Introduction
Lung cancer is one of the most commonly occurring
malignancies in China and the world (1). The early symptoms of lung cancer, of
which extensive-stage small cell lung cancer is one of the
subtypes, include hemoptysis or paraneoplastic syndromes (2). Lung cancer has one of the highest
mortality rates among malignant tumors (3). When patients present with symptoms in
the respiratory tract or with recognized tumor-associated image
symptoms, this indicates that the lung cancer has already entered
into the advanced stages or that it has metastasized (4). Under these circumstances, the
survival rate of patients is <65%, with a poor prognosis
(5,6). Therefore, further studies on the
early stages of lung tumors are in high demand. Specifically,
detection of early-stage biomarkers and characterization of
prognostic markers would provide a theoretical basis for the
diagnosis and treatment of lung cancer.
Nuclear factor of activated T cells (NFAT) c1 is a
common cell transcription factor (7). As a member of the NFAT family, NFATc1
expression has been found to be upregulated in number of solid
tumors and hematological malignancies in humans (8). It is has also been reported to be of
functional importance in a diverse array of functions, including
tumor cell differentiation, cell proliferation, apoptosis,
invasion, migration and angiogenesis, whilst also enabling tumor
cells to escape from detection by the immune system (9-12).
Specifically, NFATc1 has been demonstrated to promote cell invasion
and tumor metastasis in breast, colon and ovarian cancer (13,14).
In addition, NFATc1 has been found to be highly expressed in
pancreatic cancer tissues and cell lines, where the interaction
between NFATc1 and STAT3 promoted the severity of malignancy
(15). In ovarian cancer cells,
NFATc1 has been reported to promote both proliferation and
tumorigenesis by activating the ERK/p38 MAPK signaling pathway
(16). A number of studies have
provided evidence that NFATc1 may serve an important role in
tumorigenesis (8,14,16).
However, to the best of our knowledge, the functional role of
NFATc1 in the development and progression of lung cancer remain to
be fully elucidated. Therefore, the aim of the present study was to
determine whether NFATc1 has a role in regulating the
proliferation, invasion, migration and apoptosis of lung cancer
cells in vitro. Another aim of the present study was to
elucidate the underlying mechanism through which NFATc1 may exert
its regulatory function, specifically which cell signaling pathways
are involved.
Materials and methods
Acquisition of human specimens
All human specimens were obtained from the
Department of Thoracic Surgery of the Harbin Medical University
Cancer Hospital (Harbin, China) from August 2016 and July 2018. A
total of 30 samples from patients with lung cancer were collected,
all of which were verified using pathology. Inclusion criteria were
as follows: i) All patients with lung cancer were diagnosed based
on pathological confirmation; ii) the patients' own conditions
allowed surgical treatment; and iii) all subjects were informed
about the clinical study and signed an informed consent form.
Exclusion criteria were as follows: i) Combination of other types
of malignant tumors; ii) combination of severe cardiac, pulmonary,
hepatic and renal impairment; iii) Combination of various types of
hematological diseases; iv) systemic inflammatory diseases such as
infections and autoimmune diseases; and v) history of surgery
within the last 3 months. There were 21 males and 9 females, and
the average age of patients were 45.52±12.76 (range, 24-67 years
old). The lung cancer tissues and adjacent non-cancerous tissues
(≥2 cm from the tumor) were obtained during surgery for the
immunohistochemical and western blot analysis. The patients and
their families consented to participate and provided written
informed consent for use. All participants provided the relevant
informed consent forms, and the study protocol (approval no.
KY2021-33) was approved by the Medical Ethics Committee of the
Harbin Medical University Cancer Hospital.
Immunohistochemical staining
The tissues were fixed in 4% PFA for >24 h, and
embedded in paraffin after gradient dehydration within 1 week, 70%
alcohol 1 h, 80% alcohol 1 h, 90% alcohol 1 h, 95% alcohol 1 h
twice, absolute ethanol 30 min twice, xylene 30 min twice, paraffin
1 h twice. The tissues were cut into 4-µm sections. The slides were
dewaxed to water through xylene for 20 min twice, absolute ethanol
for 10 min twice, 95% alcohol for 5 min, 90% alcohol for 5 min, 80%
alcohol for 5 min and 70% alcohol for 5 min and distilled water for
washing. EDTA antigen repair buffer (pH 9.0) was used to repair the
antigen in the microwave oven. Use a histochemical pen to draw a
circle around the tissues, drop hydrogen peroxide into the circle
to cover the tissues incubate for 20 min at room temperature in
dark, wash the slides with PBS thrice. Incubated with 3% BSA at
room temperature for 30 min, then incubated with primary antibodies
NFAT2 (1:100; cat. no. ab2796; Abcam) overnight at 4˚C, washed with
PBS thrice. Followed by peroxidase-conjugated secondary antibody
(HRP-goat anti-mouse; 1:200; cat. no. AS1106; Wuhan Aspen
Biotechnology, Co., Ltd.) for 1 h at room temperature, washed with
PBS for 3 times. Then the tissue sections were stained with
diaminobenzidine for 5 min, washed with water, and counterstained
with hematoxylin for 3 min, all at room temperature. A light
microscope (Olympus Corporation) was used to capture images of the
representative areas.
Cell culture
The H1975 human non-small cell lung cancer cell line
(cat. no. CRL-5908™), the A549 human lung carcinoma cell
line (cat. no. CRM-CCL-185™), the NCI-H1299 human
non-small cell lung cancer cell line (cat. no.
CRL-5803™) and 293T cell (cat. no. CRL-3216™)
were obtained from American Type Culture Collection. The cells were
cultured as monolayers in RPMI-1640 or DMEM (Gibco; Thermo Fisher
Scientific, Inc.) medium including 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.), with 100 µg/ml streptomycin and 100 U/ml
penicillin (Thermo Fisher Scientific, Inc.). The cells were
cultured at 37˚C in a humidified atmosphere of 95% air and 5%
CO2.
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from the A549 and NCI-H1299
cells using Invitrogen® TRIzol™ (cat. no.
15596018; Thermo Fisher Scientific, Inc.). cDNA synthesis and
amplification were subsequently performed using a
Fermentas® RevertAid™ First Strand cDNA
Synthesis Kit (cat no. K16215; Thermo Fisher Scientific, Inc.) in
accordance with the manufacturer's protocols. FastStart™
Universal SYBR® Green (cat no. 4913914001; Roche
Diagnostics) was used for qPCR analysis, and the reactions were
performed according to a two-step method, with the following
thermocycling conditions: 50˚C for 2 min and 95˚C for 10 min;
followed 40 cycles of 95˚C for 10 sec and 60˚C for 30 sec. Primers
for qPCR were designed as follows: NFATc1 forward,
5'-CCATGAAGTCAGCGGAGGAA-3' and reverse, 5'-GAGGTCTGAAGGTTGTGGCA-3';
and GAPDH forward, 5'-GAAGGTGAAGGTCGGAGTCA-3 and reverse,
5'-TTGAGGTCAATGAAGGGGTC-3'. GAPDH was used as an internal control
and the relative expression levels of the target gene were
calculated using the 2-∆∆Cq method (17).
Short hairpin RNA (shRNA/sh)
knockdown
A lentivirus system was used to knock down NFATc1
expression. The plvx-shRNAs [short hairpin negative control
(shCtrl) and plvx-shNFATc1] were obtained from Shanghai Jikai Gene
Chemical Technology Co., Ltd. The target sequences of shNFATc1 and
shCtrl used in the present study were as follows: shNFATc1,
5'-GAATCCTGAAACTCAGAAA-3'; and shCtrl, 5'-TTCTCCGAACGTGTCACGT-3'.
The plasmid backbone used was GV493 which proved by Shanghai Jikai
Gene Chemical Technology Co., Ltd. The lentivirus system used was
the 3rd system, which included three packaging vectors (Rev, VSV-G
and RRE) and one Lenti-X plasmid, carrying the target sequence and
the control sequence. The 293T cell line was selected for
optimizing lentivirus production. In the experiment, the packing
vector (Rev, VSV-G and RRE) and the core plvx-shRNA were packaged
with lentivirus in a ratio of 1:2:4:8. Lentiviral particles were
collected by filtering with 0.45 µm then using centrifugation at
1,000 x g for 5 min at room temperature. Subsequently, the viruses
were used to infect the target A549 (MOI=10) and NCI-H1299 (MOI=5)
cells at the ratio of 1:1 for >16 h. The lentiviral transfection
efficiency was assessed after 72 h to confirm that the target gene
had been knocked down by RT-qPCR. Then 1 µg/ml puromycin was used
for selection, and 0.5 µg/ml puromycin was used for maintenance.
Time interval between transduction and subsequent experimentation
was one week.
Cell proliferation assay
An MTT assay was used to observe and compare the
proliferation of the cells. A total of 2x103 A549 and
NCI-H1299 cells were plated into 96-well plates. Then the cell
proliferation was assessed. Cells were incubated for 4 h in 20 µl 5
mg/ml MTT at 37˚C. The color was then developed by incubating the
cells in 100 µl DMSO for 2-3 min, and the absorbance was detected
at a wavelength of 490 nm using a microplate reader. Repeat
measurements were taken for 5 consecutive days. The results were
obtained from three independently performed experiments and
calculated.
Wound healing assay
A549 and NCI-H1299 cells in which NFATc1 expression
was knocked down were used for the wound healing experiments. After
incubation for 48 h at 37˚C, the cells were treated with 0.25%
trypsin, counted and plated at a density of 5x106
cells/ml in six-well plates. The cells were then incubated
overnight at 37˚C, yielding 100% confluent monolayers for wounding.
Wounds were created using a 10 µl pipette tip and images were
captured immediately and at 24 h after wounding at 37˚C, the cells
were cultured in RPMI-1640 medium with 0.5% FBS. Images were
captured using a fluorescence microscope (Olympus Corporation), and
cell area was analyzed at 0 and 24 h to calculate the migration
rate (%). The plasmid backbone used had a green fluorescence, so
images were captured using a fluorescence microscope at the same
point. Experiments were performed in triplicate, and each
experiment was repeated three times.
Colony formation assay
A549 and NCI-H1299 cells in which NFATc1 expression
was knocked down. After incubation for 48 h at 37˚C, the cells were
trypsinized with 0.25% trypsin, resuspended in the medium and
counted. The cells were then re-seeded (1,000 cells per well) and
incubated for a further 2 weeks at 37˚C. When the colonies became
visible to the naked eye, the medium was removed from the dishes
and the plates were washed once with PBS. Cell immobilization with
4% paraformaldehyde for 10 min at room temperature.
Colonies were then stained with 0.1% (w/v) crystal
violet for 6 min at room temperature on a rocking platform. The
dishes were subsequently rinsed three times with PBS and air-dried.
Images were captured using a camera, and the number of colonies
were analyzed using Image J software (National Institutes of
Health).
Cell migration and invasion
assays
Transwell assays (Corning, Inc.) were used to
measure the migration and invasion of the cells.
Lentivirus-transfected A549 and NCI-H1299 cells were cultured for
48 h at 37˚C. Before seeding, the Transwell chambers of 24-well
plates with 8 µm aperture were soaked with PBS for 5 min (the
invasion assay experiments required one more step: 80 µl
Matrigel™ was placed in the small chambers, which were
incubated in an incubator at 37˚C for 30 min). A total of
105 cells resuspended in RPMI-1640 with no FBS were
inoculated into the Transwell chambers of the 24-well plates. A
total of 0.7 ml RPMI-1640 medium containing 10% FBS was added to
the lower chamber of the 24-well plate, and the plate was incubated
at 37˚C for 48 h. Subsequently, a 4% formaldehyde solution (1
ml/well) was added to fix the cells in the wells for 10 min at room
temperature, after which time the adhering cells were removed,
followed by washing once with PBS. Subsequently, the cells were
stained with 0.1% crystal violet (1 ml/well) for 6 min at room
temperature, washed three times with PBS and dried. Images were
captured using a light microscope (Olympus Corporation), four
fields of view were imaged per chamber, and the number of migrating
or invading cells were analyzed using Image J software.
Cell cycle and apoptosis
Flow cytometric analysis was performed to assess the
effects of NFATc1 on cell cycle progression and apoptosis in A549
and NCI-H1299 cells. For cell cycle analysis, after incubation for
48 h at 37˚C, 3x106 cells (per group) were washed once
with PBS and then fixed in 70% ethanol overnight at -20˚C, prior to
being stained with propidium iodide buffer (50 mg/ml) in dark on
ice for 30 min. The numbers of cells were detected by flow
cytometry (BD Accuri™ C6 Plus; BD Biosciences) and
analyzed using Cell Quest software (version 5.1; BD
Biosciences).
For the detection of apoptosis, after incubation for
48 h at 37˚C, 1x106 cells were washed with PBS and then
once in 1X binding buffer. A total of 5 µl
allophycocyanin-conjugated Annexin V (cat. no. 88-8007-74;
Invitrogen; Thermo Fisher Scientific, Inc.) was added to the cells
and incubated for 15 min at room temperature, after washing and
resuspension in 1X binding buffer. The numbers of apoptotic cells
were detected by flow cytometry (BD Accuri™ C6 Plus; BD
Biosciences) and analyzed using Cell Quest software (version 5.1;
BD Biosciences).
Western blot analysis
Lentivirus-transfected A549 and NCI-H1299 cells were
harvested using RIPA lysis buffer (cat. no. P0013B; Beyotime
Institute of Biotechnology) supplemented with protease and
phosphatase inhibitors. The cell lysates were incubated on ice for
30 min to allow for complete homogenization to take place. After
having centrifuged the lysates at 12,000 x g at 4˚C for 10 min, the
proteins in the supernatant were collected and quantified using a
BCA protein kit (cat. no. P0010S; Beyotime Institute of
Biotechnology). A total of 50-100 µg protein were separated by 10
or 15% SDS-PAGE and then transferred onto PVDF microporous
membranes. The blots were blocked in 5% skimmed milk for 1 h at
room temperature, followed by incubation with primary antibodies
against ERK (cat. no. 9107; 1:2,000; Cell Signaling Technology,
Inc.), E-cadherin (cat. no. 14472; 1:500; Cell Signaling
Technology, Inc.), phosphorylated (p)-ERK (cat. no. 4376; 1:1,000;
Cell Signaling Technology, Inc.), N-cadherin (cat. no. ab18203;
1:500; Abcam), Bax (cat. no. ab32503; 1:2,000; Abcam), p38 (cat.
no. 8690; 1:3,000; Cell Signaling Technology, Inc.), cleaved
caspase-3 (cat. no. 9664; 1:500; Cell Signaling Technology, Inc.),
p-p38 (cat. no. 4631; 1:500; Cell Signaling Technology, Inc.),
c-Myc (cat. no. ab32072; 1:1,000; Abcam), CDK4 (cat. no. ab137675;
1:1,000; Abcam), NFAT2 (cat. no. ab2796; 1:1,000; Abcam) and GAPDH
(cat. no. ab37168; 1:5,000; Abcam) at 4˚C with gentle shaking
overnight. The next day, after washing the membranes five or six
times (5-10 min per wash) with TBS-T (0.05% Tween-20), the blots
were incubated with the corresponding HRP-conjugated secondary
antibodies (HRP-Goat anti Rabbit; cat. no. AS1107; 1:10,000; Wuhan
Aspen Biotechnology, Co., Ltd.; HRP-Goat anti Mouse; cat. no.
AS1106; 1:10,000; Wuhan Aspen Biotechnology, Co., Ltd.) at room
temperature for 1 h. After washing again and incubating the
membranes with BeyoECL Plus (cat. no. P0018S, Beyotime Institute of
Biotechnology), the blots were visualized using an ECL imaging
system, and the relative protein levels were calculated using
ImageLab software (version 5.2.1; Bio-Rad Laboratories, Inc.). The
levels of the protein of interest were normalized against those of
GAPDH.
Statistical analysis
The data were analyzed using SPSS 22.0 software (IBM
Corp.). All experiments were repeated at least three times. The
measured data are presented as the mean ± standard deviation.
Comparisons of three groups of samples were made using one-way
ANOVA followed by Dunnett's test, whereas comparisons of two groups
of samples were analyzed using an unpaired Student's t test. The
MTT results were analyzed by two-way ANOVA followed by Sidak's
multiple comparisons test. P<0.05 was considered to indicate a
statistically significant difference.
Results
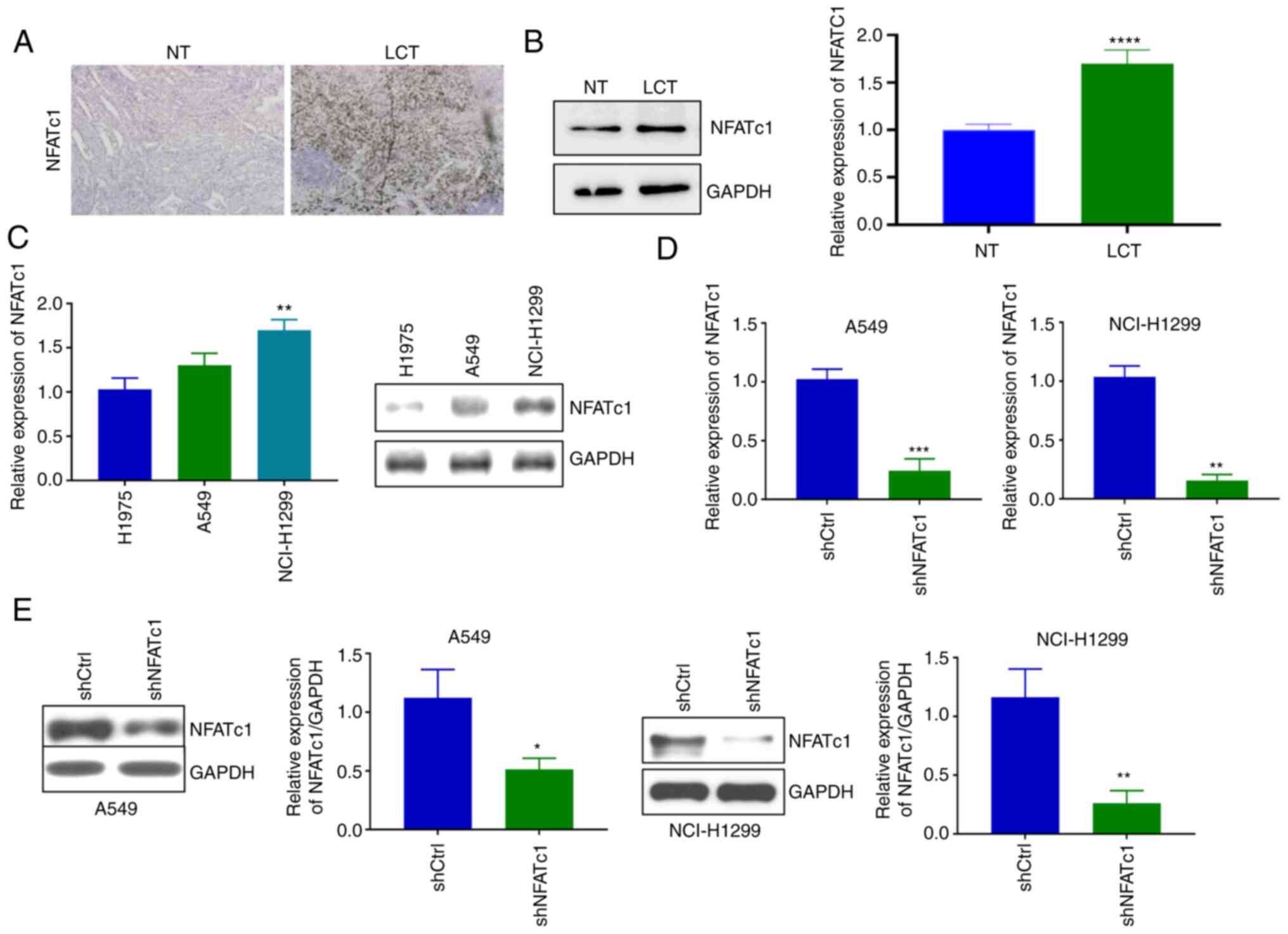
Downregulation of NFATc1 expression in
lung cancer cells by lentivirus infection
First, the expression levels of the transcription
factor NFATc1 in the Cancer and adjacent tissues of lung cancer
were examined. The results of both immunohistochemistry (Fig. 1A) and western blot analysis
(Fig. 1B) demonstrated that NFATc1
expression was markedly increased in the lung cancer tissues
compared with in normal lung tissues. Subsequent experiments
revealed that NFATc1 expression was upregulated in A549 and
NCI-H1299 cells compared with H1975 cells, but the difference
between A549 and H1975 cells was not significant (Fig. 1C). The present study aimed to
elucidate the mechanistic role of NFATc1 in lung cancer. Therefore,
NFATc1 expression was knocked down to explore its function of the
gene. The two high NFATc1-expressing cell lines (A549 and
NCI-H1299) were selected for the subsequent experiments. Compared
with those in the shCtrl group, the mRNA (Fig. 1D) and protein (Fig. 1E) expression levels of NFATc1 in
both the A549 and NCI-H1299 cell lines were significantly decreased
following lentivirus infection. After verifying the efficiency of
NFATc1 knockdown, this shNFATc1-encoding lentivirus was used for
the subsequent experiments.
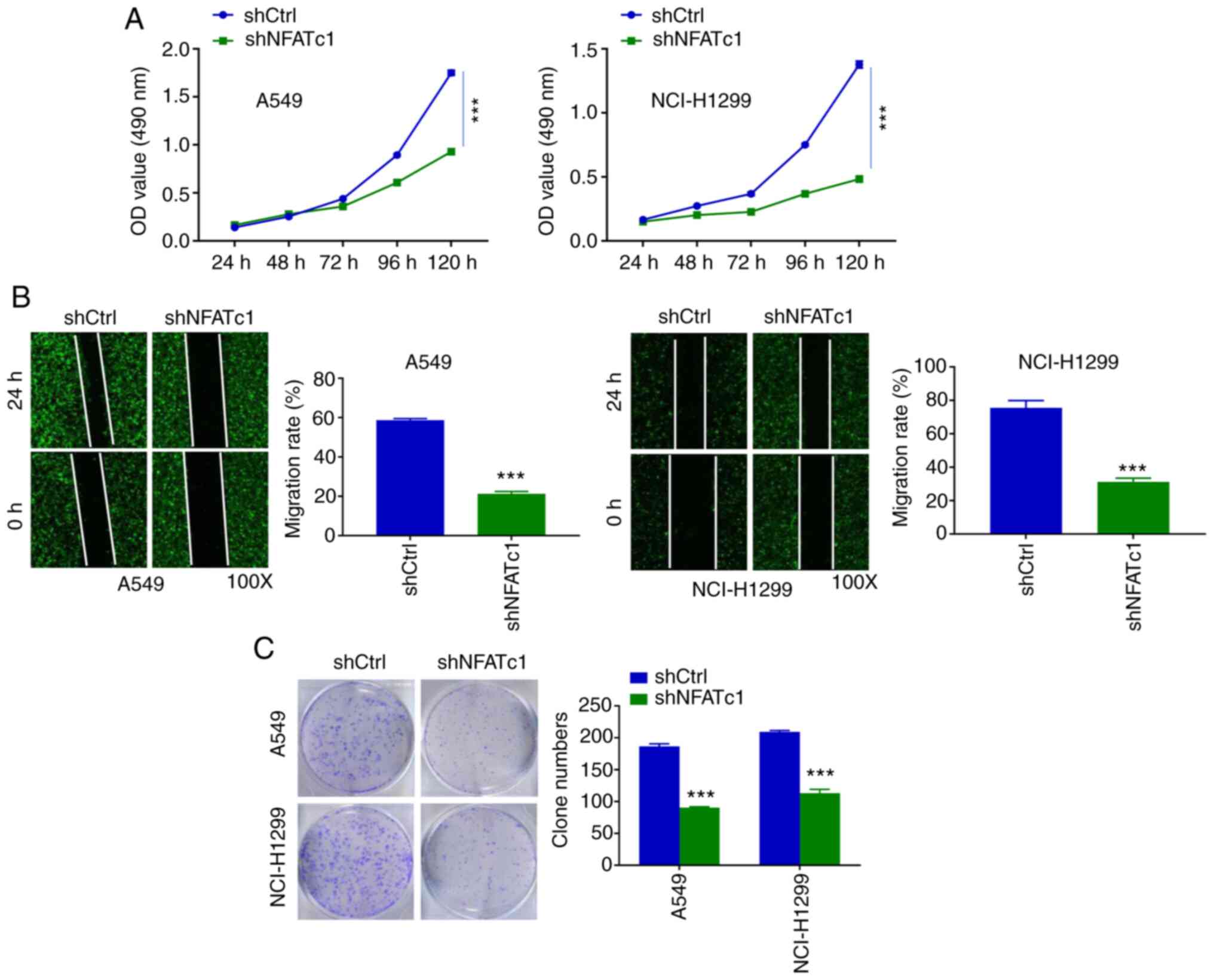
Knockdown of NFATc1 inhibits the
proliferation and colony formation of lung cancer cells
After NFATc1 knockdown, the proliferation of the
lung cancer cells was investigated. In these experiments, cell
viability was recorded for once per day for 5 days. Over the course
of the first 3 days, differences in cell viability were not readily
detectable. However, after 3 days, the differences could be
observed more easily. When compared with that in the shCtrl group,
NFATc1 knockdown caused significant inhibition of the proliferation
of A549 (from the third day) and NCI-H1299 cells (from the second
day) (Fig. 2A). Subsequently,
wound healing experiments were performed. The gap was found to be
wider in the shNFATc1 group compared with in the shCtrl group,
irrespective of whether A549 or NCI-H1299 cells were used (Fig. 2B). Similarly, the cell colony
numbers in the shNFATc1 group were significantly lower compared
with those in the shCtrl group for both cell lines (Fig. 2C).
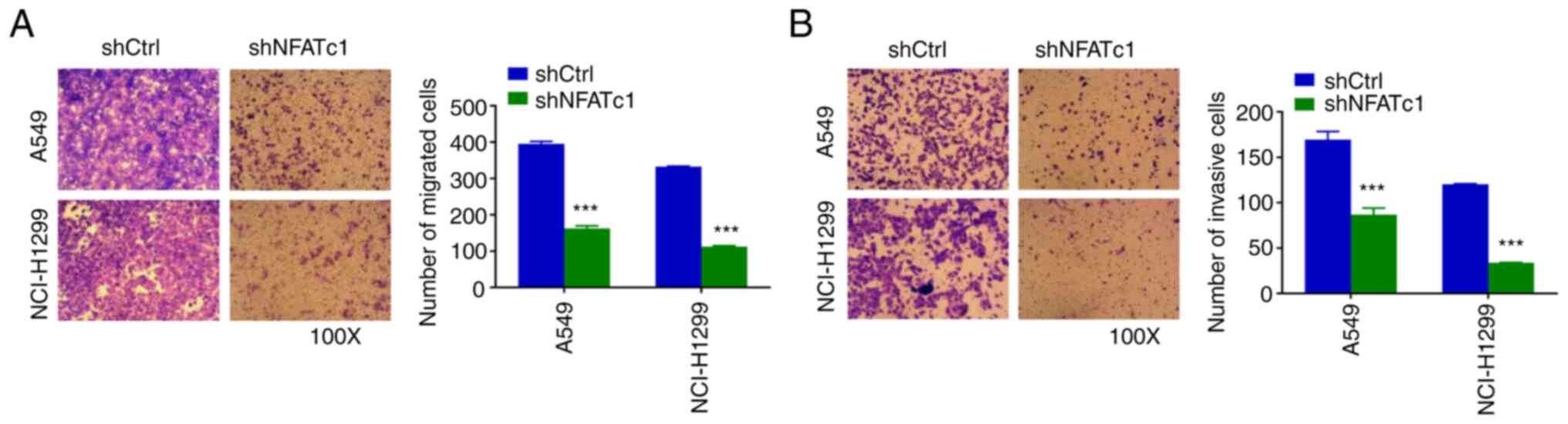
Knockdown of NFATc1 inhibits cell
migration and invasion of lung cancer cells
Transwell migration and invasion assays were
subsequently used to assess the effect of NFATc1 on cell migration
and invasion. The number of migrated cells in the shNFATc1 group
was found to be significantly decreased compared with that in the
shCtrl group (Fig. 3A).
Furthermore, to assess the invasive capability of the cells, these
Transwell experiments were repeated after Matrigel coating. The
number of invasive cells in the shNFATc1 group was likewise found
to be significantly lower compared with that in the shCtrl group
(Fig. 3B).
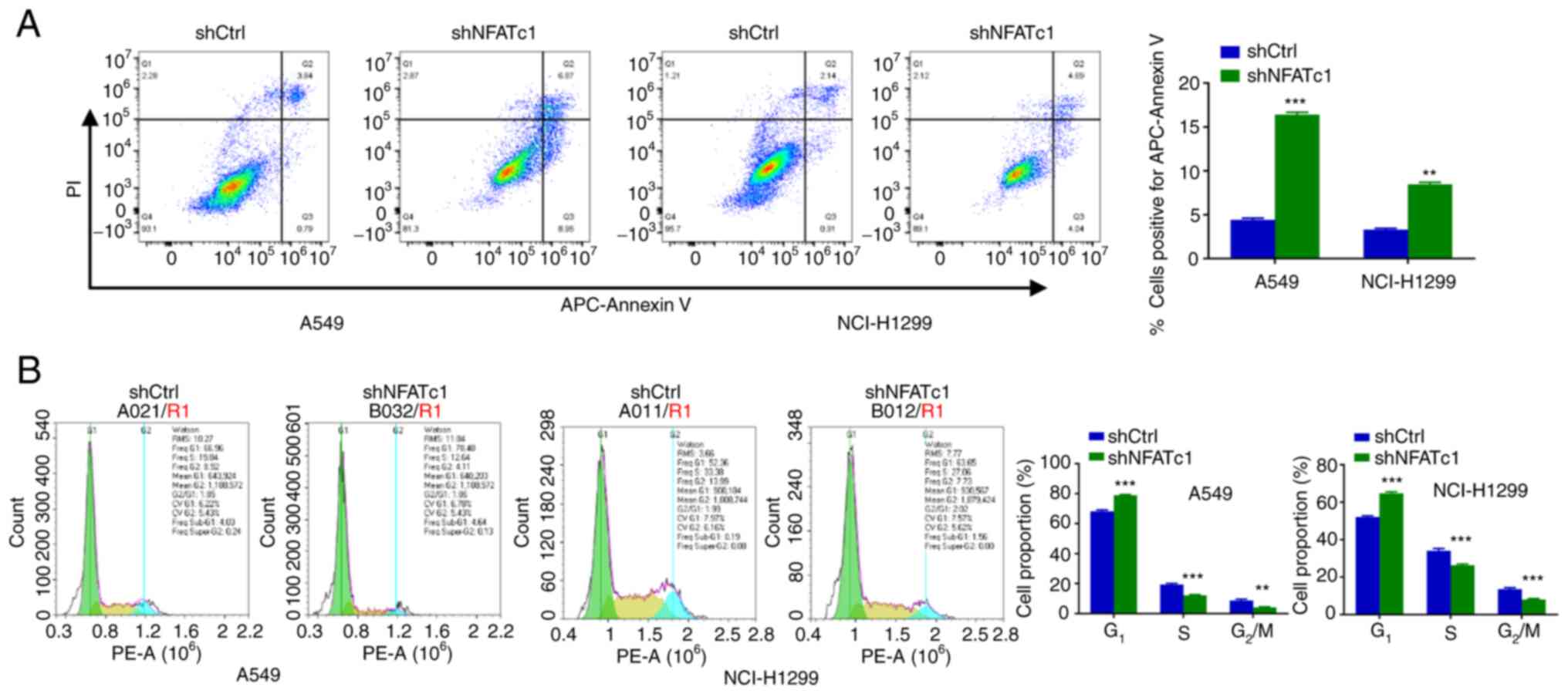
Knockdown of NFATc1 promote apoptosis
and inhibits the cell cycle progression in lung cancer cells
The effect of NFATc1 on apoptosis in lung cancer
cells was explored. The number of apoptotic cells was significantly
increased following NFATc1 knockdown compared with in the shCtrl
group (Fig. 4A). Changes in the
cell cycle were subsequently investigated. NFATc1 knockdown
resulted in a significant increase in the percentage of cells at
G1 phase and a significant decrease in the percentage of
cells at the S and G2/M phases, compared with those in
the shCtrl group (Fig. 4B). This
suggest that knockdown of NFATc1 expression in A549 and NCI-H1299
cells led to cell arrest at the G1 phase, where the
cells were unable to pass through the G1/S
checkpoint.
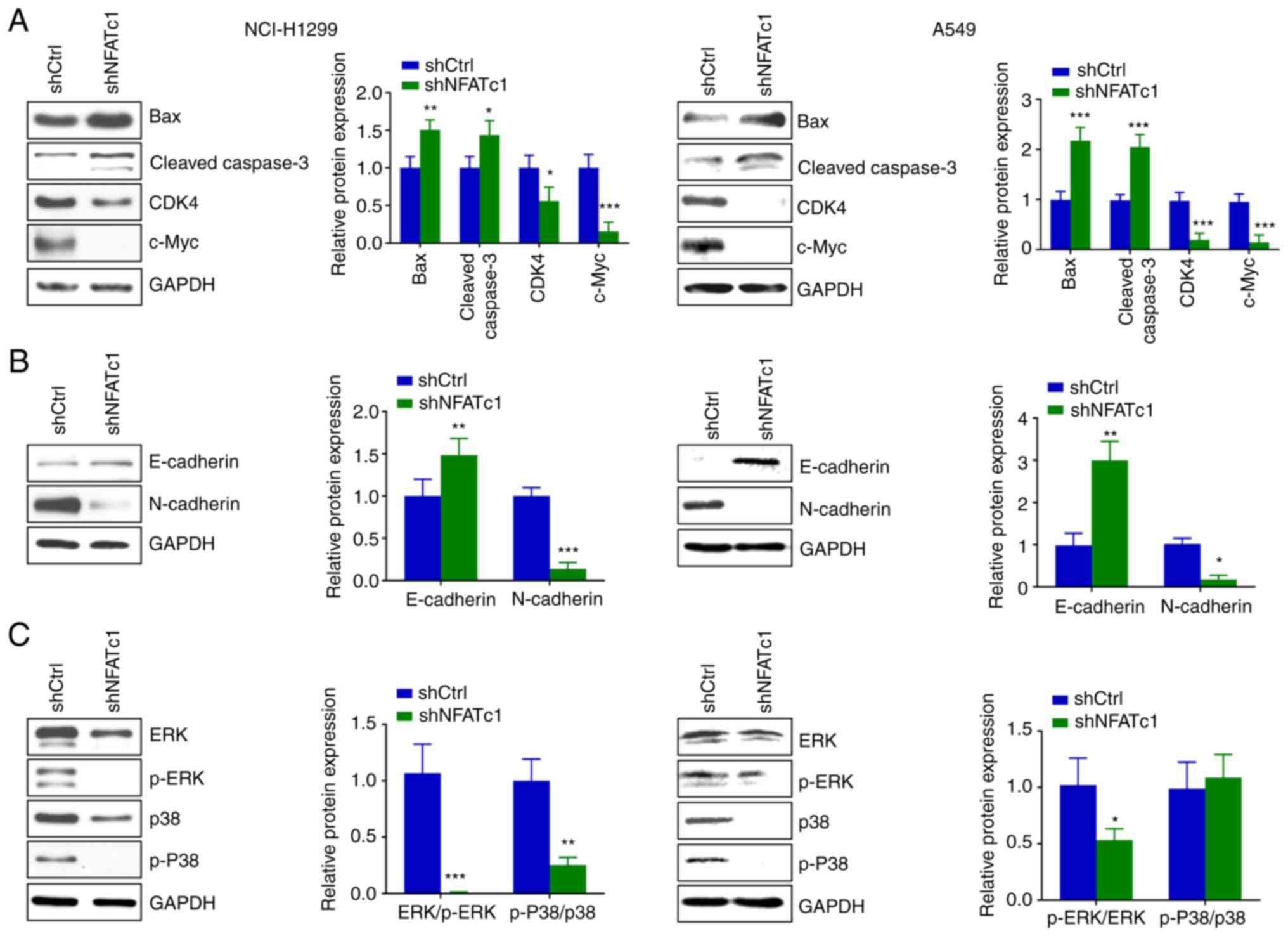
Knockdown of NFATc1 expression induces
cell apoptosis and inhibits the MAPK and EMT signaling pathways in
lung cancer cells
Western blotting revealed that knockdown of NFATc1
led to a significant increase in the protein expression levels of
Bax and cleaved caspase-3, while the expression levels of CDK4 and
c-Myc were significantly decreased, compared with those in the
shCtrl group (Fig. 5A).
Furthermore, the expression levels of EMT signaling pathway markers
were detected. Knockdown of NFATc1 expression caused a significant
increase in the protein expression levels of E-cadherin, whereas
the protein expression levels of N-cadherin were significantly
decreased, compared with those in the shCtrl group (Fig. 5B). The effects of knockdown of
NFATc1 expression on the MAPK signaling pathway were subsequently
examined. The protein expression levels of p-ERK/ERK and p-P38/p38
in the shNFATc1 group were found to be reduced compared with those
in the shCtrl group (Fig. 5C).
Discussion
NFATc1 is an important transcription factor and a
previous study has been performed to assess its role in
lymphocytes, NFATc1 are dispensable for inflammatory reactivity but
are required for effector differentiation in T cells (18). Accumulating evidence has
demonstrated that NFATc1 can mediate important regulatory roles in
mammalian tissues and in various types of malignant tumor cells
(19). At present, lung cancer is
one of the most commonly occurring malignant tumors, which has the
highest mortality rate worldwide. Even after the patients have been
treated with the standard therapies, the survival rate continues to
remain low, with a poor prognosis (5,6).
Therefore, the aim of the present study was to assess whether
NFATc1 can serve a role as a therapeutic biomarker. NFATc1 has been
previously shown to regulate the expression of DNA damage-induced
apoptosis suppressor in both pancreatic cancer and lung cancer
cells (20). Furthermore, NFATc1
exerts an important role in the activation of cytotoxic functions
in T cells (21). Based on these
findings, it was hypothesized that NFATc1 may also be a suitable
biomarker for lung cancer, providing a theoretical basis for its
diagnosis and treatment.
In the present study, a series of experiments
revealed that NFATc1 protein expression could be decreased at least
for periods up to 48 h, where knockdown of NFATc1 expression led to
the inhibition of the proliferation, invasion and migration of lung
cancer cells. In breast cancer and ovarian cells, similar results
were obtained (14,22). Following the inhibition of NFATc1
activity, cell proliferation and survival rates were markedly
inhibited (14,23). The abnormal expression of NFATc1
may lead to changes in the morphology of cells to enhance the
invasive ability of the cells in lung cancer and breast cancer
(24). In addition, NFATc1 has
been previously revealed to promote urothelial tumorigenesis
(25) and bladder cancer cell
proliferation (26). The extent of
apoptosis and cell cycle progression was also analyzed after the
lentiviral transfection in the lung cancer cells for 48 h. Although
this time period of transfection may have been too short to observe
unequivocal results, compared with that in the shCtrl group, the
shNFATc1 group exhibited an increase in the apoptotic rate. In
liver cancer cells, knockdown of NFATc1 expression resulted in the
blockade of the cell cycle at the G1/S phase (27), a result that was consistent with
the present findings in lung cancer cells.
In urinary tract urothelial carcinoma (UUTUC), a
marked increase in NFATc1 expression was observed when compared
with that in the nonneoplastic urothelium (28). In addition, a strong NFATc1
expression was markedly associated with poorer outcomes for
patients with UUTUC (28). NFATc1
is a central regulator of pancreatic cancer cell plasticity, it can
mediate NFATc1/Sox2 signaling, which holds therapeutic promise in
patients with differentiated and metastatic pancreatic cancer
(29). Considering these
observations, it is likely that NFATc1 serves a key role in a
variety of cancer types.
It has been extensively reported that NFATc1 serves
an important role in numerous signal transduction pathways,
including those associated with apoptosis, EMT and the MAPK
signaling pathway connected with tumorigenesis and development
(30,31). In addition, a previous study
revealed that NFATc1 can upregulate c-Myc expression through
activation of the ERK/p38 MAPK signaling pathway in ovarian cancer
(32). In addition, NFATc1 was
demonstrated to be recruited to the promoter of cyclooxygenase 2
(COX-2) and enhanced COX-2 transcriptional activation (33). The expression levels of COX-2 are
increased in various types of tumors, including glioblastoma
multiforme (34). MMP-2 and MMP-9
were decreased in the NFATc1-silenced cells (35). Another study previously
demonstrated that NFATc1 regulate cell proliferation, migration and
invasion by inhibiting c-Myc and pyruvate kinase M2 expression in
prostate cancer (36). c-Myc is a
proto-oncogene that serves key roles in a range of tumors and may
be regulated by NFATc1 activity. In the present study, it was
demonstrated that silencing NFATc1 expression led to the inhibition
of migration and invasion by lung cancer cells. In addition, EMT
was activated and cell apoptosis was promoted, findings that were
consistent with those of a study by He and Lu (37).
The MAPK/ERK signaling pathway serves an important
role in cell proliferation, differentiation, migration and
apoptosis (38). The ERK family of
proteins mainly participate in the processes of proliferation and
differentiation, whereas the p38 MAPK family not only have an
influence on inflammation and the stress response but can also
regulate the normal process of apoptosis (39). The protein expression levels of ERK
and p38 may not change during the time when either cells or
osteosarcoma are treated with drugs (40,41).
However, if the mRNA expression of certain components is altered,
then the expression levels of the ERK and p38 proteins may also be
affected by the components in the MAPK/ERK signaling pathway,
leading to changes in the expression levels of ERK and p38. Yang
et al (42) found that the
protein levels of ERK, p-ERK, p38 and p-p38 were all decreased when
the activation of aquaporin 5 was inhibited in human glioma cells.
Upon overexpressing NFATc1 in the SKOV3 and CaOV3 ovarian cancer
cell lines, the phosphorylation levels of ERK1/2 and p38 were found
to be increased (16). In the
present study, it was demonstrated that silencing NFATc1 not only
reduced p-ERK/ERK and p-P38/p38 protein ratio.
Overall, based on the experiments performed in the
present study, it was demonstrated that NFATc1 serves a key role in
the proliferation of lung cancer. NFATc1 was also found to be
involved in the regulation of the MAPK signaling pathway, thereby
serving a key role in lung cancer cell signaling. However, there
were number of limitations associated with the present study. Only
NFATc1 was knocked down to explore its function. It is necessary to
perform the associated overexpression experiments to further
substantiate these conclusions. Therefore, overexpression
experiments should form part of the future studies in lung cancer.
Other experiments in the future should include in vivo
assays to obtain further confirmatory evidence. Through a more
detailed study of the underlying mechanism of NFATc1 and its role
in lung cancer, these studies may provide a potential therapeutic,
treatment and target biomarker of lung cancer.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by The Fundamental
Research Funds for the Provincial Universities (grant nos.
2018-KYYWF-0539 and 2020-KYYWF-1469) and Scientific Research
Subject of Heilongjiang Health Committee (grant no. 2019-061).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FR, KZ and LC guarantee the integrity of the entire
study. FR, KZ and LC conceived and designed the study, and defined
the intellectual content. YW, FZ and SP performed the literature
research. KZ, FZ and SP performed the clinical studies. FR, KZ and
YW performed the experimental studies. FR, KZ, YW, FZ and SP
performed data acquisition and analysis. KZ, YW and FZ performed
statistical analysis. FR and KZ performed manuscript preparation
and editing. LC reviewed the manuscript. FR and LC confirm the
authenticity of all the raw data. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Medical Ethics
Committee of the Harbin Medical University Cancer Hospital
(approval no. KY2021-33; Harbin, China). The patients and their
families provided written informed consent for the use of specimens
for the present study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Miller KD, Nogueira L, Devasia T, Mariotto
AB, Yabroff KR, Jemal A, Kramer J and Siegel RL: Cancer treatment
and survivorship statistics, 2022. CA Cancer J Clin. 72:409–436.
2022.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Hage R, de la Rivière AB, Seldenrijk CA
and van den Bosch JM: Update in pulmonary carcinoid tumors: A
review article. Ann Surg Oncol. 10:697–704. 2003.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Viale PH: The American cancer society's
facts & figures: 2020 edition. J Adv Pract Oncol. 11:135–136.
2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Collins LG, Haines C, Perkel R and Enck
RE: Lung cancer: Diagnosis and management. Am Fam Physician.
75:56–63. 2007.PubMed/NCBI
|
|
5
|
Miller KD, Nogueira L, Mariotto AB,
Rowland JH, Yabroff KR, Alfano CM, Jemal A, Kramer JL and Siegel
RL: Cancer treatment and survivorship statistics, 2019. CA Cancer J
Clin. 69:363–385. 2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Herbst RS, Morgensztern D and Boshoff C:
The biology and management of non-small cell lung cancer. Nature.
553:446–454. 2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Zhao Q, Wang X, Liu Y, He A and Jia R:
NFATc1: Functions in osteoclasts. Int J Biochem Cell Biol.
42:576–579. 2010.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Qin JJ, Nag S, Wang W, Zhou J, Zhang WD,
Wang H and Zhang R: NFAT as cancer target: Mission possible?
Biochim Biophys Acta. 1846:297–311. 2014.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Liang Q, Wang Y, Lu Y, Zhu Q, Xie W, Tang
N, Huang L, An T, Zhang D, Yan A, et al: RANK promotes colorectal
cancer migration and invasion by activating the
Ca2+-calcineurin/NFATC1-ACP5 axis. Cell Death Dis.
12(336)2021.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Jiang W, Rixiati Y, Huang H, Shi Y, Huang
C and Jiao B: Asperolide A prevents bone metastatic breast cancer
via the PI3K/AKT/mTOR/c-Fos/NFATc1 signaling pathway. Cancer Med.
9:8173–8185. 2020.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Song J, Zou D, Zhao X, Chen Y, Lv F, Wang
S, Sui D, Han Q, Yang C, Wang X, et al: Bufalin inhibits human
diffuse large B-cell lymphoma tumorigenesis by inducing cell death
through the Ca2+/NFATC1/cMYC pathway. Carcinogenesis. 42:303–314.
2021.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Klein-Hessling S, Muhammad K, Klein M,
Pusch T, Rudolf R, Flöter J, Qureischi M, Beilhack A, Vaeth M,
Kummerow C, et al: NFATc1 controls the cytotoxicity of
CD8+ T cells. Nat Commun. 8(511)2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Jauliac S, López-Rodriguez C, Shaw LM,
Brown LF, Rao A and Toker A: The role of NFAT transcription factors
in integrin-mediated carcinoma invasion. Nat Cell Biol. 4:540–544.
2002.PubMed/NCBI View
Article : Google Scholar
|
|
14
|
Li L, Duan Z, Yu J and Dang HX: NFATc1
regulates cell proliferation, migration, and invasion of ovarian
cancer SKOV3 cells in vitro and in vivo. Oncol Rep. 36:918–928.
2016.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Baumgart S, Chen NM, Siveke JT, König A,
Zhang JS, Singh SK, Wolf E, Bartkuhn M, Esposito I, Heßmann E, et
al: Inflammation-induced NFATc1-STAT3 transcription complex
promotes pancreatic cancer initiation by KrasG12D. Cancer Discov.
4:688–701. 2014.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Xu W, Gu J, Ren Q, Shi Y, Xia Q and Wang
J, Wang S, Wang Y and Wang J: NFATC1 promotes cell growth and
tumorigenesis in ovarian cancer up-regulating c-Myc through
ERK1/2/p38 MAPK signal pathway. Tumour Biol. 37:4493–4500.
2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Peng SL, Gerth AJ, Ranger AM and Glimcher
LH: NFATc1 and NFATc2 together control both T and B cell activation
and differentiation. Immunity. 14:13–20. 2001.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Kawahara T, Kashiwagi E, Ide H, Li Y,
Zheng Y, Ishiguro H and Miyamoto H: The role of NFATc1 in prostate
cancer progression: Cyclosporine A and tacrolimus inhibit cell
proliferation, migration, and invasion. Prostate. 75:573–584.
2015.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Im JY, Lee KW, Won KJ, Kim BK, Ban HS,
Yoon SH, Lee YJ, Kim YJ, Song KB and Won M: NFATc1 regulates the
transcription of DNA damage-induced apoptosis suppressor. Data
Brief. 5:975–980. 2015.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Heim L, Friedrich J, Engelhardt M, Trufa
DI, Geppert CI, Rieker RJ, Sirbu H and Finotto S: NFATc1 promotes
antitumoral effector functions and memory CD8+ T-cell
differentiation during non-small cell lung cancer development.
Cancer Res. 78:3619–3633. 2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Zhou L and Xie X: RNA-binding protein
CELF2 inhibits breast cancer cell invasion and angiogenesis by
downregulating NFATc1. Exp Ther Med. 22(898)2021.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Buchholz M, Schatz A, Wagner M, Michl P,
Linhart T, Adler G, Gress TM and Ellenrieder V: Overexpression of
c-myc in pancreatic cancer caused by ectopic activation of NFATc1
and the Ca2+/calcineurin signaling pathway. EMBO J. 25:3714–3724.
2006.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Oikawa T, Nakamura A, Onishi N, Yamada T,
Matsuo K and Saya H: Acquired expression of NFATc1 downregulates
E-cadherin and promotes cancer cell invasion. Cancer Res.
73:5100–5109. 2013.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Kawahara T, Kashiwagi E, Li Y, Zheng Y,
Miyamoto Y, Netto GJ, Ishiguro H and Miyamoto H: Cyclosporine A and
tacrolimus inhibit urothelial tumorigenesis. Mol Carcinog.
55:161–169. 2016.PubMed/NCBI View
Article : Google Scholar
|
|
26
|
Kawahara T, Kashiwagi E, Ide H, Li Y,
Zheng Y, Miyamoto Y, Netto GJ, Ishiguro H and Miyamoto H:
Cyclosporine A and tacrolimus inhibit bladder cancer growth through
down-regulation of NFATc1. Oncotarget. 6:1582–1593. 2015.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Xu S, Shu P, Zou S, Shen X, Qu Y, Zhang Y,
Sun K and Zhang J: NFATc1 is a tumor suppressor in hepatocellular
carcinoma and induces tumor cell apoptosis by activating the
FasL-mediated extrinsic signaling pathway. Cancer Med. 7:4701–4717.
2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Kawahara T, Inoue S, Fujita K, Mizushima
T, Ide H, Yamaguchi S, Fushimi H, Nonomura N and Miyamoto H: NFATc1
expression as a prognosticator in urothelial carcinoma of the upper
urinary tract. Transl Oncol. 10:318–323. 2017.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Singh SK, Chen NM, Hessmann E, Siveke J,
Lahmann M, Singh G, Voelker N, Vogt S, Esposito I, Schmidt A, et
al: Antithetical NFATc1-Sox2 and p53-miR200 signaling networks
govern pancreatic cancer cell plasticity. EMBO J. 34:517–530.
2015.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Saitoh M: Involvement of partial EMT in
cancer progression. J Biochem. 164:257–264. 2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Kim EK and Choi EJ: Pathological roles of
MAPK signaling pathways in human diseases. Biochim Biophys Acta.
1802:396–405. 2010.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Wang S, Kang X, Cao S, Cheng H, Wang D and
Geng J: Calcineurin/NFATc1 pathway contributes to cell
proliferation in hepatocellular carcinoma. Dig Dis Sci.
57:3184–3188. 2012.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Iñiguez MA, Martinez-Martinez S, Punzón C,
Redondo JM and Fresno M: An essential role of the nuclear factor of
activated T cells in the regulation of the expression of the
cyclooxygenase-2 gene in human T lymphocytes. J Biol Chem.
275:23627–23635. 2000.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Qiu J, Shi Z and Jiang J: Cyclooxygenase-2
in glioblastoma multiforme. Drug Discov Today. 22:148–156.
2017.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Wang L, Wang Z, Li J, Zhang W, Ren F and
Yue W: NFATc1 activation promotes the invasion of U251 human
glioblastoma multiforme cells through COX-2. Int J Mol Med.
35:1333–1340. 2015.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Liu Y, Liang T, Qiu X, Ye X, Li Z, Tian B
and Yan D: Down-regulation of Nfatc1 suppresses proliferation,
migration, invasion, and warburg effect in prostate cancer cells.
Med Sci Monit. 25:1572–1581. 2019.PubMed/NCBI View Article : Google Scholar
|
|
37
|
He W and Lu J: MiR-338 regulates NFATc1
expression and inhibits the proliferation and
epithelial-mesenchymal transition of human non-small-cell lung
cancer cells. Mol Genet Genomic Med. 8(e1091)2020.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Sun Y, Liu WZ, Liu T, Feng X, Yang N and
Zhou HF: Signaling pathway of MAPK/ERK in cell proliferation,
differentiation, migration, senescence and apoptosis. J Recept
Signal Transduct Res. 35:600–604. 2015.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Kim EK and Choi EJ: Compromised MAPK
signaling in human diseases: An update. Arch Toxicol. 89:867–882.
2015.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Lalkovicova M, Horvathova F, Sulla I,
Mihalik J and Danielisova V: Effects of low and high deprenyl dose
on antioxidant enzyme activities in the adult rat brain. Gen
Physiol Biophys. 36:83–90. 2017.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Lin H, Hao Y, Wan X, He J and Tong Y:
Baicalein inhibits cell development, metastasis and EMT and induces
apoptosis by regulating ERK signaling pathway in osteosarcoma. J
Recept Signal Transduct Res. 40:49–57. 2020.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Yang J, Zhang JN, Chen WL, Wang GS, Mao Q,
Li SQ, Xiong WH, Lin YY, Ge JW, Li XX, et al: Effects of AQP5 gene
silencing on proliferation, migration and apoptosis of human glioma
cells through regulating EGFR/ERK/p38 MAPK signaling pathway.
Oncotarget. 8:38444–38455. 2017.PubMed/NCBI View Article : Google Scholar
|