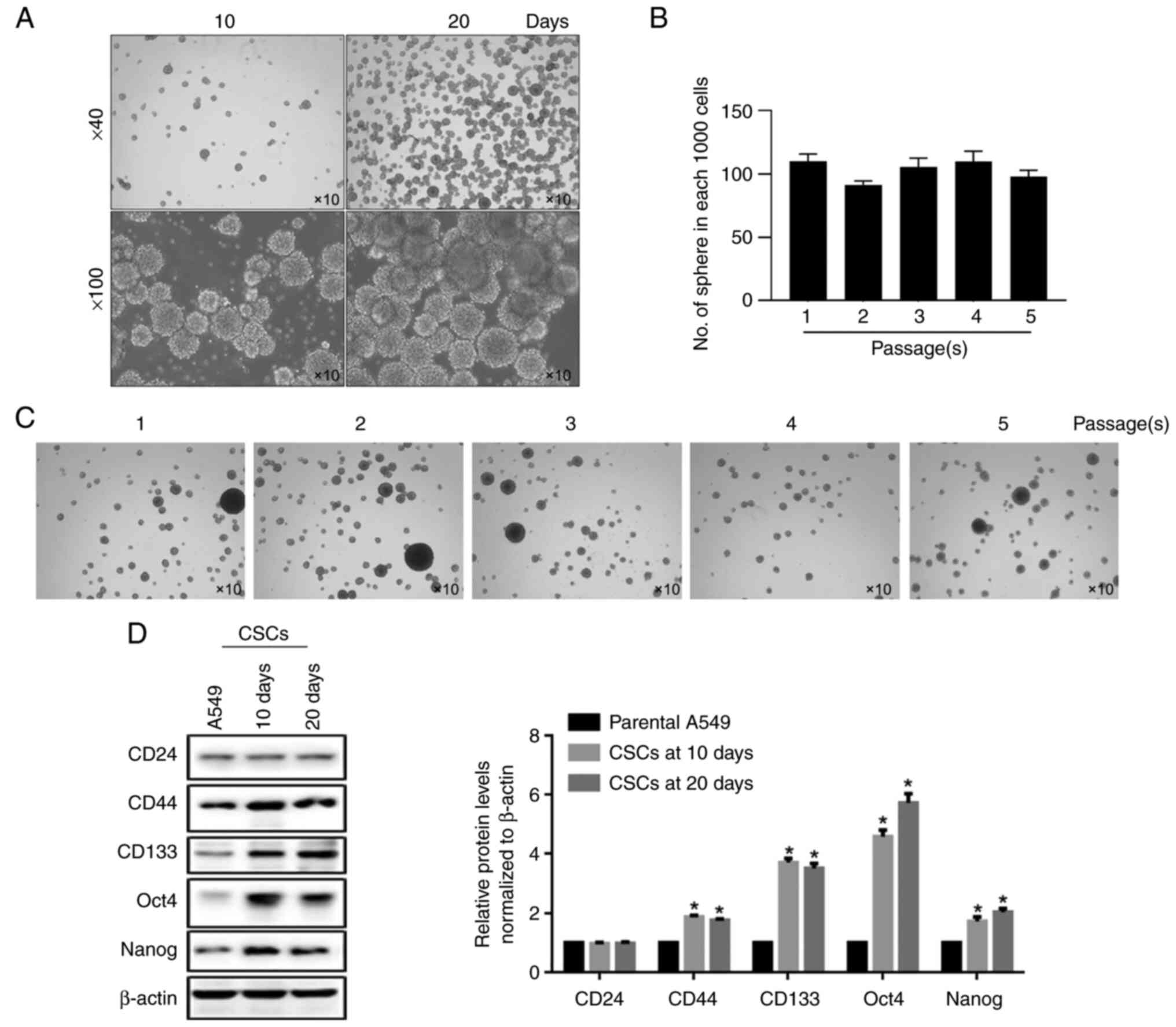
Enrichment and identification of CSCs
from A549 cells
The inhibitory effects of EVO on A549 cells promoted
speculation on whether EVO exerted similar effects on CSCs derived
from A549 cells. By considering the widely accepted method for
enriching the stem-like cells by sphere-forming ability of cancer
cells in serum-free medium, A549 cells were cultured in serum-free
DMEM/F12 supplemented with EGF, bFGF and B27 for 10 and 20 days.
Morphologically, unattached spheres were observed at both 10 and 20
days (Fig. 3A). To confirm whether
enriched spheres present self-renewal capacity, additional serial
replating experiments were performed. By passaging for five times,
no detectable decrease in the number of formed spheres in each
1,000 cells, indicates the notable self-renewal capacity of
stem-like cells enriched from A549 (Fig. 3B and C). Furthermore, several hallmarks of
stemness, including CD24, CD44, CD133, Oct4 and Nanog, were
determined by semi-quantitative western blot. As expected, compared
with parental A549 cells, CD44, CD133, Oct4 and Nanog were found
significantly upregulated in both 10- and 20-day spheres (Fig. 3D).
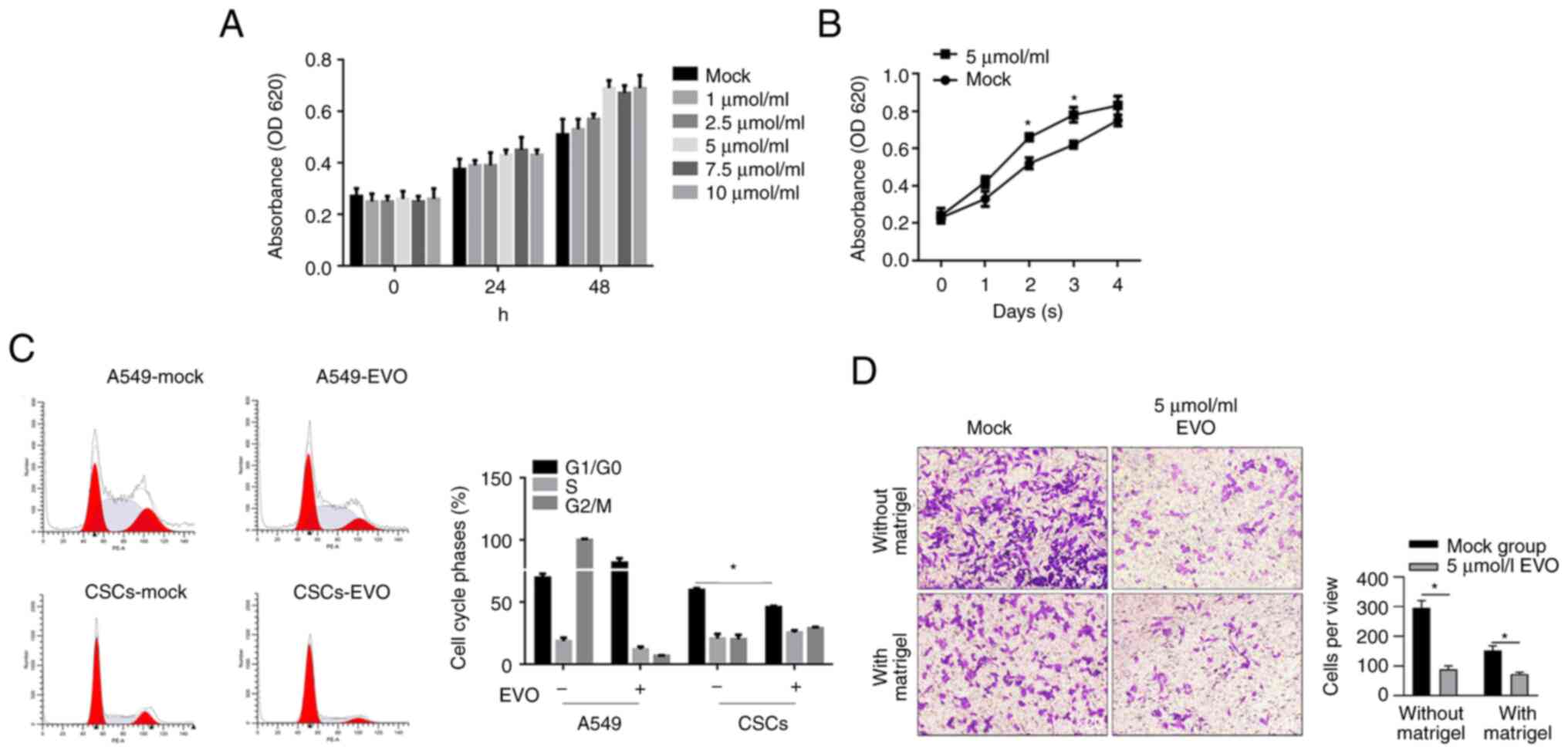
Effects of EVO treatment on
physiological processes of CSCs derived from A549 cells
By considering the inhibitory effects of different
concentration of EVO on A549's malignant behaviors including
proliferation, migration and invasion (Fig. 2), whether EVO exerted similar
effects to CSCs enriched from A549 was investigated. By performing
CCK-8 assay, it was found that 5.0, 7.5 and 10 µmol/l EVO exposure
clearly promoted cell viability after 24 and 48 h (Fig. 4A). To further confirm its effects
on proliferation of CSCs, 5 µmol/l EVO was employed. As shown in
Fig. 4B, EVO exposure
significantly promoted cell viability, which was opposite to the
expectation of the present study. In order to rule out the
possibility of promoting viability instead of promoting
proliferation, EVO-exposed CSCs were fixed with 4% paraformaldehyde
and stained with 5 µg/ml PI followed by cytometric analysis. The
results confirmed that EVO exposure decreased the proportion of
G1/G0 phase in CSCs and, by contrast,
arrested cell phase at G1/G0 in A549 cells,
which explained the opposite effects in CSCs and A549 cells
(Fig. 4C). The present study then
evaluate the effects of EVO on migration and invasion in CSCs.
Oddly, EVO treatment inhibited migration and invasion in CSCs,
similar to the effects in A549 cells (Fig. 4D).
EVO treatment inactivated EMT program
and decreased stemness of CSCs
EMT has been accepted as a key and reversible
process, which allows cancer cells to be activated during the
metastasis process (22,23). By considering the effects of EVO on
migration and invasion in CSCs enriched from A549 cells, the
present study measured the relative expressing levels of EMT
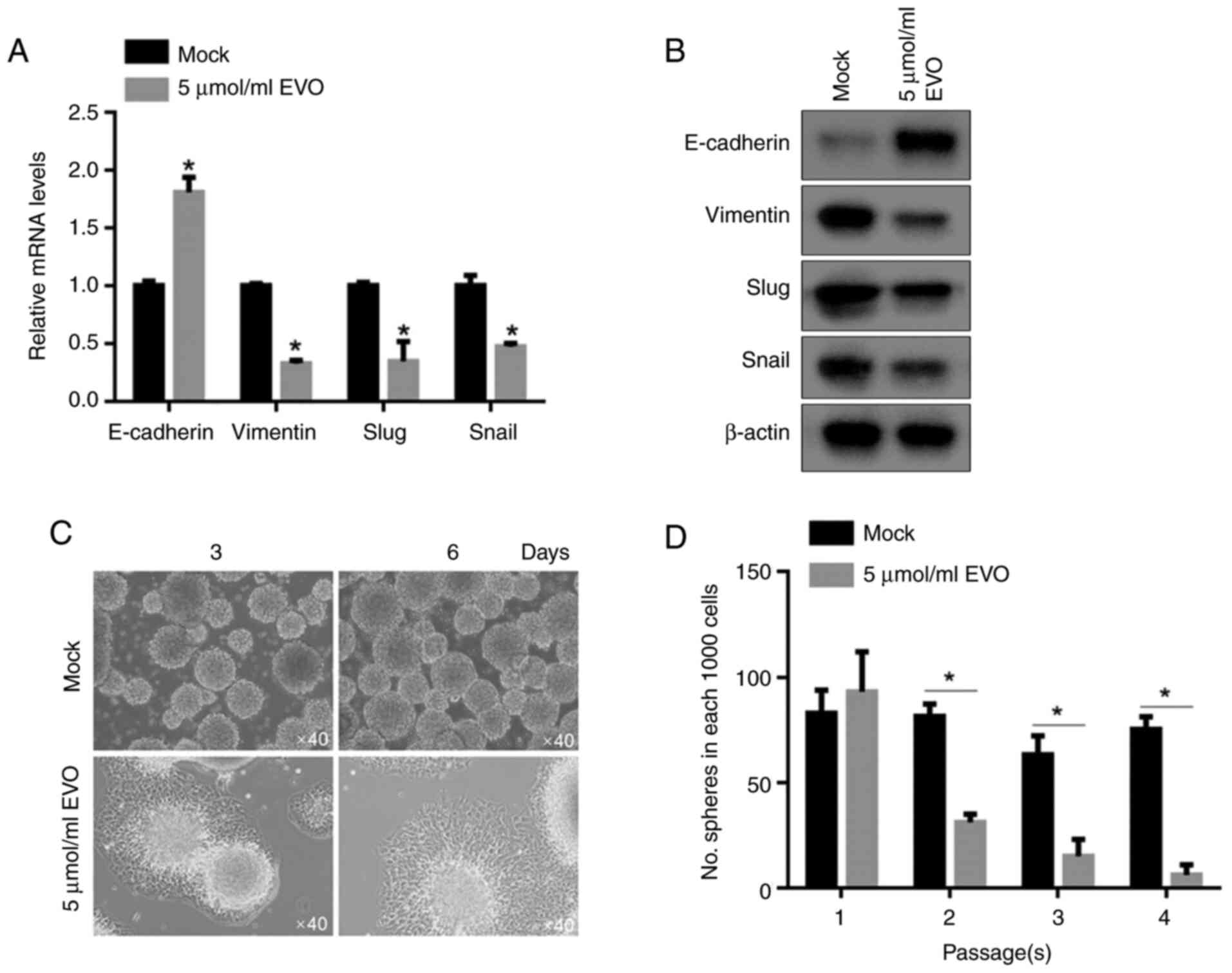
hallmarks, including E-cadherin, Vimentin, Slug and Snail. As shown
in Fig. 5A and B, both mRNA and protein levels of
E-cadherin, were upregulated and those of Vimentin, Slug and Snail
were downregulated in EVO-treated CSCs, compared to untreated CSCs,
demonstrating that EVO treatment inactivated EMT-program. It has
been reported that the changes in expression levels of these
hallmarks of EMT lead to morphological changes in the cells
(24). After being cultured in
EVO-contained medium for 3-6 days, spheres attempted to attach to
the bottom of the wells (Fig. 5C),
indicating that EVO treatment promoted cells differentiation or
inhibited maintenance of stemness. By performing self-replating
experiment, it was confirmed that EVO exposure decreased
self-renewal capacity of CSCSs (Fig.
5D).
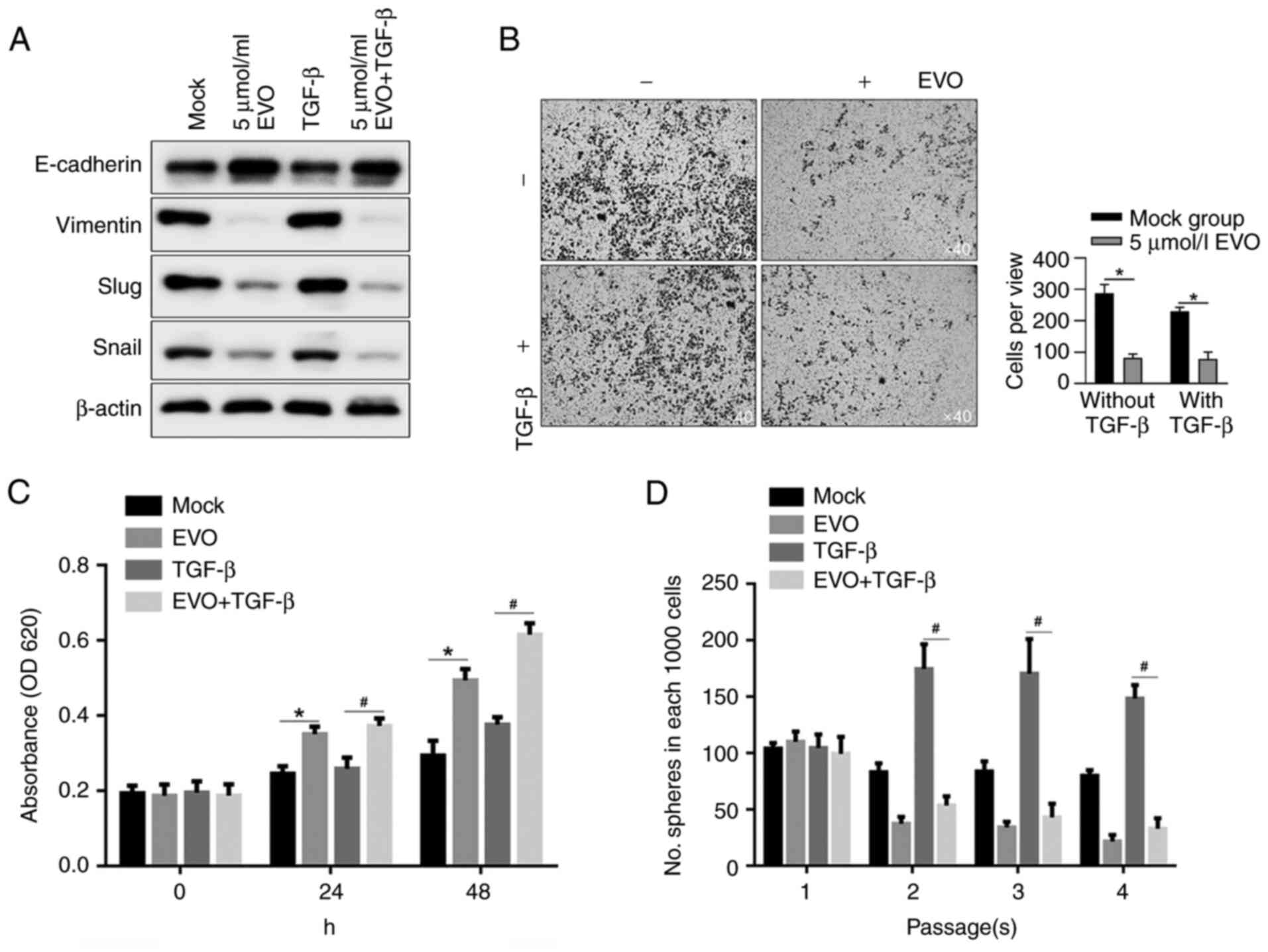
EVO treatment regulated physiological processes of
CSCs potentially via inactivating EMT. TGF-β is the most studied
growth factor and it serves a central regulating role in activating
EMT. The present study, by employing TGF-β as an EMT-activator
before EVO exposure, tried to clarify whether the effects of EVO
exposure on physiological processes of CSC by inactivating
EMT-program. By measuring the hallmarks of EMT with
semi-quantitative western blotting, it was shown that upregulation
of EMT hallmarks by TGF-β stimulation was inhibited following EVO
exposure, indicating that EVO exposure not only morphologically
inhibited migration and invasion, but also affected EMT signaling
in CSCs (Fig. 6A). By performing
Transwell assay with Matrigel, consistent results were obtained
with the expressing levels of EMT hallmarks (Fig. 6B). TGF-β-promoted cell
proliferation was also inhibited by EVO exposure in CSCs (Fig. 6C). By performing serial replating
experiments, as expected, EVO decreased self-renewal capacity in
CSCs and exerted antagonistic effect on maintaining stemness by
TGF-β stimulation (Fig. 6D).
Discussion
Although rapid advances in diagnostic and operative
techniques have been developed, due to the advanced stage of lung
cancer (8) it remains one of the
most difficult human malignancies to treat, which leads to a low
survival rate and poor quality of life. The potential roles of CSCs
in regulating malignant behaviors, including metastasis and
recurrence, have attracted attention (9,10).
Accumulating studies have demonstrated that EVO
exerts an inhibitory effect on the proliferation of several types
of cancer, including gastrointestinal (3), genitourinary tract (4), breast (5), prostate (6) and colon (7) cancer. Consistent with previous
studies, the present study found that EVO inhibited NSCLC cell
proliferation. However, EVO treatment promoted cell proliferation
and cell viability in A549-derived stem-like cells, indicating that
A549 CSCs are more resistant to EVO. Considering the contradictory
results, a concentration range of EVO exposure (1-10 µmol/l) was
set to test its effect on the malignant behaviors of A549 cells and
CSCs derived from A549 cells. The results showed that A549 cells
and CSCs derived from A549 cells presented different reactivities
to EVO. EVO exposure inhibited migrating and invasive abilities in
both A549 cells and CSCs derived from A549 cells. However, instead
of inhibiting proliferation in A549 cells, EVO exposure promoted
proliferation in CSCs. By considering that existence of CSCs
contributes to chemoresistance, exposure to EVO may be a promising
strategy to overcome chemoresistance caused by CSCs. However, as a
limitation, the effect of EVO on overcoming chemoresistance related
to the presence of CSCs was not detected, which is worth further
study.
The present study exposed non-small-cell lung cancer
cells A549 and CSCs derived from A549 to EVO and tested its
efficacy against physiological processes, including self-renewal
capacity, proliferation, migration and invasion. It was determined
that EVO exposure exerted inhibitory effects on malignant behavior
in A549 cells. In CSCs, despite promoting proliferation, EVO
exposure also inhibited malignant behaviors. It was found that EVO
treatment significantly decreased the maintenance of the
self-renewal capacity in CSCs by inactivating the EMT process. The
inducing effects of the EMT by TGF-β were confirmed in CSCs and EVO
treatment abolished the induction of TGF-β on the EMT. This finding
indicated the potential mechanism by which EVO regulates CSCs by
affecting the EMT process. Considering that all the experiments
were carried out in vitro, the exact roles of EVO on lung
epithelial cancer cells and CSCs should be investigated in
vivo.
In summary, the present results show that EVO
treatment inhibited physiological processes in A549 cells. To the
best of our knowledge, this is the first study to evaluate the
effects of EVO exposure on CSCs derived from non-small cell lung
cancer cells, and the inhibitory effects towards CSCs, potentially
by regulating EMT, were confirmed. Taken together, the results
suggested that EVO can be an promising chemoagent that exerts
inhibitory effects not only on non-small-cell lung cancer cells but
also on CSCs, which are a critical subpopulation contributing to
poor prognosis. The data may provide therapeutic suggestions on the
use of EVO in non-small-cell lung cancer-related surgery.
Acknowledgements
The authors would like to thank Mr. Hong Tao
(Sichuan University, Chengdu, China) for language editing and
suggestions for the statistical analysis.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XL and ML designed the experiments. XL, WZ and YL
performed cell culture-associated experiments. YL and ML were
responsible for data collection and performed the statistical
analysis. XL, WZ, YL and ML confirmed the authenticity of all the
raw data. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yu H, Jin H, Gong W, Wang Z and Liang H:
Pharmacological actions of multi-target-directed evodiamine.
Molecules. 18:1826–1843. 2013.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Huang DM, Guh JH, Huang YT, Chueh SC,
Chiang PC and Teng CM: Induction of mitotic arrest and apoptosis in
human prostate cancer pc-3 cells by evodiamine. J Urol.
173:256–261. 2005.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Zhao LC, Li J, Liao K, Luo N, Shi QQ, Feng
ZQ and Chen DL: Evodiamine induces apoptosis and inhibits migration
of HCT-116 human colorectal cancer cells. Int J Mol Sci.
16:27411–27421. 2015.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Kan SF, Huang WJ, Lin LC and Wang PS:
Inhibitory effects of evodiamine on the growth of human prostate
cancer cell line LNCaP. Int J Cancer. 110:641–651. 2004.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Wang KL, Hsia SM, Yeh JY, Cheng SC, Wang
PS and Wang SW: Anti-proliferative effects of evodiamine on human
breast cancer cells. PLoS One. 8(e67297)2013.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Ogasawara M, Matsubara T and Suzuki H:
Inhibitory effects of evodiamine on in vitro invasion and
experimental lung metastasis of murine colon cancer cells. Biol
Pharm Bull. 24:917–920. 2001.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Zhang C, Fan X, Xu X, Yang X, Wang X and
Liang HP: Evodiamine induces caspase-dependent apoptosis and S
phase arrest in human colon Lovo cells. Anticancer Drugs.
21:766–776. 2010.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Lynch TJ, Adjei AA, Bunn PA Jr, Eisen TG,
Engelman J, Goss GD, Haber DA, Heymach JV, Jänne PA, Johnson BE, et
al: Summary statement: Novel agents in the treatment of lung
cancer: Advances in epidermal growth factor receptor-targeted
agents. Clin Cancer Res. 12 (14 Pt 2):4365s–4371s. 2006.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001.PubMed/NCBI View
Article : Google Scholar
|
|
11
|
Wang JC and Dick JE: Cancer stem cells:
Lessons from leukaemia. Trends Cell Biol. 15:494–501.
2005.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Clarke MF, Dick JE, Dirks PB, Eaves CJ,
Jamieson CH, Jones DL, Visvader J, Weissman IL and Wahl GM: Cancer
stem cells-perspectives on current status and future directions:
AACR workshop on cancer stem cells. Cancer Res. 66:9339–9344.
2006.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Tirino V, Camerlingo R, Franco R, Malanga
D, La Rocca A, Viglietto G, Rocco G and Pirozzi G: The role of
CD133 in the identification and characterisation of
tumour-initiating cells in non-small-cell lung cancer. Eur J
Cardiothorac Surg. 36:446–453. 2009.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Chiou SH, Wang ML, Chou YT, Chen CJ, Hong
CF, Hsieh WJ, Chang HT, Chen YS, Lin TW, Hsu HS and Wu CW:
Coexpression of Oct4 and Nanog enhances malignancy in lung
adenocarcinoma by inducing cancer stem cell-like properties and
epithelial-mesenchymal transdifferentiation. Cancer Res.
70:10433–10444. 2010.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Yuan P, Kadara H, Behrens C, Tang X, Woods
D, Solis LM, Huang J, Spinola M, Dong W, Yin G, et al: Sex
determining region Y-Box 2 (SOX2) is a potential cell-lineage gene
highly expressed in the pathogenesis of squamous cell carcinomas of
the lung. PLoS One. 5(e9112)2010.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Eramo A, Lotti F, Sette G, Pilozzi E,
Biffoni M, Di Virgilio A, Conticello C, Ruco L, Peschle C and De
Maria R: Identification and expansion of the tumorigenic lung
cancer stem cell population. Cell Death Differ. 15:504–514.
2008.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Justilien V, Regala RP, Tseng IC, Walsh
MP, Batra J, Radisky ES, Murray NR and Fields AP: A. Fields, Matrix
Metalloproteinase-10 is required for lung cancer stem cell
maintenance, tumor initation and metastatic potential. PLoS One.
7(e35040)2010.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Zhang L, Wang D, Li Y, Liu Y, Xie X, Wu Y,
Zhou Y, Ren J, Zhang J, Zhu H and Su Z: CCL21/CCR7 axis contributed
to CD133+ pancreatic cancer stem-like cell metastasis via EMT and
Erk/NF-kB pathway. PLoS One. 11(e0158529)2016.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan
A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, et al: The
epithelial-mesenchymal transition generates cells with properties
of stem cells. Cell. 133:704–715. 2008.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Morel AP, Lièvre M, Thomas C, Hinkal G,
Ansieau S and Puisieux A: Generation of breast cancer stem cells
through epithelial-mesenchymal transition. PLoS One.
3(e2888)2008.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Spaderna S, Schmalhofer O, Hlubek F, Berx
G, Eger A, Merkel S, Jung A, Kirchner T and Brabletz T: A
transient, EMT-linked loss of basement membranes indicates
metastasis and poor survival in colorectal cancer.
Gastroenterology. 131:830–840. 2006.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Prall F: Tumour budding in colorectal
carcinoma. Histopathology. 50:151–162. 2007.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Tiwari N, Gheldof A, Tatari M and
Christofori G: EMT as the ultimate survival mechanism of cancer
cells. Semin Cancer Biol. 22:194–207. 2012.PubMed/NCBI View Article : Google Scholar
|