Introduction
Juvenile idiopathic arthritis (JIA) refers to
chronic inflammation of the joints in children under 16 years of
age that persists for more than six weeks (1,2). It
is the most common rheumatoid disease in childhood, characterized
by manifestations and recurrent arthritis, with a prevalence of
1/10,000(3). The pathogenesis of
JIA is not well understood and may be related to many factors, such
as infection, immunity and heredity (4). An increasing number of publications
have confirmed that JIA has a complex genetic background. One of
the most studied is human leukocyte antigen (HLA) (5). In addition, certain HLA loci
have been shown to be associated with JIA (6). The genetic associations between
non-HLA genes including protein tyrosine phosphatase
non-receptor type 22 (PTPN22) (7), Toll-like receptor 4 (TLR4)
(8), TNF-α (9) and JIA susceptibility have been
extensively studied and confirmed.
PTPN22, located in 1p13.2, encodes
lymphospecific protein tyrosine phosphatase (LYP), which is a
member of the family that inhibits T-cell activation and serves an
important role in immune homeostasis (10). A variant of the PTPN22 gene
(rs2476601) is reported to be related to multiple autoimmune
diseases, including lupus erythematosus (SLE), rheumatoid arthritis
(RA) and ankylosing spondylitis (AS) in Caucasian and Asian
populations (10-12).
Multiple studies have suggested that PTPN22 rs2476601 is
associated with JIA in Caucasian (13,14)
and Chinese Han populations (15).
However, studies have suggested that PTPN22 rs2476601 is not
polymorphic in the Chinese Han population (10,11).
Furthermore, the studies referred to the rare relationship between
other variants in the PTPN22 gene and JIA risk. Thus, it is
necessary to investigate new JIA-related loci in the PTPN22
gene.
Chronic arthritis, a common feature of RA, AS, SLE
and JIA, is suggested to be associated with dysregulation of the
immune response (16). IFN and IFN
regulatory factors serve an important role in the immune response,
as well as in the occurrence and progression of autoimmune diseases
(17). IFN regulatory factor 5
(IRF5), a member of the interferon regulatory factor family, is a
class of transcription factors that mainly regulate the expression
of IFN and IFN responsive genes (18). The IRF5 gene can induce IFN
production and participate in the expression of inflammatory
factors, which suggests that the IRF5 gene may serve a role
in the pathogenesis of autoimmune diseases (19). In addition, tyrosine kinase 2
(TYK2), a member of the receptor tyrosine kinase Janus kinase (JAK)
family, has been shown to serve an important role in the type I IFN
signaling pathway (20).
Furthermore, IRF5 and TYK2 genes variants are
suggested to be related to the susceptibility of autoimmune
diseases including SLE, RA and multiple sclerosis (MS) (21-23).
However, few studies on the genetic association between the
IRF5 and TYK2 gene variants and JIA susceptibility
have been reported.
To further evaluate the role of the variants in
PTPN22, IRF5 and TYK2 genes in the pathogenesis of JIA, a
case-control study on the genetic associations between the variants
in PTPN22, IRF5 and TYK2 genes and JIA risk was
performed in a Han Chinese population by using direct
sequencing.
Materials and methods
Subjects
The protocol of the present study was performed
following the ethical standards laid down in the 1964 Declaration
of Helsinki and its later amendments and approved by the Local
Ethics Committee of the Changshu Hospital affiliated with Soochow
University (protocol number: EC-19-024). Written informed consent
was obtained from the participants' parents before the study.
A total of 378 healthy individuals from Changshu
Hospital Affiliated to Soochow University (between 06/01/2019 and
12/31/2020) were randomly selected as the normal control group (67
male and 311 female participants), with an average age of 7.38±1.42
years, excluding heart, liver, kidney and vascular inflammatory
diseases. A total of 378 patients with JIA (68 male and 311 female
patients) were selected as the case group. All the patients were
ranked as systemic JIA. The mean age was 7.29±2.58 years and the
mean age of onset was 6.3±1.3 years. The clinical characteristics
of JIA patients and healthy controls are summarized in Table I. The diagnosis of JIA was
confirmed by typical clinical symptoms and laboratory tests
according to the International League of Associations for
Rheumatology (24), but not
European Alliance of Associations for Rheumatology (EULAR) as the
validity of the EULAR criteria is low since it often exclude
patients from subgroup classification and the possibility of having
more than one diagnosis is not negligible (25). All patients received non-steroidal
anti-inflammatory drugs and methotrexate therapy at the initial
stage of the disease. All subjects were unrelated individuals of
Han ethnic group in Jiangsu Province.
 | Table IClinical characteristics of JIA
patients and healthy controls. |
Table I
Clinical characteristics of JIA
patients and healthy controls.
| Clinical
features | JIA (mean ±
SD) | Control (mean ±
SD) | P-value |
|---|
| Sex
(Female/Male) | 311/67 | 311/67 | 1.00 |
| Age (years) | 7.29±2.58 | 7.38±1.42 | >0.05 |
| Onset age
(years) | 6.3±1.3 | - | - |
| DAS28 | 3.2±1.2 | - | - |
| RF+, % | 12.15 | - | - |
| CCP+, % | 18.54 | - | - |
| ANA+, % | 18.31 | - | - |
| ESR (mm/h) | 36.8±12.3 | 4.2±2.1 | <0.01 |
| CRP (mg/l) | 18.4±7.5 | 3.6±1.3 | <0.01 |
| IgA mg/ml | 11.1±2.7 | 2.7±1.4 | <0.01 |
| IgG mg/ml | 26.6±3.1 | 9.4±1.7 | <0.01 |
| IgM mg/ml | 6.1±1.9 | 1.9±0.7 | <0.01 |
Sample collection and DNA
extraction
A total of 2 ml fasting venous blood was collected
from the right arm of all subjects'. The blood cells were used for
DNA extraction and the blood plasma was stored at -80˚C. Genomic
DNA was extracted from 200 µl anticoagulant blood cells using a
QIAamp DNA Mini kit (Qiagen GmbH).
PCR and genotyping
Variants in the PTPN22, IRF5 and TYK2 genes
in the present study were selected according to Tang et al
(10,11,21).
The primer sequence was synthesized by Dalian Bao Biological
Engineering Co., Ltd. Genomic DNA was extracted from peripheral
leukocytes using the standard phenol-chloroform method. The total
volume of PCR amplification was 25 µl, including 12.5 µl 2X GoTap
Green Master Mix (Promega Corporation; 400 µmol/l dNTP, 3 mmol/l
MgCl2), 1.5 µl 10 µmol/l upstream and downstream primers
and 200 ng DNA, respectively. The cycle parameters were
pre-denatured at 94˚C for 5 min, then denatured at 94˚C for 30 sec,
annealed at 57˚C for 30 sec and extended at 72˚C for 30 cycles and
finally, extended at 72˚C for 5 min. The PCR was performed using
the Bio-Rad CFX384 PCR thermocycler (Bio-Rad Laboratories, Inc.).
The PCR amplification product was stained with 8% ethidium bromide
on 1% Nusieve 3:1Agrose gel. Electrophoresis was performed at 100 V
for 15 min. Markers under UV light and D2000 plus DNA ladder
(Beijing Solarbio Science & Technology Co., Ltd.) were compared
to estimate DNA molecular weight. The product was purified by
adding 60 µl of 75% ethanol. Sequencing reactions were performed
with ABI PRISM BigDye Terminator V3.1 Sequencing kit (cat. no.
4336913; Applied Biosystems; Thermo Fisher Scientific, Inc.), with
one-way sequencing primer identical to PCR forward primer. The
reaction system was supplemented with 5 µl reaction system,
including 2 µl PCR amplification product, 1 µl 1 µM one-way primer,
0.5 µl ABI PRISM BigDye and 1 µl 5X reaction mix buffer,
supplemented with ddH2O to 5 µl. The reaction conditions
of sequencing PCR were denaturation at 96˚C for 45 sec, followed by
30 cycles of 96˚C for 10 sec, 55˚C for 5 sec and 60˚C for 2.5 min.
Sequencing reaction products were purified using multiScreen
96-well filtration plates. Sequencing electrophoresis was performed
according to ABI 3730XL DNA Sequencer instructions (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The sequencing primers
of the variants are listed in the Table SI. GeneScan 3.7 Software (Applied
Biosystems; Thermo Fisher Scientific, Inc.) was used for data
analysis.
Statistical analysis
The subjects were compared with Hardy-Weinberg
equilibrium (HWE), genotype and allele frequency by four-table
χ2 test and RC contingency table χ2 test. The
relative risk was estimated by odds ratio (OR) and 95% confidence
interval (95% CI). Haplotypes were constructed with the lowest
frequency threshold (LFT) >0.01. Multiple comparison correction
was performed to adjust P-values (q value) using the Benja
Mini-Hochberg method based on the False Discovery Rate (FDR)
standard. The SHEsis software was used to analyze the data
(http://analysis.bio-x.cn/myAnalysis.php) (26). The multifactor dimensionality
reduction (MDR) 3.0.2 software was used to analyze the gene-gene
interactions (https://sourceforge.net/projects/mdr/) (27). Calculation power was obtained at
the 0.05 level of significance, assuming an OR of 1.5 (small effect
size) by using the G*Power software (www.gpower.hhu.de) (28). P<0.05 was considered to indicate
a statistically significant difference.
Results
The HWE test was performed for the frequencies of
variants in the healthy control group. The results showed that
distributions of these variants in this group were in HWE
(P>0.05), indicating that the selected subjects were
representative of the population. The results of linkage
disequilibrium showed that the PTPN22 rs1746853 and
rs3765598, as well as rs1970559 and rs3811021, were in strong
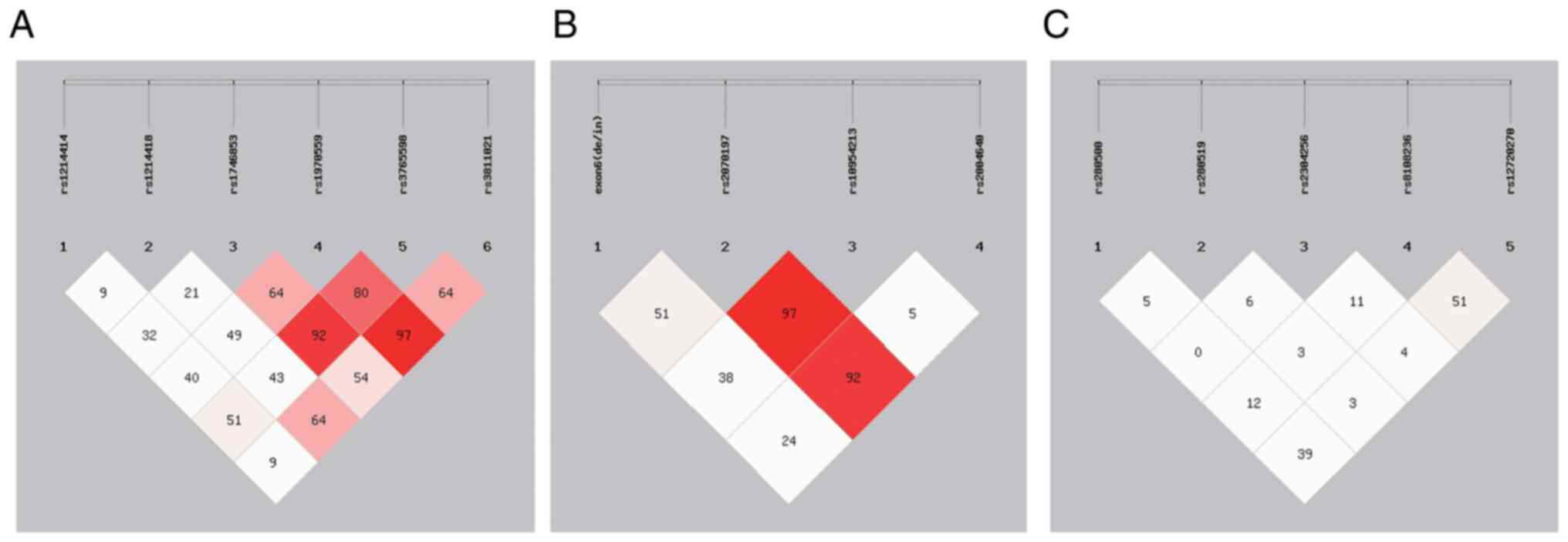
linkage (D'>85; Fig. 1A). In
addition, similar results were found for IRF5 rs2070197 and
rs10954213, as well as rs2070197 and rs2004640 (D'>85; Fig. 1B). However, the variants in the
TYK2 gene were found to be in strong linkage with each other
(D'<85; Fig. 1C). In addition,
a calculation power of 71.3% at the 5% significance level
(two-tailed) was obtained (Table
II).
| Table IIDistributions of PTPN22,
IRF5 and TYK2 genes polymorphisms in cases and
controls. |
Table II
Distributions of PTPN22,
IRF5 and TYK2 genes polymorphisms in cases and
controls.
| Gene | Polymorphisms
(A<B) (MAF, Asian) | Case
(AA/AB/BB) | Control
(AA/AB/BB) | Allele model [P, OR
(95%CI)]a | q (P
adjb) | Dominant model [P,
OR (95%CI)] | q (P
adjb) | Recessive model [P,
OR (95%CI)] | q (P
adjb) | HWE in control
group (P) | Power |
|---|
| PTPN22 | rs2476601 (T<C)
(0.0006) | 0/0/378 | 0/0/378 | - | - | - | - | - | - | 1.000 | 71.3 |
| | rs1214414 (T<C)
(0.2006) | 0/31/347 | 3/86/289 |
1.01x10-8, 0.31
(0.20-0.47) |
3.54x10-8 |
4.08x10-8, 0.29 (0.19,
0.45) |
9.25x10-8 | 0.2, 0.14 (0.01,
2.75) | 0.47 | 0.211 | |
| | rs1214418 (G<A)
(0.1508) | 5/80/293 | 3/35/340 |
2.91x10-7, 2.36 (1.61,
3.46) |
6.79x10-7 |
5.93x10-7, 2.60 (1.72,
3.92) |
2.07x10-6 | 0.48, 1.68 (0.40,
7.06) | 0.67 | 0.058 | |
| | rs1746853 (G<T)
(0.2703) | 27/133/218 | 14/90/274 |
5.20x10-6, 1.78 (1.37,
2.30) |
9.10x10-6 |
4.42x10-5, 1.93 (1.43,
2.62) |
3.09x10-4 | 0.04, 2.00 (1.03,
3.88) | 0.14 | 0.061 | |
| | rs1970559 (C<T)
(0.0665) | 0/36/342 | 0/31/347 | 0.53, 1.17 (0.72,
1.91) | 0.62 | 0.52, 1.18 (0.71,
1.95) | 0.61 | - | - | 0.410 | |
| | rs3765598 (T<C)
(0.2006) | 15/109/254 | 3/44/331 |
5.07x10-12, 3.18 (2.26,
4.47) |
3.55x10-11 |
1.29x10-10, 3.44 (2.37,
4.99) |
7.74x10-10 | 0.01, 5.17 (1.48,
17.99) | 0.07 | 0.262 | |
| | rs3811021 (C<T)
(0.2271) | 14/89/275 | 20/132/226 | 0.0003, 0.62
(0.48-0.81) | 0.0042 | 0.0002, 0.56 (0.41,
0.76) | 0.007 | 0.29, 0.69 (0.34,
1.38) | 0.51 | 0.899 | |
| IRF5 | exon6 (de/in)
(in<de) (0.4685) | 103/169/106 | 87/183/108 | 0.35, 1.10 (0.90,
1.35) | 0.47 | 0.87, 1.03 (0.75,
1.41) | 0.87 | 0.18, 1.25 (0.90,
1.74) | 0.24 | 0.576 | |
| | rs2070197 (C<T)
(0.0007) | 0/3/375 | 0/2/376 | 0.66, 1.50 (0.25,
9.01) | 0.66 | 0.66, 1.50 (0.25,
9.05) | 0.88 | - | - | 0.958 | |
| | rs10954213 (G<A)
(0.4814) | 34/116/228 | 49/197/132 |
9.12x10-10, 0.50 (0.40,
0.63) |
3.65x10-9 |
2.26x10-11, 0.35 (0.26,
0.47) |
9.04x10-11 | 0.08, 0.66 (0.42,
1.05) | 0.16 | 0.064 | |
| | rs2004640 (A<C)
(0.3245) | 20/156/202 | 10/136/232 | 0.02, 1.35 (1.06,
1.71) | 0.04 | 0.03, 1.38 (1.04,
1.85) | 0.06 | 0.07, 2.06 (0.95,
4.45) | 0.28 | 0.056 | |
| TYK2 | rs280500 (A<G)
(0.0178) | 34/196/148 | 12/124/242 |
2.31x10-11, 2.20
(1.74-2.78) | 1.16e-010 |
2.14x10-11 2.77 (2.06,
3.71) |
1.07x10-10 | 0.001, 3.01 (1.54,
5.92) | 0.005 | 0.416 | |
| | rs280519 (A<G)
(0.4938) | 93/184/101 | 124/185/69 | 0.001, 0.72 (0.58,
0.88) | 0.002 | 0.006, 0.61 (0.43,
0.87) | 0.01 | 0.01, 0.67 (0.49,
0.92) | 0.025 | 0.999 | |
| | rs2304256 (A<G)
(0.4075) | 24/159/195 | 10/89/279 |
6.15x10-10, 2.24 (1.73,
2.90) | 1.54e-09 |
1.82x10-9, 2.64 (1.95,
3.59) |
4.55x10-9 | 0.02, 2.49 (1.18,
5.29) | 0.033 | 0.371 | |
| | rs8108236 (A<G)
(0.0055) | 30/128/220 | 29/128/221 | 0.91, 1.01 (0.80,
1.28) | 0.91 | 0.94, 1.01 (0.76,
1.35) | 0.94 | 0.89, 1.04 (0.61,
1.77) | 0.89 | 0.089 | |
| | rs12720270 (G<T)
(0.3897) | 2/42/334 | 1/68/309 | 0.02, 0.63
(0.43-0.93) | 0.025 | 0.01, 0.59 (0.39,
0.89) | 0.013 | 0.7, 2.01 (0.18,
22.21) | 0.87 | 0.170 | |
For PTPN22, the rs2476601 was found to be not
polymorphic and was deleted from the subsequent analysis. All the
allelic and dominant models of the five variants (rs1214414,
rs1214418, rs1746853, rs3765598 and rs3811021) except for rs1970559
were significantly associated with JIA risk, even after correction
with the FDR method (P<0.05; Table
II). In addition, the recessive models of rs1746853 and
rs3765598 were found to be associated with JIA risk (P<0.05).
However, these significant associations disappeared after
correction with the FDR method (P>0.05; Table II).
For IRF5, significant associations were
observed between the allelic and dominant models of rs10954213 and
the allelic model of rs2004640 and JIA risk, even after correction
with the FDR method (P<0.05). By contrast, no association was
detected between the recessive models of the rs10954213 and
rs2004640 and JIA susceptibility (P>0.05). In addition, the
exon6(de/in) and rs2070197 were not significantly associated with
the risk of JIA (P>0.05; Table
II).
For TYK2, all the allelic, recessive and
dominant models of the four variants (rs280500, rs280519 and
rs2304256, except for rs8108236) were significantly associated with
JIA risk, even after correction with the FDR method (P<0.05;
Table II). At the same time, the
allelic and dominant models but not the recessive model of
rs12720270 were detected to be significantly associated with the
risk of JIA (P<0.05; Table
II).
The haplotype results revealed that the frequencies
of PTPN22 haplotypes including HC A G T C C,
HC A G T T C and HC G T T C T in
the JIA patient group were significantly different from those in
the healthy group (P<0.05). The HC A T T C T was the
most common haplotype both in cases and controls with the
frequencies of 0.661 and 0.622, respectively (Table III). For IRF5, a total of
nine haplotypes (LFT>0.01) were observed. The frequencies of
IRF5 haplotypes (HD T A A and HI T A
C) were significantly higher in the case group (P<0.05).
Furthermore, the frequency of HD T G C was found to be
significantly lower in the case group (P<0.05; Table III). For TYK2, a total of
11 haplotypes (LFT>0.01) were detected. The HA G G A
T frequency was significantly higher in the case group
(P<0.05). In comparison, the frequencies of HG A G G
T and HG G G A T were found to be significantly
lower in the case group (P<0.05; Table III).
| Table IIIHaplotypes structure and frequencies
of PTPN22, IRF5 and TYK2 genes. |
Table III
Haplotypes structure and frequencies
of PTPN22, IRF5 and TYK2 genes.
| Gene |
Haplotypesa | Case (freq) | Control (freq) | P, OR
[95%CI]b | q
(Padjc) |
|---|
| PTPN22 | C A G T C C | 0.014 | 0.036 | 0.006, 0.384
[~0.189-0.781] | 0.008 |
| | C A G T T C | 0.115 | 0.043 |
5.02x10-7, 2.809
[~1.852-4.261] | 2.00e-006 |
| | C A T T C T | 0.661 | 0.622 | 0.307, 1.123
[~0.898-1.404] | 0.307 |
| | C G T T C T | 0.068 | 0.019 |
3.62x10-6, 3.779
[~2.075-6.884] | 7.24e-006 |
| IRF5 | D T A A | 0.144 | 0.096 | 0.004, 1.578
[1.151~2.163] | 0.010 |
| | D T A C | 0.300 | 0.290 | 0.644, 1.054
[~0.844-1.314] | 0.736 |
| | D T G A | 0.029 | 0.028 | 0.918, 1.032
[~0.564-1.887] | 0.918 |
| | D T G C | 0.029 | 0.113 |
2.22x10-10, 0.235
[~0.145-0.379] | 1.77e-09 |
| | I T A A | 0.052 | 0.030 | 0.027, 1.799
[~1.063-3.047] | 0.432 |
| | I T A C | 0.256 | 0.191 | 0.002, 1.460
[~1.144-1.864] | 0.008 |
| | I T G A | 0.034 | 0.064 | 0.007, 0.513
[~0.314-0.839] | 0.140 |
| | I T G C | 0.152 | 0.185 | 0.083, 0.787
[~0.601~1.032] | 0.111 |
| TYK2 | A A G A T | 0.034 | 0.025 | 0.297, 1.374
[~0.755-2.501] | 0.363 |
| | A A G G T | 0.080 | 0.064 | 0.210, 1.285
[~0.867-1.904] | 0.330 |
| | A G G A T | 0.080 | 0.021 |
1.29x10-7, 4.102
[~2.338-7.197] | 7.095e-007 |
| | A G G G T | 0.054 | 0.062 | 0.524, 0.869
[~0.563-1.341] | 0.576 |
| | G A A A T | 0.033 | 0.022 | 0.189, 1.516
[~0.810-2.837] | 0.416 |
| | G A A G T | 0.067 | 0.052 | 0.192, 1.330
[~0.865-2.045] | 0.352 |
| | G A G A T | 0.051 | 0.074 | 0.065, 0.673
[~0.440-1.028] | 0.178 |
| | G A G G T | 0.150 | 0.275 |
4.48x10-9, 0.468
[~0.362-0.605] | 4.93e-008 |
| | G G A G T | 0.045 | 0.041 | 0.689, 1.107
[~0.673-1.819] | 0.689 |
| | G G G A T | 0.033 | 0.062 | 0.008, 0.519
[~0.316-0.852] | 0.029 |
| | G G G G T | 0.214 | 0.193 | 0.277, 1.149
[~0.894-1.478] | 0.381 |
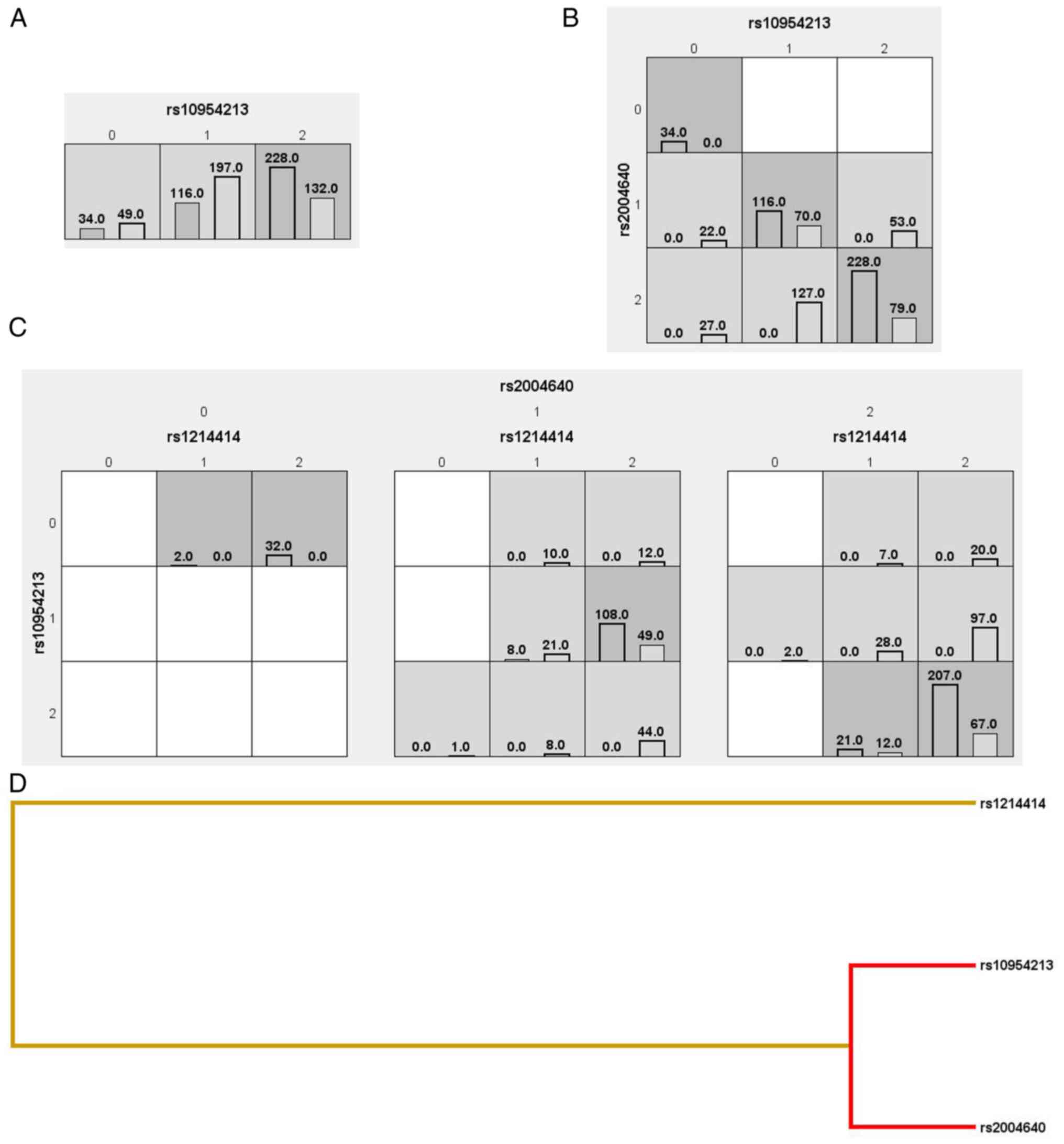
The MDR results revealed that IRF5 rs10954213
had the highest testing balanced accuracy among the tested
variants, with 61.84% testing balanced accuracy (TBA). IRF5
rs10954213, rs2004640 and PTPN22 rs1214414 suggested a
three-way interaction between PTPN22 and IRF5 genes
and JIA with 78.95% TBA and cross-validation consistency (CVC)
(10/10) (P<0.0001) (Fig.
2).
Discussion
In the present study, 378 unrelated patients with
JIA and 378 healthy controls were genotyped for the 16 variants in
PTPN22, IRF5 and TYK2 genes. The results indicated that five
variants in the PTPN22 gene, two variants in the IRF5
gene and four variants in the TYK2 gene were associated with
JIA susceptibility in the Chinese Han population. MDR analysis
suggested a three-way interaction (IRF5 rs10954213,
rs2004640 and PTPN22 rs1214414) between PTPN22 and
IRF5 genes and JIA.
The PTPN22 R620w variant (rs2476601) is found
to be associated with susceptibility to autoimmune diseases
(29,30). PTPN22 R620W causes an
alteration of arginine to tryptophan at codon 620 in the SH3
region, which downregulates the T-cell receptor (TCR) signaling
system, decreased the affinity between C-Src tyrosine kinase and
LYP, abolished the inhibition of tyrosine kinases LCK and Fyn and
then activated the TCR signaling pathway (31,32).
The genetic association between the PTPN22 R620W and
autoimmune diseases such as RA, SLE, Wegener's granulomatosis,
Type-1 diabetes, MS and JIA has been widely reported (33-36).
The PTPN22 rs2476601 has been shown to increase JIA risk in
American, Greek and Norwegian populations (14,34-38).
However, negative results have been reported in Czech, Hungarian
and Finnish populations (7,39,40).
Furthermore, meta-analysis suggests that the PTPN22
rs2476601 is a JIA susceptibility factor in Caucasian populations
(41-43).
However, no studies have been conducted on the genetic association
between the PTPN22 rs2476601 and JIA risk in the Chinese
population. In the present study, rs2476601 was found to be not
polymorphic in the Chinese Han population and no genetic
association was detected between rs2476601 and JIA susceptibility.
The discordance among the studies might be due to the different
genetic backgrounds among multiple ethnic groups.
In the present study, PTPN22 rs1214414,
rs1214418, rs1746853, rs3765598 and rs3811021 were found to be
significantly associated with JIA risk in the Chinese Han
population, which was similar to previous results reported by Tang
et al (10,11) on the relationship of PTPN22
variants and RA, SLE and AS risk. Notably, most of the selected
variants were in the introns of the PTPN22 gene and may not
affect the function of LYP. There are an increasing number of new
pathogenic variants located in introns and studies have suggested
that many disease-related intronic variants are responsible for
aberrant splice processes (44,45).
These observations indicate that the intronic variants in the
PTPN22 gene might affect the splice processes of LYP,
leading to aberrant expression of the PTPN22 and thereby
influencing the susceptibility to JIA.
IRF5 and Toll-like receptors (TLRs) form a pathway
to induce the expression of inflammatory factors (46). The lipopolysaccharide and nucleic
acid of the pathogen can be recognized by specific TLRs, causing
IRF5 to be activated, thus activating the downstream target gene
and serving the function of immune defense. At the same time,
processing and presenting foreign antigens, inducing T-cell
differentiation and activating acquired immunity (47). The IRF5 gene contains nine
exons and is located on the human genome 7q32(48). Studies have shown that loci in
IRF5, including exon 6(de/in), rs2004640, rs10954213,
rs2280714 and rs2070197 are important susceptibility factors for
autoimmune diseases (21-23).
The IRF5 rs2004640T allele creates a 5'-donor splicing site
in intron 1, resulting in the expression of untranslated exon 1B
(49). The IRF5 rs10954213
changes the polyadenylate sites of IRF5, which is related to the
mechanism of the increased IRF5 level in patients with autoimmune
diseases (50). The genetic
associations between these two variants and autoimmune diseases,
including SLE, RA, Sjogren syndrome and JIA, have been widely
reported (21,51,52).
However, no association between the IRF5 rs2004640 and JIA
risk was detected in Russian populations (53). In the present study, IRF5
rs2004640 and rs10954213 were first found to be significantly
related to the susceptibility to JIA in a Chinese Han population.
The inconsistency might be due to the relatively small sample size
and genetic backgrounds between the Russian and Chinese Han
populations. A larger number of subjects from multiple ethnicity is
necessary to confirm these results.
Several studies have demonstrated the important role
of TYK2 in the type I interferon signaling pathway. Activated TYK2
can selectively activate STAT3 and STAT5 in the downstream
substrates, initiating the JAK-STAT signaling pathway and finally
triggering the expression of the type I IFN gene (54). TYK2 is also involved in the
signaling of colony-stimulating factors, angiotensin II,
Platelet-activating factor and some cytokines, such as IL-6 and
IL-10(55). Therefore, changes in
TYK2 function may also lead to the restriction of other cytokine
signaling pathways, such as IL-10, which has been previously
reported to be associated with SLE (56). In addition, animal experiments
suggest that TYK2 gene-deficient mice are not susceptible to
experimental arthritis (57).
These studies have confirmed the importance of TYK2 in autoimmune
diseases. TYK2 gene located in human 19p13.2. Previous
linkage analyses have shown that this region is susceptible to
autoimmune diseases. Several TYK2 variants were associated
with the susceptibility to autoimmune diseases, such as SLE and RA,
and have been investigated and confirmed (21,58,59).
Significant associations have been detected between TYK2
rs280519, rs12720270, rs2304256 and SLE in the United Kingdom
(60). Similar results are
reported in a Chinese Han population conducted by Tang et al
(21). However, previous studies
found no association between the TYK2 rs12720270, rs2304256
and SLE risk in Japanese and Hong Kong populations (58,61).
Notably, only TYK2 rs34536443 has been investigated and no
genetic association with JIA risk was found in a northern Greece
population (62). In the present
study, variants reported by Tang et al (21) were selected. All the selected
variants except for rs8108236 were associated with JIA
susceptibility in the Chinese Han population. The genetic
associations between the TYK2 variants and JIA were detected
for the first time. The results indicated that TYK2
rs280500, rs280519, rs2304256 and rs12720270 might be the
susceptibility factors for JIA in a Jiangsu Chinese Han
population.
The association between haplotypes and JIA was also
analyzed in the present study to evaluate the combined influence of
multiple variants in the PTPN22, IRF5 and TYK2
genes. Three haplotypes (HC A G T C C, HC A G T T
C and HC G T T C T ) in the PTPN22 gene,
two haplotypes (HD T A A and HI T A C) in the
IRF5 gene and one haplotype (HA G G A T) in the
TYK2 gene were detected to be risk factors for JIA. In
addition, the haplotype (HD T G C) in the IRF5
gene and two haplotypes (HG A G G T and HG G G A
T) in the TYK2 gene were found to be protective
factors for JIA. This is the first study, to the best of the
authors' knowledge, on the association between JIA and haplotypes
in PTPN22, IRF5 and TYK2 genes. Nevertheless, further
investigation with a large sample size and haplotype analysis with
more variants are required to confirm the current study
results.
In addition to focusing on the effect of a single
gene on JIA, the variants of the PTPN22, IRF5 and TYK2 genes
were analyzed by the MDR method in the present study. The present
study found a three-way interaction between IRF5 rs10954213,
rs2004640 and PTPN22 rs1214414 and JIA risk, suggesting that
gene-gene interactions between PTPN22 and IRF5 genes
may increase JIA susceptibility in the Han Chinese population.
Limitations should be motioned. First, the present
study enrolled limited number of studies for analyzing the
association between the variants in PTPN22, IRF5 and
TYK2 genes and susceptibility of JIA, which may have
resulted in insufficient authority for identifying relationship of
PTPN22, IRF5 and TYK2 genes and JIA risk. Second,
although significant associations were found between haplotypes and
JIA, the protein data of the expression of three genes is
unavailable. It is impossible to distinguish the haplotype for each
individual, especially subjects with haplotypes.
In summary, the present study suggested that
PTPN22 rs1214418, rs1746853, rs3765598, IRF5
rs2004640, TYK2 rs280500 and rs2304256 might be risk factors
for JIA. In addition, PTPN22 rs1214414, rs3811021,
IRF5 rs10954213, TYK2 rs280519 and rs12720270 might
be protective factors for JIA. A three-way interaction between
IRF5 rs10954213, rs2004640 and PTPN22 rs1214414 might
be a risk factor for JIA in Han Chinese population. However, future
studies in other cohorts of patients with JIA need to be performed
to validate it.
Supplementary Material
The sequencing primers of varients in
PTPN22, IRF5 and TYK2 genes
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the Natural Science
Foundation of Changshu (grant no. csws201613).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YQ and BC participated in study design and data
collection, carried out the initial analysis and drafted the
article. ZW aided in data acquisition, data analysis and
statistical analysis. YP carried out literature search, data
acquisition and manuscript editing. ZW and YP confirm the
authenticity of all the raw data. BC and ZW performed manuscript
review. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
The protocol of this study was performed following
the ethical standards laid down in the 1964 Declaration of Helsinki
and its later amendments and approved by the Local Ethics Committee
of the Changshu Hospital affiliated to Soochow University (protocol
number: EC-19-024). Written informed consent was obtained from the
participants' parents before the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhong L, Wang W, Li J, Ma M, Gou L, Wang
C, Yu Z, Zhang T, Dong Y, Wei Q and Song H: The association of MEFV
gene mutations with the disease risk and severity of systemic
juvenile idiopathic arthritis. Pediatr Rheumatol Online J.
18(38)2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Bašić J, Vojinović J, Jevtović-Stoimenov
T, Despotović M, Cvetković T, Lazarević D, Sušić G, Milošević V,
Cvetković M and Pavlović D: The association of CAT-262C/T
polymorphism with catalase activity and treatment response in
juvenile idiopathic arthritis. Rheumatol Int. 39:551–559.
2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Simon TA, Harikrishnan GP, Kawabata H,
Singhal S, Brunner HI and Lovell DJ: Prevalence of co-existing
autoimmune disease in juvenile idiopathic arthritis: A
cross-sectional study. Pediatr Rheumatol Online J.
18(43)2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Nigrovic PA, Martínez-Bonet M and Thompson
SD: Implications of juvenile idiopathic arthritis genetic risk
variants for disease pathogenesis and classification. Curr Opin
Rheumatol. 31:401–410. 2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Smerdel A, Lie BA, Ploski R, Koeleman BP,
Førre Ø, Thorsby E and Undlien DE: A gene in the telomeric HLA
complex distinct from HLA-A is involved in predisposition to
juvenile idiopathic arthritis (JIA). Arthritis Rheum. 46:1614–1619.
2002.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Silvestri AD, Capittini C, Poddighe D,
Marseglia GL, Mascaretti L, Bevilacqua E, Scotti V, Rebuffi C, Pasi
A, Martinetti M and Tinelli C: HLA-DRB1 alleles and juvenile
idiopathic arthritis: Diagnostic clues emerging from a
meta-analysis. Autoimmun Rev. 16:1230–1236. 2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Pazár B, Gergely P, Nagy ZB, Gombos T,
Pozsonyi E, Rajczy K, Balogh Z, Sevcic K, Orbán I, Szodoray P and
Poór G: Role of HLA-DRB1 and PTPN22 genes in susceptibility to
juvenile idiopathic arthritis in Hungarian patients. Clin Exp
Rheumatol. 26:1146–1152. 2008.PubMed/NCBI
|
|
8
|
Wang Y, Chen LH, Li F, Bao M, Zeng J,
Xiang J, Luo H, Li J and Tang L: TLR4 rs41426344 increases
susceptibility of rheumatoid arthritis (RA) and juvenile idiopathic
arthritis (JIA) in a central south Chinese Han population. Pediatr
Rheumatol Online J. 15(12)2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Sprchez M, Samaca G, Iancu M, Bolba C and
Miu N: Relation of interleukin-6, TNF-alpha and interleukin-1alpha
with disease activity and severity in juvenile idiopathic arthritis
patients. Clin Lab. 58:253–260. 2012.PubMed/NCBI
|
|
10
|
Tang L, Wang Y, Zheng S, Bao M, Zhang Q
and Li J: PTPN22 polymorphisms, but not R620W, were associated with
the genetic susceptibility of systemic lupus erythematosus and
rheumatoid arthritis in a Chinese Han population. Hum Immunol.
77:692–698. 2016.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Tang L, Wang Y and Chen BF: A variant
within intron 1 of the PTPN22 gene decreases the genetic
susceptibility of ankylosing spondylitis in a central south Chinese
Han population. Scand J Rheumatol. 43:380–384. 2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Ramirez M, Quintana G, Diaz-Gallo LM,
Caminos J, Garces M, Cepeda L, Rondon F, Restrepo JF, Egea E,
Garavito G, et al: The PTPN22 C1858T variant as a risk factor for
rheumatoid arthritis and systemic lupus erythematosus but not for
systemic sclerosis in the Colombian population. Clin Exp Rheumatol.
30:520–524. 2012.PubMed/NCBI
|
|
13
|
Goulielmos GN, Chiaroni-Clarke RC,
Dimopoulou DG, Zervou MI, Trachana M, Pratsidou-Gertsi P,
Garyfallos A and Ellis JA: Association of juvenile idiopathic
arthritis with PTPN22 rs2476601 is specific to females in a Greek
population. Pediatr Rheumatol Online J. 14(25)2016.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Hinks A, Barton A, John S, Hawkins C,
Griffiths CE, Donn R, Thomson W, Silman A and Worthington J:
Association between the PTPN22 gene and rheumatoid arthritis and
juvenile idiopathic arthritis in a UK population: Further support
that PTPN22 is an autoimmunity gene. Arthritis Rheum. 52:1694–1699.
2005.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Fan ZD, Wang FF, Huang H, Huang N, Ma HH,
Guo YH, Zhang YY, Qian XQ and Yu HG: STAT4 rs7574865 G/T and PTPN22
rs2488457 G/C polymorphisms influence the risk of developing
juvenile idiopathic arthritis in Han Chinese patients. PLoS One.
10(e0117389)2015.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Hinks A, Eyre S, Ke X, Barton A, Martin P,
Flynn E and Packham J: Childhood Arthritis Prospective Study
(CAPS); UKRAG Consortium; BSPAR Study Group. et al: Overlap of
disease susceptibility loci for rheumatoid arthritis and juvenile
idiopathic arthritis. Ann Rheum Dis. 69:1049–1053. 2010.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Zhang J: Paradoxical roles of
interferon-gamma in autoimmune disease. Exp Rev Clin Immunol.
3:35–38. 2007.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Eames HL, Corbin AL and Udalova IA:
Interferon regulatory factor 5 in human autoimmunity and murine
models of autoimmune disease. Transl Res. 167:167–182.
2016.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Matta B and Barnes BJ: Coordination
between innate immune cells, type I IFNs and IRF5 drives SLE
pathogenesis. Cytokine. 132(154731)2020.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Chang Y, Xu S and Ding K: Tyrosine kinase
2 (TYK2) allosteric inhibitors to treat autoimmune diseases. J Med
Chem. 62:8951–8952. 2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Tang L, Wan P, Wang Y, Pan B, Wang Y and
Chen B: Genetic association and interaction between the IRF5 and
TYK2 genes and systemic lupus erythematosus in the Han Chinese
population. Inflamm Res. 64:817–824. 2015.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Peng Y, Chen B, Sheng X and Qian Y:
Polymorphisms in IRF5 and TYK2 genes are associated with rheumatoid
arthritis in a Chinese Han population. Med Sci Monit.
27(e928455)2021.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Tang L, Chen B, Ma B and Nie S:
Association between IRF5 polymorphisms and autoimmune diseases: A
meta-analysis. Genet Mol Res. 13:4473–4485. 2014.PubMed/NCBI View Article : Google Scholar
|
|
24
|
ILAR 2001. Abstracts of the 20th congress
of the international league of associations for rheumatology.
Edmonton, Alberta, Canada. August 26-30, 2001. J Rheumatol Suppl.
63:1–120. 2001.PubMed/NCBI
|
|
25
|
Berntson L, Fasth A, Andersson-Gäre B,
Kristinsson J, Lahdenne P, Marhaug G, Nielsen S, Pelkonen P and
Svensson E: Nordic Study Group. Construct validity of ILAR and
EULAR criteria in juvenile idiopathic arthritis: A population based
incidence study from the Nordic countries. International league of
associations for rheumatology. European league against rheumatism.
J Rheumatol. 28:2737–2743. 2002.PubMed/NCBI
|
|
26
|
Shi YY and He L: SHEsis, a powerful
software platform for analyses of linkage disequilibrium, haplotype
construction, and genetic association at polymorphism loci. Cell
Res. 15:97–98. 2005.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Hahn LW, Ritchie MD and Moore JH:
Multifactor dimensionality reduction software for detecting
gene-gene and gene-environment interactions. Bioinformatics.
19:376–382. 2003.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Kang H: Sample size determination and
power analysis using the G*Power software. J Educ Eval Health Prof.
18(17)2021.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Bottini N and Peterson EJ: Tyrosine
phosphatase PTPN22: Multifunctional regulator of immune signaling,
development, and disease. Ann Rev Immunol. 32:83–119.
2014.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Lee YH, Rho YH, Choi SJ, Ji JD, Song GG,
Nath SK and Harley JB: The PTPN22 C1858T functional polymorphism
and autoimmune diseases-a meta-analysis. Rheumatology (Oxford).
46:49–56. 2007.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Gregersen PK, Lee HS, Batliwalla F and
Begovivh AB: PTPN22: Setting thresholds for autoimmunity. Semin
Immunol. 18:214–223. 2006.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Carlton V, Hu X, Chokkalingam AP, Schrodi
SJ, Brandon R, Alexander HC, Chang M, Catanese JJ, Leong DU, Ardlie
KG, et al: PTPN22 genetic variation: Evidence for multiple variants
associated with rheumatoid arthritis. Am J Hum Genet. 77:567–581.
2005.PubMed/NCBI View
Article : Google Scholar
|
|
33
|
Jagiello P, Aries P, Arning L, Wagenleiter
SEN, Csernok E, Hellmich B, Gross WL and Epplen JT: The PTPN22 620W
allele is a risk factor for Wegener's granulomatosis. Arthritis
Rheum. 52:4039–4043. 2005.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Onengut-Gumuscu S, Ewens KG, Spielman RS
and Concannon P: A functional polymorphism (1858C/T) in the PTPN22
gene is linked and associated with type I diabetes in multiplex
families. Genes Immun. 5:678–680. 2004.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Skórka A, Bednarczuk T, Bar-Andziak E,
Nauman J and Ploski R: Lymphoid tyrosine phosphatase (PTPN22/LYP)
variant and Graves' disease in a Polish population: Association and
gene dose-dependent correlation with age of onset. Clin Endocrinol
(Oxf). 62:679–682. 2005.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Wu H, Wan S, Qu M, Ren B, Liu L and Shen
H: The relationship between PTPN22 R620W polymorphisms and the
susceptibility to autoimmune thyroid diseases: An updated
meta-analysis. Immunol Invest. 2:438–451. 2022.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Thompson SD, Sudman M, Ramos PS, Marion
MC, Ryan M, Tsoras M, Weiler T, Wagner M, Keddache M, Haas JP, et
al: The susceptibility loci juvenile idiopathic arthritis shares
with other autoimmune diseases extend to PTPN2, COG6, and ANGPT1.
Arthritis Rheum. 62:3265–3276. 2010.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Viken MK, Amundsen SS, Kvien TK, Boberg
KM, Gilboe IM, Lilleby V, Sollid LM, Førre OT, Thorsby E, Smerdel A
and Lie BA: Association analysis of the 1858C/T polymorphism in the
PTPN22 gene in juvenile idiopathic arthritis and other autoimmune
diseases. Genes Immun. 6:271–273. 2005.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Cinek O, Hradsky O, Ahmedov G, Slavcev A,
Kolouskova S, Kulich M and Sumnik Z: No independent role of the
-1123 G>C and +2740 A>G variants in the association of PTPN22
with type 1 diabetes and juvenile idiopathic arthritis in two
Caucasian populations. Diabetes Res Clin Pract. 76:297–303.
2007.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Seldin MF, Shigeta R, Laiho K, Li H, Saila
H, Savolainen A, Leirisalo-Repo M, Aho K, Tuomilehto-Wolf E,
Kaarela K, et al: Finnish case-control and family studies support
PTPN22 R620 W polymorphism as a risk factor in rheumatoid
arthritis, but suggest only minimal or no effect in juvenile
idiopathic arthritis. Genes Immun. 6:720–722. 2005.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Di Y, Zhong S, Wu L, Li Y and Sun N: The
association between PTPN22 Genetic polymorphism and juvenile
idiopathic arthritis (JIA) susceptibility: An updated
meta-analysis. Iran J Public Health. 44:1169–1175. 2015.PubMed/NCBI
|
|
42
|
Lee YH, Bae SC and Song GG: The
association between the functional PTPN22 1858 C/T and MIF 173 C/G
polymorphisms and juvenile idiopathic arthritis: A meta-analysis.
Inflamm Res. 61:411–415. 2012.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Hinks A, Worthington J and Thomson W: The
association of PTPN22 with rheumatoid arthritis and juvenile
idiopathic arthritis. Rheumatology (Oxford). 45:365–368.
2006.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Baralle D and Baralle M: Splicing in
action: Assessing disease causing sequence changes. J Med Genet.
42:737–748. 2005.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Buratti E, Baralle M and Baralle FE:
Defective splicing, disease and therapy: Searching for master
checkpoints in exon definition. Nucleic Acids Res. 34:3494–3510.
2006.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Schoenemeyer A, Barnes BJ, Mancl ME, Latz
E, Goutagny N, Pitha PM, Fitzgerald KA and Golenbock DT: The
interferon regulatory factor, IRF5, is a central mediator of
toll-like receptor 7 signaling. J Biol Chem. 280:17005–17012.
2005.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Takaoka A, Yanai H, Kondo S, Duncan G,
Negishi H, Mizutani T, Kano SI, Honda K, Ohba Y, Mak TW and
Taniguchi T: Integral role of IRF-5 in the gene induction programme
activated by toll-like receptors. Nature. 434:243–249.
2005.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Miceli-Richard C, Comets E, Loiseau P,
Puechal X, Hachulla E and Mariette X: Association of an IRF5 gene
functional polymorphism with Sjgren's syndrome. Arthritis Rheum.
56:3989–3994. 2007.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Tada Y, Kondo S, Aoki S, Koarada S, Inoue
H, Suematsu R, Ohta A, Mak TW and Nagasawa K: Interferon regulatory
factor 5 is critical for the development of lupus in MRL/lpr mice.
Arthritis Rheum. 63:738–748. 2011.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Graham DS, Manku H, Wagner S, Reid J,
Timms K, Gutin A, Lanchbury JS and Vyse TJ: Association of IRF5 in
UK SLE families identifies a variant involved in polyadenylation.
Hum Mol Genet. 16:579–591. 2007.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Kristjansdottir G, Sandling JK, Bonetti A,
Roos IM, Milani L, Wang C, Gustafsdottir SM, Sigurdsson S, Lundmark
A, Tienari PJ, et al: Interferon regulatory factor 5 (IRF5) gene
variants are associated with multiple sclerosis in three distinct
populations. J Med Genet. 45:362–369. 2008.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Reykande SE, Rezaei A, Sadr M, Shabani M,
Varzaneh FN, Ziaee V and Rezaei N: Association of interferon
regulatory factor 5 (IRF5) gene polymorphisms with juvenile
idiopathic arthritis. Clin Rheum. 37:2661–2665. 2018.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Fedorov ES, Krylov MY, Salugina SO,
Samarkina EY and Latypova AN: STAT4 rs7574865 G/T and IRF5
rs2004640 G/T polymorphisms as markers of predisposition to
juvenile idiopathic arthritis. What can genetics give to understand
its heterogeneity? Modern Rheumatol J. 13:55–60. 2019.
|
|
54
|
Schindler C, Levy DE and Decker T:
JAK-STAT signaling: From interferons to cytokines. J Biol Chem.
282:20059–20063. 2007.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Kisseleva T, Bhattacharya S, Braunstein J
and Schindler CW: Signaling through the JAK/STAT pathway, recent
advances and future challenges. Gene. 285:1–24. 2002.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Chong WP, Ip WK, Wong WHS, Lau CS, Chan TM
and Lau YL: Association of interleukin-10 promoter polymorphisms
with systemic lupus erythematosus. Genes Immun. 5:484–492.
2004.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Shaw MH, Boyartchuk V, Wang S,
Karaghiosoff M, Ragimbeau J, Pellegrini S, Muller M, Dietrich WF
and Yap GS: A natural mutation in the Tyk2 pseudokinase domain
underlies altered susceptibility of B10.Q/J mice to infection and
autoimmunity. Proc Natl Acad Sci USA. 100:11594–11599.
2003.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Kyogoku C, Morinobu A, Nishimura K,
Sugiyama D, Hashimoto H, Tokano Y, Mimori T, Terao C, Matsuda F,
Kuno T and Kumagai S: Lack of association between tyrosine kinase 2
(TYK2) gene polymorphisms and susceptibility to SLE in a Japanese
population. Mod Rheumatol. 19:401–406. 2009.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Mohamadhosseini A, Mansouri R, Javinani A,
Ganjouei AA, Akhlaghi M, Aslani S, Hamzeh E, Jamshidi A, Ahmadzadeh
N and Mahmoudi M: Single nucleotide polymorphism of TYK2 gene and
susceptibility to rheumatoid arthritis in iranian population.
Avicenna J Med Biotechnol. 11:187–191. 2019.PubMed/NCBI
|
|
60
|
Graham D, Akil M and Vyse TJ: Association
of polymorphisms across the tyrosine kinase gene, TYK2 in UK SLE
families. Rheumatology (Oxford). 46:927–930. 2007.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Li P, Chang YK, Shek KW and Lau YL: Lack
of association of TYK2 gene polymorphisms in Chinese patients with
systemic lupus erythematosus. J Rheumatol. 38:177–178.
2011.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Zervou M, Dimopoulou DG, Eliopoulos E,
Trachana M, Pratsidou-Gkertsi P, Andreou A, Sidiropoulos P,
Spandidos DA, Garyfallos A and Goulielmos GN: Τhe genetics of
juvenile idiopathic arthritis: Searching for new susceptibility
loci. Mol Med Rep. 16:8793–8798. 2017.PubMed/NCBI View Article : Google Scholar
|