1. Introduction
Oxidative stress is caused by the disruption of the
balance between oxidative and antioxidant responses in the body.
When exposed to noxious stimuli, the body is in a state of
oxidative stress, and the level of free radicals is elevated,
leading to apoptosis and tissues damage (1). Inflammation is the fundamental
response of the body to injury and infection. However, if
inflammation is not controlled, various proinflammatory factors
(Interleukins, Tumor Necrosis Factor, etc.) released by persistent
inflammation can induce further pathological responses (2). Previous research shows that oxidative
stress and inflammation are associated with the occurrence and
development of a number of diseases such as diabetes,
cardiovascular disease and cancer (3-5).
Therefore, studies have been attempting to reveal additional
effective drugs with antioxidant and anti-inflammatory effects. As
an important source of drugs, natural products serve a key role in
the identification and development of new drugs (6). Therefore, finding new antioxidant and
anti-inflammatory drugs from natural products may become an
important research direction in the future. An overview of the
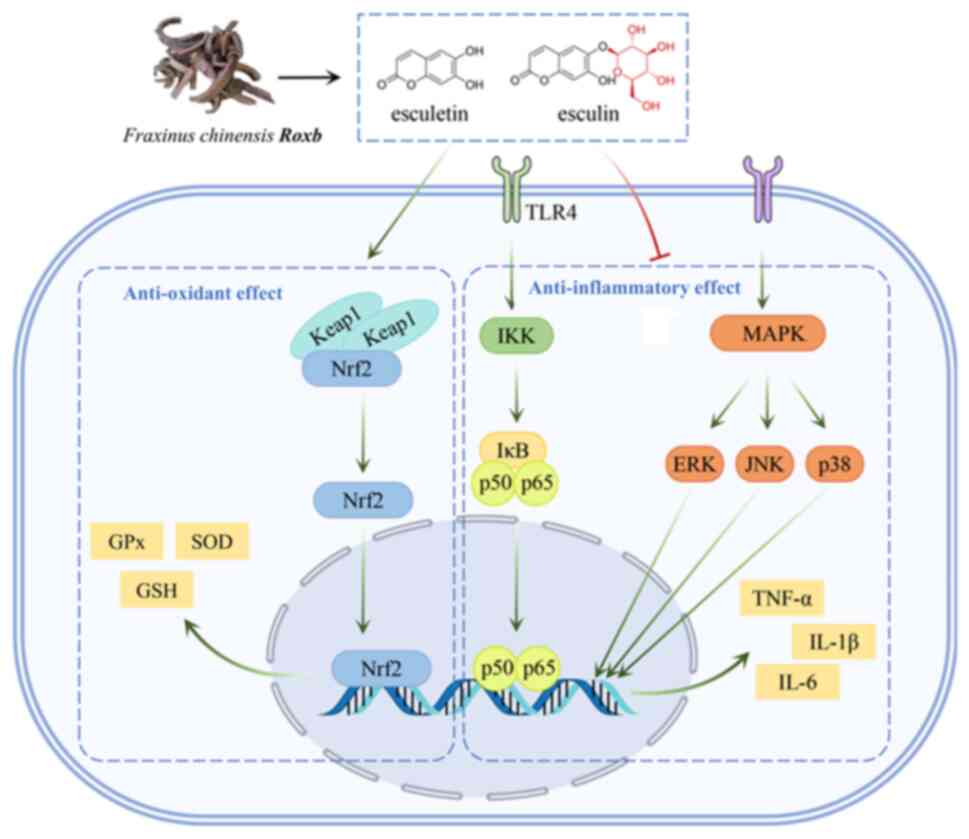
antioxidant and anti-inflammatory effects of Fraxinus chinensis
Roxb in Fig. 1.
Fraxinus chinensis Roxb is a deciduous tree,
which is distributed worldwide and has important medicinal value.
In Asia, the bark of Fraxinus is a commonly used traditional
Chinese medicine called Qinpi (7).
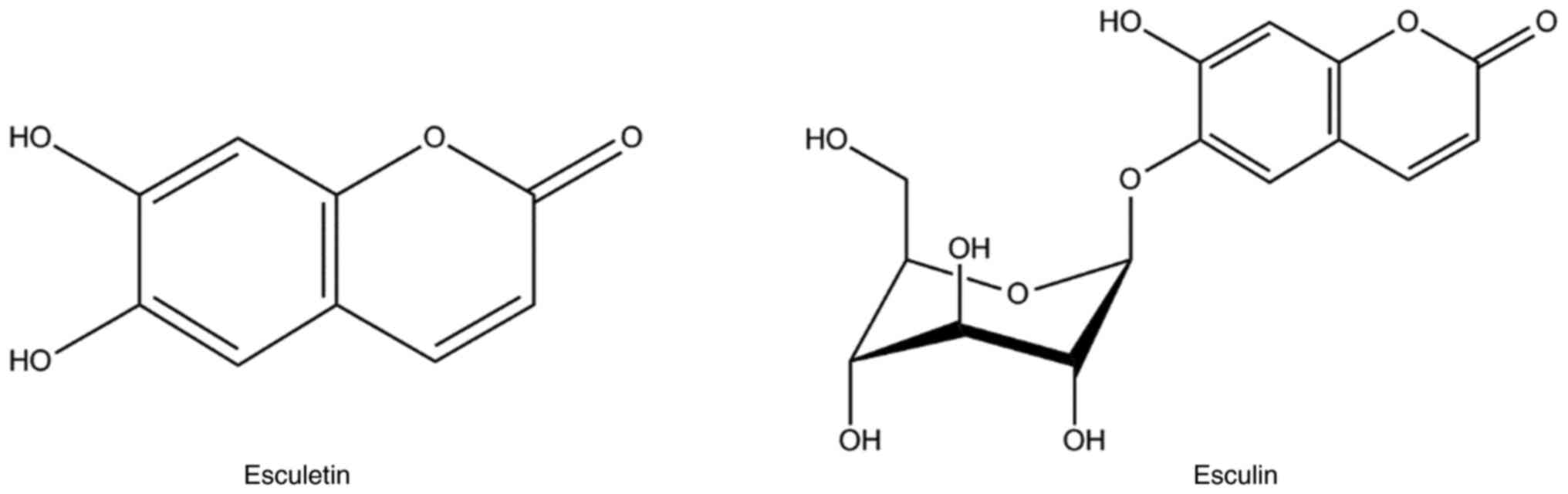
The bark of Fraxinus contains 1.0~1.4%, with esculetin and
its glycoside form, esculin, being the most studied active
ingredients (Fig. 2) (8). The present study shows that esculetin
and esculin have a wide range of pharmacological activities,
including antioxidant, anti-inflammatory, antibacterial, antitumor,
antidiabetic, anti-atherosclerotic, and immunomodulatory effects.
And esculetin and esculin are expected to be used in the treatment
of various diseases, such as cancer, diabetes and neurodegenerative
diseases. Further studies have shown that esculetin and esculin
have prominent antioxidant and anti-inflammatory effects, which may
underlie their broad pharmacological activities (9,10).
Antioxidant effects of esculetin and esculin are associated with
the activation of the nuclear factor erythroid-derived factor
2-related factor 2 (Nrf2) signaling pathway (11). As a key signaling pathway for
antioxidation, the Nrf2 signaling pathway can regulate the
expression levels of various endogenous antioxidant proteins, such
as superoxide dismutase (SOD), glutathione peroxidase (GPx) and
glutathione (GSH) (11,12). The anti-inflammatory effects of
esculetin and esculin are associated with the inhibition of nuclear
factor κ-B (NF-κB) and mitogen-activated protein kinase (MAPK), two
important inflammatory signaling pathways. In various inflammatory
models, esculetin and esculin can reduce the expression levels of a
number of proinflammatory factors such as tumor necrosis factor-α
(TNF-α), interleukin (IL)-1β and IL-6, thereby inhibiting the
development of inflammation (13,14).
In summary, as natural products, esculin and esculetin may be
promising candidates for the potential use as complementary and
alternative medicines. The present study aimed to systematically
review studies that investigated the anti-inflammatory and
antioxidant properties of esculetin and esculin, as well as to
provide current and useful information.
2. Antioxidant activities of esculin and
esculetin
It is considered that the oxidative and antioxidant
reactions of the normal body are in a state of dynamic equilibrium.
Oxidative stress is caused by the breakdown of this balance and the
accumulation of oxidative substances such as reactive oxygen
species (ROS) or reactive nitrogen species such as
O2•−, NO•, ONOO− and
OH• (15). A study
confirmed that esculin and esculetin effectively scavenge free
radicals (16). In addition,
experimental results show that esculin and esculetin can improve
various pathological processes that are associated with oxidative
stress. This is associated with the activation of the Nrf2
signaling pathway and the promotion of the expression of various
antioxidant proteins such as GSH and SOD (15,16).
Antioxidant activities of esculetin.
Free radical scavenging effects of esculetin
At present, a number of studies have determined the
scavenging effect of esculetin on 2,2-Diphenyl-1-picrylhydrazyl
(DPPH), but there are differences between the results. Wang et
al (16) find that the
clearance rates of DPPH with different concentrations of esculetin
(0.02, 0.05, 0.08 and 0.1 mg/ml) are 28.13, 59.94, 79.68 and 100%,
respectively. The SC50 values of esculetin for
scavenging DPPH as determined by Jeong et al (17) and Kim et al (18) are both 14.68 µM. The
SC50 value of esculetin for scavenging DPPH as
determined by Vianna et al (19) is 25.18 µM, but is determined by Lee
et al (20) to be 40 µM. In
addition to scavenging DPPH, esculetin also has a scavenging effect
on hydroxyl radicals with an SC50 value of 0.091 mg/ml (16). Hsia et al (21) also find that esculetin (50-80 µM)
inhibits hydroxyl radical formation in washed human platelets. In
addition, Lee et al (22)
demonstrated that esculetin has a scavenging effect on superoxide
anion radicals with an SC50 value of 0.6 µg/ml. In addition,
esculetin inhibits peroxidase-induced oxidation in a
non-competitive manner with a Ki value of 9.5 µM in the
methemalbumin-H2O2-tetramethylbenzidine pseudoperoxidase system
(23). By scavenging the free
radicals, esculetin (10-100 mg/ml) protects human dermal
fibroblasts from UVB-induced oxidative stress damage (22). Furthermore, by scavenging the
O2-accumulation caused by H2O2, esculetin (100 µM)
protects human leukemia NB4 cells from oxidative stress damage
(24,25).
Regulated antioxidant protein action
of esculetin
Studies using cells further confirm the antioxidant
effect of esculetin, which regulates the expression of various
antioxidant proteins (26-28).
For example, in L-buthionine-sulfoximine-induced mouse cortical
cells, esculetin (10-100 µM) increases GPx, glutathione reductase
(GR) and GSH activities and inhibits oxidized glutathione
(glutathione disulfide; GSSG) accumulation (26). In addition, esculetin (10 µM) can
inhibit the accumulation of ROS in Helicobacter pylori
urease-induced human vascular endothelial cells by promoting the
expression of heme oxygenase-1 (HO-1) (27). Additionally, esculetin (10-20 µM)
can increase the levels of aspartate aminotransferase (AST),
alanine aminotransferase (ALT), SOD and GSH and inhibit the
accumulation of ROS and malondialdehyde (MDA) in tert-butyl
hydroperoxide (t-BHP)-induced HepG2 cells (28,29).
Further studies show that the antioxidant activity
of esculetin is associated with the regulation of multiple
signaling pathways, such as inhibiting the NF-κB and MAPK signaling
pathways (13,14,30).
The Nrf2 signaling pathway is the primary defense mechanism for
inhibiting oxidative stress, and it regulates the expression of a
number of important antioxidant proteins such as HO-1, SOD and GPx.
A large number of studies show that the antioxidant effect of
esculetin is associated with the activation of the Nrf2 signaling
pathway (15,30,31).
Sen et al (30) find that
esculetin (10-20 µM) reduces the production of ROS in the high
glucose-induced human renal tubular epithelial HK-2 cells and that
the Nrf2 signaling pathway is activated. Furthermore, esculetin
(20-500 µM) can induce SOD expression in the leukemia NB4 cells by
activating the Nrf2 signaling pathway (31).
Scavenging ROS effects of
esculetin
Accumulation of ROS can activate the mitochondrial
apoptotic pathway and induce apoptosis, and esculetin can block
this process by scavenging ROS (32). In zinc oxide nanoparticles-induced
human neuroblastoma SH-SY5Y cells, esculetin (10-30 µM) can
decrease GPx and GSH depletion by inhibiting lipoxygenase
expression, which ameliorates the ROS accumulation-induced
apoptosis (33). Esculetin (0.1-10
µM) can also protect Aβ25-35-induced human neuroblastoma SH-SY5Y
cells from apoptosis caused by ROS accumulation by increasing SOD,
GSH and catalase (CAT) expression levels, which reduces caspase-3,
cytochrome c and Bax expression levels and increases Bcl-2
expression levels (34,35). Additionally, esculetin (5-20 µM)
increases SOD levels and decreases ROS levels and thus decreases
lactate dehydrogenase (LDH), Bax and cleaved caspase-3 levels in
hypoxia/reoxygenation-induced myocardial H9c2 cells (36).
In H2O2-induced human corneal epithelial cells,
esculetin (20-100 µM) activates the Nrf2 signaling pathway,
inducing the expression of multiple antioxidant genes, including
HO-1, quinone 1 (NQO1), glutamate cysteine ligase modifier subunit
(GCLM), SOD1 and SOD2, thereby inhibiting ROS generation and
accumulation (37). By activating
the Nrf2 pathway and promoting the expression of HO-1, SOD, GPx and
CAT, esculetin (12.5-100 µM) inhibits the production and
accumulation of ROS in 3T3-L1 preadipocytes (38). Furthermore, in H2O2-induced Caco2
cells, esculetin (100-300 µg/ml) activates the Nrf2 pathway and
increases the expression of SOD, CAT and GPx (39). Esculetin (10-25 µM) also protects
human hepatoma HepG2 cells from hydrogen peroxide-induced oxidative
damage by activating the Nrf2 signaling pathway and promoting the
expression of NQO1 and glutathione (40). Additionally, esculetin (10-100 µM)
can restore depleted GSH levels and inhibit the production of ROS
and MDA in ethanol-induced HepG2 cells by activating the Nrf2
signaling pathway (41).
Amelioration of oxidative stress damage by
esculetin. The results of mouse studies further show that
esculetin can improve oxidative stress and associated injuries,
such as nervous system injury, liver injury and kidney injury: For
example, esculetin can improve nervous system injury caused by
oxidative stress (42-45).
Esculetin (0.5% w/w) also ameliorates
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced neurotoxicity
in the substantia nigra pars compacta in mice by preventing
nitrosative stress, increasing GSH levels, inhibiting the
activation of caspase 3 and preventing neuronal apoptosis (42). Additionally, esculetin (20-80
mg/kg) inhibits tBCCAO-induced activation of the mitochondrial
apoptosis pathway and autophagy in mice by reducing the expression
levels of autophagy-related factors (such as Bnip3, Beclin1, Pink1
and parkin) and apoptosis-related factors (such as p53, Bax and
caspase 3) by activating the Nrf2 signaling pathway (43). In addition, esculetin (25 mg/kg)
decreases immobility time and climbing time in the forced swim task
elicited in acute restraint stress-induced rats by restoring the
activities of antioxidant enzymes including CAT, SOD, GPx and GSH,
and reducing the levels of GSSG in rat cortical areas (44). Another study shows that esculetin
can inhibit oxidative stress in liver injury (45). In CCl4-induced liver injury rats,
esculetin (35 mg/kg) increases SOD and CAT levels and decreases MDA
levels (45). Furthermore, in
N-nitrosodiethylamine-induced liver injury rats, esculetin (0.5%
w/w) ameliorates hepatic lipid peroxidation by restoring the total
glutathione levels and increasing the levels of NQO1, HO-1 and
glutathione S-transferase Pi (GSTP1) (46). Esculetin can also inhibit oxidative
stress in kidney injury. In high-fat diet-induced kidney injury
mice, esculetin (40 mg/kg) increases the expression levels of SOD,
CAT, GPx, glutathione-S-transferase (GST), non-enzymic antioxidants
vitamin C and GSH, and reduces the levels of thiobarbituric acid
reactive substances (TBARS), lipid hydroperoxides and co-conjugated
diene (47).
Other antioxidant oxidative effects of
esculetin
In addition, He et al (36) find that esculetin (5-20 µM)
protects hypoxia/reoxygenation-induced myocardial H9c2 cells from
oxidative stress-induced apoptosis. This protective effect is
produced by activating the Janus kinase/signal transducer and
activator of transcription (JAK2/STAT3) pathway (36). Notably, studies show that
activation of the Nrf2 pathway is associated with the extracellular
regulated protein kinases (ERK) signaling pathway. Esculetin (5 µM)
can activate the Nrf2 signaling pathway and promote the expression
of NQO1 by activating the ERK signaling pathway in H2O2-induced
C2C12 myoblasts (48,49).
The results of animal studies further show that
esculetin can improve the oxidative stress caused by
ischemia-reperfusion. For example, esculetin (20-40 mg/kg) restores
SOD activity and reduces the levels of MDA in myocardial
ischemia-reperfusion rats (50).
In addition, esculetin can improve the oxidative stress in
testicular tissue damage. Esculetin (50 mg/kg) ameliorates
AlCl3-induced rat testicular tissue damage by activating the Nrf-2
signaling pathway, increasing the activities of GSH, GPx and CAT
and preventing the activation of the apoptotic pathway (51).
In conclusion, the antioxidant effect of esculetin
stems from increasing the expression levels of endogenous
antioxidants such as NQO1, HO-1, GSH and SOD by activating the Nrf2
signaling pathway, which enables the free radicals generated by the
body to be scavenged in time to avoid lipid oxidation and
apoptosis.
Antioxidant activities of esculin.
Free radical scavenging effects of esculin
Free radical scavenging experiments show that
esculin has scavenging effects on a variety of free radicals
including O2•-, NO• and DPPH, and the SC50 values are 69.27 µg/ml,
8.56 µg/ml and 0.141 µM, respectively (52,53).
Similar to the antioxidant mechanism of esculetin, the activation
of the Nrf2 signaling pathway is also associated with the
antioxidant effect of esculin. Studies demonstrate that esculin
(50-200 µmol/l) can disrupt the interaction of the Kelch-like
ECH-associated protein 1 (Keap1), a key negative regulator of the
Nrf2 signaling pathway, with Nrf2 to activate the Nrf2 signaling
pathway (54,55).
Regulated antioxidant protein action
of esculin
Esculin has significant antioxidant modulating
effects and ameliorates oxidative stress in kidney injury. Esculin
(150 mg/kg) promotes the expression of SOD, GR and CAT in
prooxidant aflatoxin B-1-induced nephrotoxicity mice (56). Similarly, esculin can inhibit the
oxidative stress in pancreatic injury (57). Esculin (10-50 mg/kg) increases the
expression levels of GPx, SOD and CAT in streptozotocin
(STZ)-induced type 2 diabetic mice (57).
Scavenging ROS effects of esculin
The scavenging ROS effects of esculin have been
widely reported. For example, esculin (1-100 µM) protects
dopamine-induced human neuroblastoma SH-SY5Y cells from ROS
accumulation-induced apoptosis by increasing SOD and GSH expression
levels and inhibiting the expression of cytochrome c,
apoptosis-inducing factor, p53, Bax and caspase 3(58). Myeloperoxidase is able to mediate
oxidative stress by promoting ROS production and promoting
inflammation-related signaling pathways (59). In human neuroblastoma SH-SY5Y
cells, esculin reduces the dopamine-induced ROS overproduction,
scavenges ROS and enhances the activities of SOD and GSH (58).
Amelioration of oxidative stress
damage by esculin
Experiments using cells further confirm the
antioxidant effect of esculin. For example, esculin (20-100 µM) can
protect linoleic acid hydroperoxide-induced human umbilical vein
endothelial cells from free radical-induced oxidative stress damage
(60).
The results of experiments using mice further show
that esculin can decrease oxidative stress and the associated
damage. Esculin can inhibit oxidative stress in liver injury, as
esculin (10-40 mg/kg) can activate the Nrf2 signaling pathway to
increase HO-1 expression levels and decrease the MDA content in
lipopolysaccharide (LPS)/D-galactosamine-induced acute liver injury
mice (61). Another study
demonstrates that the esculin can improve oxidative stress in
gastric injury, as in ethanol-induced gastric mucosal injury rats,
esculin (12.5-50 mg/kg) increases the SOD levels and decreases the
MDA levels (52).
Other antioxidant oxidative effects of
esculin
In addition, esculin can decrease oxidative stress
in colitis. For example, esculin (5-25 mg/kg) promotes the
expression of GPx and partially ameliorates the intestinal injury
in rats with 2,4,6-trinitrobenzenesulfonic acid (TNBS)-induced
colitis (53). Furthermore,
studies associated with acute lung injury (62) and diabetic nephropathy (63) demonstrate the antioxidant activity
of esculin. Esculin can improve oxidative stress in arthritis and,
in adjuvant-induced arthritis rats, esculin (10-40 mg/kg) promotes
the expression of GPx (64).
In conclusion, esculin, as the glycoside form of
esculetin, has similar antioxidant activities to esculetin. By
activating the Nrf2 signaling pathway and promoting the expression
of various endogenous antioxidants such as HO-1, GSH and SOD,
esculin serves a protective role in the process of various tissue
injuries.
3. Anti-inflammatory activities of esculetin
and esculin
Inflammation is the basic response of the body to
injury and infection. However, if inflammation is not controlled,
persistent inflammatory responses can induce new pathological
responses (such as tissue damage, fibrosis, metabolic disorders,
cardiovascular disease, etc.) and lead to the development of
various diseases, such as cancer, metabolic diseases,
neurodegenerative diseases and vascular diseases (65). Previous research shows that
esculetin and esculin can inhibit the expression of proinflammatory
factors, such as TNF-α, IL-1β and IL-6 in various inflammatory
models (13,14,61).
This is associated with the inhibition of inflammatory signaling
pathways, such as NF-κB and MAPK.
Anti-inflammatory activities of
esculetin
Studies using cells show that esculetin can inhibit
the expression of various inflammatory markers (66). For example, esculetin can inhibit
the production of nitric oxide (NO) in IL-1β stimulated rat
hepatocytes with an IC50 value of 34 µM (66). Additionally, esculetin (10-100
µg/ml) inhibits IL-6 production in TNF-α stimulated human
osteosarcoma MG-63 cells (67).
Esculetin (1-10 µg/ml) can also inhibit the release of inflammatory
mediators, such as NO, TNF-α, IL-1β, IL-6 and monocyte
chemoattractant protein-1 (MCP-1) in LPS-induced RAW 264.7 cells
(68-71).
Further studies demonstrate that the anti-inflammatory effect of
esculetin is associated with inhibiting the NF-κB signaling pathway
(72-77).
Sun et al (72) demonstrate
that esculetin (10-40 µM) inhibits the histamine-induced production
of IL-6, IL-8 and mucin-5AC in human nasal epithelial cells, which
is partly mediated by inhibition of the NF-κB signaling pathway
(72). Esculetin (12 µg/ml) also
inhibits the expression of inducible isoform of NO synthase (iNOS),
cyclooxygenase 2 (COX-2), TNF-α and IL-1β in LPS-induced RAW 264.7
cells by inhibiting the NF-κB signaling pathway (73). Additionally, esculetin (12.5-25
µg/ml) inhibits the activation of the NF-κB signaling pathway in
TNF-α induced vascular smooth muscle cells, which reduces the
expression levels of mitochondrial membrane potential-9 (MMP-9) and
the binding activity of activating protein-1 (AP-1) (74,75).
Furthermore, esculetin (30-200 µM) reduces the IL-1β, COX-2 and
TNF-α levels in trimethyltin-induced human neuroblastoma SH-SY5Y
cells by inhibiting the NF-κB signaling pathway (76). In addition to inhibiting the NF-κB
signaling pathway, esculetin can also decrease the inflammatory
response by inhibiting the MAPK signaling pathway. In LPS-induced
human retinal pigment epithelium cells, esculetin (0.78-50 µM)
decreases the expression levels of IL-1β, IL-12, TNF receptor and
TNF-related apoptosis-inducing ligand by inhibiting the ERK1/2 and
NF-κB signaling pathways (77). In
addition, lipoxygenases are a family of non-heme iron-containing
enzymes and are involved in the synthesis of inflammatory cytokines
such as leukotrienes. A previous study demonstrated that the
inhibition rate of esculetin on soybean 15-lipoxygenase at a
concentration of 28.1 µM is 40.50% (78). Esculetin can also inhibit the
activity of 5-lipoxygenase with an IC50 value of 6.6 µM (79).
The results of study with mice further indicate that
esculetin can exhibit an anti-inflammatory effect in various
inflammatory models (61,63,74).
This suggests that esculetin may be promising for the treatment of
a variety of inflammation-related diseases, such as ulcerative
colitis, chronic kidney disease, cardiovascular disease.
Lung inflammation
A study confirms that esculetin can improve the
inflammatory response in lung injury (80). Esculetin (20-40 mg/kg) reduce the
histopathological changes (such as alveolar wall thickening,
interstitial edema, and pulmonary congestion) and the infiltration
of inflammatory cells, and inhibits the production of TNF-α, IL-1β,
IL-6, IL-23 and myeloperoxidase (MPO) in LPS-induced acute lung
injury mice. This is associated with the inhibition of the
activation of the AKT/ERK/NF-κB and retinoid-related orphan nuclear
receptor γt (RORγt)/IL-17 signaling pathways (80,81).
In polyhexamethylene guanidine-induced pulmonary fibrosis mice,
esculetin (10 mg/kg) antagonizes macrophage infiltration,
ameliorates alveolar epithelial barrier disruption and reduces the
levels of MMP-9, macrophage inflammatory protein 2, IL-8 and IL-1β
(82).
Esculetin decreases the inflammatory response in
asthma. For example, esculetin (20 mg/kg) reduces the levels of
IL-4, IL-5, IL-13, IL-17A, immunoglobulin E (IgE), GATA binding
protein 3 and RORγt in the lung tissue of ovalbumin-induced asthma
mice (83). In addition, esculetin
(10 mg/kg) improves airway inflammation, and inhibits airway
eosinophilia, collagen fiber deposition and goblet cell hyperplasia
in mice exposed to particles <10 µm. This is associated with the
inhibition of the activation of the toll-like receptor 4 (TLR4)
signaling pathway and the expression of COX-2 and nitric oxide
synthase 2(84).
Inflammation of the gastrointestinal
tract
Esculetin can reduce the inflammatory response in
gastrointestinal injury. For example, esculetin (100 mg/kg)
decreases the levels of COX-2, iNOS and cytokine-induced neutrophil
chemoattractant-3 in TNBS-induced colitis rats (85). Furthermore, esculetin (5 mg/kg) can
inhibit the production of NO, TNF-α, IL-6, iNOS and
nucleotide-binding oligomerization domain, leucine-rich repeat and
pyrin domain-containing in DSS-induced colitis rats by inhibiting
the activation of the NF-κB and MAPK signaling pathways (86,87).
Additionally, esculetin (20-40 mg/kg) inhibits the expression of
iNOS, TNF-α and IL-6 in ethanol-induced gastric injury rats by
inhibiting the activation of the NF-κB signaling pathway (88).
Inflammation in liver injury
Esculetin decreases the inflammatory response in
liver injury. For example, esculetin (0.01%, w/w) can downregulate
the expression of inflammatory genes including TLR4, myeloid
differentiation primary response gene 88 (MyD88), NF-κB, TNF-α and
IL-6, and decrease the levels of blood glycated hemoglobin, TNF-α,
IL-6 and MCP-1 in high-fat diet-induced fatty liver mice (89).
Neuroinflammation
Esculetin can reduce neuroinflammation. For example,
esculetin (5-25 mg/kg) can inhibit the zinc oxide
nanoparticles-induced expression of IL-1β and TNF-α in rat brain
tissue (90). Furthermore,
esculetin (20-40 mg/kg) ameliorates LPS-induced neuroinflammation
and depression-like behavior in mice by reducing the levels of
IL-6, IL-1β, TNF-α, iNOS and COX-2 in serum and the hippocampus by
inhibiting the activation of the NF-κB signaling pathway (91,92).
Skin-related inflammation
Esculetin decreases skin-related inflammation. In
house dust mite (Dermatophagoides farinae extract) and
2,4-dinitrochlorobenzene-induced dermatitis mice, esculetin (2-50
mg/kg) reduces serum IgE, IgG2a and histamine levels, reduce the
infiltration of inflammatory cells, and inhibits the production of
TNF-α, interferon-γ (IFN-γ), IL-4, IL-13, IL-17 and IL-31 by
inhibiting the STAT1 and NF-κB signaling pathways (93). Furthermore, in imiquimod-induced
psoriasis-like mouse, esculetin (50-100 mg/kg) was able to
significantly reduce the mRNA levels of pro-inflammatory cytokines
(including IL-6, IL-17A, IL-22, IL-23, TNF-α and IFN-γ) in the skin
of mice, and inhibited the phosphorylation of IKK alpha and P65 in
the skin of psoriasis, consequently, inhibited the NF-KB signaling
pathway (94).
Other inflammation
Esculetin can improve the inflammatory response in
other pathological processes. For example, esculetin (20-60 mg/kg)
can inhibit the production of inflammatory factors such as IL-1β,
IL-6, TNF-α, C-C motif chemokine ligand 2 and iNOS in
Escherichia coli-induced sepsis mice by inhibiting the NF-κB
and STAT1/STAT3 signaling pathways (95). In a rabbit model in which the
primary lacrimal gland, sclerotial gland, and cornea were removed
to induce dry eye, esculetin (0.05% of the diet) was able to
inhibit the expression of IL-1α, IL-1β, and TNF-α by inhibiting the
ERK1/2 signaling pathway and thereby suppressing IL-1α, IL-1β, and
TNF-α (96). Furthermore,
esculetin (10-20 mg/kg) inhibits the expression of TNF-α and IL-6
in isoproterenol induced myocardial toxicity in rats by inhibiting
the activation of the NF-κB signaling pathway (97). Additionally, esculetin (2.5-10
mg/kg) can reduce reserpine-induced fibromyalgia in mice, which is
associated with the inhibition of monoamine oxidase-A activity, and
downregulated the levels of IL-1β, TNF-α and TBARS (98). In summary, the aforementioned
studies show that esculetin can reduce the expression levels of
various proinflammatory factors and inhibit the infiltration of
inflammatory cells in a number of inflammatory models. In addition
to inhibiting the two most common inflammatory signaling pathways,
NF-κB and MAPK, the anti-inflammatory effect of esculetin is also
associated with inhibiting the STAT signaling pathway.
Anti-inflammatory activity of
esculin
Cellular research shows that esculin (25-200 µM)
reduces the levels of TNF-α, IL-1β and IL-6 in free fatty
acid-induced HepG2 cells (99).
Further studies show that the anti-inflammatory effect of esculin
is associated with the inhibition of the MAPK and NF-κB signaling
pathways. For example, esculin (300-500 µM) can reduce the
expression levels of IL-1β, TNF-α and iNOS in LPS-stimulated
RAW264.7 cells by inhibiting the NF-κB signaling pathway (100). In LPS-stimulated RAW264.7 cells,
esculin (10-50 µM) also inhibits the p38/MAPK signaling pathway and
downregulates the MMP-9 level and AP-1 binding activity (101). Additionally, esculin (1 µg-1 mg)
can inhibit the p38 MAPK, c-Jun N-terminal kinase (JNK) and ERK1/2
signaling pathways and reduce the TNF-α and IL-6 expression levels
in mouse peritoneal macrophages (102).
Studies using animals further indicates that esculin
can exhibit an anti-inflammatory effect in various inflammatory
models (95,96,99,100). This suggests that esculin may be
promising for the treatment of a variety of inflammation-related
diseases (including ulcerative colitis, chronic kidney
cardiovascular disease).
Lung inflammation
Esculin can reduce lung injury-related inflammation.
For example, esculin (10-50 mg/kg) decreases the levels of NO and
TNF-α in hyperoxia-induced lung injury rats (61). Additionally, esculin (20-40 mg/kg)
can inhibit the TLR4/MyD88/NF-κB signaling pathway and downregulate
the expression of TNF-α, IL-1β and IL-6 in LPS-induced acute lung
injury mice (103).
Inflammation of the gastrointestinal
tract
Esculin can reduce the inflammatory response in
gastrointestinal injury. For example, esculin (5-50 mg/kg) inhibits
the activation of the NF-κB signaling pathway and downregulates the
levels of IL-1β and TNF-α in DSS-induced colitis mice (100). In ethanol-induced gastric injury
mice, esculin (5-20 mg/kg) also inhibits the activation of the
NF-κB signaling pathway and downregulates the production of NO,
TNF-α and IL-6(104).
Inflammation in liver injury
Esculin can reduce the inflammatory response in
liver injury. For example, esculin (10-40 mg/kg) inhibits the NF-κB
signaling pathway and reduces the levels of TNF-α and IL-1β in
LPS/D-galactosamine-induced acute liver injury mice (62). In methionine choline-deficient
diet-induced non-alcoholic steatohepatitis mice, esculin (20-40
mg/kg) also inhibits the NF-κB signaling pathway, downregulates the
expression of TNF-α and IL-6, and increases the expression level of
the silent information regulator 1(99).
Neuroinflammation
Esculin can improve nervous system inflammation. For
example, esculin (50 mg/kg) inhibits the expression of IL-12, IL-6
and TNF-α in the hippocampus of reserpine-induced depression mice
(105). Furthermore, esculin
(20-40 mg/kg) inhibits the activation of the NF-κB signaling
pathway and the expression of IL-6 and TNF-α in LPS-induced
depressed mice hippocampus, which increases the number of
selectively activated microglia and decreases the number of
classically activated microglia (106). In addition, esculin (30-90 mg/kg)
can inhibit the MAPK signaling pathway and reduce the levels of
TNF-α, IL-6, intracellular adhesion molecule-1 (ICAM-1) and MCP-1
to improve memory impairment in STZ-induced diabetic rats (63).
Kidney inflammation
Esculin can improve the inflammatory response in
renal injury. For example, esculin (25-100 mg/kg) inhibits the
TLR4/MyD88 signaling pathway and downregulates the expression of
IL-1β, IL-6, TNF-α, MCP-1 and ICAM-1 in LPS-induced acute kidney
injury mice (107). Similarly,
esculin (30-90 mg/kg) decreases the expression levels of IL-1β,
IL-6, ICAM-1 and NO in the kidney tissue of STZ-induced diabetic
rats (108).
Other inflammation
Esculin can also improve the inflammatory response
in other pathological processes. For example, esculin (10-40 mg/kg)
significantly improved body weight, decreased paw volume, and the
levels of IL-1β, TNF-α, and IL-6 levels in adjuvant-induced
arthritis rats compared to arthritic controls (64). Additionally, esculin (5-20 mg/kg)
can ameliorate xylene-induced rat paw swelling by inhibiting the
activation of the p38, MAPK, JNK and ERK1/2 signaling pathways and
decreasing TNF-α and IL-6 levels (102). Furthermore, in LPS-induced sepsis
mice, esculin (30 mg/kg) inhibits the NF-κB signaling pathway and
reduces the levels of TNF-α and IL-6(109). In conclusion, esculin (the
glycoside form of esculetin) has anti-inflammatory effects and the
mechanisms of action are similar to esculetin, inhibiting the
activation of the NF-κB and MAPK signaling pathways and inhibiting
the expression of various proinflammatory factors.
4. Conclusion and future perspectives
In conclusion, esculetin and esculin have been
extensively studied, and studies utilizing cells and/or animals
show that both possess a variety of pharmacological activities,
including anti-inflammatory, antioxidant, anti-tumor, antidiabetic,
antibacterial, antiviral, anti-atherosclerotic and immunomodulatory
pharmacological effects (110).
The antioxidant effects of esculetin and esculin have been shown in
a variety of diseases, their underlying mechanisms of action and
biological activities include scavenging of free radicals,
promotion of the expression of endogenous antioxidant proteins
(such as SOD, GPx, and GSH), modulating the nuclear factor
erythroid 2-related factor 2 pathway, regulating the cell cycle,
inhibiting tumor cell proliferation and migration, etc.. Further
studies reveal that this is associated with the activation of the
Nrf2 signaling pathway (Fig. 3).
The anti-inflammatory effects of esculetin and esculin are
associated with the inhibition of two important inflammatory
signaling pathways, NF-κB and MAPK. In various models of
inflammation, including diabetes, liver injury, tumors, bacterial,
fungal, neurological, lung and respiratory inflammation, both
esculetin and esculin are able to reduce the expression of various
proinflammatory factors, such as TNF-α, IL-1β and IL-6, thereby
inhibiting the tissues damage caused by inflammation (Fig. 3, Table
I and II). In conclusion,
esculetin and esculin both have antioxidant and anti-inflammatory
effects, and may be used for the treatment or improvement of
various diseases in the future.
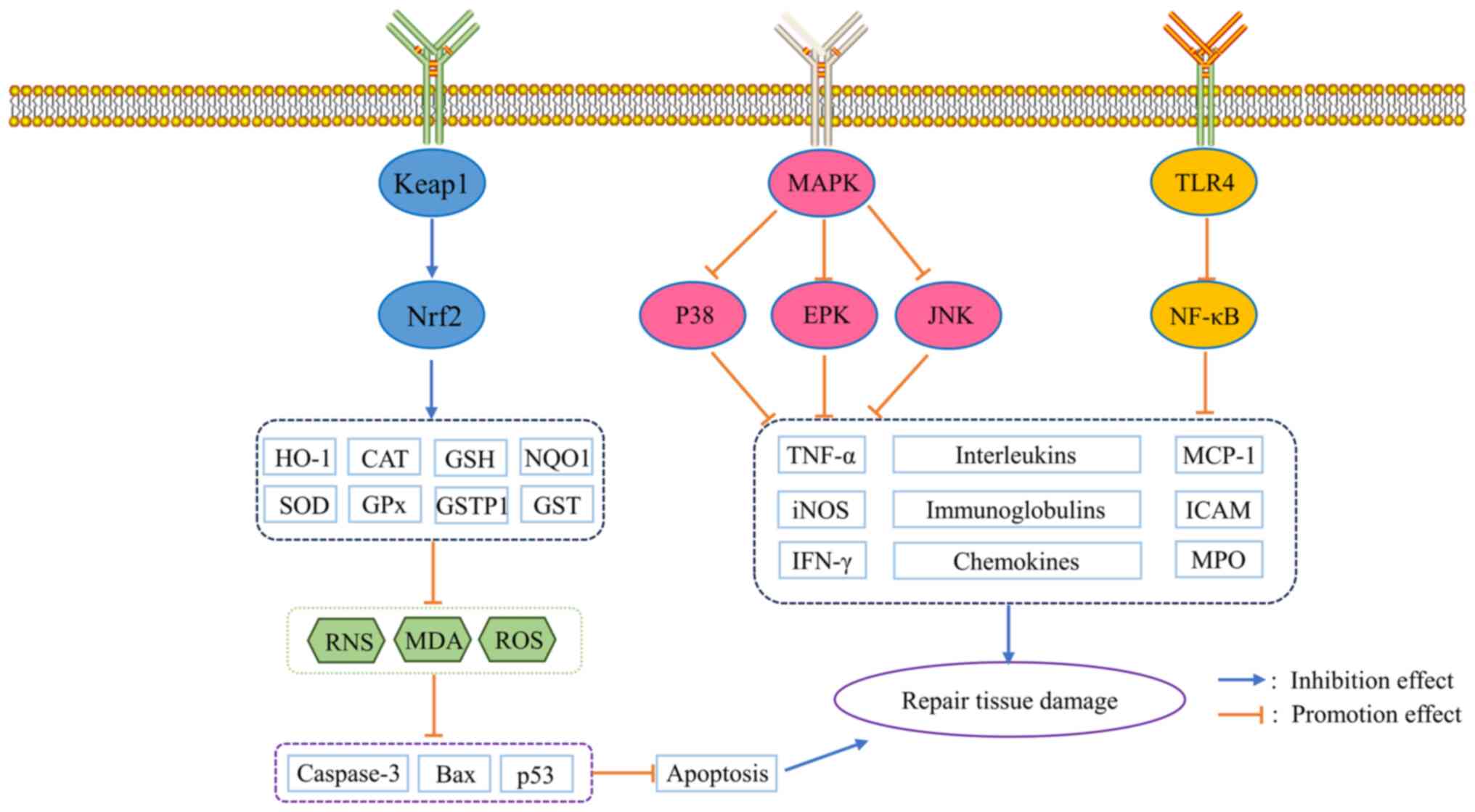 | Figure 3Schematic diagram of the antioxidant
and anti-inflammatory mechanisms of esculetin and esculin. TLR4,
toll-like receptor 4; Keap1, Kelch-like ECH-associated protein 1;
Nrf2, nuclear factor erythroid 2-related factor 2; HO-1, heme
oxygenase-1; CAT, chloramphenicol acetyltransferase; GSH,
glutathione; NQO1, NADPH quinone oxidoreductase; SOD, superoxide
dismutase; GPx, glutathione peroxidase; GSTP1, glutathione
S-transferase Pi; GST, glutathione-S-transferase; RNS, reactive
nitrogen species; MDA, malondialdehyde; ROS, reactive oxygen
species; ERK, extracellular regulated protein kinase; JNK, c-Jun
N-terminal kinase; MCP-1, monocyte chemoattractant protein-1; iNOS,
inducible isoform of nitric oxide synthase; ICAM, intracellular
adhesion molecule; MPO, myeloperoxidase. |
 | Table IAntioxidant and anti-inflammatory
effects of esculetin. |
Table I
Antioxidant and anti-inflammatory
effects of esculetin.
| A, Antioxidant |
|---|
| Detail | Cell
lines/model | Dosage | (Refs.) |
|---|
| Increase GPx, GR
and GSH activities and inhibit GSSG accumulation |
L-buthionine-sulfoximine-induced mouse
cortical cells | 10-100 µM | (26) |
| Promote the
expression of HO-1 | Helicobacter
pylori urease-induced human vascular endothelial cells | 10 µM | (27) |
| Increase the levels
of AST, ALT, SOD and glutathione and inhibit the accumulation of
ROS and MDA | t-BHP-induced HepG2
cells | 10-20 µM | (28,29) |
| Decrease GPx and
GSH depletion by inhibiting lipoxygenase expression | Zinc oxide
nanoparticles-induced human neuroblastoma SH-SY5Y cells | 10-30 µM | (33) |
| Increase SOD, GSH
and CAT expression levels, which inhibit activation of
mitochondrial apoptosis pathway |
Aβ25-35-induced human
neuroblastoma SH-SY5Y cells | 0.1-10 µM | (34,35) |
| Increase SOD levels
and decrease ROS levels and thus, inhibit activation of
mitochondrial apoptosis pathway |
Hypoxia/reoxygenation-induced myocardial
H9c2 cells | 5-20 µM | (36) |
| Induce SOD
expression levels by activating the Nrf2 signaling pathway | Leukemia NB4
cells | 20-500 µM | (31) |
| Activate the Nrf2
signaling pathway and induce the expression levels of multiple
antioxidant genes, including HO-1, NQO1, GCLM, SOD1 and SOD2 |
H2O2-induced human
corneal epithelial cells | 20-100 µM | (37) |
| Activate the Nrf2
pathway and promote the expression of HO-1, SOD, GPx and CAT | 3T3-L1
preadipocytes | 12.5-100 µM | (38). |
| Activate the Nrf2
pathway and increase the expression levels of SOD, CAT and GPx |
H2O2-induced Caco2
cells | 100-300 µg/ml | (39) |
| Activate Nrf2
signaling pathway and promote the expression of NQO1 and
glutathione | Hydrogen
peroxide-induced human hepatoma HepG2 cells | 10-25 µM | (40) |
| Restore the
depleted GSH level and inhibit the production of ROS and MDA by
activating the Nrf2 signaling pathway | Ethanol-induced
HepG2 cells | 10-100 µM | (41) |
| Activate the Nrf2
signaling pathway and promote expression of NQO1 by activating the
ERK signaling pathway |
H2O2-induced C2C12
myoblasts | 5 µM | (48,49) |
| Activate JAK2/STAT3
pathway |
Hypoxia/reoxygenation-induced myocardial
H9c2 cells | 5-20 µM | (36) |
| Prevent nitrosative
stress, increase GSH levels and inhibit the activation of caspase
3 |
1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced
neurotoxicity in mice | 0.5% w/w | (42) |
| Reduce the
expression levels of autophagy-related factors (such as Bnip3,
Beclin1, Pink1 and parkin) and apoptosis-related factors (such as
p53, Bax and caspase 3) by activating the Nrf2 signaling
pathway | tBCCAO-induced
mice | 20-80 mg/kg | (43) |
| Decrease immobility
time and climbing time by restoring the antioxidant enzyme
activities of CAT, SOD, GPx and GSH, and reducing GSSG levels | Forced swim task
elicited in acute restraint stress-induced rats | 25 mg/kg | (44) |
| Increase SOD and
CAT levels, and decrease MDA levels |
CCl4-induced liver injury
rats | 35 mg/kg | (45) |
| Restore total
glutathione levels and increase the levels of NQO1, HO-1 and
GSTP1 |
N-nitrosodiethylamine-induced liver injury
rats | 0.5% w/w | (46) |
| Increase expression
levels of SOD, catalase, GPx, GST, non-enzymic antioxidants vitamin
C and GSH, and reduce levels of TBARS, lipid hydroperoxides and
co-conjugated diene | High-fat
diet-induced kidney injury mice | 40 mg/kg | (47) |
| Restore SOD
activity and reduce the amount of MDA | Myocardial
ischemia-reperfusion rats | 50 mg/kg | (50) |
| Activate the Nrf-2
signaling pathway, and increase the activities of GSH, GPx and
CAT |
AlCl3-induced rat testicular
tissue damage | 50 mg/kg | (51) |
| B,
Anti-inflammatory |
| Detail | Cell
lines/model | Dosage | (Refs.) |
| Inhibit the
production of NO | IL-1β stimulated
rat hepatocytes | IC50, 34
µM | (66) |
| Inhibit IL-6
production | TNF-α stimulated
human osteosarcoma MG-63 cells | 10-100 µg/ml | (67) |
| Inhibit the release
of NO, TNF-α, IL-1β, IL-6 and MCP-1 | LPS-induced RAW
264.7 cells | 1-10 µg/ml | (68-71) |
| Inhibit the
production of IL-6, IL-8 and mucin-5AC by inhibition of the NF-κB
signaling pathway | Histamine-induced
human nasal epithelial cells | 10-40 µM | (72) |
| Inhibit expression
of iNOS, COX-2, TNF-α and IL-1β by inhibiting the NF-κB signaling
pathway | LPS-induced RAW
264.7 cells | 12 µg/ml | (73) |
| Inhibit activation
of the NF-κB signaling pathway and reduce expression of MMP-9 and
the binding activity of AP-1 | TNF-α induced
vascular smooth muscle cells | 12.5-25 µg/ml | (74,75) |
| Reduce the IL-1β,
COX-2 and TNF-α levels by inhibiting the NF-κB signaling
pathway |
Trimethyltin-induced human neuroblastoma
SH-SY5Y cells | 30-200 µM | (76) |
| Decrease the
expression levels of IL-1β, IL-12, TNF receptor and TRAIL by
inhibiting the ERK1/2 and NF-κB signaling pathways | LPS-induced human
retinal pigment epithelium cells | 0.78-50 µM | (77) |
| Inhibit the
production of TNF-α, IL-1β, IL-6, IL-23, myeloperoxidase and MPO by
inhibiting the AKT/ERK/NF-κB and RORγt/IL-17 signaling
pathways | LPS-induced acute
lung injury mice | 20-40 mg/kg | (80,81) |
| Reduce the levels
of MMP-9, macrophage inflammatory protein 2, IL-8 and IL-1β | Polyhexamethylene
guanidine-induced pulmonary fibrosis mice | 10 mg/kg | (82) |
| Reduce the levels
of IL-4, IL-5, IL-13, IL-17A, IgE, GATA3 and RORγt | Ovalbumin-induced
asthma mice | 20 mg/kg | (83) |
| Inhibit the TLR4
signaling pathway and the expression levels of COX-2 and NOS2 | uPM10-exposed
mice | 10 mg/kg | (84) |
| Decrease the levels
of COX-2, iNOS and CINC-3 | TNBS-induced
colitis rats | 100 mg/kg | (85) |
| Inhibit the
production of NO, TNF-α, IL-6, iNOS and NLRP3 by inhibiting the
NF-κB and MAPK signaling pathways | DSS-induced colitis
rats | 5 mg/kg | (86,87) |
| Inhibit the
expression of iNOS, TNF-α and IL-6 by inhibiting the NF-κB
signaling pathway | Ethanol-induced
gastric injury rats | 20-40 mg/kg | (88) |
| Downregulate the
expression of Tlr4, Myd88, Nfkb, Tnfα,
Il6, blood HbA1c, TNF-α, IL-6 and MCP-1 | HFD-induced fatty
liver mice | 0.01% w/w | (89) |
| Inhibit the
expression of IL-1β and TNF-α | Zinc oxide
nanoparticles-induced rats | 5-25 mg/kg | (90) |
| Reduce the levels
of IL-6, IL-1β, TNF-α, iNOS and COX-2 by inhibiting the NF-κB
signaling pathway | LPS-induced
mice | 20-40 mg/kg | (91,92) |
| Reduce serum IgE,
IgG2a and histamine levels, and inhibit the production of TNF-α,
IFN-γ, IL-4, IL-13, IL-17 and IL-31 by inhibiting the STAT1 and
NF-κB signaling pathways | House dust mite
(dermatophagoides farinae extract) and
2,4-dinitrochlorobenzene-induced dermatitis mice | 2-50 mg/kg | (93) |
| Reduce the levels
of IL-6, IL-17A, IL-22, IL-23, TNF-α and IFN-γ by inhibiting the
NF-κB signaling pathway | Imiquimod-induced
psoriasis-like mice | 50-100 mg/kg | (94) |
| Inhibit the
expression of IL-1β, IL-6, TNF-α, CCL2 and iNOS by inhibiting the
NF-κB and STAT1/STAT3 signaling pathways | Escherichia
coli-induced sepsis mice | 20-60 mg/kg | (95) |
| Inhibit the
expression of IL-1α, IL-1β and TNF-α by inhibiting the ERK1/2
signaling pathway | Dry eye
rabbits | 0.05% of the
diet | (96) |
| Inhibit the
expression of TNF-α and IL-6 by inhibiting the NF-κB signaling
pathway | Isoproterenol
induced myocardial toxicity rats | 10-20 mg/kg | (97) |
| Inhibit monoamine
oxidase-A activity, and downregulate the IL-1β, TNF-α and TBARS
levels | Reserpine-induced
fibromyalgia mice | 2.5-10 mg/kg | (98) |
| Table IIAntioxidant and anti-inflammatory
effects of esculin. |
Table II
Antioxidant and anti-inflammatory
effects of esculin.
| A, Antioxidant |
|---|
| Detail | Cell
lines/model | Dosage | (Refs.) |
|---|
| Increase SOD and
GSH expression levels and inhibit the expression of cytochrome c,
apoptosis-inducing factor, p53, Bax and caspase 3 | Dopamine-induced
human neuroblastoma SH-SY5Y cells | 1-100 µM | (58) |
| Activate the Nrf2
signaling pathway to increase HO-1 expression levels and decrease
the MDA content |
LPS/D-galactosamine-induced acute liver
injury mice | 10-40 mg/kg | (61) |
| Increase SOD levels
and decrease MDA levels | Ethanol-induced
gastric mucosal injury rats | 12.5-50 mg/kg | (104) |
| Promote the
expression of GPx | Adjuvant-induced
arthritis rats | 10-40 mg/kg | (64) |
| Promote the
expression of GPx | TNBS-induced
colitis rats | 5-25 mg/kg | (53) |
| Promote the
expression of SOD, GR and catalase | Prooxidant
aflatoxin B-1-induced nephrotoxicity mice | 150 mg/kg | (56) |
| Increase the
expression levels of GPx, SOD and catalase | STZ-induced type 2
diabetic mice | 10-50 mg/kg | (57) |
| B,
Anti-inflammatory |
| Detail | Cell
lines/model | Dosage | (Refs.) |
| Reduce the levels
of TNF-α, IL-1β and IL-6 | Free fatty
acid-induced HepG2 cells | 25-200 µM | (99) |
| Reduce the
expression levels of IL-1β, TNF-α and iNOS by inhibiting the NF-κB
signaling pathway | LPS-stimulated
RAW264.7 cells | 300-500 µM | (100) |
| Inhibit p38/MAPK
signaling pathway and downregulate MMP-9 levels and AP-1 binding
activity | LPS-stimulated
RAW264.7 cells | 10-50 µM | (101) |
| Inhibit p38/MAPK,
JNK and ERK1/2 signaling pathways and reduce the TNF-α and IL-6
expression levels | Mouse peritoneal
macrophages | 1 µg-1 mg | (102) |
| Inhibit the
expression levels of IL-12, IL-6 and TNF-α | Reserpine-induced
depression mice | 50 mg/kg | (105) |
| Inhibit the
activation of the NF-κB signaling pathway and the expression of
IL-6 and TNF-α | LPS-induced
depressed mice | 20-40 mg/kg | (106) |
| Inhibit the MAPK
signaling pathway and reduce the levels of TNF-α, IL-6, ICAM-1 and
MCP-1 | STZ-induced
diabetic rats | 30-90 mg/kg | (63) |
| Decrease the levels
of NO and TNF-α | Hyperoxia-induced
lung injury rats | 10-50 mg/kg | (61) |
| Inhibit the
TLR4/MyD88/NF-κB signaling pathway and downregulate the expression
levels of TNF-α, IL-1β and IL-6 | LPS-induced acute
lung injury mice | 20-40 mg/kg | (103) |
| Inhibit the NF-κB
signaling pathway and downregulate the levels of IL-1β and
TNF-α | DSS-induced colitis
mice | 5-50 mg/kg | (100) |
| Inhibit the NF-κB
signaling pathway and downregulate the production of NO, TNF-α and
IL-6 | Ethanol-induced
gastric injury mice | 5-20 mg/kg | (104) |
| Inhibit the
TLR4/MyD88 signaling pathway and downregulate the expression of
IL-1β, IL-6, TNF-α, MCP-1 and ICAM-1 | LPS-induced acute
kidney injury mice | 25-100 mg/kg | (107) |
| Decrease the
expression levels of IL-1β, IL-6, ICAM-1 and NO | STZ-induced
diabetic rats | 30-90 mg/kg | (108) |
| Inhibit the NF-κB
signaling pathway and reduce the levels of TNF-α and IL-1β |
LPS/D-galactosamine-induced acute liver
injury mice | 10-40 mg/kg | (62) |
| Inhibit the NF-κB
signaling pathway, downregulate the expression of TNF-α and IL-6,
and increase the expression of the silent information regulator
1 | Non-alcoholic
steatohepatitis mice | 20-40 mg/kg | (99) |
| Decrease the levels
of IL-1β, TNF-α and IL-6 | Adjuvant-induced
arthritis rats | 10-40 mg/kg | (64) |
| Inhibit p38/MAPK,
JNK and ERK1/2 signaling pathways and decrease the TNF-α and IL-6
levels | Xylene-induced rat
paw swelling | 5-20 mg/kg | (102) |
| Inhibit the NF-κB
signaling pathway and reduce the levels of TNF-α and IL-6 | LPS-induced sepsis
mice | 30 mg/kg | (109) |
Currently, the most studied components in the
Chinese medicine Qinpi are esculetin and esculin, both of which
belong to the coumarin group of components. It's worth looking into
esculetin and esculin have relatively abundant experimental data
from animals (including mice, rats, rabbits, etc.) in the treatment
of a variety of diseases related to inflammation and oxidative
stress, and therefore, these two components have a better prospect
of clinical application. In the future research, the clinical study
can be carried out to determine the esculetin and esculin in the
clinical environment. antioxidant and anti-inflammatory activities.
The development of esculetin and esculin as novel drugs for the
treatment of inflammation and other associated diseases has the
potential to advance the development of natural drug formulations
for the treatment of inflammation.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the Natural Science
Foundation of Sichuan Province (grant no. 2023NSFSC1803),
Innovative topics of Affiliated Sport Hospital of CDSU (grant no.
LCCX22B01), and the Project of Sichuan Provincial Administration of
TCM (grant nos. 2023MS271, 2023MS268).
Availability of data and materials
Not applicable.
Authors' contributions
SJ conceived and revised the review. YT and QW
performed the literature. LZ and KW drafted the review article and
revised the manuscript. CW and MW reviewed and helped to modify the
manuscript. Data authentication is not applicable. All authors have
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen Z and Zhong C: Oxidative stress in
Alzheimer's disease. Neurosci Bull. 30:271–281. 2014.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Müller N, Myint AM and Schwarz MJ:
Inflammation in schizophrenia. Adv Protein Chem Struct Biol.
88:49–68. 2012.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Liu J, Li Y, Sun C, Liu S, Yan Y, Pan H,
Fan M, Xue L, Nie C, Zhang H, et al: Geniposide reduces cholesterol
accumulation and increases its excretion by regulating the
FXR-mediated liver-gut crosstalk of bile acids. Pharmacol Res.
152(104631)2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Fernández-Sánchez A, Madrigal-Santillán E,
Bautista M, Esquivel-Soto J, Morales-González A, Esquivel-Chirino
C, Durante-Montiel I, Sánchez-Rivera G, Valadez-Vega C and
Morales-González JA: Inflammation, oxidative stress, and obesity.
Int J Mol Sci. 12:3117–3132. 2011.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Yang J, Xiang D, Xiang D, He W, Liu Y, Lan
L, Li G, Jiang C, Ren X, Liu D and Zhang C: Baicalin protects
against 17α-ethinylestradiol-induced cholestasis via the sirtuin
1/hepatic nuclear receptor-1α/farnesoid X receptor pathway. Front
Pharmacol. 10(1685)2020.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Butler MS, Robertson AAB and Cooper MA:
Natural product and natural product derived drugs in clinical
trials. Nat Prod Rep. 31:1612–1661. 2014.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Sarfraz I, Rasul A, Jabeen F, Younis T,
Zahoor MK, Arshad M and Ali M: Fraxinus: A plant with
versatile pharmacological and biological activities. Evid Based
Complement Alternat Med. 2017(4269868)2017.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Hong S and Kim MM: Aesculetin inhibits
cell invasion through inhibition of MMP-9 activity and antioxidant
activity. J Life Sci. 26:673–679. 2016.
|
|
9
|
Li CX, Li JC, Lai J and Liu Y: The
pharmacological and pharmacokinetic properties of esculin: A
comprehensive review. Phytother Res. 36:2434–2448. 2022.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Zhang L, Xie Q and Li X: Esculetin: A
review of its pharmacology and pharmacokinetics. Phytother Res.
36:279–298. 2022.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Lin Q, Wang W, Li X, Lun J and Shao T:
Ameliorative effect of esculetin against streptozotocin-induced
experimental dementia via activation of Nrf2/HO-1 axis and
suppression of NF-κB. Lat Am J Pharm. 36:399–407. 2017.
|
|
12
|
Medina ME, Galano A and Alvarez-Idaboy JR:
Theoretical study on the peroxyl radicals scavenging activity of
esculetin and its regeneration in aqueous solution. Phys Chem Chem
Phys. 16:1197–1207. 2014.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Zhen AX, Piao MJ, Kang KA, Fernando PDSM,
Kang HK, Koh YS and Hyun JW: Esculetin prevents the induction of
matrix metalloproteinase-1 by hydrogen peroxide in skin
keratinocytes. J Cancer Prev. 24:123–128. 2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Grover J and Jachak SM: Coumarins as
privileged scaffold for anti-inflammatory drug development. RSC
Adv. 5:38892–38905. 2015.
|
|
15
|
Diniz BS, Mendes-Silva AP, Silva LB,
Bertola L, Vieira MC, Ferreira JD, Nicolau M, Bristot G, da Rosa
ED, Teixeira AL and Kapczinski F: Oxidative stress markers
imbalance in late-life depression. J Psychiatr Res. 102:29–33.
2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Wang K, Zhang Y, Ekunwe S, Yi X, Liu X,
Wang H and Pan YM: Antioxidant activity and inhibition effect on
the growth of human colon carcinoma (HT-29) cells of esculetin from
Cortex Fraxini. Med Chem Res. 20:968–974. 2011.
|
|
17
|
Jeong GS, Yoon KH, Kim HC, Oh SH, Kim M,
Kang DG, Lee HS and Kim YC: Cytoprotective constituents of the stem
barks of Fraxinus rhynchophylla on mouse hippocampal HT22 cells and
their antioxidative activity. Korean J Pharmacogn. 38:287–290.
2007.
|
|
18
|
Kim HC, An RB, Jeong GS and Kim YC:
1,1-Diphenyl-2-picrylhydrazyl radical scavenging compounds of
Fraxini Cortex. Nat Prod Sci. 11:150–154. 2005.
|
|
19
|
Vianna DR, Bubols G, Meirelles G, Silva
BV, Da Rocha A, Lanznaster M, Monserrat JM, Garcia SC, Von Poser G
and Eifler-Lima VL: Evaluation of the antioxidant capacity of
synthesized coumarins. Int J Mol Sci. 13:7260–7270. 2012.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Lee JM, Tseng TH and Lee YJ: An efficient
synthesis of neoflavonoid antioxidants based on Montmorillonite
K-10 catalysis. Synthesis. 2247–2254. 2001.
|
|
21
|
Hsia CW, Lin KC, Lee TY, Hsia CH, Chou DS,
Jayakumar T, Velusamy M, Chang CC and Sheu JR: Esculetin, a
coumarin derivative, prevents thrombosis: Inhibitory signaling on
PLCγ2-PKC-AKT activation in human platelets. Int J Mol Sci.
20(2731)2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Lee BC, Lee SY, Lee HJ, Sim GS, Kim JH,
Kim JH, Cho YH, Lee DH, Pyo HB, Choe TB, et al: Anti-oxidative and
photo-protective effects of coumarins isolated from Fraxinus
chinensis. Arch Pharm Res. 30:1293–1301. 2007.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Potapovich MV, Metelitsa DI and Shadyro
OI: Antioxidant activity of hydroxy derivatives of coumarin. Prikl
Biokhim Mikrobiol. 48:282–288. 2012.PubMed/NCBI(In Russian).
|
|
24
|
Rubio V, García-Pérez AI, Herráez A,
Tejedor MC and Diez JC: Esculetin modulates cytotoxicity induced by
oxidants in NB4 human leukemia cells. Exp Toxicol Pathol.
69:700–712. 2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Rubio V, García-Pérez AI, Tejedor MC,
Herráez A and Diez JC: Esculetin neutralises cytotoxicity of t-BHP
but Not of H2O2 on human leukaemia NB4 cells.
Biomed Res Int. 2017(9491045)2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Lee CR, Shin EJ, Kim HC, Choi YS, Shin T
and Wie MB: Esculetin inhibits N-methyl-D-aspartate neurotoxicity
via glutathione preservation in primary cortical cultures. Lab Anim
Res. 27:259–263. 2011.PubMed/NCBI View Article : Google Scholar
|
|
27
|
de Jesus Souza M, De Moraes JA, Da Silva
VN, Helal-Neto E, Uberti AF, Scopel-Guerra A, Olivera-Severo D,
Carlini CR and Barja-Fidalgo C: Helicobacter pylori urease
induces pro-inflammatory effects and differentiation of human
endothelial cells: Cellular and molecular mechanism. Helicobacter.
24(e12573)2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Abdallah H, Farag M, Osman S, Kim DH, Kang
K, Pan CH and Abdel-Sattar E: Isolation of major phenolics from
Launaea spinosa and their protective effect on HepG2 cells damaged
with t-BHP. Pharm Biol. 54:536–541. 2016.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Hong S, Heo H, Lee H, Lee M, Lee J and
Park JH: Protective effects of the methanol extract from calyx of
Diospyros kaki on alcohol-induced liver injury. J Korean Soc Food
Sci Nutr. 50:339–346. 2021.
|
|
30
|
Sen Z, Weida W, Jie M, Li S, Dongming Z
and Xiaoguang C: Coumarin glycosides from Hydrangea paniculata slow
down the progression of diabetic nephropathy by targeting Nrf2
anti-oxidation and smad2/3-mediated profibrosis. Phytomedicine.
57:385–395. 2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Rubio V, García-Pérez AI, Herráez A and
Diez JC: Different roles of Nrf2 and NFKB in the antioxidant
imbalance produced by esculetin or quercetin on NB4 leukemia cells.
Chem Biol Interact. 294:158–166. 2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Gao Y, Shi W, Yao H, Ai Y, Li R, Wang Z,
Liu T, Dai W, Xiao X, Zhao J, et al: An integrative pharmacology
based analysis of refined liuweiwuling against liver injury: A
novel component combination and hepaprotective mechanism. Front
Pharmacol. 12(747010)2021.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Kim JH, Jeong MS, Kim DY, Her S and Wie
MB: Zinc oxide nanoparticles induce lipoxygenase-mediated apoptosis
and necrosis in human neuroblastoma SH-SY5Y cells. Neurochem Int.
90:204–214. 2015.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Gao JL and Gong XY: Esculetin attenuates
neurotoxicity induced by Aβ25-35 in SH-SY5Y cells via
inhibiting oxidative stress and mitochondria-mediated apoptosis.
Acta Pol Pharm. 75:1177–1185. 2018.
|
|
35
|
Pruccoli L, Morroni F, Sita G, Hrelia P
and Tarozzi A: Esculetin as a bifunctional antioxidant prevents and
counteracts the oxidative stress and neuronal death induced by
amyloid protein in SH-SY5Y cells. Antioxidants (Basel).
9(551)2020.PubMed/NCBI View Article : Google Scholar
|
|
36
|
He Y, Li C, Ma Q and Chen S: Esculetin
inhibits oxidative stress and apoptosis in H9c2 cardiomyocytes
following hypoxia/reoxygenation injury. Biochem Biophys Res Commun.
501:139–144. 2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Zhang Y, An Y, He X, Zhang D and He W:
Esculetin protects human corneal epithelial cells from oxidative
stress through Nrf-2 signaling pathway. Exp Eye Res.
202(108360)2021.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Kim Y and Lee J: Esculetin inhibits
adipogenesis and increases antioxidant activity during adipocyte
differentiation in 3T3-L1 cells. Prev Nutr Food Sci. 22:118–123.
2017.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Shi T, Li T, Jiang X, Jiang X, Zhang Q,
Wang Y, Zhang Y, Wang L, Qin X, Zhang W and Zheng Y: Baicalin
protects mice from infection with methicillin-resistant
Staphylococcus aureus via alleviating inflammatory response. J
Leukoc Biol. 108:1829–1839. 2020.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Subramaniam SR and Ellis EM:
Esculetin-induced protection of human hepatoma HepG2 cells against
hydrogen peroxide is associated with the Nrf2-dependent induction
of the NAD(P)H: Quinone oxidoreductase 1 gene. Toxicol Appl
Pharmacol. 250:130–136. 2011.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Lee J, Yang J, Jeon J, Jeong HS, Lee J and
Sung J: Hepatoprotective effect of esculetin on ethanol-induced
liver injury in human HepG2 cells and C57BL/6J mice. J Funct Foods.
40:536–543. 2018.
|
|
42
|
Subramaniam SR and Ellis EM:
Neuroprotective effects of umbelliferone and esculetin in a mouse
model of Parkinson's disease. J Neurosci Res. 91:453–461.
2013.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Xu BR, Zhu LY, Chu J, Ma Z, Fu Q, Wei W,
Deng X and Ma S: Esculetin improves cognitive impairments induced
by transient cerebral ischaemia and reperfusion in mice via
regulation of mitochondrial fragmentation and mitophagy. Behav
Brain Res. 372(112007)2019.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Martín-Aragón S, Villar Á and Benedí J:
Age-dependent effects of esculetin on mood-related behavior and
cognition from stressed mice are associated with restoring brain
antioxidant status. Prog Neuropsychopharmacol Biol Psychiatry.
65:1–16. 2016.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Atmaca M, Bilgin HM, Obay BD, Diken H,
Kelle M and Kale E: The hepatoprotective effect of coumarin and
coumarin derivates on carbon tetrachloride-induced hepatic injury
by antioxidative activities in rats. J Physiol Biochem. 67:569–576.
2011.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Subramaniam SR and Ellis EM: Umbelliferone
and esculetin protect against N-nitrosodiethylamine-induced
hepatotoxicity in rats. Cell Biol Int. 40:761–769. 2016.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Prabakaran D and Ashokkumar N: Protective
effect of esculetin on hyperglycemia-mediated oxidative damage in
the hepatic and renal tissues of experimental diabetic rats.
Biochimie. 95:366–373. 2013.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Han MH, Park C, Lee DS, Hong SH, Choi IW,
Kim GY, Choi SH, Shim JH, Chae JI, Yoo YH and Choi YH:
Cytoprotective effects of esculetin against oxidative stress are
associated with the upregulation of Nrf2-mediated NQO1 expression
via the activation of the ERK pathway. Int J Mol Med. 39:380–386.
2017.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Arora R, Sawney S, Saini V, Steffi C,
Tiwari M and Saluja D: Esculetin induces antiproliferative and
apoptotic response in pancreatic cancer cells by directly binding
to KEAP1. Mol Cancer. 15(64)2016.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Weiwei T, Ting Z, Chunhua M and Hongyan L:
Suppressing receptor-interacting protein 140: A new sight for
esculetin to treat myocardial ischemia/reperfusion injury. RSC Adv.
6:112117–112128. 2016.
|
|
51
|
Türk E, Ozan Tekeli I, Özkan H, Uyar A,
Cellat M, Kuzu M, Yavas I, Alizadeh Yegani A, Yaman T and Güvenç M:
The protective effect of esculetin against aluminium
chloride-induced reproductive toxicity in rats. Andrologia.
53(e13930)2021.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Owczarek A, Kolodziejczyk-Czepas J,
Woźniak-Serwata J, Magiera A, Kobiela N, Wąsowicz K and Olszewska
MA: Potential activity mechanisms of aesculus hippocastanum bark:
Antioxidant effects in chemical and biological in vitro models.
Antioxidants (Basel). 10(995)2021.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Witaicenis A, Seito LN, da Silveira Chagas
A, de Almeida LD Jr, Luchini AC, Rodrigues-Orsi P, Cestari SH and
Di Stasi LC: Antioxidant and intestinal anti-inflammatory effects
of plant-derived coumarin derivatives. Phytomedicine. 21:240–246.
2014.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Raghunath A, Nagarajan R, Sundarraj K,
Palanisamy K and Perumal E: Identification of compounds that
inhibit the binding of Keap1a/Keap1b Kelch DGR domain with Nrf2
ETGE/DLG motifs in zebrafish. Basic Clin Pharmacol Toxicol.
125:259–270. 2019.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Hassanein EHM, Sayed AM, Hussein OE and
Mahmoud AM: Coumarins as modulators of the Keap1/Nrf2/ARE signaling
pathway. Oxid Med Cell Longev. 2020(1675957)2020.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Naaz F, Abdin MZ and Javed S: Protective
effect of esculin against prooxidant aflatoxin B1-induced
nephrotoxicity in mice. Mycotoxin Res. 30:25–32. 2014.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Yao Y, Zhao X, Xin J, Wu Y and Li H:
Coumarins improved type 2 diabetes induced by high-fat diet and
streptozotocin in mice via antioxidation. Can J Physiol Pharmacol.
96:765–771. 2018.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Zhao DL, Zou LB, Lin S, Shi JG and Zhu HB:
Anti-apoptotic effect of esculin on dopamine-induced cytotoxicity
in the human neuroblastoma SH-SY5Y cell line. Neuropharmacology.
53:724–732. 2007.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Wispriyono B, Jalaludin J, Kusnoputranto
H, Pakpahan S, Aryati GP, Pratama S, Librianty N, Rozaliyani A,
Taufik FF and Novirsa R: Glutathione (GSH) and superoxide dismutase
(SOD) levels among junior high school students induced by indoor
particulate matter 2.5 (PM2.5) and nitrogen dioxide (NO2) exposure.
J Public Health Res. 10(2372)2021.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Kaneko T, Baba N and Matsuo M: Protection
of coumarins against linoleic acid hydroperoxide-induced
cytotoxicity. Chem Biol Interact. 142:239–254. 2003.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Qi SP and Yu JC: Protective effect of
esculin in hyperoxic lung injury in neonatal rats. La Am J Pharm.
35:1177–1181. 2016.
|
|
62
|
Liu A, Shen Y, Du Y, Chen J, Pei F, Fu W
and Qiao J: Esculin prevents
Lipopolysaccharide/D-Galactosamine-induced acute liver injury in
mice. Microb Pathog. 125:418–422. 2018.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Song Y, Wang X, Qin S, Zhou S, Li J and
Gao Y: Esculin ameliorates cognitive impairment in experimental
diabetic nephropathy and induces anti-oxidative stress and
anti-inflammatory effects via the MAPK pathway. Mol Med Rep.
17:7395–7402. 2018.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Zheng L, Yang L, Wang Z, Chen C and Su Y:
Protective effect of Esculin in adjuvant-induced arthritic (AIA)
rats via attenuating pro-inflammatory cytokines and oxidative
stress. Cell Mol Biol (Noisy-le-grand). 61:1–5. 2015.PubMed/NCBI
|
|
65
|
Lee H, Lee JH, Koh SJ and Park H:
Bidirectional relationship between atopic dermatitis and
inflammatory bowel disease: A systematic review and meta-analysis.
J Am Acad Dermatol. 83:1385–1394. 2020.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Nakajima A, Yamamoto Y, Yoshinaka N, Namba
M, Matsuo H, Okuyama T, Yoshigai E, Okumura T, Nishizawa M and
Ikeya Y: A new flavanone and other flavonoids from green perilla
leaf extract inhibit nitric oxide production in interleukin
1β-treated hepatocytes. Biosci Biotechnol Biochem. 79:138–146.
2015.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Kim AR, Jin Q, Jin HG, Ko HJ and Woo ER:
Phenolic compounds with IL-6 inhibitory activity from Aster yomena.
Arch Pharm Res. 37:845–851. 2014.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Wang F, Jia QW, Yuan ZH, Lv LY, Li M,
Jiang ZB, Liang DL and Zhang DZ: An anti-inflammatory C-stiryl
iridoid from Camptosorus sibiricus Rupr. Fitoterapia. 134:378–381.
2019.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Hu J, Zhao L, Li N, Yang Y, Qu T, Ren H,
Cui X, Tao H, Chen Z and Peng Y: Investigation of the active
ingredients and pharmacological mechanisms of Porana sinensis
Hemsl. Against rheumatoid arthritis using network pharmacology and
experimental validation. PLoS One. 17(e0264786)2022.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Lee MW, Kang SJ, Park M, Yoon JH, Han BH,
Choi SE and Lee MW: Anti-oxidative and nitric oxide production
inhibitory activities of phenoliccompounds from the fruits of
actinidia arguta. Nat Prod Sci. 12:221–225. 2006.
|
|
71
|
Kim Y, Park Y, Namkoong S and Lee J:
Esculetin inhibits the inflammatory response by inducing heme
oxygenase-1 in cocultured macrophages and adipocytes. Food Funct.
5:2371–2377. 2014.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Sun B, Wang B and Xu M: Esculetin inhibits
histamine-induced expression of inflammatory cytokines and mucin in
nasal epithelial cells. Clin Exp Pharmacol Physiol. 46:821–827.
2019.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Hong SH, Jeong HK, Han MH, Park C and Choi
YH: Esculetin suppresses lipopolysaccharide-induced inflammatory
mediators and cytokines by inhibiting nuclear factor-κB
translocation in RAW 264.7 macrophages. Mol Med Rep. 10:3241–3246.
2014.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Lee SJ, Lee US, Kim WJ and Moon SK:
Inhibitory effect of esculetin on migration, invasion and matrix
metalloproteinase-9 expression in TNF-α-induced vascular smooth
muscle cells. Mol Med Rep. 4:337–341. 2011.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Xie C, Li Y, Gao J and Wang Y: Esculetin
regulates the phenotype switching of airway smooth muscle cells.
Phytother Res. 33:3008–3015. 2019.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Song WJ, Yun JH, Jeong MS, Kim KN, Shin T,
Kim HC and Wie MB: Inhibitors of lipoxygenase and cyclooxygenase-2
attenuate trimethyltin-induced neurotoxicity through regulating
oxidative stress and pro-inflammatory cytokines in human
neuroblastoma SH-SY5Y cells. Brain Sci. 11(1116)2021.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Ozal SA, Turkekul K, Gurlu V, Guclu H and
Erdogan S: Esculetin protects human retinal pigment epithelial
cells from lipopolysaccharide-induced inflammation and cell death.
Curr Eye Res. 43:1169–1176. 2018.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Torres R, Mascayano C, Núñez C, Modak B
and Faini F: Coumarins of haplopappus multifolius and derivative as
inhibitors of lox: Evaluation in-vitro and docking studies. J Chil
Chem Soc. 58:2027–2030. 2013.
|
|
79
|
Kwon OS, Choi JS, Islam MN, Kim YS and Kim
HP: Inhibition of 5-lipoxygenase and skin inflammation by the
aerial parts of Artemisia capillaris and its constituents. Arch
Pharm Res. 34:1561–1569. 2011.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Lee HC, Liu FC, Tsai CN, Chou AH, Liao CC
and Yu HP: Esculetin ameliorates lipopolysaccharide-induced acute
lung injury in mice via modulation of the AKT/ERK/NF-κB and
RORγt/IL-17 pathways. Inflammation. 43:962–974. 2020.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Chen T, Guo Q, Wang H, Zhang H, Wang C,
Zhang P, Meng S, Li Y, Ji H and Yan T: Effects of esculetin on
lipopolysaccharide (LPS)-induced acute lung injury via regulation
of RhoA/Rho Kinase/NF-кB pathways in vivo and in vitro. Free Radic
Res. 49:1459–1468. 2015.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Oh SY, Kim YH, Kang MK, Lee EJ, Kim DY, Oh
H, Kim SI, Na W and Kang YH: Aesculetin attenuates alveolar injury
and fibrosis induced by close contact of alveolar epithelial cells
with blood-derived macrophages via IL-8 signaling. Int J Mol Sci.
21(5518)2020.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Hongyan L: Esculetin attenuates Th2 and
Th17 responses in an ovalbumin-induced asthmatic mouse model.
Inflammation. 39:735–743. 2016.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Oh SY, Kim YH, Kang MK, Lee EJ, Kim DY, Oh
H, Kim SI, Na W, Kang IJ and Kang YH: Aesculetin inhibits airway
thickening and mucus overproduction induced by urban particulate
matter through blocking inflammation and oxidative stress involving
TLR4 and EGFR. Antioxidants (Basel). 10(494)2021.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Yum S, Jeong S, Lee S, Kim W, Nam J and
Jung Y: HIF-prolyl hydroxylase is a potential molecular target for
esculetin-mediated anti-colitic effects. Fitoterapia. 103:55–62.
2015.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Wang SK, Chen TX, Wang W, Xu LL, Zhang YQ,
Jin Z, Liu YB and Tang YZ: Aesculetin exhibited anti-inflammatory
activities through inhibiting NF-кB and MAPKs pathway in vitro and
in vivo. J Ethnopharmacol. 296(115489)2022.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Witaicenis A, Luchini AC, Hiruma-Lima CA,
Felisbino SL, Justulin LA Jr, Garrido-Mesa N, Utrilla P, Gálvez J
and Di Stasi LC: Mechanism and effect of esculetin in an
experimental animal model of inflammatory bowel disease. Eur J
Inflamm. 11:433–446. 2013.
|
|
88
|
Xu J, Wang H, Wen H, Zhang J and Wu A:
Evaluation of antioxidant, antiulcer, and analgesic activities of
esculetin. La Am J Pharm. 40:1584–1591. 2021.
|
|
89
|
Choi RY, Ham JR and Lee MK: Esculetin
prevents non-alcoholic fatty liver in diabetic mice fed high-fat
diet. Chem Biol Interact. 260:13–21. 2016.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Song WJ, Kim J, Shin T, Jeong MS, Kim KN,
Yun JH and Wie MB: Esculetin and fucoidan attenuate autophagy and
apoptosis induced by zinc oxide nanoparticles through modulating
reactive astrocyte and proinflammatory cytokines in the rat brain.
Toxics. 10(194)2022.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Zhu L, Nang C, Luo F, Pan H, Zhang K, Liu
J, Zhou R, Gao J, Chang X, He H, et al: Esculetin attenuates
lipopolysaccharide (LPS)-induced neuroinflammatory processes and
depressive-like behavior in mice. Physiol Behav. 163:184–192.
2016.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Sulakhiya K, Keshavlal GP, Bezbaruah BB,
Dwivedi S, Gurjar SS, Munde N, Jangra A, Lahkar M and Gogoi R:
Lipopolysaccharide induced anxiety- and depressive-like behaviour
in mice are prevented by chronic pre-treatment of esculetin.
Neurosci Lett. 611:106–111. 2016.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Jeong NH, Yang EJ, Jin M, Lee JY, Choi YA,
Park PH, Lee SR, Kim SU, Shin TY, Kwon TK, et al: Esculetin from
Fraxinus rhynchophylla attenuates atopic skin inflammation by
inhibiting the expression of inflammatory cytokines. Int
Immunopharmacol. 59:209–216. 2018.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Chen Y, Zhang Q, Liu H, Lu C, Liang CL,
Qiu F, Han L and Dai Z: Esculetin ameliorates psoriasis-like skin
disease in mice by inducing CD4+Foxp3+
regulatory T cells. Front Immunol. 9(2092)2018.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Cheng YJ, Tian XL, Zeng YZ, Lan N, Guo LF,
Liu KF, Fang HL, Fan HY and Peng ZL: Esculetin protects against
early sepsis via attenuating inflammation by inhibiting NF-κB and
STAT1/STAT3 signaling. Chin J Nat Med. 19:432–441. 2021.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Jiang D, Liu X and Hu J: Topical
administration of Esculetin as a potential therapy for experimental
dry eye syndrome. Eye (Lond). 31:1724–1732. 2017.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Pullaiah CP, Nelson VK, Rayapu S, G V NK
and Kedam T: Exploring cardioprotective potential of esculetin
against isoproterenol induced myocardial toxicity in rats: In vivo
and in vitro evidence. BMC Pharmacol Toxicol. 22(43)2021.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Singh L, Kaur A, Garg S, Singh AP and
Bhatti R: Protective effect of esculetin, natural coumarin in mice
model of fibromyalgia: Targeting pro-inflammatory cytokines and
MAO-A. Neurochem Res. 45:2364–2374. 2020.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Yang XD, Chen Z, Ye L, Chen J and Yang YY:
Esculin protects against methionine choline-deficient diet-induced
non-alcoholic steatohepatitis by regulating the Sirt1/NF-κ B p65
pathway. Pharm Biol. 59:922–932. 2021.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Tian X, Peng Z, Luo S, Zhang S, Li B, Zhou
C and Fan H: Aesculin protects against DSS-induced colitis though
activating PPARγ and inhibiting NF-кB pathway. Eur J Pharmacol.
857(172453)2019.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Choi HJ, Chung TW, Kim JE, Jeong HS, Joo
M, Cha J, Kim CH and Ha KT: Aesculin inhibits matrix
metalloproteinase-9 expression via p38 mitogen activated protein
kinase and activator protein 1 in lipopolysachride-induced RAW264.7
cells. Int Immunopharmacol. 14:267–274. 2012.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Niu X, Wang Y, Li W, Zhang H, Wang X, Mu
Q, He Z and Yao H: Esculin exhibited anti-inflammatory activities
in vivo and regulated TNF-α and IL-6 production in LPS-stimulated
mouse peritoneal macrophages in vitro through MAPK pathway. Int
Immunopharmacol. 29:779–786. 2015.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Tianzhu Z and Shumin W: Esculin inhibits
the inflammation of LPS-induced acute lung injury in mice via
regulation of TLR/NF-κB pathways. Inflammation. 38:1529–1536.
2015.PubMed/NCBI View Article : Google Scholar
|
|
104
|
Li W, Wang Y, Wang X, Zhang H, He Z, Zhi
W, Liu F and Niu X: Gastroprotective effect of esculin on
ethanol-induced gastric lesion in mice. Fundam Clin Pharmacol.
31:174–184. 2017.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Kim YR, Park BK, Seo CS, Kim NS and Lee
MY: Antidepressant and anxiolytic-like effects of the stem bark
extract of fraxinus rhynchophylla hance and its components in a
mouse model of depressive-like disorder induced by reserpine
administration. Front Behav Neurosci. 15(650833)2021.PubMed/NCBI View Article : Google Scholar
|
|
106
|
Chen T, Zheng M, Li Y, Liu S and He L: The
role of CCR5 in the protective effect of Esculin on
lipopolysaccharide-induced depressive symptom in mice. J Affect
Disord. 277:755–764. 2020.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Cheng X, Yang Y, Li W, Liu M, Zhang S,
Wang Y and Du G: Esculin alleviates acute kidney injury and
inflammation induced by LPS in mice and its possible mechanism. J
Chin Pharm Sci. 29:322–332. 2020.
|
|
108
|
Wang YH, Liu YH, He GR, Lv Y and Du GH:
Esculin improves dyslipidemia, inflammation and renal damage in
streptozotocin-induced diabetic rats. BMC Complement Altern Med.
15(402)2015.PubMed/NCBI View Article : Google Scholar
|
|
109
|
Li W, Wang Y, Wang X, He Z, Liu F, Zhi W,
Zhang H and Niu X: Esculin attenuates endotoxin shock induced by
lipopolysaccharide in mouse and NO production in vitro through
inhibition of NF-κB activation. Eur J Pharmacol. 791:726–734.
2016.PubMed/NCBI View Article : Google Scholar
|
|
110
|
Cai T and Cai B: Pharmacological
activities of esculin and esculetin: A review. Medicine
(Baltimore). 102(e35306)2023.PubMed/NCBI View Article : Google Scholar
|