Introduction
Rheumatoid arthritis (RA) is a chronic, systemic
autoimmune disease (1-3),
which is characterized by pain and swelling, stiffness and
deformity of the joints, and this can lead to disability in severe
cases. In vivo, it manifests as persistent synovitis,
systemic inflammation and the generation of autoantibodies
(4), elevated levels of
pro-inflammatory cytokines and inflammatory mediators such as
interleukins IL-1β, IL-6, IL-8, TNF-α, chemokines and interferons
from synovial tissue (5-7),
leading to the development of disease and even accumulation of
peripheral organs (8). The
impaired joint function and dysfunction induced by RA limit a
patient's ability to move freely and seriously affects daily life,
resulting in increased psychological stress, reduced quality of
life and a heavy financial burden (9,10).
Research into the pathogenesis of RA is not yet
fully understood, but it is generally accepted that its development
is associated with genetics, environmental factors and immune
dysregulation (11). Associated
pathogenic mechanisms include an imbalance between Th1 and Th2
cells and Th17/Treg cells, leading to an inflammatory response in
the synovium and activation of synovial fibroblasts, leading to the
development of RA (12,13). Fibroblastic synovial cells (FLS), a
common type of cell found in synovial joints, serves an important
role in the development of RA and the over-proliferation of FLS has
been reported to be a significant factor in joint damage in RA
(14,15). MH7A cells are an RA synovial
fibroblast cell line and an established in vitro cell model
for the study of RA (16). An
early step in the development of RA is the activation of synovial
fibroblasts, which causes local autoimmune cells to infiltrate the
synovial tissues in response and promote the release of
pro-inflammatory cytokines leading to ongoing synovial
inflammation. Toll-like receptor (TLR), an important pattern
recognition receptor mediating natural immunity, mediates signaling
pathways that play an important role in the development of
inflammation (17). During the
development of inflammation, NLR family pyrin domain-containing 3
(NLRP3), NF-κB and other inflammatory pathways are activated as a
result of the excitement of TLR4 on the cell membrane (18). TLR4 primarily plays a role in
triggering an immune response and phagocytosis of bacteria when
substances that cause activation of MH7A cells bind to TLR4 and
activate NF-κB phosphorylation and thus translocation to the
nucleus, thereby initiating NLRP3 inflammatory vesicles (19). Inflammasomes were first proposed by
Tschopp in 2002(20), and among
the several subtypes of inflammasomes, the NLPR3 inflammasome is
one of the most extensively studied; a multi-class protein complex
thought to be widely involved in the body's inflammatory and immune
responses, it is widely present in immune cells, including
granulocytes, antigen-presenting cells (APCs), macrophages, T and B
lymphocytes (21) and in
inflammatory vesicles, by activating Caspase-1, IL-β and IL-8 and
initiating an inflammatory response (22,23).
Strychnine, the dried mature seeds of Strychnos
nurvomica L. (family, Strychnosaceae), has a long history of
use in Chinese medicine and has been widely used in China for
swelling and pain in joints (24,25).
In modern times, strychnine is also widely used in Chinese medicine
to treat diseases such as cancer and orthopedic and inflammatory
conditions (26-28).
The primary pharmacological component of strychnine are alkaloids,
which account for 1.5-5% of the total chemical composition
(29), with strychnine being the
most abundant and potent component (30). Alkaloid constituents in strychnine
have strong and long-lasting analgesic effects. Brucine is not only
a medicinal component of strychnine but is also a toxic
constituent. It has toxic effects on the nervous, immune, urinary
and digestive systems (31-34).
These toxic effects are the main reason that the widespread use of
strychnine in clinical practice is limited (35-37).
It has been demonstrated that strychnine
significantly inhibits TNF-induced proliferation of HFLS-RA through
activation of the JNK signaling pathway (25). The mechanism of action of
strychnine and Tripterygium wilfordii in the treatment of RA
is through the blocking of the angiogenic mediator cascade by
targeting multiple interactions (38). Research has confirmed that
Atractylodes macrocephala extract has an antagonistic effect on the
intestinal absorption of strychnine (39). A study has confirmed the
antagonistic effect of Atractylodes macrocephala extract on
the intestinal absorption of stilbene (39). The above studies show strychnine's
potential in the clinical treatment of RA.
Methotrexate is an anti-folate oncology drug but has
been revealed to treat RA as early as 1951(40). It also has a specific role; as a
first-line anti-rheumatic drug, it modulates the function of the
inflammatory cells involved in rheumatoid arthritis (41). Because it is highly efficacious and
well-tolerated, is beneficial in the vast majority of patients and,
if used early in life, will achieve the same results as other
biological agents (42).
Therefore, the present study selected methotrexate as a control
drug.
In the current clinical treatment of RA,
pharmaceuticals remain the primary option of treatment, primarily
non-steroidal anti-inflammatory drugs, anti-rheumatic drugs,
glucocorticoids and biological agents, amongst others; however,
these drugs generally have other issues such as a high risk of
relapse, complicated administration instructions resulting in poor
adherence, development of drug resistance and expense. Botanicals
and their extracts have been of great interest in the treatment of
RA due to their high efficacy and lower risk of side effects. In
the present study, HFLS cells were used as a blank control group
and MH7A cells, which have a greater migratory and invasive
capacity compared with HFLS cells (43), were used as a model group to
investigate whether strychnine with Atractylodes
macrocephala extract could affect the proliferation of MH7A
cells and whether it was associated with the TLR4/NF-κB/NLRP3
pathway.
Materials and methods
Reagents
Strychnine and Atractylodes macrocephalae
were purchased from Shandong Provincial Hospital of Traditional
Chinese Medicine (Jinan, China). Strychnine (cat. no. DS0026;
Chengdu Desite Biotechnology Co., Ltd.), Brucine (cat. no. DM0019;
Chengdu Desite Biotechnology Co., Ltd.) Atractylenolide II (cat.
no. DB0015; Chengdu Desite Biotechnology Co., Ltd.) and
Methotrexate (cat. no. H31020644; Tonghua Maoxiang
Pharmaceutical).
Cell culture
MH7A cells (cat. no. BNCC358158; BeNa Culture
Collection; Beijing Beina Chunglian Institute of Biotechnology)
were cultured in H-DMEM (cat. no. 12100046; Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) and 1% penicillin mix (cat. no. P1400-100;
Beijing Solarbio Science & Technology Co., Ltd.) at 37˚C in a
humidified incubator supplied with 5% CO2. HFLS cells
(cat. no. JNO171-71; Jennio Biotech Co., Ltd.) were cultured in
DMEM (cat. no. SH30243.01; Hyclone; Cytiva) supplemented with 10%
FBS and 1% penicillin mix at 37˚C in a humidified incubator
supplied with 5% CO2. Both types of cells were purchased
as the second-generation cell lines and passed through once in
about 48 h, with subsequent experiments starting when both types of
cells reached the 7th generation.
High-performance liquid chromatography
(HPLC)-tandem mass spectrometry (MS/MS) analysis of chemical
composition
The same batch of Strychnine and Atractylodes
macrocephala was weighed, and 20 g Strychnine and 1,000 g
Atractylodes macrocephala were weighed in 10 times the amount of
water (w/v) (in the current study, 20 g of Strychnine and 200 g of
water, 1,000 g of Atractylodes macrocephala and 10,000 g of water
were used). The same method was adopted for a common decoction of
20 g Strychnine and 120 g Atractylodes macrocephala, and 20
g Strychnine and 240 g Atractylodes macrocephala. The drug
was added to a round-bottom flask, about half of its capacity, and
water was added to soak the surface of the restorative material by
1-2 cm, heated at 100˚C, refluxed and decocted and kept boiling for
1 h. After cooling to room temperature, a 0.22 microporous membrane
was used for filtration. After decoction, 3 ml of the decoction was
suspended in a 10-ml volumetric flask (3:7 ratio of decoction to
HPLC grade methanol), diluted to the required scale using
HPLC-grade methanol (cat. no. 34860; Sigma-Aldrich; Merck KGaA) and
transferred to a centrifuge tube at 25˚C, 1,610 x g for 15 min.
When the centrifugation was completed, the supernatant was
collected, the missing solution (the discarded precipitated
fraction of liquid) was made up with methanol, the solution was
filtered through a 0.22-µm microporous membrane and 2 µl of liquid
was used for mass spectrometry analysis.
The strychnine, brucine, and Atractylenolide II were
accurately weighed (all 1 mg) and dissolved in 1 ml HPLC grade
methanol to obtain the ‘master mix’ of each substance.
Subsequently, different proportions of the master mix were
accurately aspirated and placed in the same 5-ml volumetric flask
and mixed thoroughly to obtain the mixed reference substance
solution for subsequent standard curve plotting. The concentrations
of the resulting controls are presented in Table I.
 | Table IControl component concentrations. |
Table I
Control component concentrations.
| | Control mixture
number |
|---|
| Component | 1 | 2 | 3 | 4 | 5 | 6 |
|---|
| Strychnine,
µg/ml | 0.292 | 0.584 | 1.167 | 2.334 | 4.668 | 9.336 |
| Brucine, µg/ml | 0.272 | 0.545 | 1.089 | 2.178 | 4.356 | 8.712 |
| Atractylodes II,
µg/ml | 0.260 | 0.519 | 1.038 | 2.076 | 4.152 | 8.304 |
The study was carried out on a TSQ Quantis
triple-stage quadrupole mass (TSQ QUANTIS; Thermo Fisher
Scientific, Inc.) with a Waters Symmetry-C18 (4.6x75 mm; 3.5 µm;
Waters Corporation) column. The mobile phase was 1% formic acid
(A)-acetonitrile (B) with gradient elution (0-35 min, 95-65% B;
35-40 min, 65% B; 40-55 min, 65-5% B; 55-56 min, 5-95% B; 56-60
min, 95% B) at a flow rate of 0.2 ml/min, column temperature of
30˚C and injection volume of 2.0 µl. The ion source was carried out
using positive ion electrospray ionization (ESI) source with the
sheath gas (N2) flow rate set to 1.2 l/min; the
auxiliary gas (N2) was set to a flow rate of 0.09 l/min;
the collision gas was argon (Ar) assigned to a flow rate of 2 m
Torr; the atomization temperature was 300˚C; the atomizer pressure
of 35 psi, the capillary temperature was 280˚C; and the spray
voltage is 3,500 V. The scanning method was Selective Response
Monitoring) for each ion. Strychnine m/z 395.16-243.97 with a
collision energy of 37.73 eV, Brucine m/z 335.11-184 with a
collision energy of 38.66 eV and Atractylenolide II m/z 232.3-158.1
with a collision energy of 17.93 eV. Data were acquired using
Xcalibur software 4.1 (Thermo Fisher Scientific, Inc.), peak areas
were calculated and standard curves were plotted using the
concentration and peak area of the markers. AM group=the mixed
standard. SS group=the liquid obtained by decoction of strychnine
(20 g); S:A (1:6) group=liquid from decoction of strychnine (20 g)
and Atractylodes macrocephala (120 g); S:A (1:12) liquid
from decoction of strychnine (20 g) and Atractylodes
macrocephala (240 g).
CCK-8 assay
Strychnine and Atractylenolide II were accurately
weighed, and the two drugs were dissolved in the culture medium
(H-DMEM medium containing 10% FBS and 1% penicillin-streptomycin)
to the desired concentration. The Strychnine concentrations were
130, 390 and 780 µg/ml, while the concentrations of Atractylenolide
II were 2, 20 and 40 µg/ml, followed by filtration through a
0.22-µm microporous filter membrane. Cells in the logarithmic phase
of growth were digested for 1 min in a 5% CO2 incubator
at 37˚C with trypsin-EDTA digest (0.25%; cat. no. T1300-100;
Beijing Solarbio Science & Technology Co., Ltd.) and made into
cell suspensions. Subsequently, 100 µl of each of the
aforementioned solutions were added to a 96-well plate and 100 µl
of medium was used as a blank control. The plate was incubated
overnight before dividing into seven groups: i) Control group; ii)
strychnine 130 µg/ml group; iii) strychnine 390 µg/ml group; iv)
strychnine 780 µg/ml group; v) Atractylenolide II 2 µg/ml; vi)
Atractylenolide II 20 µg/ml; and vii) Atractylenolide II 40 µg/ml.
After incubation for 0, 12, 24 or 48 h, a Cell Counting Kit-8
(CCK-8; cat. no. CP002; Signalway Antibody LLC) and serum-free
H-DMEM were mixed in a 1:10 volume ratio, and 100 µl was added per
well, after which cells were incubated for 1 h. Subsequently, the
absorbance at 450 nm was measured using an enzyme marker (cat. no.
DNM-9602; Beijing Prolong New Technology Co., Ltd.), and the values
for each plate were recorded. A total of three replicates were used
for each experimental group.
Flow cytometry analysis
Cells were divided into 7 groups: i) Control group
(HLFS); ii) model group (MH7A); iii) methotrexate group (MTX, 0.1
mg/ml); iv) strychnine group (780 µg/ml); v) Atractylodes
macrocephala group (12 µg/ml); vi) strychnine with
Atractylodes macrocephala 1:6 group (S:A 1:6); and vii)
strychnine with Atractylenolide II (S:A 1:6 component 780-12 µg/ml)
group. After adding drugs, the above groups were put in a
humidified incubator at 37˚C supplied with 5% CO2 for 24
h. Among them, the strychnine and Atractylodes macrocephala
group (S:A 1:6) consisted of two drugs, strychnine and
Atractylodes macrocephala, two drugs co-decocted in a ratio
of 1:6. The strychnine and atractylenolide II (S:A 1:6 component
780-12 µg/ml) group consisted of only two extracts, strychnine and
Atractylenolide II, configured together. The media was removed from
the drug-treated groups of cell cultures, the cells were washed
with PBS, and the cells were digested using trypsin-EDTA digest.
The cells were observed to be rounding under the microscope and the
wall of the flask was gently tapped to observe when the cells fell
off in a quicksand pattern. Medium was added to the flask to
terminate the digestion. The cell cultures from the previous step
were collected, mixed slightly and transferred to a centrifuge
tube, centrifuged at 1,000 x g, 25˚C for 5 min and then resuspend
in PBS and drop resuspended onto the cell counting chamber for
manual counting under a microscope. Between 50-100,000 resuspended
cells were collected, centrifuged at 1,000 x g, 25˚C for 5 min, the
supernatant was discarded and 200 µl Annexin V-FITC conjugate (cat.
no. C1062S; Beyotime Institute of Biotechnology) was added and
gently resuspended, mixed gently and incubated for 15 min at 4˚C in
the dark. Subsequently, 5 µl propidium iodide staining solution was
added, gently mixed and incubated for 5 min at 4˚C, while a tube
without Annexin V-FITC and PI was used as the negative control.
Flow cytometry analysis was performed using a BD Accuri C/6 flow
cytometer (Becton, Dickinson and Company). Annexin V-FITC
fluoresces green, corresponding to the FL1 detection channel; PI
fluoresces red, corresponding to the FL2 detection channel. The
analysis was performed using FlowJo 7.6.1 software (FlowJo LLC). A
total of five replicates were used for each experimental group.
Immunofluorescence analysis
The dilution, batch number and supplier of the
primary antibodies used were: TLR4 (1:800; cat. no. ab22048;
Abcam), NF-κBP65, (1:1,200; cat. no. ab16502; Abcam) and NLRP3
(1:800; cat. no. ab214185; Abcam). The secondary antibodies used
were IgG (Fluorescent dye FITC coupling; 1:100; cat. no. ZF-0316;
OriGene Technologies, Inc.). The cells were digested with 0.25%
trypsin-EDTA in a 5% CO2 37˚C incubator for 1 min; the
digested cell suspension was dropped onto a slide placed in the
culture dish. After waiting for ~30 min for the cells to adhere to
the wall, the medium (H-DMEM medium containing 10% FBS and 1%
penicillin-streptomycin) was added and incubated in a 37˚C 5%
CO2 incubator for 2 h. The slide was removed after 2 h
and rinsed three times with PBS for 2 min each time. Cells were
fixed using 4% paraformaldehyde for 30 min at room temperature and
washed, permeabilized using 0.5% Triton X-100 (in DPBS) for 20 min,
followed by treatment with 3% methanolic hydrogen peroxide for 20
min at room temperature and then placed in PBS. A Pap Pen was used
to draw circles surrounding the cells being examined on the slide
to avoid the running and spreading of the fluid during staining.
Goat serum sealant (no. ZLI-9022; ZSGB) was added dropwise for 20
min at room temperature to block the cells. The primary antibody
was added dropwise and cells were incubated overnight at 4˚C,
removed the next day and left for 20 min before adding the
fluorescently labeled secondary antibody (IgG) dropwise. Cells were
incubated in the dark for 20 min at 37˚C, placed in PBS, washed
with water, air-dried and sealed with neutral resin. Overall, three
replicates were used for each experimental group. Cells were
observed using a fluorescence microscope at a magnification of 100x
and 400x. Analysis of gray value with Image J 1.8.0 software
(National Institutes of Health). An ‘immunohistochemical score
(IHS)’ was calculated for each group by multiplying the percentage
of stained cells with the score for staining intensity, using the
following criteria: A=Grading the number of positive cells (0-1%=0;
1-10%=1; 10-50%=2; 50-80%=3; 80-100%=4); B=grading of the intensity
of color development of positive cells [0 (negative), 1 (weakly
positive), 2 (positive), 3 (strongly positive)]. Therefore, IHS=A x
B.
Reverse transcription-quantitative PCR
(RT-qPCR) assay
Total RNA was extracted from the tissue samples
using an ultra-pure RNA extraction reagent (cat. no. 9108; Takara
Bio, Inc.). Using the property that nucleic acids have strong
absorption ~260 nm and proteins have strong absorption ~280 nm, the
A260/A280 ratio of RNA was determined using a Nanodrop 2000
ultra-micro spectrophotometer; the ideal RNA purity A260/A280
should be in the range of 1.9-2.1. The residual genomic DNA in the
RNA was digested using a gDNA Eraser (cat. no. RR047A; Takara Bio,
Inc.). RT was performed using a Reverse Transcription kit according
to the manufacturer's protocol (cat. no. DRRO47A; Takara Bio,
Inc.). qPCR was performed using a SYBR Premix Ex Taq kit (cat. no.
DRR420A; Takara Bio, Inc.) and amplification procedure: 95˚C for 30
sec, (95˚C for 5 sec; 60˚C for 34 sec) x40 cycles. The data were
analyzed using the 2-ΔΔCq method (44). Amplification was performed on an
ABI 7500 fluorescent qPCR instrument. The sequences of the primers
were: TLR4 forward, 5'-CAGGATGATGTCTGCCTCGC-3', and reverse,
5'-TGGTTTAGGGCCAAGTCTCC-3'; IκB kinase (IKKβ) forward,
5'-CTAAGGTGGAAGTGGCCCTC-3', and reverse,
5'-CTGGATCCTACAAGGGACCG-3'; NF-κB p65 forward,
5'-TGAACCAGGGCATACCTGTG-3', and reverse 5'-CCCCTGTCACTAGGCGAGTT-3';
NLRP3 forward, 5'-GCTGGCATCTGGGGAAACCT-3', and reverse,
5'-GGTCCTTAGGCTTCGGTCCA-3'; TNF-α forward,
5'-GCTGCACTTTGGAGTGATCG-3' and reverse, 5'-TCACTCGGGGTTCGAGAAGA-3';
and GAPDH forward, 5'-GCAAATTCCATGGCACCGTC-3' and reverse,
5'-AGCATCGCCCCACTTGATTT-3'. A total of five replicates were used
for each experimental group.
Western blotting
The dilutions, batch numbers and suppliers of the
antibodies used were: TLR4 (1:800; cat. no. ab22048; Abcam),
p-NLRP3 (1:600; cat. no. AF4320; Affinity Biosciences), NLRP3
(1:800; cat. no. ab214185; Abcam), phosphorylated (p-)NF-κB P65
(1:600; cat. no. ab86299; Abcam), TNF-α (1:1,000; cat. no. ab6671;
Abcam), IKKβ (1:800; cat. no. ab124957; Abcam), NF-κB P65 (1:1,200;
cat. no. ab16502; Abcam) and β-actin (1:1,000; cat. no. TA-09;
OriGene Technologies, Inc.). The cells were collected, centrifuged
at 1,000 x g for 5 min at 25˚C to remove the original culture
fluid, washed with PBS and lysed using 200 µl lysis solution (0.303
g Tris base, 0.4383 g NaCl, 0.05 g SDS, 40 ml H2O; pH
adjusted to 8.0 with HCl to a fixed volume of 50 ml) supplemented
with PMSF. After centrifuging at 1,200 x g, 15˚C for 10 min to
remove the cell debris, the protein concentration of each sample
was determined using the Coomassie Blue Staining method (45), gently mixed with 5x protein gel
loading buffer, denatured at 95˚C for 10 min and stored at -80˚C
until required. The extracted total protein samples were removed
from the -80˚C freezer and immediately inserted into ice and left
to melt; according to the results of protein quantification, the
corresponding volume of total protein was added to each lane (~30
µg protein/lane). Total protein was separated by SDS-PAGE on a 10%
gel and transferred to a PVDF membrane. The PVDF membrane was
carefully removed and placed in a sealing solution consisting of
100 ml 1X TBST (cat. no. T1085; Beijing Solarbio Science &
Technology Co., Ltd.) and 5 g of skimmed milk powder (20:1) for 1 h
at room temperature using a shaker with slow shaking, and then
incubated with the primary antibodies followed by the secondary
antibody is horseradish peroxidase labelled goat anti-rabbit IgG
(1:3,000; cat. no. ZB-2301; OriGene Technologies, Inc.), slow
shaking at room temperature and away from light for 60 min.
Analysis of gray values was performed using ImageJ 1.8.0 software
(National Institutes of Health). The visualization reagents used
are BeyoECL Plus luminescent liquids (cat. no. P0018S; Beyotime
Institute of Biotechnology). A total of five replicates were used
for each experimental group.
Statistical analysis
Data are expressed as mean ± standard deviation. All
experiments were repeated at least three times. Differences between
multiple groups were assessed using one-way analysis of variance
and the Bonferroni post hoc test. When the data contains
non-parametric data, the data are tested for normality. When the
data conform to the normal distribution, the Kruskal-Wallis test
was used for analysis. If the Kruskal-Wallis was significant, the
Bonferroni method was used for analysis. All statistical analyses
were performed in SPSS version 26.0 software (IBM Corp). P<0.05
was considered to indicate a statistically significant
difference.
Results
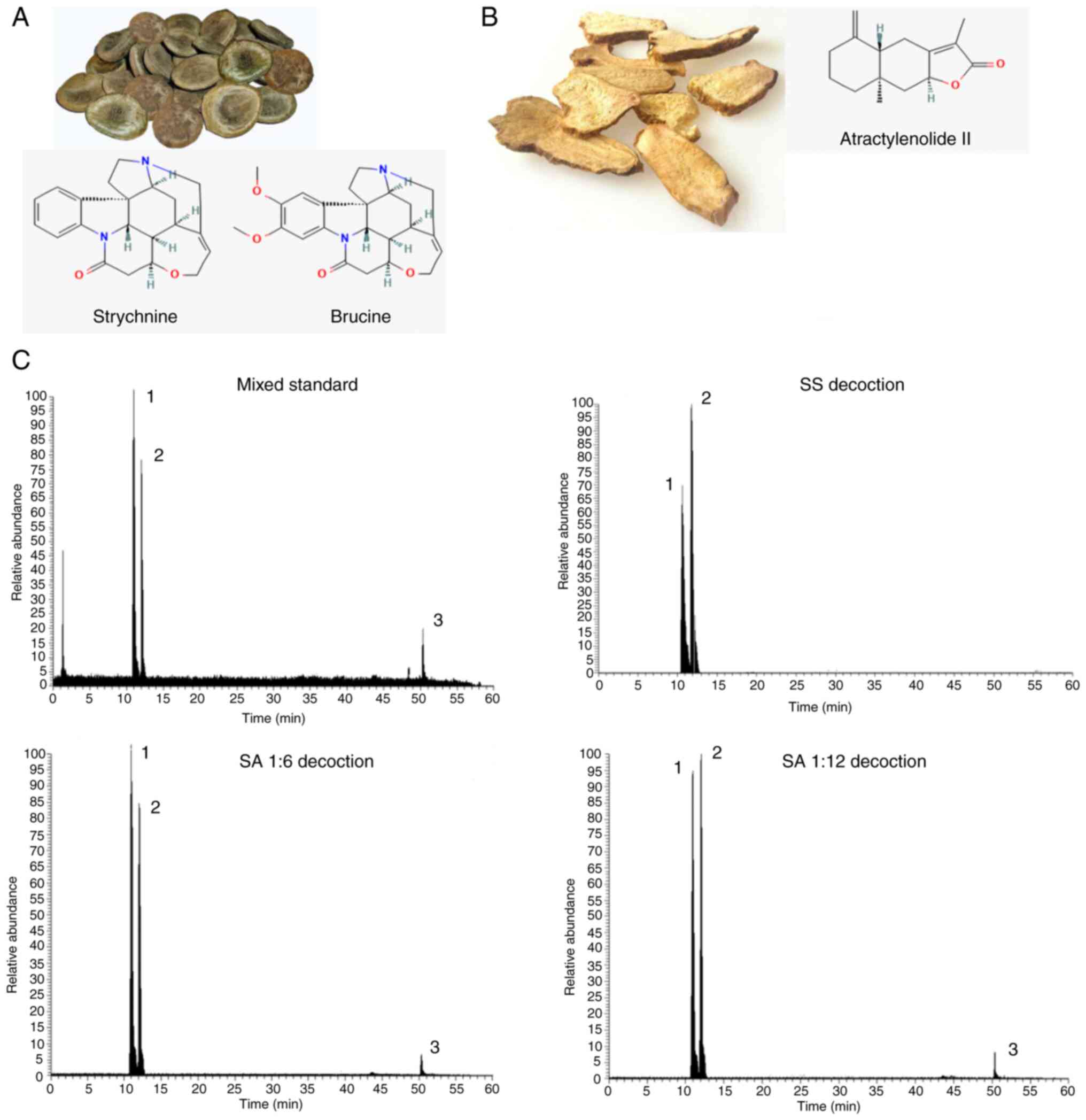
The use of strychnine in combination
with Atractylodes macrocephala can reduce the strychnine
content
To determine the changes in the contents of
strychnine, brucine and Atractylenolide II before and after the
combination of Strychnos and Atractylodes macrocephala, the
horizontal coordinate was set as the concentration of the control,
the vertical coordinate was set as the peak area and the standard
curves of the three chemical components were calculated and the
linear regression was calculated. The regression equations and
results of three chemical components were calculated respectively:
y=2E+06x+88958, R2=0.9986; y=2E+06x-273931, R2=0.9996;
y=115567x-4142.5, R2=0.9993. The value of R2 was
calculated to determine whether the standard curve has a good
linear relationship. That is, the relationship between the
concentration and the readout indicated a linear relationship, and
when the R2>0.997, the standard curve has a good
linear relationship. From the aforementioned results, it was
determined that the contents of strychnine, brucine and
Atractylenolide II could be ascertained from the linear curve
relationship established. A quantity of the control was injected
six times in succession according to the aforementioned method and
the concentrations of the three controls were subsequently
measured. By calculating the relative standard deviation, the
results exhibited <4% deviance, demonstrating that the method
had good precision, reproducibility and accuracy. Precise
quantities of each group of samples were injected three times and
the external standard method was used to calculate the strychnine,
brucine and Atractylenolide II levels. The results are presented in
Table II. Compared with the SS
group, the SA (1:6) and SA (1:12) groups contained lower levels of
strychnine and brucine. The chromatograms of each group were
obtained using HPLC-MS/MS (Fig.
1). Overall, the combination of strychnine and Atractylodes
macrocephala could reduce the content of toxic components in
strychnine and improve the safety of the medication.
| Table IIQuantitative results of the effective
components of brucine, strychnine and Atractylenolide. |
Table II
Quantitative results of the effective
components of brucine, strychnine and Atractylenolide.
| Sample | Brucine, µg/ml | Strychnine,
µg/ml | Atractylenolide II
µg/ml |
|---|
| SS | 129.212 | 127.234 | Not detected |
| AM | 0.222 | 0.0785 | 0.341 |
| SA (1:6) | 108.261 | 101.411 | 2.086 |
| SA (1:12) | 105.935 | 100.583 | 3.353 |
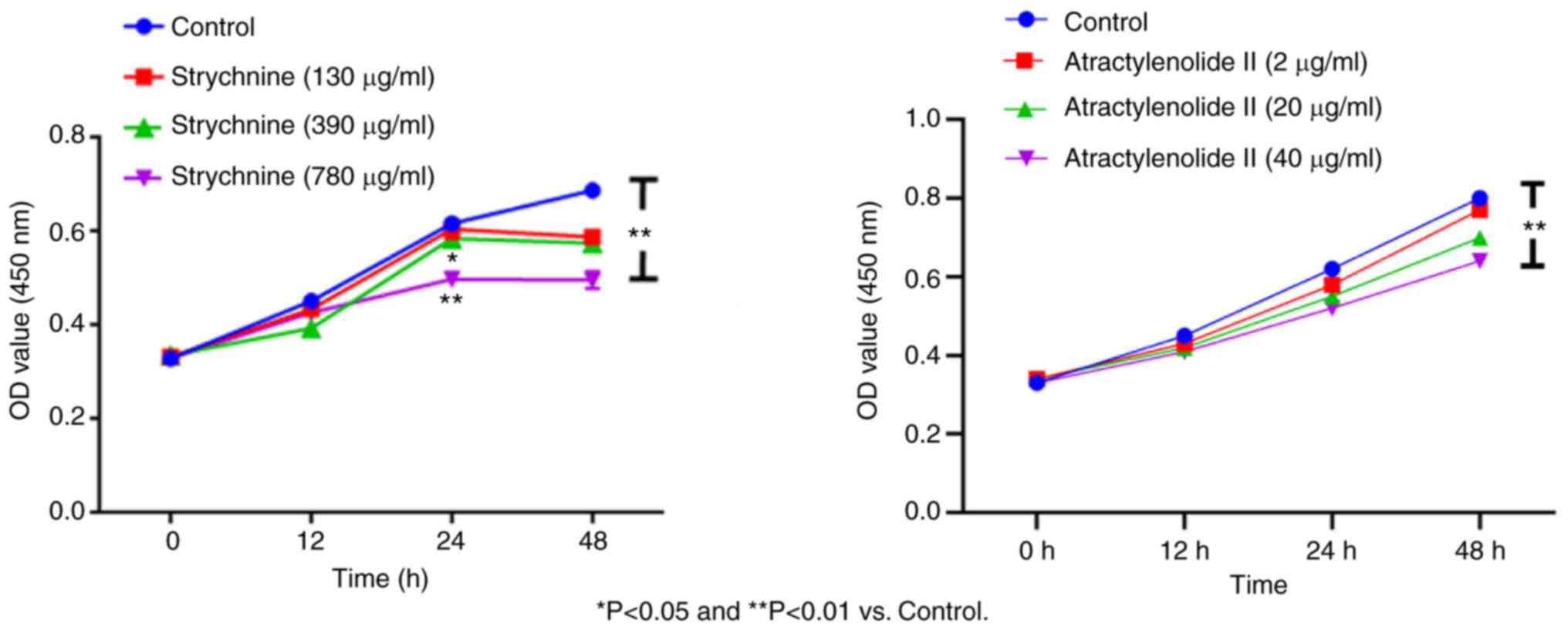
MH7A cell proliferation is inhibited
by strychnine
When MH7A cells were treated with 780 µg/ml
strychnine for 24 h, there was a significant decrease in the
proliferation of cells compared with the control group (P<0.01),
and the effect was time-dependent with treatment for 48 h showing a
further decrease. However, given the bigger decrease in the number
of cells after 48, 24 h of treatment was used for all subsequent
experiments. When MH7A cells were treated with 2, 20 and 40 µg/ml
of Atractylenolide II for 12, 24 and 48 h, the cell proliferation
demonstrated an upward trend compared with the control, which was
significant for the 40 µg/ml of Atractylenolide II group at 48 h
(P<0.01). This effect was also time-dependent, as no significant
inhibitory trend was observed within 48 h of treatment, indicating
that Atractylenolide II did not affect cell proliferation. Combined
with the inhibitory effect of strychnine on cell proliferation, 24
h treatment was used for all subsequent experiments (Fig. 2). These results indicated that high
doses of strychnine intervention MH7A cells at 48 h significantly
impacted cell proliferation and, therefore, strychnine has some
toxicity to cells.
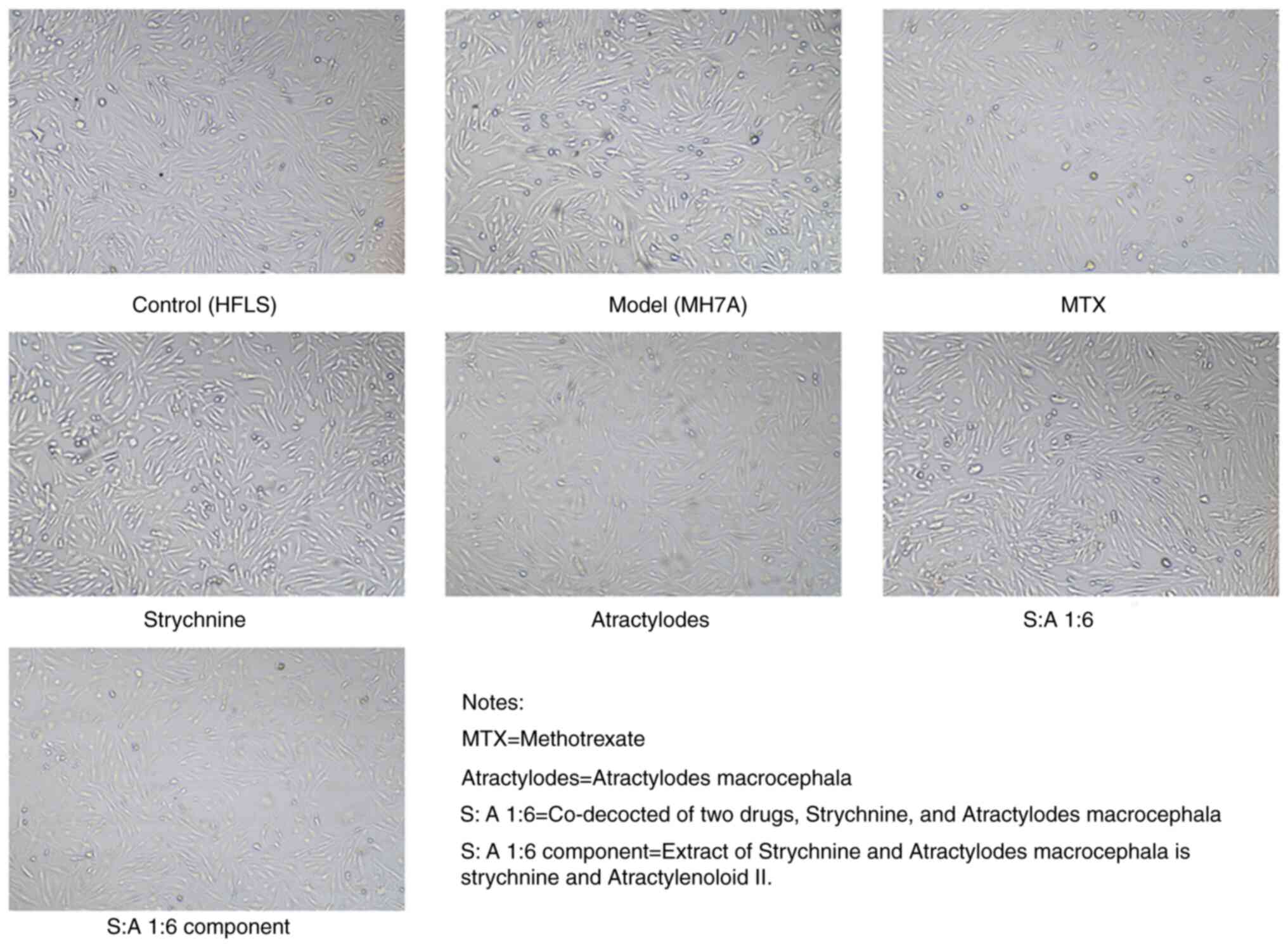
Strychnine promotes apoptosis of
synovial cells when combined with Atractylodes macrocephala
The cells started to adhere to the wall after 3-6 h
of incubation; the morphology of the cells appeared fibrous before
adherence to the wall. After 24 h of incubation, the distribution
of cells indicated a certain direction and regularity. Both HFLS
cells and MH7A cells possessed fibroblast-like characteristics,
being spindle-shaped. There were no markedly different
morphological changes in the cells under bright light microscopy
after the addition of the drug treatment (Fig. 3).
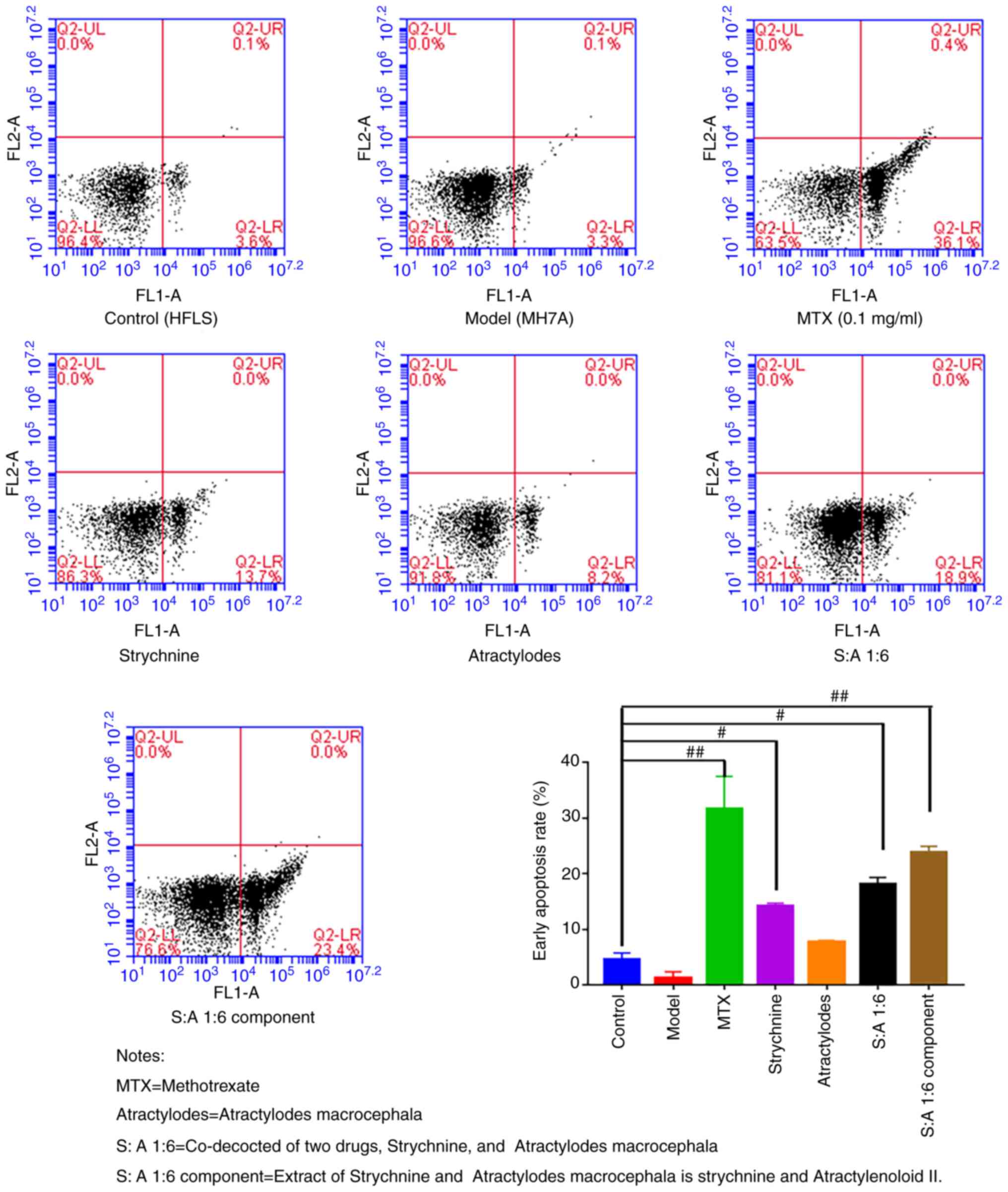
The results of the flow cytometry assay revealed
that the rate of early apoptosis was low in the Control and Model
groups and did not differ significantly from each other. After
treatment with MTX, strychnine, S-A1:6 and S-A1:6 component, the
apoptosis rate of MH7A cells was significantly higher compared with
that of the control group (P<0.05 or P<0.01; Fig. 4). These results indicated that the
drugs in each group had no significant effect on the cell
morphology but could significantly promote the apoptosis of MH7A
cells after the intervention of two drugs.
The strychnine and Atractylodes
macrocephala combination inhibits the expression of TLR4, NF-κB and
NLRP3
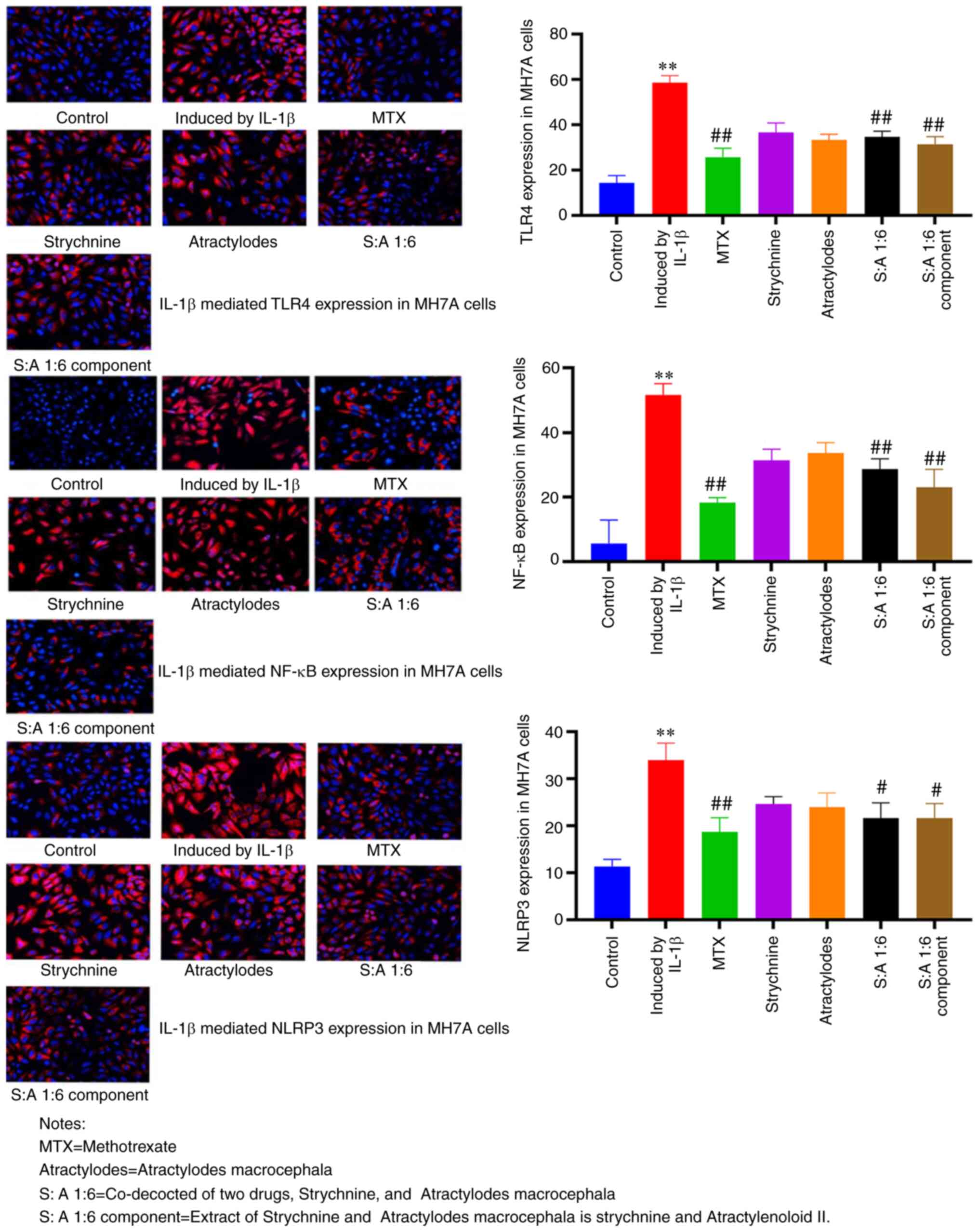
Compared with the control group, the expression
levels of TLR4, NF-κB and NLRP3 were significantly enhanced in MH7A
cells treated with IL-1β (P<0.01; Fig. 5). TLR4 expression was predominantly
observed at the cell membrane, NLRP3 expression predominantly in
the cytoplasm and NF-κB present at both. The number of positive
cells was significantly reduced after MTX, S-A 1:6 and S-A 1:6
component treatment compared with the IL-1β-induced group
(P<0.05 or P<0.01; Fig. 5).
These results showed that the expression levels of TLR4, NF-κB and
NLRP3 were significantly increased in MH7A cells induced by IL-1β,
and the expression was significantly decreased after drug
intervention. This demonstrated that the combination of the two
drugs had the potential to inhibit the expression of TLR4, NF-κB
and NLRP3.
The mRNA expression levels of TLR4,
NF-κB and NLRP3 are significantly reduced after drug
intervention
Compared with the control group, the mRNA expression
of TLR4, IKKβ, NF-κB, NLRP3 and TNF-α were significantly increased
in IL-1β-induced MH7A cells (P<0.01; Fig. 6). Following MTX, S-A 1:6 and S-A
1:6 component treatment, expression levels of TLR4, IKKβ, NF-κB and
NLRP3 were significantly reduced (P<0.01; Fig. 6). The results showed that the
expression levels of TLR4, IKKβ, NF-κB, NLRP3 and TNF-α were
significantly enhanced in IL-1β-mediated MH7A cells, and the
expression levels were significantly decreased after the
combination of the two drugs. This indicated that the combination
of the two drugs has the potential to reduce the expression of
these factors.
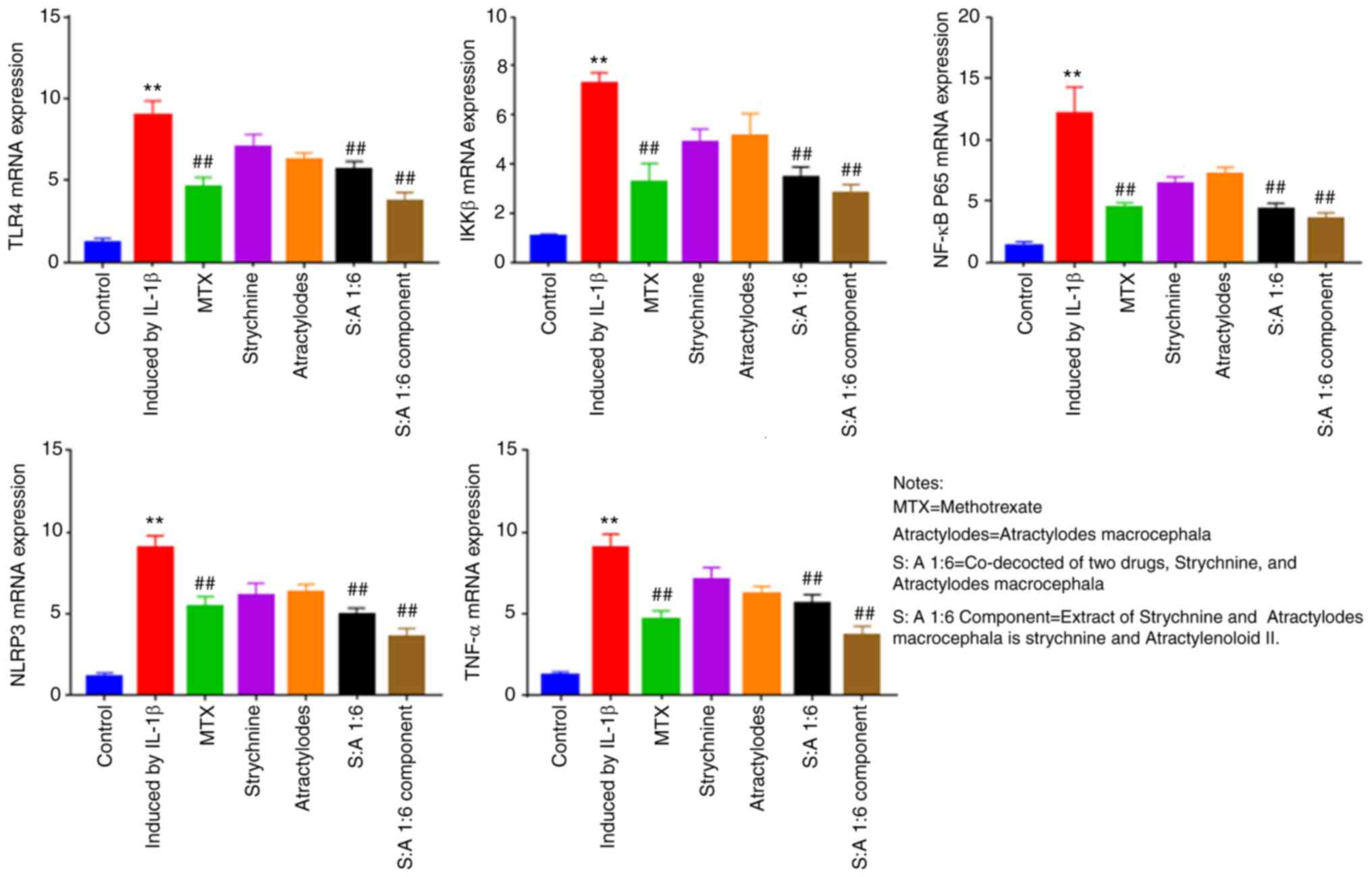 | Figure 6Effect of strychnine with
Atractylodes macrocephala on IL-1β-mediated mRNA expression
of TLR4, IKKβ, NF-κBp65, NLRP3 and TNF-α in MH7A cells. n=5.
**P<0.01 vs. Control; ##P<0.01 vs.
Induced by IL-1β. TLR4, Toll-like receptor 4; IKK, IκB kinase;
NF-κBp65; NLRP3, NLR family pyrin domain-containing 3; TNF-α
expression; MTX, Strychnine, Atractylodes macrocephala; S:A
1:6, co-decoction of two drugs, Strychnine and Atractylodes
macrocephala; S:A 1:6 component group, extract of strychnine
and Atractylenoloid II. |
Phosphorylation of TLR4 and NF-κB is
significantly reduced following treatment with strychnine combined
with white Atractylodes macrocephala
Compared with the control group, IL-1β-induced total
protein expression levels of TLR4, IKKβ and NF-κB were
significantly increased in MH7A cells (P<0.01; Fig. 7). The results showed that the
ratios of pNF-κBP65/NF-κBP65 and pNLRP3/NLRP in MH7 A cells
mediated by IL-1β were lower than those in the Control group. After
drug intervention, the ratios of pNF-κBP65/NF-κBP65 and pNLRP3/NLRP
increased. In addition, the phosphorylation levels of NF-κB and
NLRP3 were elevated in the IL-1β-induced group compared with the
Control group (P<0.01; Fig. 7).
By contrast, expression levels of TLR4, p-NF-κB, NF-κB, p-NLRP3,
TNF-α and IKKβ were significantly reduced after MTX, S-A 1:6 and
S-A 1:6 component treatment (P<0.05 or P<0.01; Fig. 7). In addition, expression of NLRP3
was significantly reduced after MTX and N-A1:6 treatment (P<0.5;
Fig. 7). The above results showed
that the combination of the two drugs could significantly inhibit
the expression of TLR4, IKKβ and NF-κB in IL-1β cells.
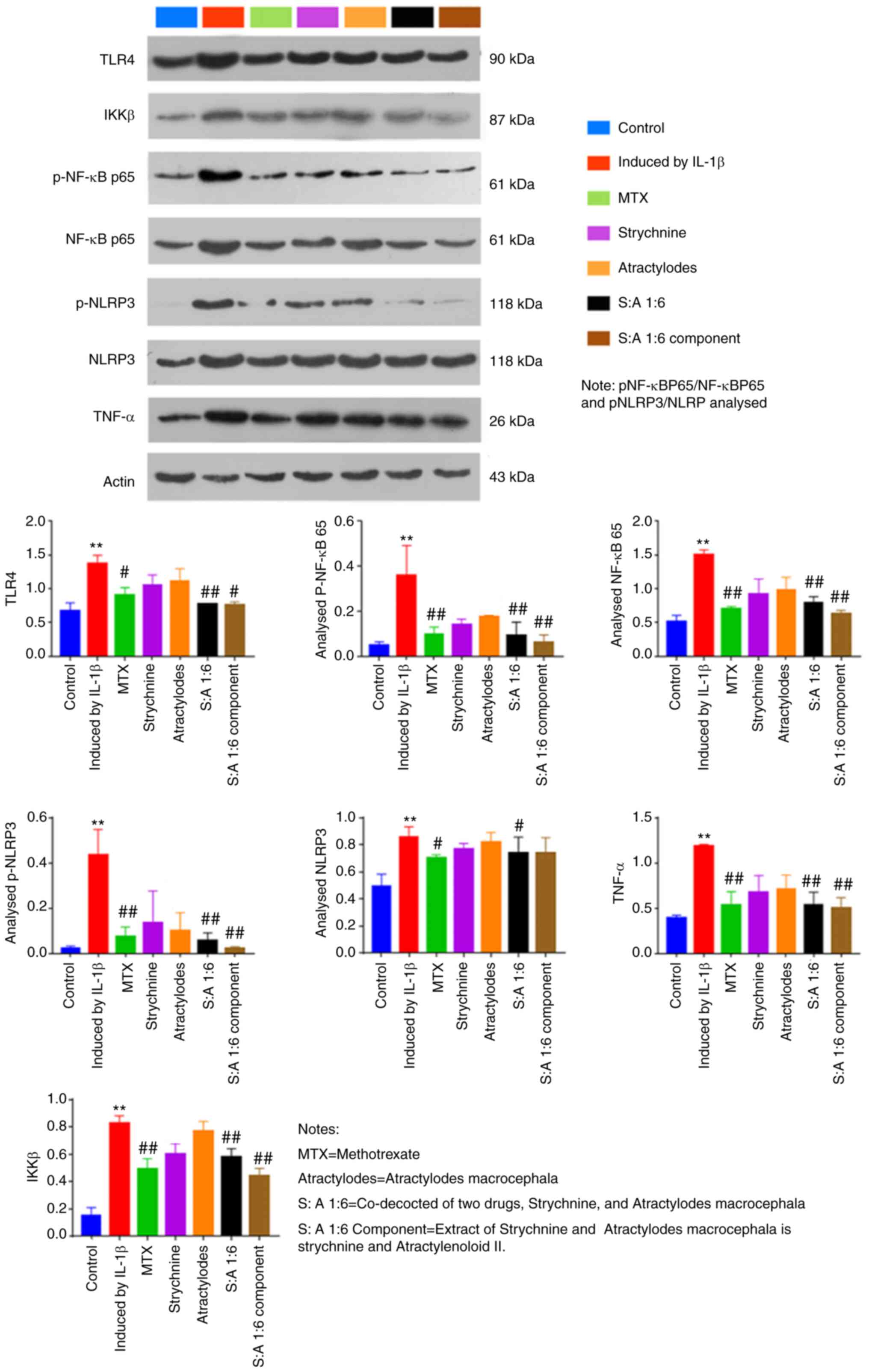 | Figure 7Effect of the strychnine and
Atractylodes macrocephala combination on IL-1β-mediated
expression of TLR4, NF-κB, IKKβ, NF-κB p65, p-NF-κB p65, NLRP3,
p-NLRP3 and TNF-α in MH7A cells. n=5. **P<0.01 vs.
Control; #P<0.05 and ##P<0.01 vs.
Induced by IL-1β. TLR4, Toll-like receptor 4; IKK, IκB kinase;
p-NF-κB p65; NF-κB p65. NLRP3, NLR family pyrin domain-containing
3; p-, phosphorylated; TNF-α expression;, MTX, methotrexate;
Strychnine; Atractylodes macrocephala; S: A 1:6 co-decocted
of two drugs co-decocted of two drugs, Strychnine, and
Atractylodes macrocephala, S: A 1:6 extract of Strychnine
and Atractylodes macrocephala is strychnine and
Atractylenoloid II component group. |
Discussion
RA is a chronic, multifactorial inflammatory immune
disease, the primary symptom of which is synovitis (46). In the early stages of the disease,
pain and swelling of the joints are the primary manifestations,
and, as the disease progresses, the joints become increasingly
stiff and deformed. In severe cases this can lead to disability and
thus seriously affect the quality of life of a patient; in
addition, the cost of lengthy treatment adds to the burden of
living. At present, the complexity of the pathogenesis of RA means
that the current understanding of this disease and its underlying
causes is incomplete, resulting in unsatisfactory treatment
outcomes (47,48). Although non-steroidal
anti-inflammatory drugs, glucocorticosteroids and biological agents
(such as etanercept, infliximab and adalimumab.) have their own
advantages in the treatment of RA, they also have a slow onset of
action, are prone to relapse when stopped, are expensive, can
induce gastrointestinal reactions and increase the risk of
cardiovascular disease (49). It
is therefore important to actively investigate alternative
therapeutic approaches, such as complementary and alternative
medicines, in addition to conventional medication.
Strychnine has been used in traditional Chinese
medicines for >600 years and is widely used for rheumatism,
swelling and pain (50,51). However, as toxic drugs, strychnine
and brucine are both active ingredients and primary components that
can cause poisoning and, in severe cases, it may lead to death
(51). Therefore, in clinical
practice, it is important to configure strychnine correctly to
reduce its toxic component content. The combination of
Atractylodes macrocephala and strychnine can reduce the
level of toxic components of strychnine and enhance its analgesic
and anti-inflammatory effects.
To the best of our knowledge, the present study was
the first to reveal through CCK-8 experiments that when cells were
treated with 780 µg/ml strychnine for 24 h, cell viability was
significantly reduced compared with the control group, and this
effect was time-dependent. Data from subsequent flow cytometry
experiments demonstrated significant differences in the apoptotic
rate of MH7A cells in the Control group following treatment with
MTX, Strychnine, S:A 1:6 and S:A 1:6 component, indicating that the
combination promoted apoptosis in synovial cells. In the follow-up
experimental data, treatment with strychnine with Atractylodes
macrocephala extract reduced the inflammatory response via
IL-1β-induced activation of TLR4, NF-κB and NLRP3 inflammatory
vesicles in MH7A cells.
TLR4/NF-κB/NLRP3 has been indicated to be involved
in the inflammatory response and apoptosis (52,53).
TLR-mediated signaling pathways are needed in the development of
inflammation, and they play an integral role in the innate immune
system as one of the most important pattern recognition systems
(54), and modulation of TLR4
signaling pathways may have potential therapeutic advantages in the
treatment of RA.
The present study demonstrated that in
IL-1β-mediated MH7A cells, the TLR4 receptor was primarily
expressed at the cell membrane surface. It has been established
that when TLR4 is activated it induces inflammatory cells to
secrete large quantities of inflammatory cytokines such as TNF-α,
IL-1β and IL-18 (55,56). IL-1β orchestrates the immune
response at the local and systemic level, stimulating cells to
secrete other inflammatory cytokines (57). From the aforementioned experiments,
it was hypothesized that strychnine combined with Atractylodes
macrocephala extract is a potent anti-inflammatory agent and
its inhibitory effect on inflammation may be associated with the
TLR4 signaling pathway.
When IL-1β binds to the TLR4 receptor, the NF-κB
pathway is activated, and this is associated with pro-inflammatory
cytokine production (58).
Therefore, NF-κB is considered a key target in the treatment of
inflammatory diseases (59). IKK
is a common transcription factor-activating protein, composed of
IKKα, IKKβ and the IKK-related kinases TBK1 and IKKε (60). These kinases are considered to be
major regulators of inflammation and innate immunity through the
control of transcription factors such as NF-κB (61,62).
When the TLR4 signaling pathway is activated by IL-1β and
inflammatory factors are released to act on IKK, the heterodimeric
protein NF-κB is released following phosphorylation of the IκBα
protein bound to NF-κB in the cytoplasmic matrix. This allows it to
be specifically recognized by E3 ubiquitin ligase, which then tags
it with the degradation signal K48-type polyubiquitin chains, which
are subsequently specifically recognized and degraded by the 26S
proteasome (63). In the present
study, strychnine combined with Atractylodes macrocephala
extract significantly inhibited IL-1β-induced activation of NF-κB
and NLRP3 phosphorylation in MH7A cells, suggesting that Strychnine
combined with Atractylodes macrocephala extract may reduce
the levels of pro-inflammatory cytokines through the
TLR4/NF-κB/NLRP3 pathway.
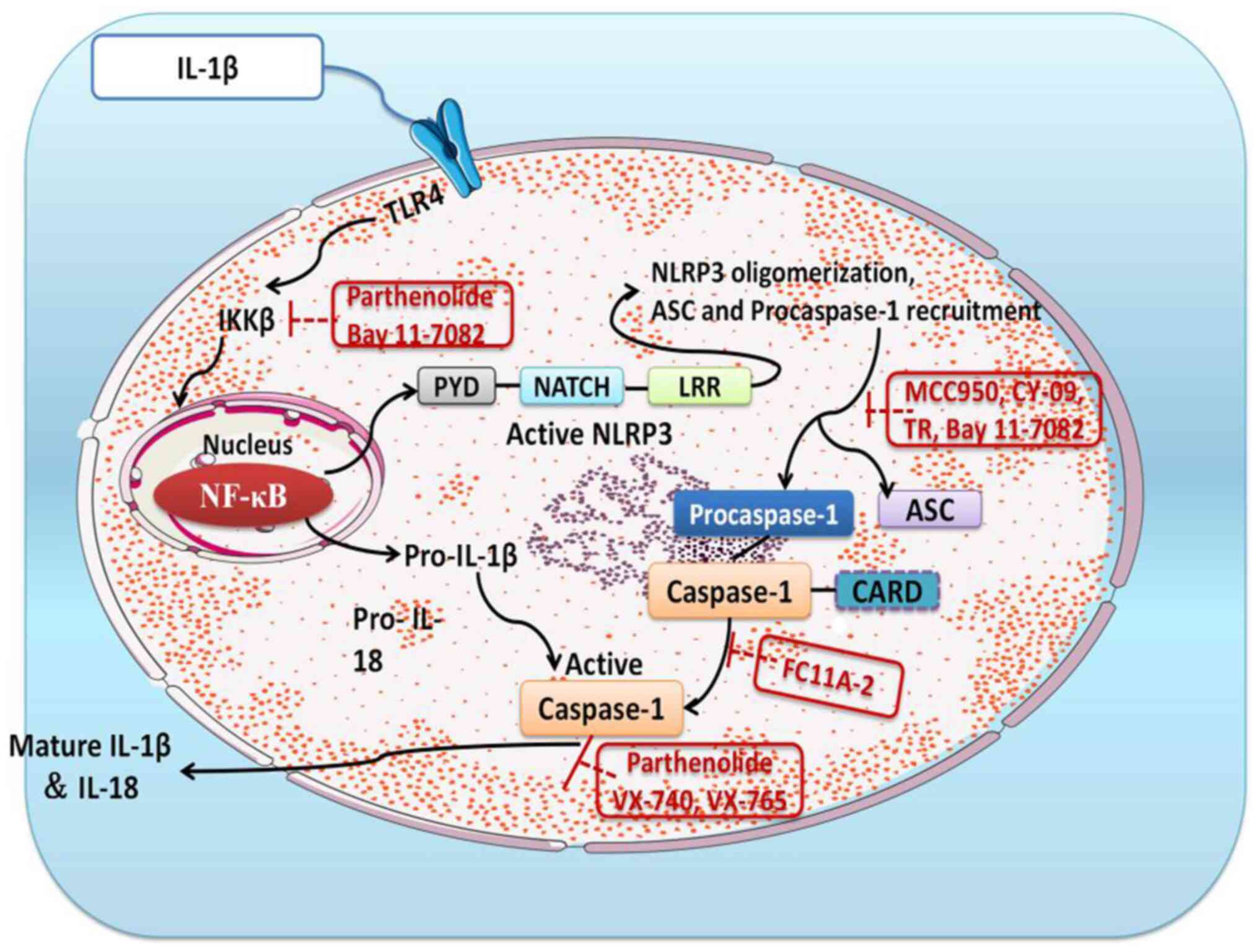
The NLR family of proteins are inflammatory vesicles
present in germline-encoded pattern recognition receptors (64) and include NLRP1, NLRP3 and NLRP4
(65,66). Of these, NLRP3 has been extensively
studied on the previous and consists of three components, NLRP3,
ASC and caspase-1 (67-69).
There is evidence that NLRP3 regulates the production of
pro-inflammatory cytokines (70).
The activation of caspase-1 is conditioned by the conversion of
pro-IL-1β to mature IL-1β, which is dependent on NLPR3 inflammatory
vesicles (71). When it is
activated, the NLPR3 protein will induce translocation and activate
pro-caspase-1 upon binding to the ASC adapter (72). The present study revealed that
strychnine with Atractylodes macrocephala extract could
effectively inhibit TLR4, pNF-κBp65, NF-κBp65, pNLRP3, TNF-κ and
IKKβ protein expression levels in MH7A cells and, thus, it was
hypothesized that this may be due to downregulation of NLPR3
protein expression and inhibition of ASC adapter and caspase-1
activity. Strychnine combined with Atractylodes macrocephala
extract was able to reduce IL-1β-stimulated production of other
inflammatory factors (such as IL-1β and IL-18) in MH7A cells, and
the combination of the two inhibited NLRP3 inflammatory vesicles. A
simplified overview of the above signaling pathway is presented in
Fig. 8.
In conclusion, the enhanced inhibitory effect of
Atractylodes macrocephala extract with strychnine on the
inflammatory response of MH7A cells was identified in the present
study. The results revealed that Atractylodes macrocephala
extract with strychnine promoted apoptosis of synovial cells and
inhibited the expression of TLR4, the NF-κB signaling pathway,
NLRP3 and activation of caspase-1 under IL-1β induction. These
results suggested that strychnine combined with Atractylodes
macrocephala extract exerted its anti-inflammatory effects by
inhibiting the NF-κB signaling pathway and NLRP3 inflammatory
vesicles. This suggested that strychnine in combination with
Atractylodes macrocephala extract may have potential in the
treatment of RA as well as other inflammation-related diseases.
Future experiments are planned to establish an
animal model of rheumatoid arthritis by adjuvant in Wistar rats to
obtain an inflammatory model group. The administration groups will
grouped in the same way as in the cellular experiments, and the
efficacy of the combination of the two will first be verified using
pharmacodynamic experiments, such as foot and plantar swelling
scoring and hot plate in rats. The changes of inflammatory index
factors in rat serum will then analyzed by ELISA assay to verify
whether strychnine and Atractylodes macrocephala had
anti-inflammatory effects. Finally, the relevant indexes will be
detected using immunohistochemistry using the NF-κB inhibitor Bay
11-7082 (73-75).
RT-qPCR assay and western blotting will be performed to verify
whether the combination of strychnine and Atractylodes
macrocephala has anti-inflammatory effects through
TLR4/NF-κB/NLRP3 signaling pathway in treating RA, and to seek
evidence that Strychnos and Atractylodes macrocephala II are
the main active ingredients in the treatment.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YG, XDL and YT conceived and designed the study. YG
and DX performed the experiments and analyzed the results. YG, DX
and XDL were involved in editing the graphs and drafting the
manuscript. YT and XDL confirm the authenticity of all the raw
data. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent to participate
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Liu W, Zhang Y, Zhu W, Ma C, Ruan J, Long
H and Wang Y: Sinomenine inhibits the progression of rheumatoid
arthritis by regulating the secretion of inflammatory cytokines and
monocyte/macrophage subsets. Front Immunol. 9(2228)2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Korczowska I: Rheumatoid arthritis
susceptibility genes: An overview. World J Orthop. 5:544–549.
2014.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Khurana R and Berney SM: Clinical aspects
of rheumatoid arthritis. Pathophysiology. 12:153–165.
2005.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Scott DL, Wolfe F and Huizinga TW:
Rheumatoid arthritis. Lancet. 376:1094–1108. 2010.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Kotake S, Sato K, Kim KJ, Takahashi N,
Udagawa N, Nakamura I, Yamaguchi A, Kishimoto T, Suda T and
Kashiwazaki S: Interleukin-6 and soluble interleukin-6 receptors in
the synovial fluids from rheumatoid arthritis patients are
responsible for osteoclast-like cell formation. J Bone Miner Res.
11:88–95. 1996.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Koopman FA, Chavan SS, Miljko S, Grazio S,
Sokolovic S, Schuurman PR, Mehta AD, Levine YA, Faltys M, Zitnik R,
et al: Vagus nerve stimulation inhibits cytokine production and
attenuates disease severity in rheumatoid arthritis. Proc Natl Acad
Sci USA. 113:8284–8289. 2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Thomas R and Cope AP: Oxford textbook of
rheumatology. Oxford University Press, Oxford, UK, 2015.
|
|
8
|
McInnes IB and Schett G: The pathogenesis
of rheumatoid arthritis. N Engl J Med. 365:2205–2219.
2011.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Syngle D, Singh A and Verma A: Impact of
rheumatoid arthritis on work capacity impairment and its
predictors. Clin Rheumatol. 39:1101–1109. 2020.PubMed/NCBI View Article : Google Scholar
|
|
10
|
van den Bemt BJ, Zwikker HE and van den
Ende CH: Medication adherence in patients with rheumatoid
arthritis: A critical appraisal of the existing literature. Expert
Rev Clin Immunol. 8:337–351. 2012.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Korani S, Korani M, Butler AE and Sahebkar
A: Genetics and rheumatoid arthritis susceptibility in Iran. J Cell
Physiol. 234:5578–5587. 2019.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Lee SY, Cho ML, Oh HJ, Ryu JG, Park MJ,
Jhun JY, Park MK, Stone JC, Ju JH, Hwang SY, et al:
Interleukin-2/anti-interleukin-2 monoclonal antibody immune complex
suppresses collagen-induced arthritis in mice by fortifying
interleukin-2/STAT5 signaling pathways. Immunology. 137:305–316.
2012.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Cooles FA, Isaacs JD and Anderson AE: Treg
cells in rheumatoid arthritis: An update. Curr Rheumatol Rep.
15(352)2013.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Pan F, Zhu L, Lv H and Pei C: Quercetin
promotes the apoptosis of fibroblast-like synoviocytes in
rheumatoid arthritis by upregulating lncRNA MALAT1. Int J Mol Med.
38:1507–1514. 2016.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Yang J, Zhao F and Nie J: Anti-rheumatic
effects of Aconitum leucostomum Worosch. On human fibroblast-like
synoviocyte rheumatoid arthritis cells. Exp Ther Med. 14:453–460.
2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
McInnes IB and Schett G: Pathogenetic
insights from the treatment of rheumatoid arthritis. Lancet.
389:2328–2337. 2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Zhu YG and Qu JM: Toll like receptors and
inflammatory factors in sepsis and differential expression related
to age. Chin Med J (Engl). 120:56–61. 2007.PubMed/NCBI
|
|
18
|
Qing YF, Zhang QB, Zhou JG and Jiang L:
Changes in toll-like receptor (TLR)4-NFκB-IL1β signaling in male
gout patients might be involved in the pathogenesis of primary
gouty arthritis. Rheumatol Int. 34:213–220. 2014.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Chang YY, Jean WH, Lu CW, Shieh JS, Chen
ML and Lin TY: Nicardipine inhibits priming of the NLRP3
inflammasome via suppressing LPS-induced TLR4 expression.
Inflammation. 43:1375–1386. 2020.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Martinon F, Burns K and Tschopp J: The
inflammasome: A molecular platform triggering activation of
inflammatory caspases and processing of proIL-beta. Mol Cell.
10:417–426. 2002.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Agostini L, Martinon F, Burns K, McDermott
MF, Hawkins PN and Tschopp J: NALP3 forms an IL-1beta-processing
inflammasome with increased activity in Muckle-Wells
autoinflammatory disorder. Immunity. 20:319–325. 2004.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Schroder K and Tschopp J: The
inflammasomes. Cell. 140:821–832. 2010.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Strowig T, Henao-Mejia J, Elinav E and
Flavell R: Inflammasomes in health and disease. Nature.
481:278–286. 2012.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Xu XY, Cai BC, Pan Y and Wang TS:
Pharmacokinetics of the alkaloids from the processed seeds of
Strychnos nux-vomica in rats. Yao Xue Xue Bao. 38:458–461.
2003.PubMed/NCBI(In Chinese).
|
|
25
|
Tang M, Zhu WJ, Yang ZC and He CS: Brucine
inhibits TNF-α-induced HFLS-RA cell proliferation by activating the
JNK signaling pathway. Exp Ther Med. 18:735–740. 2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Shu G, Mi X, Cai J, Zhang X, Yin W, Yang
X, Li Y, Chen L and Deng X: Brucine, an alkaloid from seeds of
Strychnos nux-vomica Linn., represses hepatocellular
carcinoma cell migration and metastasis: The role of hypoxia
inducible factor 1 pathway. Toxicol Lett. 222:91–101.
2013.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Rao PS and Prasad MN: Strychnos
nux-vomica root extract induces apoptosis in the human multiple
myeloma cell line-U266B1. Cell Biochem Biophys. 66:443–450.
2013.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Chen J, Qu Y, Wang D, Peng P, Cai H, Gao
Y, Chen Z and Cai B: Pharmacological evaluation of total alkaloids
from nux vomica: Effect of reducing strychnine contents. Molecules.
19:4395–4408. 2014.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Li Y, Liu Y, Shao Y, Zhang F, Liu W, Liang
X and Chen L: Mechanism of action of strychni semen for treating
rheumatoid arthritis and methods for attenuating the toxicity. Comb
Chem High Throughput Screen. 25:587–606. 2022.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Guo R, Wang T, Zhou G, Xu M, Yu X, Zhang
X, Sui F, Li C, Tang L and Wang Z: Botany, phytochemistry,
pharmacology and toxicity of Strychnos nux-vomica L.: A
review. Am J Chin Med. 46:1–23. 2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Qin JM, Yin PH, Li Q, Sa ZQ, Sheng X, Yang
L, Huang T, Zhang M, Gao KP, Chen QH, et al: Anti-tumor effects of
brucine immuno-nanoparticles on hepatocellular carcinoma. Int J
Nanomedicine. 7:369–379. 2012.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Chen J, Hou T, Fang Y, Chen ZP, Liu X, Cai
H, Lu TL, Yan GJ and Cai BC: HPLC determination of strychnine and
brucine in rat tissues and the distribution study of processed
semen strychni. Yakugaku Zasshi. 131:721–729. 2011.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Dai J, Liu J, Zhang M, Yu Y and Wang J:
Network toxicology and molecular docking analyses on strychnine
indicate CHRM1 is a potential neurotoxic target. BMC Complement Med
Ther. 22(273)2022.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Otter J and D'Orazio JL: Strychnine
toxicity. In: StatPearls [Internet]. Treasure Island (FL):
StatPearls Publishing, 2022.
|
|
35
|
Lu L, Huang R, Wu Y, Jin JM, Chen HZ,
Zhang LJ and Luan X: Brucine: A review of phytochemistry,
pharmacology, and toxicology. Front Pharmacol.
11(377)2020.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Zhao C, Li E, Wang Z, Tian J, Dai Y, Ni Y,
Li F, Ma Z and Lin R: Nux vomica exposure triggered liver injury
and metabolic disturbance in zebrafish larvae. Zebrafish.
15:610–628. 2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Fan Y, Liu S, Chen X, Feng M, Song F and
Gao X: Toxicological effects of nux vomica in rats urine and serum
by means of clinical chemistry, histopathology and 1H
NMR-based metabonomics approach. J Ethnopharmacol. 210:242–253.
2018.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Li Y, Wang J, Xiao Y, Wang Y, Chen S, Yang
Y, Lu A and Zhang S: A systems pharmacology approach to investigate
the mechanisms of action of semen strychni and Tripterygium
wilfordii Hook F for treatment of rheumatoid arthritis. J
Ethnopharmacol. 175:301–314. 2015.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Lü D, Jiang Q, Zhang J, Zeng R, Liao Z and
Liang X: Effect of baizhu (rhizoma atractylodis macrocephalae)
extract on intestinal absorption of brucine and strychnine in vitro
and in situ. J Tradit Chin Med. 40:562–570. 2020.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Emery P, Breedveld FC, Hall S, Durez P,
Chang DJ, Robertson D, Singh A, Pedersen RD, Koenig AS and
Freundlich B: Comparison of methotrexate monotherapy with a
combination of methotrexate and etanercept in active, early,
moderate to severe rheumatoid arthritis (COMET): A randomised,
double-blind, parallel treatment trial. Lancet. 372:375–382.
2008.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Friedman B and Cronstein B: Methotrexate
mechanism in treatment of rheumatoid arthritis. Joint Bone Spine.
86:301–307. 2019.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Alarcón GS, Tracy IC and Blackburn WJ Jr:
Methotrexate in rheumatoid arthritis. Toxic effects as the major
factor in limiting long-term treatment. Arthritis Rheum.
32:671–676. 1989.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Yu Z, Liu H, Fan J, Chen F and Liu W:
MicroRNA-155 participates in the expression of LSD1 and
proinflammatory cytokines in rheumatoid synovial cells. Mediators
Inflame. 2020(4092762)2020.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Brunelle JL and Green R: Coomassie blue
staining. Methods Enzymol. 541:161–167. 2014.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Dörner T, Vital EM, Ohrndorf S, Alten R,
Bello N, Haladyj E and Burmester G: A narrative literature review
comparing the key features of musculoskeletal involvement in
rheumatoid arthritis and systemic lupus erythematosus. Rheumatol
Ther. 9:781–802. 2022.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Radu AF and Bungau SG: Management of
rheumatoid arthritis: An overview. Cells. 10(2857)2021.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Chauhan K, Jandu JS, Goyal A and Al-Dhahir
MA: Rheumatoid arthritis. In: StatPearls. StatPearls Publishing,
Treasure Island, FL, 2022.
|
|
49
|
Zhong Y, Lai D, Zhang L, Lu W, Shang Y and
Zhou H: The effects of moxibustion on PD-1/PD-L1-related molecular
expression and inflammatory cytokine levels in RA rats. Evid Based
Complement Alternat Med. 2021(6658946)2021.
|
|
50
|
Li S, Chu Y, Zhang R, Sun L and Chen X:
Prophylactic neuroprotection of total glucosides of paeoniae radix
alba against semen strychni-induced neurotoxicity in rats:
Suppressing oxidative stress and reducing the absorption of toxic
components. Nutrients. 10(514)2018.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Tong HF, Chan CY, Ng SW and Mak TWL:
Strychnine poisoning due to traditional Chinese medicine: A case
series. F1000Res. 10(924)2021.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Cao B, Wang T, Qu Q, Kang T and Yang Q:
Long noncoding RNA SNHG1 promotes neuroinflammation in Parkinson's
disease via regulating miR-7/NLRP3 pathway. Neuroscience.
388:118–127. 2018.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Bachmaier K, Toya S, Gao X, Triantafillou
T, Garrean S, Park GY, Frey RS, Vogel S, Minshall R, Christman JW,
et al: E3 ubiquitin ligase Cblb regulates the acute inflammatory
response underlying lung injury. Nat Med. 13:920–926.
2007.PubMed/NCBI View
Article : Google Scholar
|
|
54
|
Xiang P, Chen T, Mou Y, Wu H, Xie P, Lu G,
Gong X, Hu Q, Zhang Y and Ji H: NZ suppresses TLR4/NF-κB signalings
and NLRP3 inflammasome activation in LPS-induced RAW264.7
macrophages. Inflamm Res. 64:799–808. 2015.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Pålsson-McDermott EM and O'Neill LAJ:
Signal transduction by the lipopolysaccharide receptor, Toll-like
receptor-4. Immunology. 113:153–162. 2004.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Wang Y, Cui Y, Cao F, Qin Y, Li W and
Zhang J: Ganglioside GD1a suppresses LPS-induced pro-inflammatory
cytokines in RAW264.7 macrophages by reducing MAPKs and NF-κB
signaling pathways through TLR4. Int Immunopharmacol. 28:136–145.
2015.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Malik A and Kanneganti TD: Function and
regulation of IL-1α in inflammatory diseases and cancer. Immunol
Rev. 281:124–137. 2018.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Zusso M, Lunardi V, Franceschini D,
Pagetta A, Lo R, Stifani S, Frigo AC, Giusti P and Moro S:
Ciprofloxacin and levofloxacin attenuate microglia inflammatory
response via TLR4/NF-kB pathway. J Neuroinflammation.
16(148)2019.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Wu XL, Liou CJ, Li ZY, Lai XY, Fang LW and
Huang WC: Sesamol suppresses the inflammatory response by
inhibiting NF-κB/MAPK activation and upregulating AMP kinase
signaling in RAW 264.7 macrophages. Inflamm Res. 64:577–588.
2015.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Antonia RJ, Hagan RS and Baldwin AS:
Expanding the view of IKK: New substrates and new biology. Trends
Cell Biol. 31:166–178. 2021.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Wu J and Chen ZJ: Innate immune sensing
and signaling of cytosolic nucleic acids. Annu Rev Immunol.
32:461–488. 2014.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Helgason E, Phung QT and Dueber EC: Recent
insights into the complexity of Tank-binding kinase 1 signaling
networks: the emerging role of cellular localization in the
activation and substrate specificity of TBK1. FEBS Lett.
587:1230–1237. 2013.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Di Rita A, Peschiaroli A, D Acunzo P,
Strobbe D, Hu Z, Gruber J, Nygaard M, Lambrughi M, Melino G,
Papaleo E, et al: HUWE1 E3 ligase promotes PINK1/PARKIN-independent
mitophagy by regulating AMBRA1 activation via IKKα. Nat Commun.
9(3755)2018.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Takeuchi O and Akira S: Pattern
recognition receptors and inflammation. Cell. 140:805–820.
2010.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Lamkanfi M and Dixit VM: Mechanisms and
functions of inflammasomes. Cell. 157:1013–1022. 2014.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Sharma D and Kanneganti TD: The cell
biology of inflammasomes: Mechanisms of inflammasome activation and
regulation. J Cell Biol. 213:617–629. 2016.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Li Z, Guo J and Bi L: Role of the NLRP3
inflammasome in autoimmune diseases. Biomed Pharmacother.
130(110542)2020.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Mangan MSJ, Olhava EJ, Roush WR, Seidel
HM, Glick GD and Latz E: Targeting the NLRP3 inflammasome in
inflammatory diseases. Nat Rev Drug Discov. 17:588–606.
2018.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Shao BZ, Xu ZQ, Han BZ, Su DF and Liu C:
NLRP3 inflammasome and its inhibitors: A review. Front Pharmacol.
6(262)2015.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Luo YP, Jiang L, Kang K, Fei DS, Meng XL,
Nan CC, Pan SH, Zhao MR and Zhao MY: Hemin inhibits NLRP3
inflammasome activation in sepsis-induced acute lung injury,
involving heme oxygenase-1. Int Immunopharmacol. 20:24–32.
2014.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Sun L, Ma W, Gao W, Xing Y, Chen L, Xia Z,
Zhang Z and Dai Z: Propofol directly induces caspase-1-dependent
macrophage pyroptosis through the NLRP3-ASC inflammasome. Cell
Death Dis. 10(542)2019.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Chao LK, Lin CH, Chiu HW, Wong WT, Chiu
HW, Tasi YL, Kuo YH, Chiu YC, Liu ML, Ho CL and Hua KF:
Peroxyauraptenol inhibits inflammation and NLRP3 inflammasome
activation by inhibiting reactive oxygen species generation and
preserving mitochondrial integrity. J Agric Food Chem.
63:1210–1219. 2015.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Lee J, Rhee MH, Kim E and Cho JY: BAY
11-7082 is a broad-spectrum inhibitor with anti-inflammatory
activity against multiple targets. Mediators Inflamm.
2012(416036)2012.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Xia ZB, Meng FR, Fang YX, Wu X, Zhang CW,
Liu Y, Liu D, Li GQ, Feng FB and Qiu HY: Inhibition of NF-κB
signaling pathway induces apoptosis and suppresses proliferation
and angiogenesis of human fibroblast-like synovial cells in
rheumatoid arthritis. Medicine (Baltimore).
97(e10920)2018.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Zahid A, Li B, Kombe AJK, Jin T and Tao J:
Pharmacological inhibitors of the NLRP3 inflammasome. Front
Immunol. 10(2538)2019.PubMed/NCBI View Article : Google Scholar
|