Introduction
Colorectal cancer (CRC) is the third most prevalent
type of cancer worldwide and the second largest cause of
cancer-related mortality (1).
Among the individuals newly diagnosed with CRC, 20% of them present
with metastatic disease upon presentation to the physician and
another 25% will develop metastases following the manifestation of
localized disease (2). The 5-year
survival rate of patients with CRC who are diagnosed at an early
stage is 90%, and patients with distant metastases exhibit a 5-year
survival rate of 10% (3). Studies
have revealed that mortality from CRC may be reduced through
screening and early detection (4).
In recent years, a number of novel biomarkers have been identified
and applied for early diagnosis, personalized treatment selection
and the prognosis of CRC, with advances being made in research on
various aspects of CRC (5).
Regardless of the progress made in CRC diagnosis and the
determination of patient prognosis (6), the underlying molecular mechanisms
require further investigation.
The cell adhesion molecule close homolog of L1
(CHL1) belongs to the cell adhesion molecule L1 (L1CAM) gene family
(7), which is located at 3p26 and
is involved in certain neurological diseases (8). Recently, CHL1 was defined to be
involved in signal transduction pathways, as well as in the
development of various human cancers, including breast cancer, CRC,
bladder cancer and lung cancer (9,10).
The downregulation of CHL1 expression has been observed in a
variety of human tumors, including neuroblastoma, and breast and
nasopharyngeal cancer (9,11,12).
The downregulation of CHL1 has been shown to inhibit the growth and
metastasis of nasopharyngeal carcinoma tumors by suppressing the
PI3K/AKT signaling pathway (11).
Ognibene et al (12)
indicated that the CHL1 gene plays an tumor suppressor role in
human adult neuroblastoma. Qin et al (13) revealed that a reduced CHL1
expression predicted a poor prognosis of patients with renal clear
cell carcinoma. However, Senchenko et al (9) detected the upregulated expression of
the CHL1 gene in lung, ovarian, uterine, liver and tracheal
cancers, in contrast to findings on CHL1 in the majority of other
cancer types, including nasopharyngeal carcinoma tumors, renal
clear cell carcinoma, neuroblastoma and bladder cancer (9,11,12).
Yang et al (14) also
revealed that CHL1 promoted the proliferation, metastasis and
migration of human glioma cells both in vivo and in
vitro. These findings indicate that CHL1 expression may play a
bidirectional regulatory role in cancer. Concerning the role of
CHL1 in CRC, Yu et al (15)
revealed that microRNA-21-5p induced colon adenocarcinoma cell
proliferation and invasion by targeting CHL1. They defined CHL1 as
a tumor suppressor gene in colon cancer (15). However, only a limited number of
studies have reported the role of CHL1 in CRC, and to the best of
our knowledge, no studies to date have directly explored its
specific functions and mechanisms of action in CRC. Therefore, the
present study aimed to evaluate the specific biological functions
and mechanisms of CHL1 in order to provide a theoretical basis for
the use of CHL1 as a biological target in CRC.
Materials and methods
Cell lines and cell culture
CRC cell lines (HT29, SW480, SW620 and HCT116) were
purchased from Cellcook Biotech Co., Ltd. The company performed
short tandem repeat validation on all of the aforementioned cells.
The corresponding catalogue numbers were as follows: HT-29, cat.
no. CC0504; HCT-116, cat. no. CC0506; SW480, cat. no. CC0505; and
SW620, cat. no. CC0503. The cells were cultured in Dulbecco's
modified Eagle's medium (Gibco; Thermo Fisher Scientific, Inc.),
including 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) and penicillin/streptomycin (Gibco; Thermo Fisher
Scientific, Inc.). The cells were then maintained in a cell
incubator of 5% CO2 at 37˚C.
Cell transfection
General Biologicals Co., Ltd. constructed and
synthesized the overexpression plasmid pcDNA3.1-CHL1 (CHL1 group,
the name of the gene of interest is CHL1), pcDNA3.1-RELA (RELA
group, the name of the gene of interest is RELA), and the control
vector pcDNA3.1 [normal control (NC) group, Invitrogen, Carlsbad,
CA, USA]. These plasmids (5 µg) were transfected into the cells
using Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's instructions. After 6 h of
incubation at 37˚C, culture was continued with the replacement of
complete medium and continued for 48 h prior to further
experiments.
Organoid culture
Organoid culture is a 3D culture model. In contrast
to 2D culture, a matrix gel is used as a 3D culture scaffold to
maintain the 3D structure. The culture plate is of a low-adsorption
type, and the 3D culture is more closely related to the in
vivo environment in comparison with the 2D culture,
particularly concerning cell-cell interactions. In the present
study, organoid culture was performed based on the descriptions of
a previous study (16). The HT29
and SW480 cells were digested into single cells and transfected
with pcDNA3.1-CHL1 (pcDNA3.1-RELA) or the control vector for 2 h,
Subsequently, the cells were resuspended with CRC organoid culture
medium (Orgen Biotech) and resuspended 1:1 with matrix glue (BD
Biosciences). Following coagulation, the organoid culture medium
was added to 48-well plates (Biofil) and placed in a cell incubator
of 5% CO2 at 37˚C to observe the growth of organoids
daily.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the HT29, SW480, SW620
and HCT116 cells using TRizol® reagent (Invitrogen;
Thermo Fisher Scientific, Inc.), and the concentration of RNA was
detected using a NanoDrop spectrophotometer (Thermo Fisher
Scientific, Inc.). The cDNA was prepared using the PrimeScript RT
kit (Takara Bio, Inc.), and the expression level of CHL1 was
examined following the instructions of the SYBR-Green Quantitative
PCR kit (Takara Bio, Inc.). The thermocycling conditions were as
follows: Initial denaturation for 2 min at 95˚C, followed by 40
cycles of 15 sec at 95˚C and 30 sec at 60˚C. Relative gene
expression was determined using the 2-IICq method
(17). GAPDH was used as an
internal control. The primer sequences were as follows: CHL1
forward, 5'-TGGAAAACCGATCACGGAGG-3' and reverse,
5'-TCAGCTCCCGGCTCAAATAC-3'; and GAPDH forward,
5'-GGTGAAGGTCGGAGTCAACG-3' and reverse,
5'-CAAAGTTGTCATGGATGHACC-3'.
Western blotting
Total proteins were extracted from the transfected
cells (HT-29 and SW480) using RIPA lysis buffer (MilliporeSigma). A
BCA protein assay kit (Beyotime Institute of Biotechnology) was
used to measure the protein concentration. A total quantity of 20
µg of protein was then separated using 12% SDS-PAGE and then
transferred to the polyvinylidene fluoride membranes
(MilliporeSigma). After blocking the membrane with 5% skimmed milk
for 1 h at 25˚C, it was incubated overnight with primary antibody
E-cadherin (cat. no. 20874-1-AP, Proteintech Group, Inc., dilution
1:20,000), N-cadherin (cat. no. 22018-1-AP, Proteintech Group,
Inc., dilution 1:2,000), CHL1 (cat. no. 25250-1-AP, Proteintech
Group, Inc., dilution 1:500), p-p65 (cat. no. AP0475; ABclonal,
Inc., dilution 1:500), p65 (cat. no. 10745-1-AP, Proteintech Group,
Inc., dilution 1:2,000) and GAPDH (cat. no. 10494-1-AP, Proteintech
Group, Inc., dilution 1:6,000) at 4˚C. Subsequently, after washing
the membrane with Tris-buffered saline wash buffer with 5% Tween-20
(TBST) for thrice (5 min per time), the membrane was incubated with
horseradish peroxidase-conjugated secondary antibodies [Anti-Rabbit
IgG (H+L), cat. no. SA00004-2, Proteintech Group, Inc., dilution
1:50,000] at room temperature for 2 h. The membrane was washed
thrice with TBST (10 min per time) and visualized by ECL
chemiluminescent reagent (cat. no. PK10001, Proteintech Group,
Inc.) to acquire images. The gray level of protein bands was
determined using ImageJ software (version 1.8.0, National
Institutes of Health). GAPDH was used as an internal reference.
Cell Counting Kit-8 (CCK-8)
The CCK-8 assay (Beyotime Institute of
Biotechnology) was used to detect cell proliferation. The
transfected cells (HT-29 and SW480) were collected by digestion and
inoculated in 96-well plates (Biofil) at a density of
5x103 cells/well. A total quantity of 20 µl of the CCK-8
reagent was added to each well of cells after 24, 48 and 72 h of
incubation and then incubated for 4 h at 37˚C. A microplate reader
(Bio-Rad 168-1130 iMark, Bio-Rad Laboratories, Inc.) at 450 nm was
used to determine the optical density (OD) of each well.
Clone formation assay
Clone formation reflects cell population dependence
and proliferation ability (18).
In the present study, cells (HT-29 and SW480) were digested with
0.25% trypsin (Thermo Fisher Scientific, Inc.) and prepared into a
cell suspension, adjusted to 1,000 cells/ml. They were seeded in
six-well plates (Biofil) at a density of 50, 100 and 200
cells/well. The culture medium was changed every 3 days. The
culture was terminated when there was obvious colony formation in
the culture dish. The cells were stained with 0.1% crystal violet
(Beijing Solarbio Science & Technology Co., Ltd.) for 20 min at
25˚C after 15 min of fixation with 4% paraformaldehyde at 25˚C.
Colonies containing >50 cells were counted and analyzed.
Transwell assay
Transwell assay was used to measure cell migration
and invasion. The upper chamber of the Transwell (8.0 µm pore
polycarbonate membrane insert, Corning, Inc.) was supplemented with
100 µl cell suspension and 700 µl medium that contained 10% FBS was
added to the lower chamber for cell migration assay. The
non-migratory cells in the upper chamber were removed following 24
h of incubation at 37˚C. Cells that migrated to the lower chamber
were fixed with 4% paraformaldehyde for 30 min and stained with
0.1% crystalline violet (Beijing Solarbio Science & Technology
Co., Ltd.) for 12 h at 25˚C. The cells were washed with
phosphate-buffered saline and then observed under a light
microscope (Guangzhou Micro-shot Technology Co., Ltd.). Cell
invasion assay was performed with the upper chamber pre-coated with
Matrigel (BD Biosciences) for 10 h at 37˚C. The other operations,
from the upper chamber of the Transwell supplemented with cell
suspension, until the migrated cells were obtained, were consistent
with those of the migration assay.
Immunofluorescence (IF) staining
IF staining was performed as previously described
(19). Cells that were cultured on
coverslips were fixed in 4% paraformaldehyde (cat. no. P1110,
Solarbio life sciences, Inc.) for 15 min at 37˚C, permeabilized
with 0.3% Triton X-100 (cat no. T8200, Beijing Solarbio Science
& Technology Co., Ltd.), and closed with 5% bovine serum (cat.
no. SW3015, Beijing Solarbio Science & Technology Co., Ltd.).
The cells were incubated with antibodies E-cadherin (cat. no.
20874-1-AP, Proteintech Group, Inc., dilution 1:200) or N-cadherin
(cat. no. 22018-1-AP, Proteintech Group, Inc., dilution 1:100)
overnight at 4˚C and then coupled with Alexa Fluor 594 secondary
antibody (cat. no. ab150080, 1:500; Abcam) for 1 h at 25˚C. The
nuclei were stained with DAPI (cat no. C0065, Beijing Solarbio
Science & Technology Co., Ltd.) for 10 min at 25˚C and images
were observed under a fluorescence inverted microscope (Leica DM
IL, Leica Microsystems GmbH).
Tumor xenograft experiment in nude
mice
A total of 12 female BALB/c nude mice (4 to 6 weeks
old; SPF-grade; weighing 18-22 g) were purchased from Guangzhou
Ruige Biotechnology Co., Ltd. The mice were kept in specific
pathogen-free conditions (temperature, 21-23˚C; 12:12 h light-dark
cycle; humidity, 50%) with adequate food and water provided
throughout the entire experimental process. A total of 12 mice were
randomly divided into the NC and CHL1 groups, with 6 mice in each
group. The CHL1-overexpressing and control HT29 stable strains (the
HT-29 cells were used as they had a lower expression of CHL1
compared with the SW480 cells) were respectively resuspended, and
the cell suspension (5x106 cells/each) was
subcutaneously injected into the right axilla of the nude mice. The
tumor size was measured with calipers from day 5 after subcutaneous
injection, and the tumor volume was calculated using the following
formula: Volume=(length x width2)/2. After 35 days, all
nude mice were euthanized using the cervical dislocation method
(mouse breath cessation and loss of response to external stimuli,
loss of heartbeat, breath cessation, pupil dilation). The tumors
were then removed and weighed. Animal health and behavior were
monitored every day. In the case that the tumor volume was
>1,500 mm3, the mice were to be sacrificed with
CO2 (no mouse had a tumor volume >1,500
mm3 during the experiment, therefore no mouse was
sacrificed with CO2). There was no mouse death before
the tumor was removed. The committee of Guangzhou Forevergen
Biosciences Animal Center approved our experiments (Approval no.
IACUC-AEWC-F2023021920). All experiments were performed under the
IACUC Handbook (Third Edition) and reported following ARRIVE
guidelines.
Hematoxylin and eosin (H&E)
staining
All nude mice were euthanized using the cervical
dislocation method, and CRC tissues were fixed in 4%
paraformaldehyde for 24 h at 4˚C. The tissues were placed in an
embedding box, rinsed in running water for 30 min, dehydrated in
ethanol, and embedded in wax immersion in xylene. They were then
cut into 5-µm-thick sections in a pathology slicer (HM340E, Thermo
Fisher Scientific, Inc.) and dried in a thermostat for 20 min at
60˚C. All the following steps were performed at 25˚C. The sections
were deparaffinized in xylene I and II for 10 min, and then placed
into gradient ethanol dehydration and washed with distilled water
in sequence. They were stained with H&E (cat no. G1005, Wuhan
Servicebio Technology Co., Ltd.) for 2 min, and then sequentially
placed in 95% ethanol I and II for 5 min each, anhydrous ethanol I
and II for 5 min each to dehydrate, xylene I and II for transparent
sections, and then removed to air-dry before sealing them with
neutral gum. The histopathologic structures were observed under a
light microscope (Guangzhou Micro-shot Technology Co., Ltd.).
RNA sequencing (RNA-seq)
Library construction and sequencing were performed
as previously described (20,21).
The TruSeq Stranded Total RNA Library Prep kit (cat. no.
RS-122-2302, Illumina, Inc.) was used to construct cDNA libraries
in a strand-specific manner from 4 µg of DNase-treated RNA.
Libraries were quality tested and quantified using a BioAnalyzer
2100 system (Agilent Technologies, Inc.) and RT-qPCR. Total RNA was
then fragmented, cDNA synthesis was performed, and the connectors
were ligated to double-stranded cDNA. Sequencing was conducted on
the Illumina NovaSeq 6000 and HiSeq X Ten platforms (150 nt
paired-end sequencing, Illumina, Inc.). Short reads, adaptors and
low-quality bases were discarded from raw data using the FASTX
Toolkit (Version 0.0.13; hannonlab.cshl.edu/fastx_toolkit/). Clean reads were
aligned to the human GRCh38 genome using HISAT2 (version 2.2.1).
The genes were further analyzed.
Screening of differentially expressed
genes (DEGs)
DEGs in CHL1-overexpressing cells compared with
those of the NC group were determined using the ‘edgeR’ of
Bioconductor (version 3.4) (20).
All gene expression data were log2-transformed. DEGs were
identified with a false discovery rate corrected P<0.05 and |log
fold-change (FC)|>1. The intersection of DEGs was obtained using
Venny 2.1.0 (Juan Carlos Oliveros BioinfoGP, CNB-CSIC, https://bioinfogp.cnb.csic.es/tools/venny/).
Gene Ontology (GO) and Kyoto
Encyclopedia of Genes and Genomes (KEGG)
A widely used web-based genomic functional
annotation tool (DAVID, https://david.ncifcrf.gov/) was used for data
annotation analysis (21).
Molecular function and pathway analyses of the DEGs were performed
using GO analysis and KEGG pathway analysis. GO analysis included
biological processes, cellular components and molecular functions.
P<0.05 was considered to indicate a statistically significant
difference.
Statistical analysis
SPSS version 20.0 software (IBM Corp.) was used for
statistical analyses. Data are expressed as the mean ± standard
deviation. An unpaired Student's t-test was used for comparisons
between two groups, and one-way ANOVA followed by Tukey's post hoc
test was used for comparisons among multiple groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
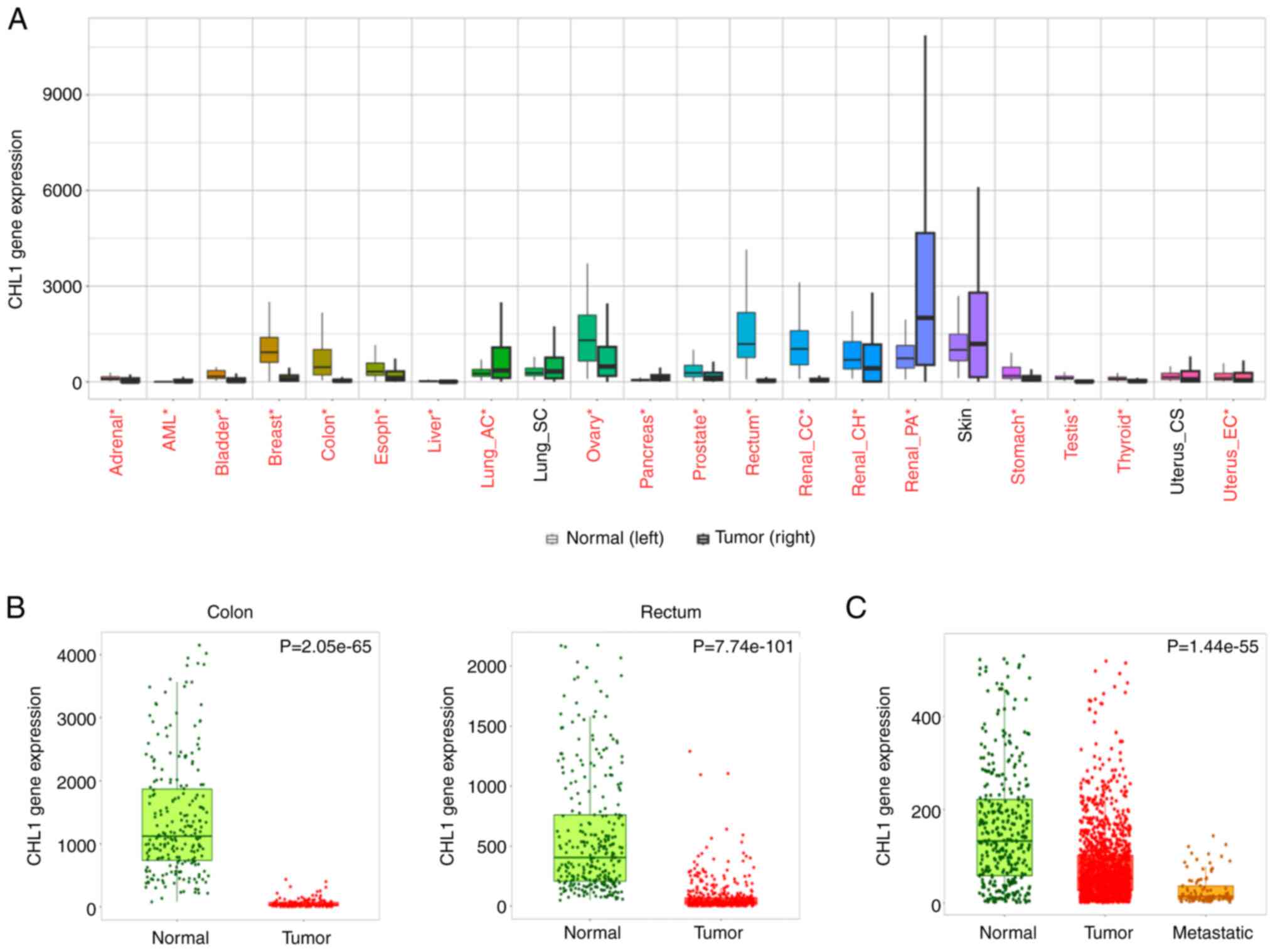
CHL1 is downregulated in CRC
tissues
The gene expression of CHL1 in CRC tissues was
assessed using the TNMplot online database (https://tnmplot.com/analysis/) to determine the
expression of CHL1 in CRC tissues. CHL1 expression was
significantly lower in CRC cancer tissues than in normal tissues,
and even lower in metastatic colon cancer tissues, as shown in
Fig. 1A-C (P<0.05). These
results indicated the involvement of CHL1 in CRC progression.
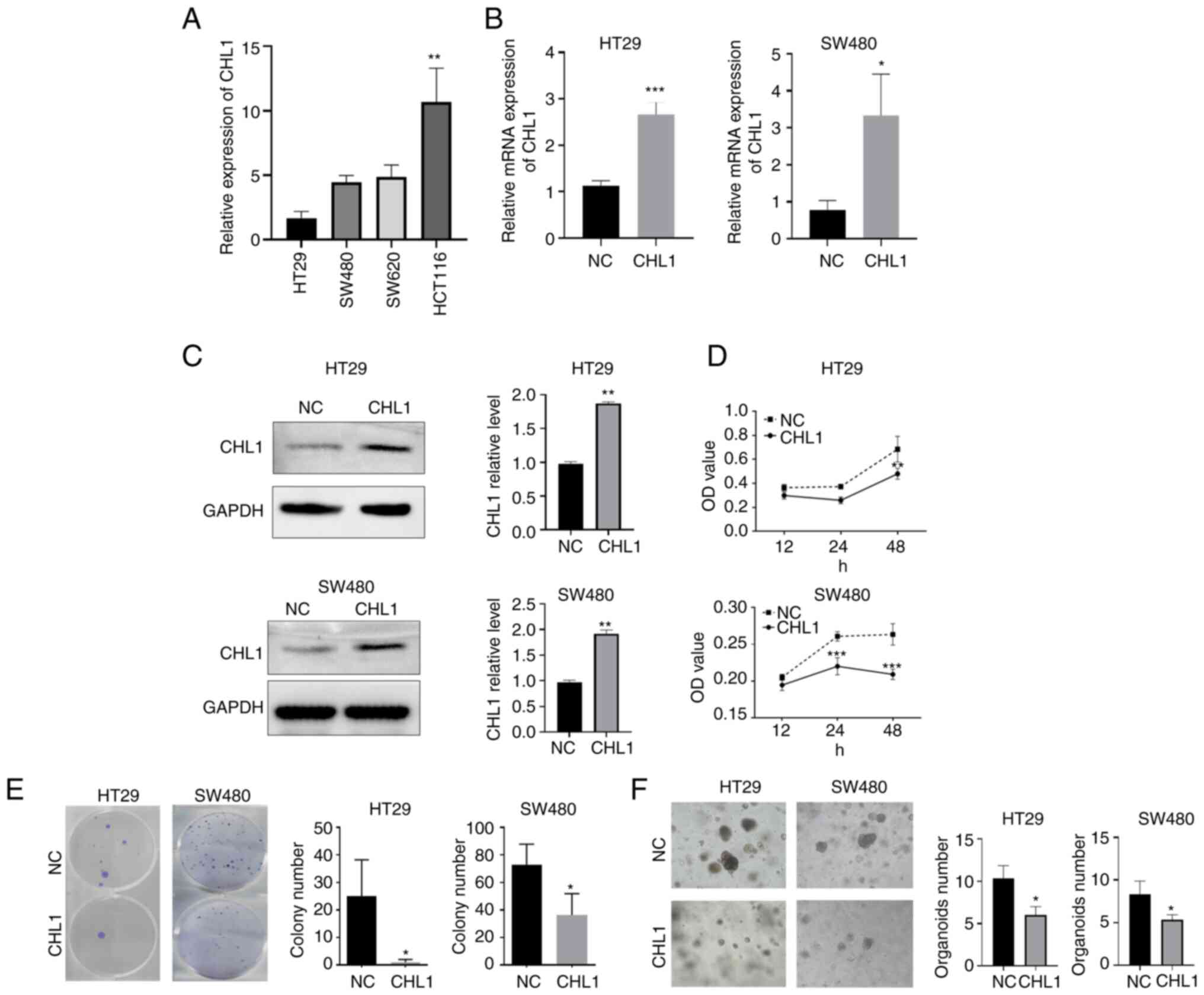
CHL1 inhibits CRC cell
proliferation
CHL1 expression in CRC cell lines was first
examined, in order to determine the function of CHL1 in CRC. The
results of RT-qPCR revealed that the expression of CHL1 was reduced
to a greater extent in HT29 and SW480 cells, among the CRC cell
lines analyzed (Fig. 2A).
Therefore, CHL1 overexpression plasmids were transfected into HT29
and SW480 cells. The results revealed significantly elevated CHL1
mRNA and protein expression levels in the CRC cells following the
transfection with overexpression plasmids (P<0.05; Fig. 2B and C). CCK-8 assay revealed that the HT29 and
SW480 cell proliferative ability was significantly decreased
following the induction of CHL1 overexpression, as compared with
the NC group (P<0.05; Fig. 2D).
Similarly, the results of colony formation assay further confirmed
that CHL1 overexpression significantly inhibited the colony
formation ability of the HT29 and SW480 cells (P<0.05; Fig. 2E). The growth of the HT29 and SW480
cell organoids was also significantly restrained, due to the
transfection with CHL1 overexpression plasmid (P<0.05; Fig. 2F).
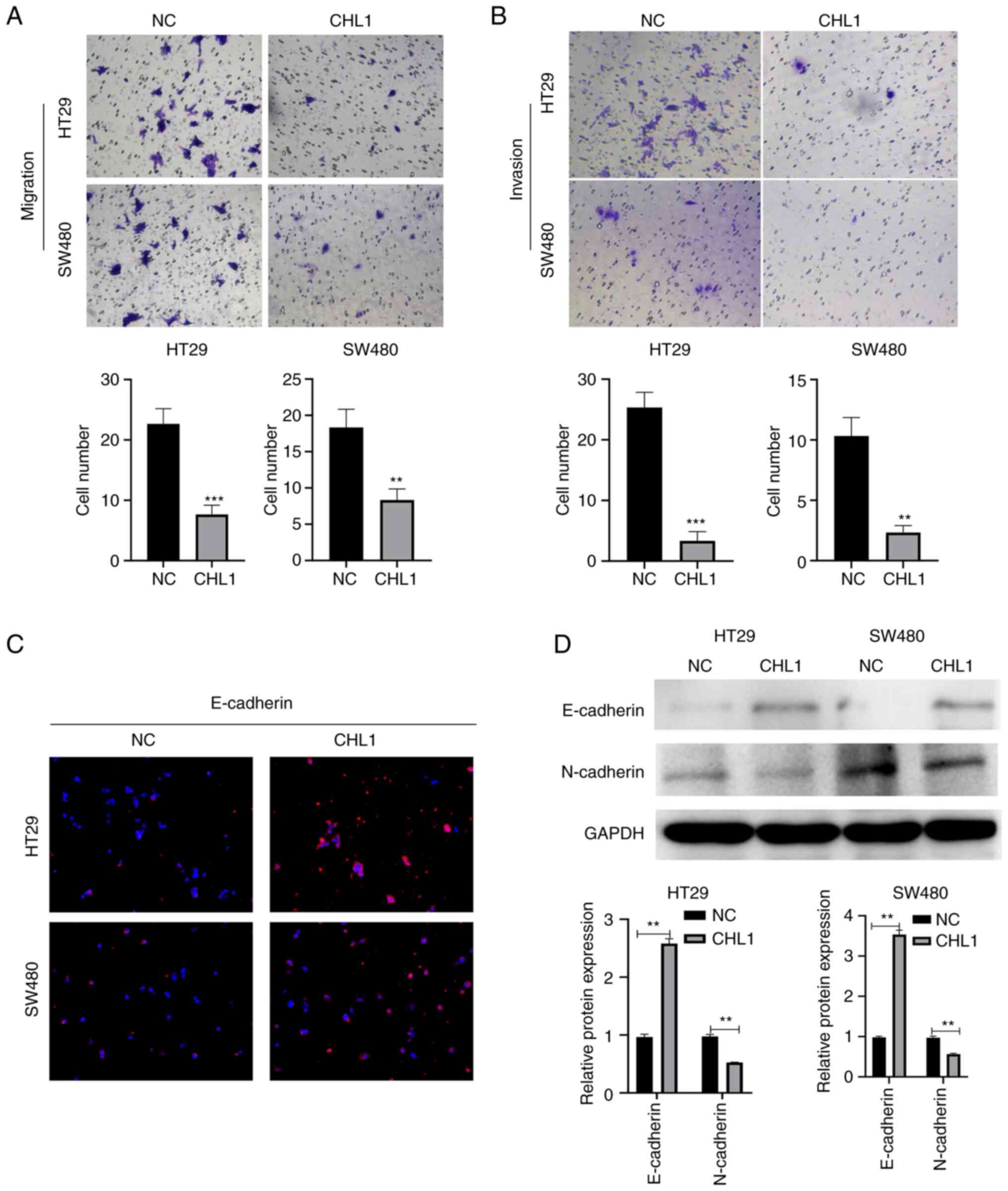
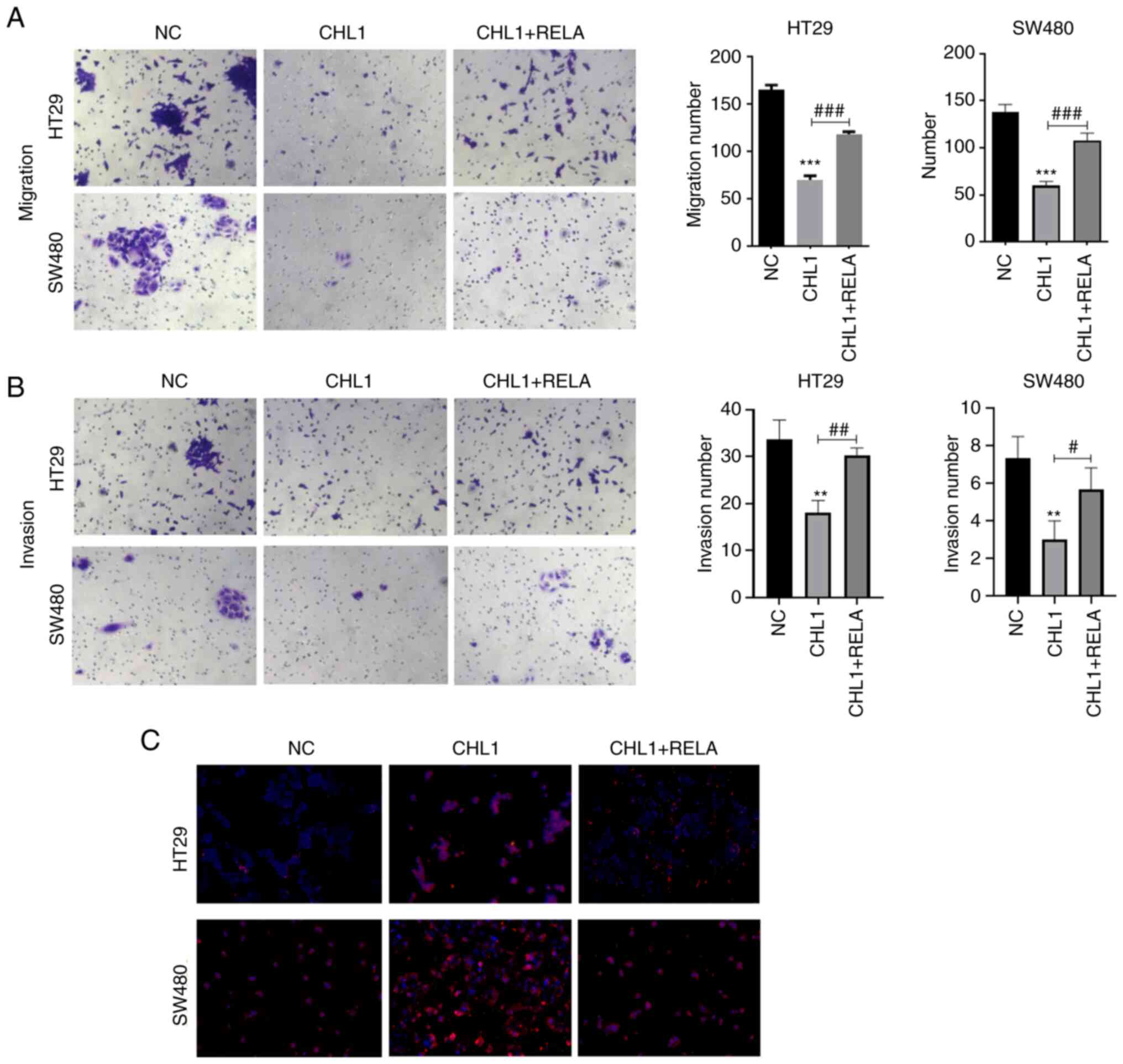
CHL1 inhibits CRC cell migration and
invasion
The cell migratory and invasive ability was then
evaluated using Transwell assay. The results illustrated in
Fig. 3A and B indicated that the HT29 and SW480 cell
migratory and invasive ability was significantly suppressed in the
CHL1 group, in comparison with the NC group (P<0.05).
Immunofluorescence staining also revealed that the protein
expression level of E-cadherin was significantly increased
following the induction of CHL1 overexpression (Fig. 3C). Additionally, the results of
western blotting also revealed that CHL1 overexpression
significantly upregulated E-cadherin and downregulated N-cadherin
expression in HT29 and SW480 cells (Fig. 3D). These results indicated that
CHL1 upregulation inhibits the migratory and invasive ability of
CRC cells.
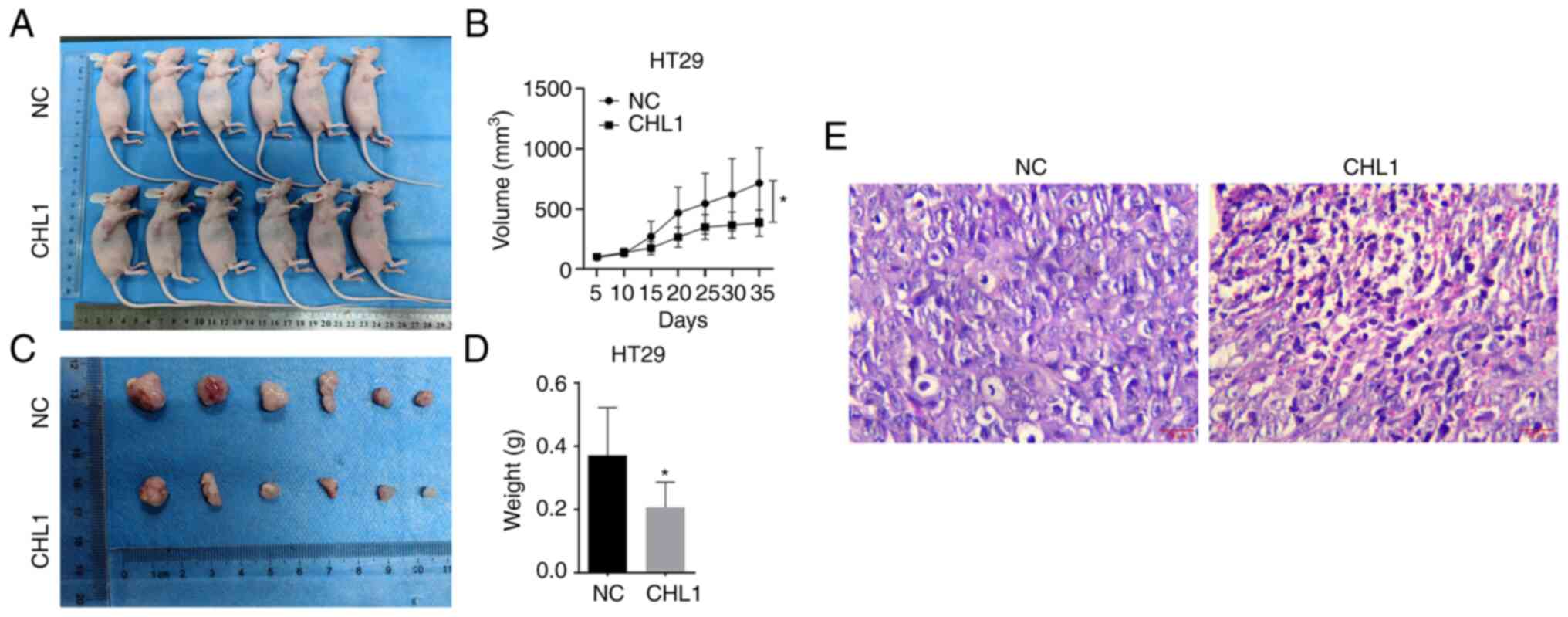
CHL1 overexpression suppresses CRC
cell tumorigenesis in vivo
HT29 cells that stably expressed CHL1 or control
HT29 cells were inoculated into BALB/c mice to investigate whether
CHL1 expression affects the tumorigenicity of CRC cells. CHL1
overexpression significantly suppressed tumor volume and weight in
the HT29 cell-induced xenograft tumor model (Fig. 4A-D). Additionally, H&E staining
revealed the closely arranged cancer cells in the NC group and the
sparsely arranged cancer cells in the group overexpressing CHL1
(Fig. 4E).
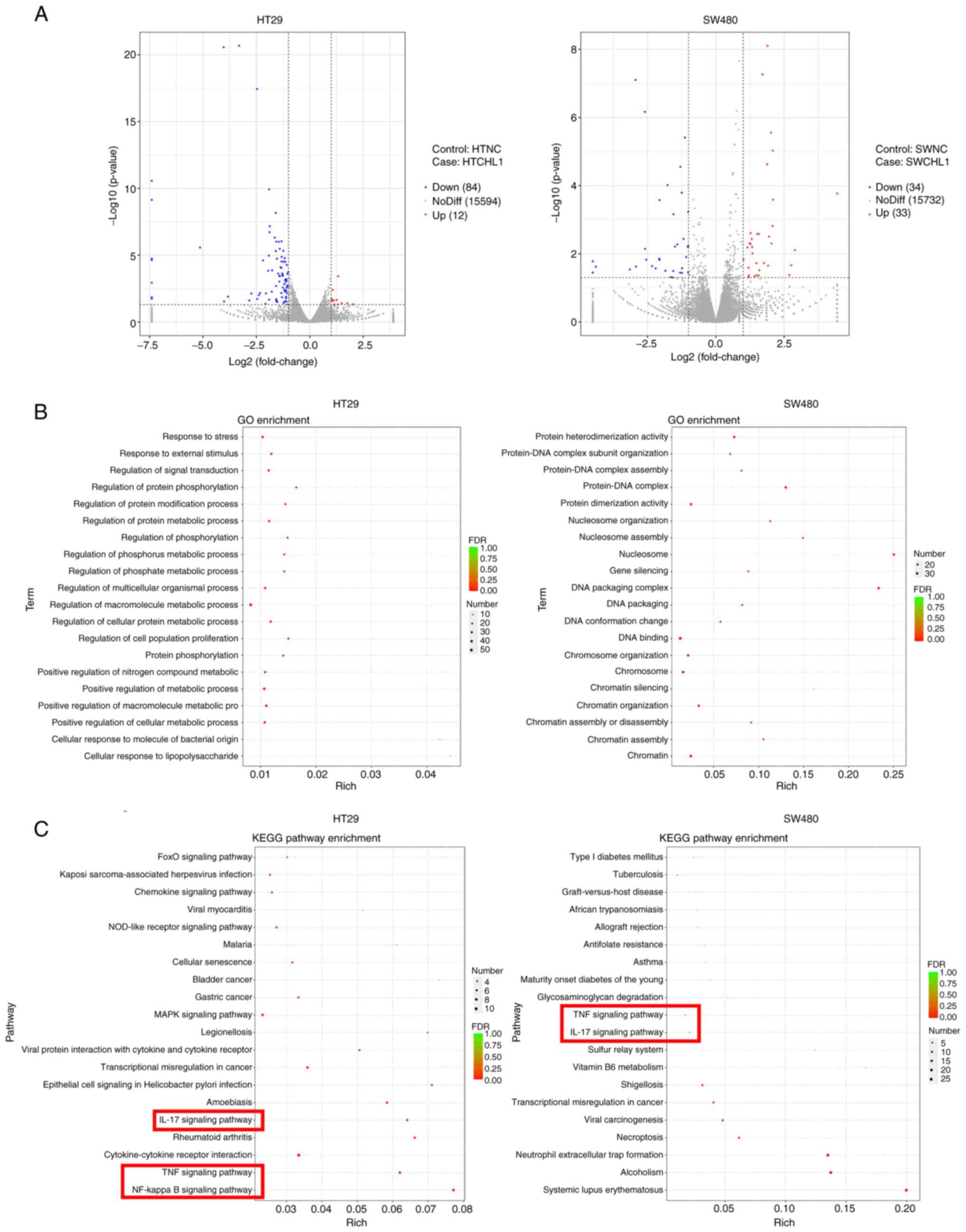
DEG screening
The downstream genes regulated by CHL1 were screened
using RNA-seq to further determine the mechanisms through which
CHL1 regulates the molecular mechanisms of CRC cell function. The
HT29 and SW480 cells were transfected with CHL1 overexpression
plasmids, followed by RNA-seq analysis. The results revealed that a
total of 96 DEGs were identified in the HT29-CHL1 group, of which
12 were upregulated and 84 were downregulated, in comparison with
the NC group. There were 67 DEGs identified in SW480-CHL1 cells,
including 33 upregulated and 34 downregulated DEGs. The cluster
volcano of the DEGs is presented in Fig. 5A. GO functional and KEGG analyses
were performed to further elucidate the biological functions of the
DEGs. The results of GO analysis (Fig.
5B) revealed that the DEGs were mainly enriched in the
regulation of macromolecule metabolic process, stress response and
signal transduction regulation in HT29-CHL1 cells. The DEGs were
mainly enriched in DNA binding, chromosome and protein dimerization
activity in the SW480 cells (Fig.
5B). KEGG analysis revealed that the DEGs of the HT29-CHL1
cells (Fig. 5C) were enriched in
the MAPK, cytokine-cytokine receptor interaction, NF-κB signaling
pathway, tumor necrosis factor (TNF) signaling pathway. The DEGs of
the SW480 cells were enriched in systemic lupus erythematosus,
alcoholism, neutrophil extracellular trap formation. The common
pathways were the interleukin (IL)-17 and TNF signaling pathways,
in which the downstream key protein was NF-κB (Fig. 5C).
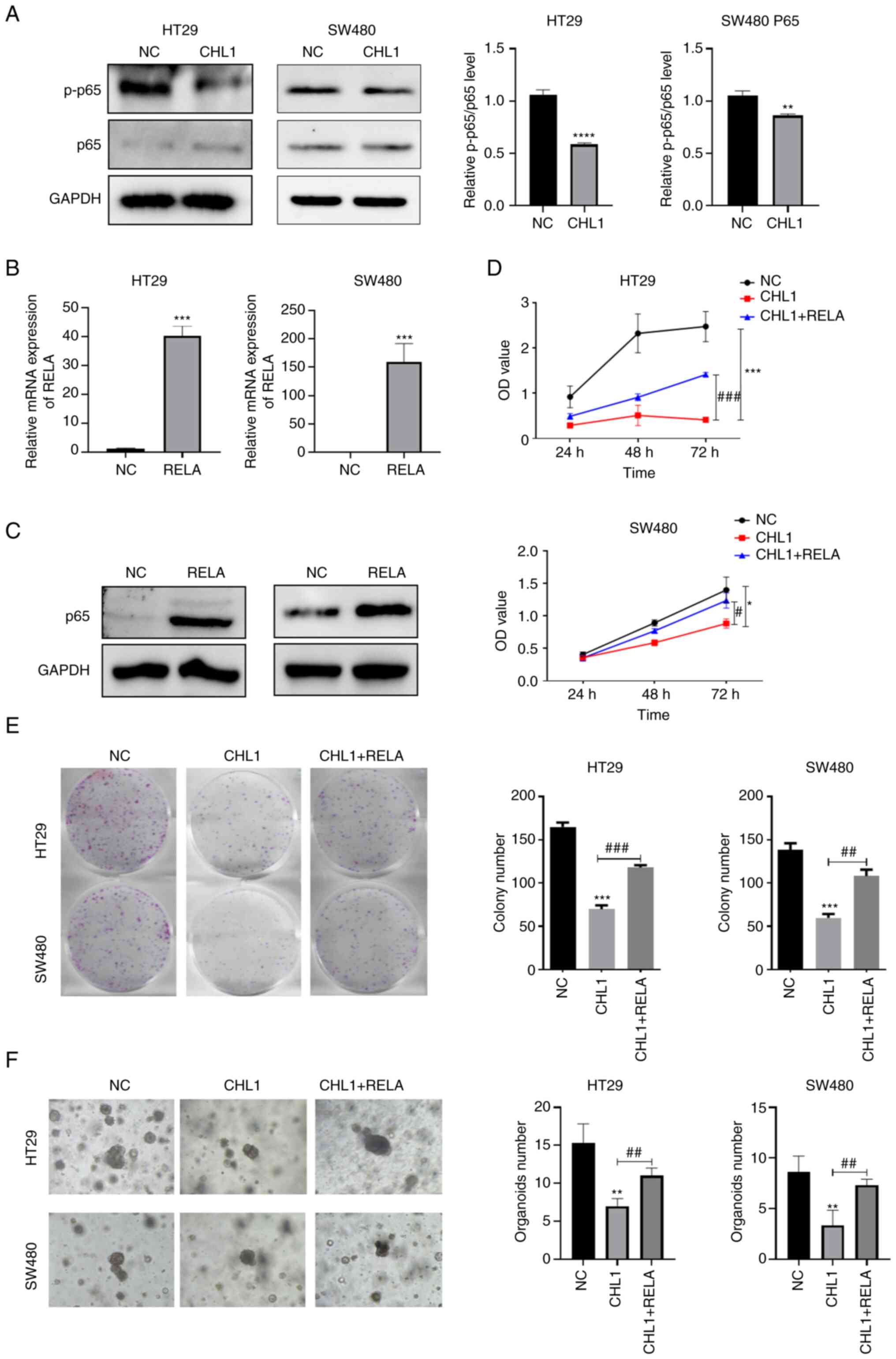
CHL1 inhibits CRC cell proliferation,
migration and invasion by regulating the NF-κB pathway
Functional enrichment analysis revealed a
significantly enriched NF-κB pathway in both the
CHL1-overexpressing HT29 and SW480 cells. Therefore, it was
hypothesized that CHL1 may play a key role in CRC cells by
regulating the NF-κB pathway. The results presented in Fig. 6A indicated a significantly
decreased expression of p-p65 and a p-p65/p65 ratio in
CHL1-overexpressing CRC cells. These results indicated that CHL1
overexpression inhibited NF-κB pathway activity. Subsequently, p65
expression was induced by transfection with pcDNA3.1-RELA
overexpression plasmid in CHL1-overexpressing CRC cells. The
results of RT-qPCR and western blotting revealed (Fig. 6B and C) significantly upregulated mRNA and
protein levels of p65 in the RELA group (P<0.05). Furthermore,
CCK-8, cell cloning and Transwell assays (Figs. 6D and E, and 7A
and B) revealed that p65
overexpression partially counteracted the inhibitory effects of
CHL1 overexpression on the proliferation, clone formation,
migration and invasion of HT29 and SW480 cells. Additionally, the
growth of HT29 and SW480 cell organoids was increased in the CHL1 +
RELA group, as compared with the CHL1 group (Fig. 6F). Additionally, the IF experiments
revealed a higher positive protein expression of E-cadherin in the
CHL1 + RELA group than in the CHL1 group (Fig. 7C).
Discussion
Cancer manifestation occurs due to changes in
genome, providing tumor cells with a selective advantage over
normal cells. These changes cause cancer cell development by
altering cell phenotypes, including proliferation and invasion
(22,23). CHL1 expression has been reported to
be downregulated in breast cancer cells, inhibiting cancer cell
proliferation and invasion (24).
Furthermore, CHL1 deletion promotes esophageal squamous cell
carcinoma cell proliferation and metastasis (25). At present, it is known that CHL1 is
abnormally expressed in CRC (15);
however, the specific regulatory mechanisms involved in CRC
tumorigenesis remain unknown. The present study revealed that CHL1
overexpression significantly inhibited the proliferative and colony
formation ability of CRC cells. Furthermore, CHL1 overexpression
significantly inhibited the organoid-formation ability of CRC
cells. Additionally, in vivo xenograft tumor experiments
revealed that CHL1 overexpression significantly reduced the HT-29
cell-induced tumor growth rate. These results indicated that CHL1
overexpression inhibited CRC cell proliferation and tumorigenic
capacity in vitro and in vivo, and that CHL1
functioned as a tumor suppressor in CRC.
E-cadherin is a key component of adherence junctions
and is essential for cell adhesion and cellular epithelial
phenotype maintenance (26). Tumor
progression has often been associated with the loss of E-cadherin
function and a transition to a more motile and aggressive phenotype
(27). E-cadherin is considered as
a tumor suppressor gene in colon tumorigenesis (28). N-cadherin is an
epithelial-mesenchymal transition (EMT)-related gene, and Wang
et al (29) confirmed that
cinobufacini inhibited CRC progression by promoting the expression
of the EMT-related protein, E-cadherin, while inhibiting the
expression of the EMT-related protein, N-cadherin. Similarly, the
results of the present study revealed a significantly upregulated
E-cadherin expression and downregulated N-cadherin expression in
HT29 and SW480 cells, following the induction of CHL1
overexpression. Moreover, Transwell assay revealed that CHL1
overexpression significantly suppressed CRC cell migration and
invasion.
RNA-seq analysis of CHL1-overexpressing HT29 and
SW480 cells was performed to further determine the molecular
mechanisms by which CHL1 regulates CRC cell function. Recently,
RNA-seq technology has appeared as a powerful method for analyzing
colorectal cancer transcriptomes (30). Zhai et al (31) identified 434 DEGs in colon cancer
and normal tissue samples using RNA-seq, providing molecular
markers for colon cancer metastasis and prognosis. The present
study identified 96 DEGs in CHL1-overexpressing HT29 cells and 67
DEGs in SW480-CHL1 cells, as compared with the NC group.
Furthermore, another notable finding that was obtained through GO
and KEGG analyses, was that the IL-17 and TNF signaling pathway, in
which the downstream key protein was NF-κB, were significantly
enriched both in CHL1-overexpressing HT29 and SW480 cells. Studies
have revealed that NF-κB plays a crucial role in tumor
proliferation, migration and invasion, which has become a hot spot
target in tumor diagnosis and therapy studies (32,33).
The NF-κB transcription factor has five members in mammals, of
which p65 is the most abundantly expressed in cells (34). TNF-α activates NF-κB signaling by
translocating the p65 DNA-binding factor to the nucleus, which in
turn regulates the transcription of various genes that are involved
in cell survival, invasion, and metastasis (35,36).
It has been reported that CRC metastasis and invasion are closely
related to the NF-κB signaling pathway (37). Wang et al (38) reported that PHD2 exerts an
anti-cancer effect by attenuating NF-κB activity in colon cancer
xenograft mice. However, no research has reported on NF-κB
signaling pathway regulation by CHL1 in CRC. The present study
revealed significantly reduced p65 phosphorylation and p-p65/p65
ratio, as well as inactivated NF-κB signaling pathway following
CHL1 overexpression. Previously, Crocin has been reported to
inhibit angiogenesis and CRC cell metastasis by blocking the
TNF-α/NF-κB/VEGF pathway (39).
This was consistent with the findings of the present study. The
results of the present study confirmed that CHL1 regulated the
TNF-α/NF-κB signaling pathway in CRC.
The inactivation NF-κB has inhibits the growth of
cells in various types of cancer, including chronic myelogenous
leukemia, breast cancer and CRC (40). Therefore, the present study further
determined whether CHL1 regulates the cellular function of CRC
through the NF-κB signaling pathway. The inhibitory effects of CHL1
overexpression on HT29 and SW480 cell proliferation, colony
formation, organoid growth, migration and invasion were reversed
following the upregulation of p65 expression. Additionally, the
promotion of E-cadherin expression was restrained. The
aforementioned results indicate that CHL1 inhibits CRC cell
proliferation, migration and invasion by regulating the NF-κB
signaling pathway.
HCT116 and SW620 cells have been reported to present
with greater metastatic potential in comparison with SW480
(41,42). However, according to the results of
the present study, it was initially observed that the expression of
CHL1 was relatively reduced in HT29 and SW480, and not in HCT116
and SW620 cells. Also, no significant difference in the expression
of CHL1 in SW480 and SW620 cells was observed. Therefore, the HT29
and SW480 cell lines were selected for inducing CHL1 overexpression
herein, in order to explore its effects and mechanisms. It was
speculated that in addition to CHL1 expression levels, the cell
migratory ability was also regulated by other possible regulatory
mechanisms, which warrant further investigations in order to
explore these in HCT116 and SW620 cells.
However, the present study has with certain
limitations. Clinical samples for the verification of the impact of
CHL1 expression alteration on CRC clinicopathology and prognosis
were not evaluated. Additionally, the specific effects of CHL1
knockdown and the mechanisms through which it is involved in
malignant CRC progression require further investigation.
Experiments involving animals and organoids are necessary for the
confirmation of the effects of CHL1 expression on CRC tumor
metastasis in vivo and in vitro. Moreover, related
functional and mechanistic analyses of additional DEGs were not
performed. In the future, the authors aim to conduct additional
in-depth studies on related targets including IL-17 and TNF
signaling pathways. The transient introduction of NF-κB regulates
other pathways to influence cell proliferation; this thus requires
further investigation.
In conclusion, the present study demonstrates that
CHL1 overexpression inhibits CRC cell proliferation, migration and
invasion, being also associated with the activation of the NF-κB
signaling pathway. Furthermore, the present study may provide a
novel molecular target for CRC clinical diagnosis and therapy.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the President
Foundation of Nanfang Hospital, Southern Medical University (grant
no. 2020B0170), the Guangdong Provincial Key Laboratory of
Precision Medicine for Gastrointestinal Cancer (grant no.
2020B121201004) and the Guangdong Provincial Major Talents Project
(grant no. 2019JC05Y361).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request. The RNA sequencing data were uploaded to Bioproject
(Bioproject no. PRJNA987357; https://www.ncbi.nlm.nih.gov/bioproject/PRJNA987357/).
Authors' contributions
MB and FJ contributed to the writing and preparation
of the original draft. MB, SL, YZ, XD and RW contributed to the
design of the methodology, data validation and formal analysis. RW
contributed to the visualization of the experimental results. RW,
FJ and SL contributed to data curation, and confirm the
authenticity of all the raw data. All authors contributed to the
conceptualization of the study, and critically revised the original
manuscript. All the authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
The experiments of the present study were approved
by the committee of Guangzhou Forevergen Biosciences Animal Center
(approval no. IACUC-AEWC-F2023021920). All experiments were
performed in accordance with the IACUC handbook (Third Edition) and
reported in accordance with ARRIVE guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Baidoun F, Elshiwy K, Elkeraie Y, Merjaneh
Z, Khoudari G, Sarmini MT, Gad M, Al-Husseini M and Saad A:
Colorectal cancer epidemiology: Recent trends and impact on
outcomes. Curr Drug Targets. 22:998–1009. 2021.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Biller LH and Schrag D: Diagnosis and
treatment of metastatic colorectal cancer: A review. JAMA.
325:669–685. 2021.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Coppedè F, Lopomo A, Spisni R and Migliore
L: Genetic and epigenetic biomarkers for diagnosis, prognosis and
treatment of colorectal cancer. World J Gastroenterol. 20:943–956.
2014.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Kanth P and Inadomi JM: Screening and
prevention of colorectal cancer. BMJ. 374(n1855)2021.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Zygulska AL and Pierzchalski P: Novel
diagnostic biomarkers in colorectal cancer. Int J Mol Sci.
23(852)2022.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Pidíkova P, Reis R and Herichova I: miRNA
clusters with down-regulated expression in human colorectal cancer
and their regulation. Int J Mol Sci. 21(4633)2020.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Holm J, Hillenbrand R, Steuber V, Bartsch
U, Moos M, Lübbert H, Montag D and Schachner M: Structural features
of a close homologue of L1 (CHL1) in the mouse: A new member of the
L1 family of neural recognition molecules. Eur J Neurosci.
8:1613–1629. 1996.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Tam GW, van de Lagemaat LN, Redon R,
Strathdee KE, Croning MD, Malloy MP, Muir WJ, Pickard BS, Deary IJ,
Blackwood DH, et al: Confirmed rare copy number variants implicate
novel genes in schizophrenia. Biochem Soc Trans. 38:445–451.
2010.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Senchenko VN, Krasnov GS, Dmitriev AA,
Kudryavtseva AV, Anedchenko EA, Braga EA, Pronina IV, Kondratieva
TT, Ivanov SV, Zabarovsky ER and Lerman MI: Differential expression
of CHL1 gene during development of major human cancers. PLoS One.
6(e15612)2011.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Cai X, Hu B, Liu S, Liu M, Huang Y, Lei P,
Zhang Z, He Z, Zhang L and Huang R: Overexpression of close homolog
of L1 enhances the chemosensitivity of lung cancer cells via
inhibition of the Akt pathway. Oncol Lett. 20(111)2020.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Chen J, Jiang C, Fu L, Zhu CL, Xiang YQ,
Jiang LX, Chen Q, Liu WM, Chen JN, Zhang LY, et al: CHL1 suppresses
tumor growth and metastasis in nasopharyngeal carcinoma by
repressing PI3K/AKT signaling pathway via interaction with Integrin
β1 and Merlin. Int J Biol Sci. 15:1802–1815. 2019.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Ognibene M, Pagnan G, Marimpietri D,
Cangelosi D, Cilli M, Benedetti MC, Boldrini R, Garaventa A,
Frassoni F, Eva A, et al: CHL1 gene acts as a tumor suppressor in
human neuroblastoma. Oncotarget. 9:25903–25921. 2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Qin M, Gao X, Luo W, Ou K, Lu H, Liu H and
Zhuang Q: Expression of CHL1 in clear cell renal cell carcinoma and
its association with prognosis. Appl Immunohistochem Mol Morphol.
30:209–214. 2022.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Yang Z, Xie Q, Hu CL, Jiang Q, Shen HF,
Schachner M and Zhao WJ: CHL1 is expressed and functions as a
malignancy promoter in glioma cells. Front Mol Neurosci.
10(324)2017.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Yu W, Zhu K, Wang Y, Yu H and Guo J:
Overexpression of miR-21-5p promotes proliferation and invasion of
colon adenocarcinoma cells through targeting CHL1. Mol Med.
24(36)2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Wang Z, Wang Q, Xu G, Meng N, Huang X,
Jiang Z, Chen C, Zhang Y, Chen J, Li A, et al: The long noncoding
RNA CRAL reverses cisplatin resistance via the miR-505/CYLD/AKT
axis in human gastric cancer cells. RNA Biol. 17:1576–1589.
2020.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Franken NA, Rodermond HM, Stap J, Haveman
J and van Bree C: Clonogenic assay of cells in vitro. Nat Protoc.
1:2315–2319. 2006.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Hu X, Wang P, Qu C, Zhang H and Li L:
Circular RNA Circ_0000677 promotes cell proliferation by regulating
microRNA-106b-5p/CCND1 in non-small cell lung cancer.
Bioengineered. 12:6229–6239. 2021.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43(e47)2015.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Dennis G Jr, Sherman BT, Hosack DA, Yang
J, Gao W, Lane HC and Lempicki RA: DAVID: Database for annotation,
visualization, and integrated discovery. Genome Biol.
4(P3)2003.PubMed/NCBI
|
|
22
|
Fearon ER: Molecular genetics of
colorectal cancer. Annu Rev Pathol. 6:479–507. 2011.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Jafarzadeh M and Soltani BM: MiRNA-Wnt
signaling regulatory network in colorectal cancer. J Biochem Mol
Toxicol. 35(e22883)2021.PubMed/NCBI View Article : Google Scholar
|
|
24
|
He LH, Ma Q, Shi YH, Ge J, Zhao HM, Li SF
and Tong ZS: CHL1 is involved in human breast tumorigenesis and
progression. Biochem Biophys Res Commun. 438:433–438.
2013.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Tang H, Jiang L, Zhu C, Liu R, Wu Y, Yan
Q, Liu M, Jia Y, Chen J, Qin Y, et al: Loss of cell adhesion
molecule L1 like promotes tumor growth and metastasis in esophageal
squamous cell carcinoma. Oncogene. 38:3119–3133. 2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Mendonsa AM, Na TY and Gumbiner BM:
E-cadherin in contact inhibition and cancer. Oncogene.
37:4769–4780. 2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Canel M, Serrels A, Frame MC and Brunton
VG: E-cadherin-integrin crosstalk in cancer invasion and
metastasis. J Cell Sci. 126:393–401. 2013.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Daulagala AC, Bridges MC and Kourtidis A:
E-cadherin beyond structure: A signaling hub in colon homeostasis
and disease. Int J Mol Sci. 20(2756)2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Wang J, Cai H, Liu Q, Xia Y, Xing L, Zuo
Q, Zhang Y, Chen C, Xu K, Yin P and Chen T: Cinobufacini inhibits
colon cancer invasion and metastasis via suppressing Wnt/β-catenin
signaling pathway and EMT. Am J Chin Med. 48:703–718.
2020.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Yamada A, Yu P, Lin W, Okugawa Y, Boland
CR and Goel A: A RNA-sequencing approach for the identification of
novel long non-coding RNA biomarkers in colorectal cancer. Sci Rep.
8(575)2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Zhai X, Xue Q, Liu Q, Guo Y and Chen Z:
Colon cancer recurrence-associated genes revealed by WGCNA
co-expression network analysis. Mol Med Rep. 16:6499–6505.
2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Wu Y and Zhou BP:
TNF-alpha/NF-kappaB/Snail pathway in cancer cell migration and
invasion. Br J Cancer. 102:639–644. 2010.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Yan T, Tan Y, Deng G, Sun Z, Liu B, Wang
Y, Yuan F, Sun Q, Hu P, Gao L, et al: TGF-β induces GBM mesenchymal
transition through upregulation of CLDN4 and nuclear translocation
to activate TNF-α/NF-κB signal pathway. Cell Death Dis.
13(339)2022.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Karin M, Cao Y, Greten FR and Li ZW:
NF-kappaB in cancer: From innocent bystander to major culprit. Nat
Rev Cancer. 2:301–310. 2002.PubMed/NCBI View
Article : Google Scholar
|
|
35
|
Inoue J, Gohda J, Akiyama T and Semba K:
NF-kappaB activation in development and progression of cancer.
Cancer Sci. 98:268–274. 2007.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Pozniak PD, White MK and Khalili K:
TNF-α/NF-κB signaling in the CNS: Possible connection to EPHB2. J
Neuroimmune Pharmacol. 9:133–141. 2014.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Seo GS: The role of NF-kappaB in colon
cancer. Korean J Gastroenterol. 57:3–7. 2011.PubMed/NCBI View Article : Google Scholar : (In Korean).
|
|
38
|
Wang L, Niu Z, Wang X, Li Z, Liu Y, Luo F
and Yan X: PHD2 exerts anti-cancer and anti-inflammatory effects in
colon cancer xenografts mice via attenuating NF-κB activity. Life
Sci. 242(117167)2020.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Bakshi HA, Quinn GA, Nasef MM, Mishra V,
Aljabali AAA, El-Tanani M, Serrano-Aroca Á, Webba Da Silva M,
McCarron PA and Tambuwala MM: Crocin inhibits angiogenesis and
metastasis in colon cancer via TNF-α/NF-kB/VEGF pathways. Cells.
11(1502)2022.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Haefner B: NF-kappa B: Arresting a major
culprit in cancer. Drug Discov Today. 7:653–663. 2002.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Nairon KG, DePalma TJ, Zent JM, Leight JL
and Skardal A: Tumor cell-conditioned media drives collagen
remodeling via fibroblast and pericyte activation in an in vitro
premetastatic niche model. iScience. 25(104645)2022.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Subauste MC, Kupriyanova TA, Conn EM, Ardi
VC, Quigley JP and Deryugina EI: Evaluation of metastatic and
angiogenic potentials of human colon carcinoma cells in chick
embryo model systems. Clin Exp Metastasis. 26:1033–1047.
2009.PubMed/NCBI View Article : Google Scholar
|