Introduction
Breast carcinoma stands as a prevalent malignancy
impacting women across the globe (1). Every year, there are 2.3 million new
cases and 685,000 deaths. North America, Europe and Australia have
the highest incidence rate, while Asia and parts of Africa have
lower incidence rates (2). It is
worth noting that although relatively rare, BC can also affect men.
This malignant tumor typically presents as a lump or lump in the
breast, often accompanied by changes in shape, size or skin texture
(3). The intricacy of molecular
mechanisms that govern tumorigenesis and progression accounts for
the heterogeneity inherent in breast cancer. From a molecular
standpoint, such variability creates challenges regarding the
choice of therapeutic strategies and the disease prognosis
(4). The genomic landscapes of
HR+, HER2+ and triple-negative cancer cells,
as well as the tumor immune microenvironment, often lead to
different immune infiltrations and functions (5). Immune cells play a crucial role in
the microenvironment of breast cancer, affecting the growth,
metastasis and response to the treatment of tumors. The in-depth
study of the interaction between immune cells and breast cancer
reveals new therapeutic targets, opening up broad possibilities for
developing new therapeutic strategies (6,7). The
ultimate goal of tumor immunotherapy with specific targets is to
trigger antitumor immune responses, leading to clinical regression
and/or recurrence of tumors. In Bergman Phase I to III trials,
there are many types of specific tumor immunotherapies involving a
wide range of tumor types. Due to slow progress, the response to
cancer vaccines and other cancer immunotherapies may take several
months or longer to emerge (8).
Although immune-targeted therapies have achieved early successes in
cancers such as melanoma and lung cancer, the progress of
immune-targeted treatment for breast cancer has been relatively
slow (9). Therefore, it is
necessary to explore the immune infiltration-associated molecular
mechanisms of the progression of breast cancer to improve the
survival rate of patients with breast cancer.
Heterogeneous nuclear RNAs are the primary
transcripts of RNA polymerase II in eukaryotes (10). Heterogeneous nuclear
ribonucleoproteins (HNRNPs) are the most abundant nuclear proteins
in higher eukaryotes and are a class of commonly recognized
RNA-binding proteins (11). HNRNPs
play important roles in key cellular processes such as
transcription, post-transcriptional modifications and translation.
The dysregulation of HNRNPs is a key factor in cancer development
and drug resistance. HNRNPs promote the diversity of tumor and
immune-related abnormal proteomes by controlling alternative
splicing and translation; they can also promote the expression of
cancer-related genes by regulating transcription factors, directly
binding to DNA or promoting chromatin remodeling (12). The HNRNPAB subfamily is the core
member of HNRNPs. In recent years, the biological value of HNRNPAB
has been discussed to some extent (13). The biogenesis mechanism of HNRNPAB,
which penetrates into all aspects of cellular RNA metabolism,
involves DNA binding, RNA splicing and transport and translation
and stability of mRNA (14). The
expression of HNRNPAB changes depending on cancer type, suggesting
its role in tumorigenesis (15-17).
However, the role and immune mechanism of HNRNPAB in breast cancer
are still unclear. Thus, the present study aimed to investigate the
role of HNRNPAB in breast cancer progression. Publicly accessible
databases and cellular assays were employed to assess the
prognostic and predictive importance of HNRNPAB expression, with a
concurrent investigation into its potential as a diagnostic
biomarker and therapeutic candidate for breast cancer.
Materials and methods
Cell culture protocol
MDA-MB-231, the breast cancer cell line, was
procured from Wuhan Servicebio Technology Co., Ltd. These cells
were maintained in DMEM medium (Gibco; Thermo Fisher Scientific,
Inc.) supplemented with 10% FBS. The cell cultures were incubated
in a humidified 5% CO2 environment at a constant
temperature of 37˚C.
HNRNPAB gene knockdown
A 6-well plate was prepared by plating
5x105 MDA-MB-231 breast cancer cells per well, followed
by a 24 h stabilization period in the incubator. The siRNA
sequences used in the present study were as follows: HNRNPAB-siRNA
sense strand: 5'-GGAGAGGUCGUUGACUGUAdTdT-3', antisense strand:
5'-UACAGUCAACGACCUCUCCdAdA-3'; negative control (NC)-siRNA sense
strand: 5'-UUCUCCGAACGUGUCACGUdTdT-3', antisense strand:
5'-ACGUGACACGUUCGGAGAAdTdT-3'. Subsequently, NC-siRNA and
HNRNPAB-siRNA (50 nm) (Tianjin Sheweisi Biotechnology Co., Ltd.)
were individually introduced into the MDA-MB-231 cells at 37˚C for
24-72 h using Lipofectamine™ 3000 reagent (Thermo Fisher
Scientific, Inc.) in strict accordance with the manufacturer's
guidelines. At 8 h post-transfection, the transfection medium was
replaced with fresh DMEM containing 10% FBS. The efficiency of
siRNA transfection was quantified and the results are presented
with corresponding SD values.
Reverse transcription-quantitative PCR
(RT-qPCR)
For this assay, MDA-MB-231 cells following siRNA
transfection were selected. The culture medium was aspirated and
lysis buffer (Wuhan Servicebio Technology Co., Ltd.) was added to
the cells. Total RNA was isolated using a commercial RNA extraction
kit (Tiangen Biotech Co., Ltd.). After quantifying the RNA
concentration, reverse transcription into cDNA was performed using
the ABScript II RT Master Mix for qPCR with gDNA Remover (ABclonal
Biotech Co., Ltd.) as per the manufacturers' instructions. The
synthesized cDNA was stored long-term at -80˚C. Fluorescence qPCR
was conducted on a LightCycler480 real-time fluorescence
quantitative PCR instrument (Roche Diagnostics GmbH), with cDNA
loaded according to the protocol of the Genious 2X SYBR Green Fast
qPCR Mix kit (ABclonal Biotech Co., Ltd.). The standardized thermal
cycling conditions were set as follows: Initial pre-denaturation at
95˚C for 3 min; followed by 40 cycles of denaturation at 95˚C for
15 sec, annealing at 60˚C for 20 sec and extension at 72˚C for 20
sec. Melting curve analysis was subsequently performed from 65˚C to
95˚C to verify the specificity of amplification products. The
primer sequences used were as follows: HNRNPAB forward,
5'-TTTGGCGAGTTTGGGGAGATT-3' and reverse,
5'-GCCATACTGCTGCTGATAGAC-3'; GAPDH forward,
5'-GGAGCGAGATCCCTCCAAAAT-3' and reverse,
5'-GGCTGTTGTCATACTTCTCATGG-3'. GAPDH served as the internal
reference gene and the 2-ΔΔCq method was applied to
calculate the relative mRNA expression levels (18). All experimental reactions were
conducted in triplicate to ensure reproducibility.
Cell Counting Kit-8 (CCK-8) assay
Logarithmic-phase MDA-MB-231 cells transfected with
either siRNA or NC-siRNA were digested with trypsin for 2 min and
then seeded into a 96-well plate at a density of 8x104
cells per ml. After incubation for 24, 48 and 72 h in a 5%
CO2 incubator at 37˚C, the 96-well plate was removed.
The culture medium was discarded and a mixture of DMEM and CCK-8
solution (Jiangsu Kaiji Biotechnology Co., Ltd.) at a 9:1 ratio was
added to each well. After 1 h, the optical density value of the
cells was measured at a wavelength of 450 nm. The cell
proliferation rate was calculated, with the blank knockdown group
designated as the control.
Cell migration and invasion assay
(Transwell chambers)
Firstly, the upper chamber of a Transwell insert
(Corning, Inc.) was coated at 37˚C for 2 h with 100 µl 10% Matrigel
(BD Biosciences) diluted in serum-free DMEM, followed by a 2 h
incubation in a cell culture incubator. Cells were then seeded into
the upper chamber of the 8 µm pore-sized Transwell at a density of
5x104 cells per well. The lower chamber was filled with
750 µl culture medium containing 10% FBS and the cells were
incubated for 48 h at 37˚C in a 5% CO2 environment.
After incubation, the medium in the upper chamber was discarded.
Cells remaining in the chamber were fixed at room temperature with
4% paraformaldehyde for 30 min and stained with 0.1% crystal violet
at room temperature for 15 min. The number of cells was counted in
three randomly selected fields of view per chamber using an
inverted light microscope.
Public database data acquisition
HNRNPAB expression levels in BRCA tissues from The
Cancer Genome Atlas (TCGA; https://portal.gdc.cancer.gov/) and Genotype-Tissue
Expression (GTEx) databases were retrieved through the Gene
Expression Profiling Interactive Analysis 2 platform (19-21).
TCGA pan-cancer (PANCAN) dataset was acquired from the UCSC Xena
browser (https://xenabrowser.net/), which offers
a standardized and comprehensive compilation of pan-cancer
datasets. Immunohistochemistry data for HNRNPAB was obtained from
The Human Protein Atlas (HPA) database (https://www.proteinatlas.com) (22) which characterizes the expression
pattern of HNRNPAB protein in both BRCA tissues and normal breast
tissues. Prognostic survival analysis of BRCA sample data was
conducted using the Kaplan-Meier Plotter online tool (http://kmplot.com/analysis/) (23) to explore the association between
HNRNPAB and the prognosis of patients with BRCA. The cBioportal
(https://www.cbioportal.org) (24) and TCGA databases were utilized to
investigate HNRNPAB-associated gene alterations in patients with
breast cancer. Immunotherapy outcomes and immune cell infiltration
profiles of patients with breast cancer were collected from The
Cancer Immunome Database (TCIA; https://tcia.at)
and the influence of HNRNPAB on immune checkpoint inhibitor
efficacy was predicted using the Immune Phenotype Score (IPS)
(25). In addition, the
StromalScore, ImmuneScore and ESTIMATEScore for BRCA samples in
TCGA database was computed using the ‘estimate’ package in R
software (26) and the immune cell
types associated with HNRNPAB expression levels were
summarized.
Gene Set Enrichment Analysis
(GSEA)
To further characterize differentially expressed
genes and elucidate the functions of potential target genes, gene
function enrichment analyses were performed using HALLMARK
(https://www.gsea-msigdb.org/gsea/msigdb/index.jsp),
REACTOME (https://reactome.org/) and Kyoto
Encyclopedia of Genes and Genomes (KEGG; https://www.kegg.jp/) databases. The ‘ClusterProfiler’
package (version 3.18.0) in R software was employed to analyze
pathway enrichment, thereby enhancing the understanding of
mRNA-associated carcinogenic mechanisms (27-29).
Gene matrix transposed files were downloaded from the Molecular
Signatures Database (https://www.gsea-msigdb.org/gsea/msigdb/) and GSEA was
applied to conduct enrichment analyses for HALLMARK, REACTOME and
KEGG gene sets (30-32).
Genes with |log2 fold change| >1 and adjusted
P<0.05 were defined as differentially expressed genes, and
enriched terms with adjusted P<0.05 were considered
statistically significant.
Statistical analysis
A total of three independent experiments were
performed for each assay (n=3) to guarantee the reliability and
reproducibility of the experimental findings. Experimental data are
presented as the mean ± SD, which is used to illustrate the central
tendency and degree of dispersion of the results. For comparisons
among multiple groups, one-way ANOVA was employed and Tukey's post
hoc test was subsequently conducted to identify the specific
differences between individual groups. The paired Student's t-test
was utilized for comparisons between two groups. Pearson's
correlation analysis was applied to evaluate the linear correlation
between relevant indicators. All statistical analyses were carried
out with SPSS (version 26.0; IBM Corp.) software with P<0.05
considered to indicate a statistically significant difference.
Results
Association between HNRNPAB mRNA
expression and clinicopathological parameters/clinical prognosis in
patients with BRCA
To comprehensively characterize HNRNPAB expression
patterns across diverse tissues of patients with BRCA and its
association with clinicopathological features, the standardized
pan-cancer dataset TCGA TARGET GTEx was retrieved (PANCAN;
n=19,131; genes=60,499) from the UCSC Xena browser. HNRNPAB was
found to be significantly upregulated in 24 tumor types, whereas
downregulation was detected in 2 tumor types (Fig. 1A). Differential analysis of normal
and tumor tissues from TCGA and GTEx databases further demonstrated
that HNRNPAB expression was significantly higher in BRCA tissues
compared with normal breast tissues (Fig. 1B). Immunohistochemical data of
HNRNPAB from the HPA database demonstrated that at the protein
level, HNRNPAB expression was elevated in BRCA tissues compared
with normal breast tissues (Fig.
1C and D). Overall survival
analysis revealed that high HNRNPAB mRNA expression was closely
associated with unfavorable prognosis in patients with BRCA
(Fig. 1E). As breast cancer
progressed to advanced stages, the median expression level of
HNRNPAB exhibited a declining trend (Fig. 1F). The number of lymph node
metastases in patients with breast cancer showed a positive
association with increased median HNRNPAB expression (Fig. 1G). Patients with BRCA aged 21-40
years displayed a marked increase in median HNRNPAB expression,
with elevated levels also observed in other age groups compared
with normal tissue (Fig. 1H).
HNRNPAB expression was found to be notable higher in invasive
ductal carcinoma and medullary carcinoma compared with normal
tissues, while a mild elevation was noted in other histological
subtypes (Fig. 1I). In addition,
Pearson's correlation was calculated between HNRNPAB expression and
ploidy across a number of tumors and thus identified a significant
positive correlation in BRCA. The expression of HNRNPAB positively
correlated with tumor ploidy in breast cancer, suggesting that
further investigation is warranted (Fig. 1J).
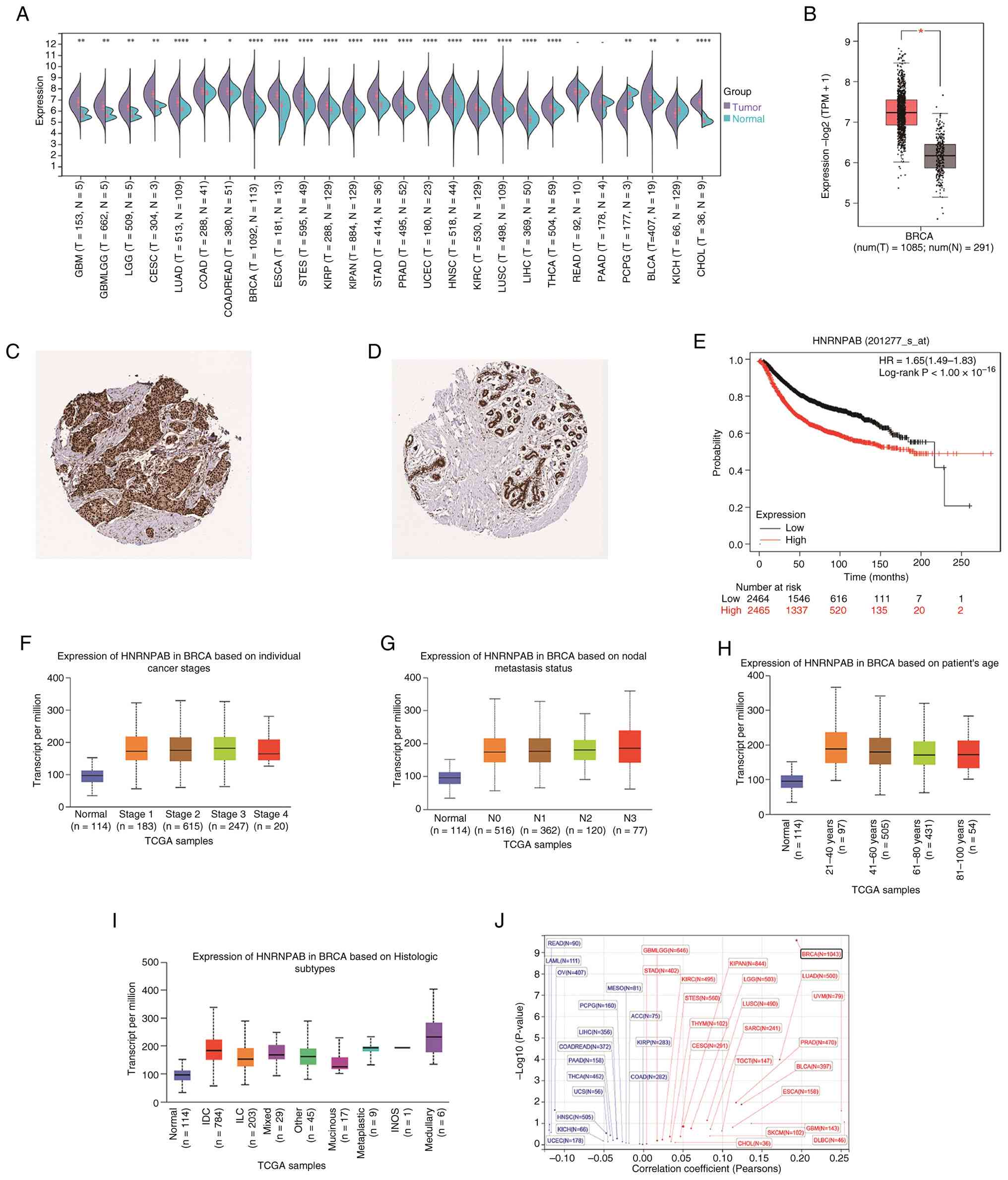 | Figure 1Expression of HNRNPAB and its
association with clinical prognosis of patients with breast cancer.
(A) Differential expression of HNRNPAB in different cancer types
based on pan-cancer analysis. (B) Expression of HNRNPAB in BRCA
tissue samples in TCGA database is higher than compared with
paracancerous tissues of breast cancer. HNRNPAB
immunohistochemistry images obtained from the Human Protein Atlas
database show that the expression level of HNRNPAB protein in (C)
breast cancer tissues is higher compared with that in (D)
paracancerous tissues of breast cancer. (E) The influence of
HNRNPAB expression on the prognosis of patients with breast cancer
analyzed by the Kaplan-Meier method. Box plots describing the
expression analysis of HNRNPAB according to (F) stage, (G) lymph
node metastasis, (H) age and (I) breast cancer histology. (J)
Scatter plot showing the correlation between HNRNPAB expression and
ploidy in various tumors, with red indicating a positive
correlation and blue indicating a negative correlation. HNRNPAB,
heterogeneous nuclear ribonucleoprotein A/B; TPM, transcripts per
million; HR, hazard ratio; TCGA, The Cancer Genome Atlas; IDC,
invasive ductal carcinoma; ILC, invasive lobular carcinoma; INOS,
infiltrating carcinoma not otherwise specified.
*P<0.05, **P<0.01 and
****P<0.0001. |
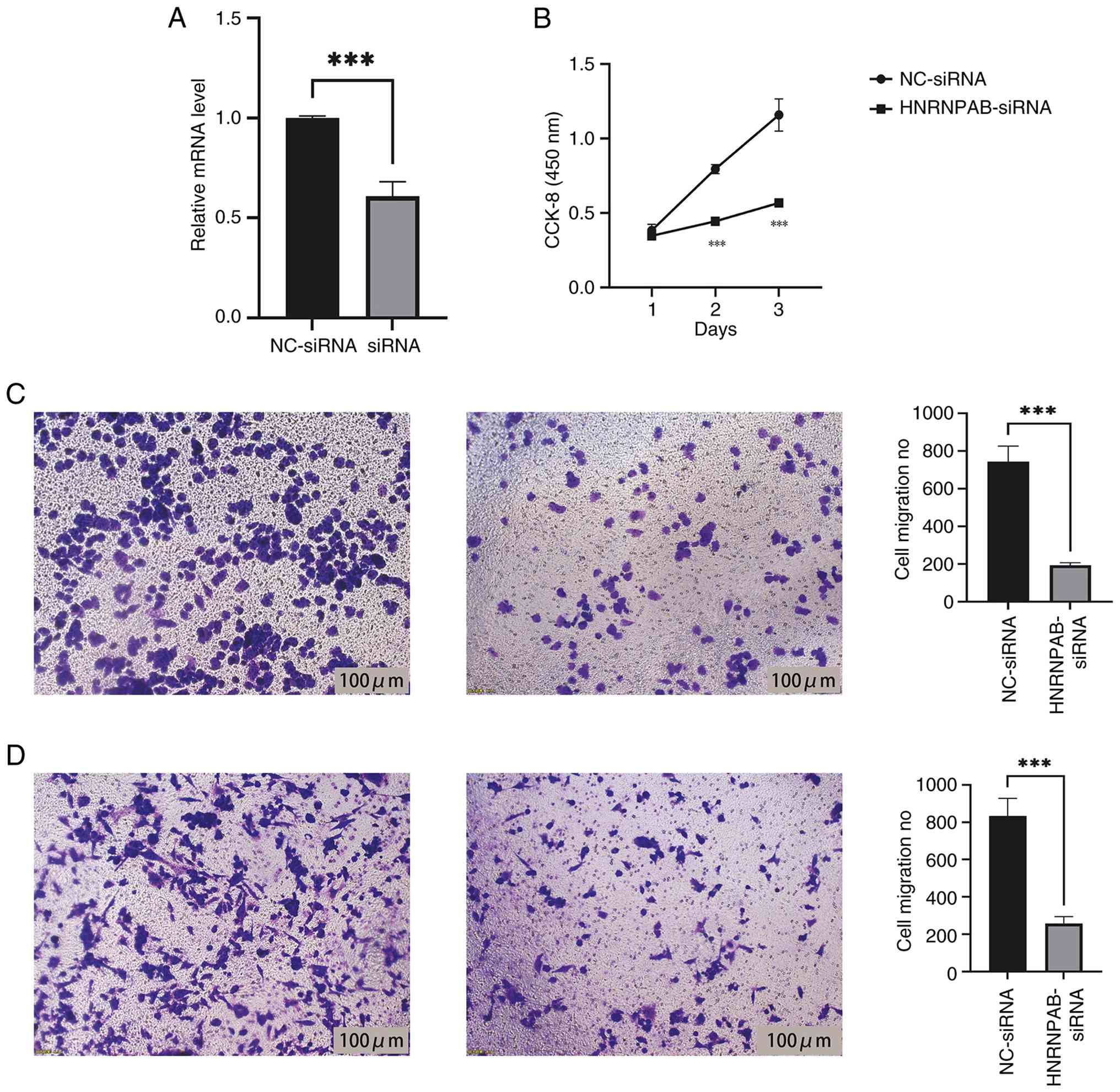
Function of HNRNPAB in proliferation,
migration and invasion of breast cancer cells
Pre-transfection with siRNA targeting HNRNPAB
effectively reduced HNRNPAB levels in MDA-MB-231 cells, leading to
a significant decrease in intracellular HNRNPAB content (Fig. 2A). Proliferation, migration and
invasion assays were performed to compare two groups, namely the
MDA-MB-231 siRNA control group and the MDA-MB-231 HNRNPAB-siRNA
group. The CCK-8 assay results indicated that knockdown of HNRNPAB
expression significantly inhibited the proliferation of breast
cancer cells (Fig. 2B). Transwell
migration and invasion experiments further demonstrated that
HNRNPAB promoted a significant decrease in cancer cell migration
and invasion (Fig. 2C and D).
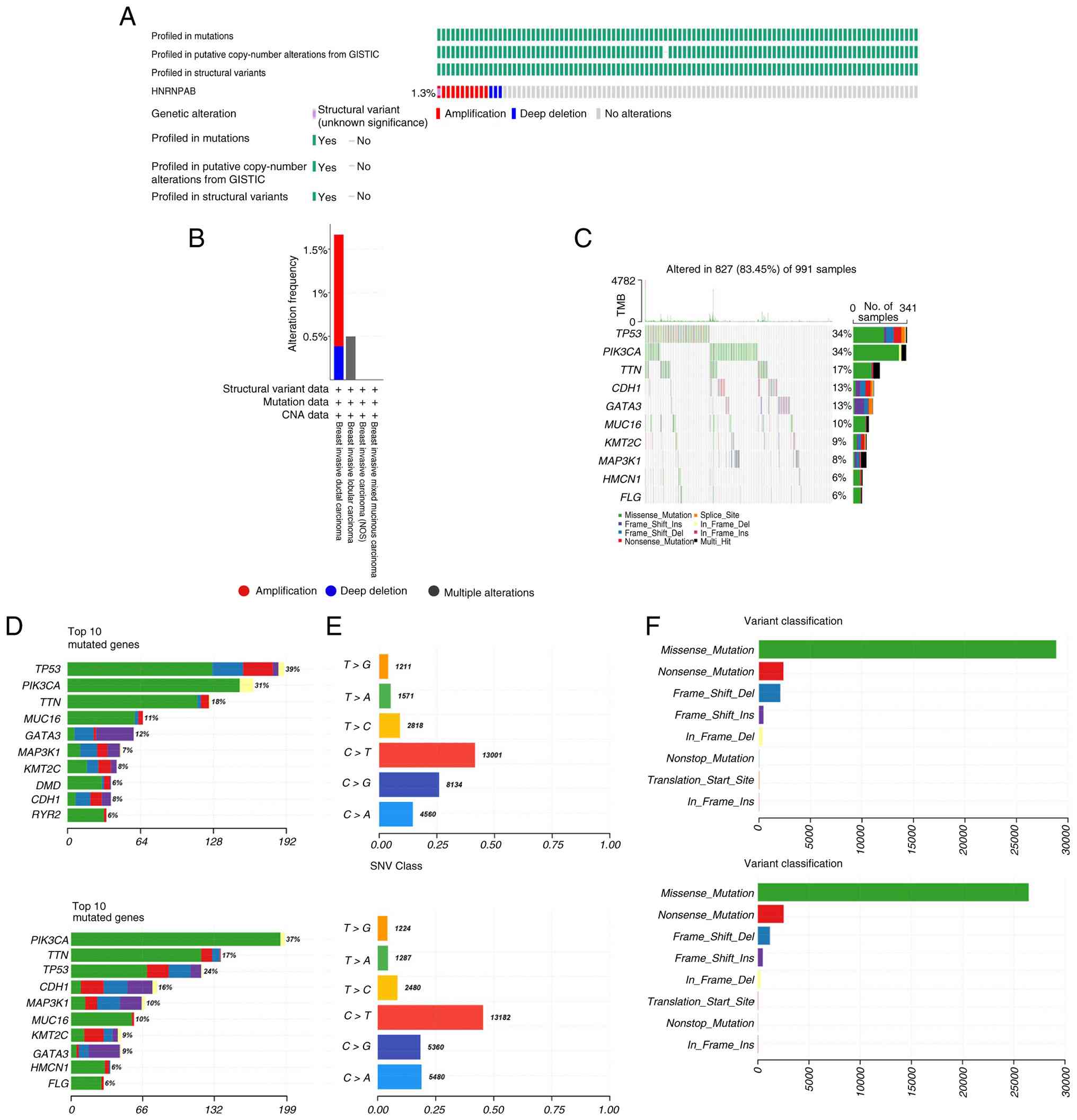
Genetic alteration landscape of
HNRNPAB
To explore the genetic architecture and
transcriptional variations of HNRNPAB, the cBioportal database was
utilized. With regard to genetic alterations of HNRNPAB, 1.3% of
patients exhibited modifications, with amplification being the
predominant type of alteration in cancer (Fig. 3A). Among varying breast cancer
subtypes, invasive ductal carcinoma exhibited the highest frequency
of HNRNPAB mutations, followed by invasive lobular carcinoma. By
contrast, invasive mucinous breast carcinoma and breast invasive
carcinoma (not otherwise specified) displayed minimal HNRNPAB
mutation rates (Fig. 3B). Analysis
of the breast tumor cohort landscape revealed that distinct HNRNPAB
expression levels were associated with specific genetic
alterations. The high HNRNPAB expression group exhibited
significant differences in the mutation status of PIK3CA, titin
(TTN) and tumor protein p53 (TP53), whereas the low expression
group exhibited higher mutation frequencies of these three genes.
Therefore, mutations in PIK3CA, TP53 and TTN may affect the
progression of breast cancer regulated by HNRNPAB (Fig. 3C and D). In the high HNRNPAB expression group,
C>G mutations ranked second and C>A mutations ranked third.
Conversely, in the low expression group, C>A mutations were the
second most common, surpassing C>G mutations (Fig. 3E). Differences were also observed
in variation classifications between the two groups: In the high
HNRNPAB group, ‘Nonstop_Mutation’ ranked fifth (surpassing
‘Translation_Start_Site’ mutations at sixth), while in the low
expression group, ‘Translation_Start_Site’ mutations occupied the
fifth position (surpassing ‘Nonstop_Mutations’ at sixth; Fig. 3F). These discrepancies may be
attributed to altered HNRNPAB expression; however, further studies
are required to clarify their impact on breast cancer progression,
which will enhance the general understanding of HNRNPAB
function.
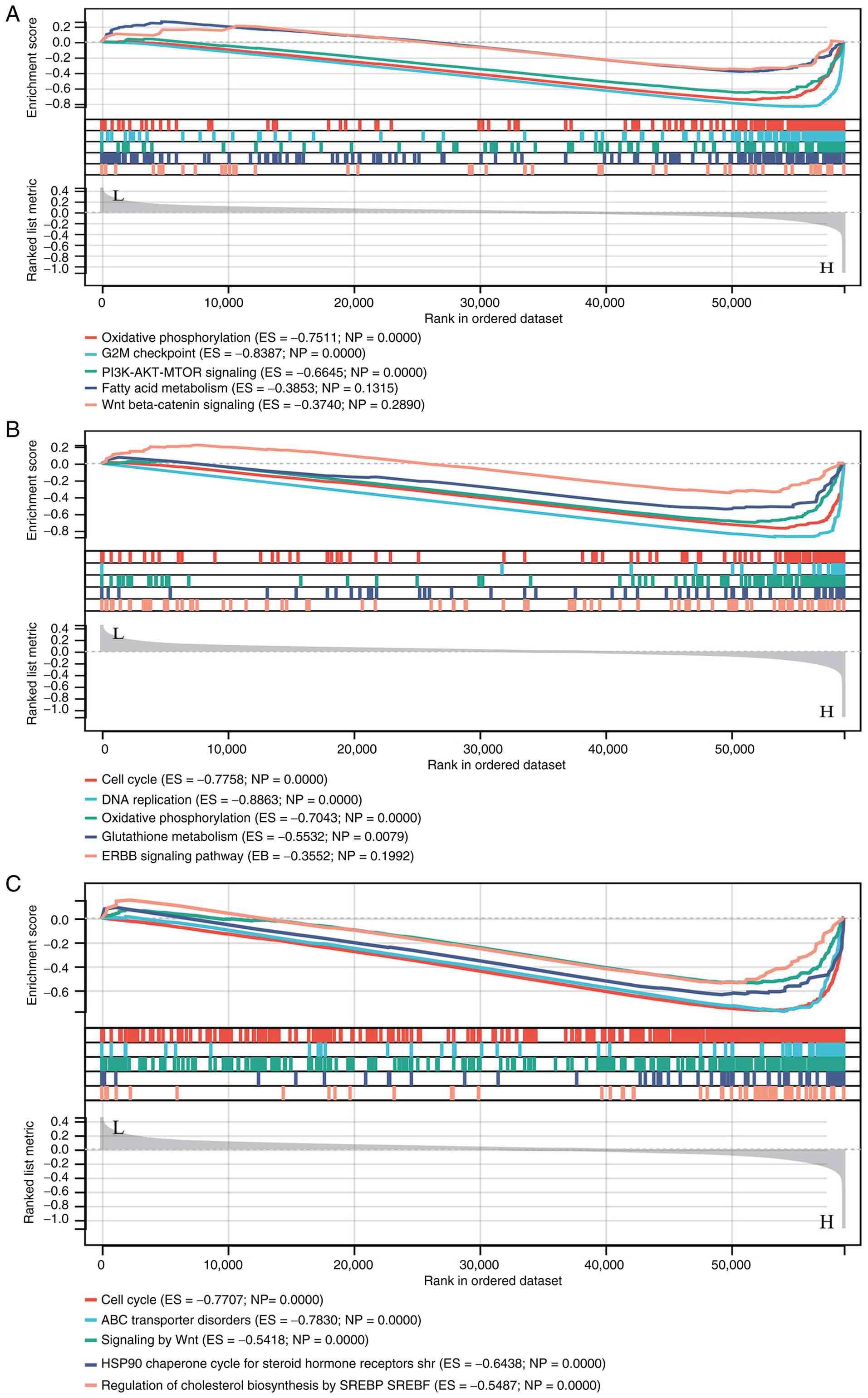
HNRNPAB-associated enrichment
analyses
HALLMARK enrichment analysis suggested that HNRNPAB
may participate in ‘OXIDATIVE_PHOSPHORYLATION’, ‘G2M_CHECKPOINT’,
‘PI3K_AKT_MTOR_SIGNALING’, ‘FATTY_ACID_METABOLISM’ and
‘WNT_BETA_CATENIN_SIGNALING’ (Fig.
4A). KEGG enrichment analysis indicated that HNRNPAB is
primarily involved in ‘CELL_CYCLE’, ‘DNA_REPLICATION’,
‘OXIDATIVE_PHOSPHORYLATION’, ‘GLUTATHIONE_METABOLISM’ and the
‘ERBB_SIGNALING_PATHWAY’ (Fig.
4B). REACTOME enrichment analysis results demonstrated that
HNRNPAB may be involved in ‘CELL_CYCLE’,
‘ABC_TRANSPORTER_DISORDERS’, ‘SIGNALING_BY_WNT’,
‘HSP90_CHAPERONE_CYCLE_FOR_STEROID_HORMONE_RECEPTORS_SHR’ and
‘REGULATION_OF_CHOLESTEROL_BIOSYNTHESIS_BY_SREBP_SREBF’ (Fig. 4C).
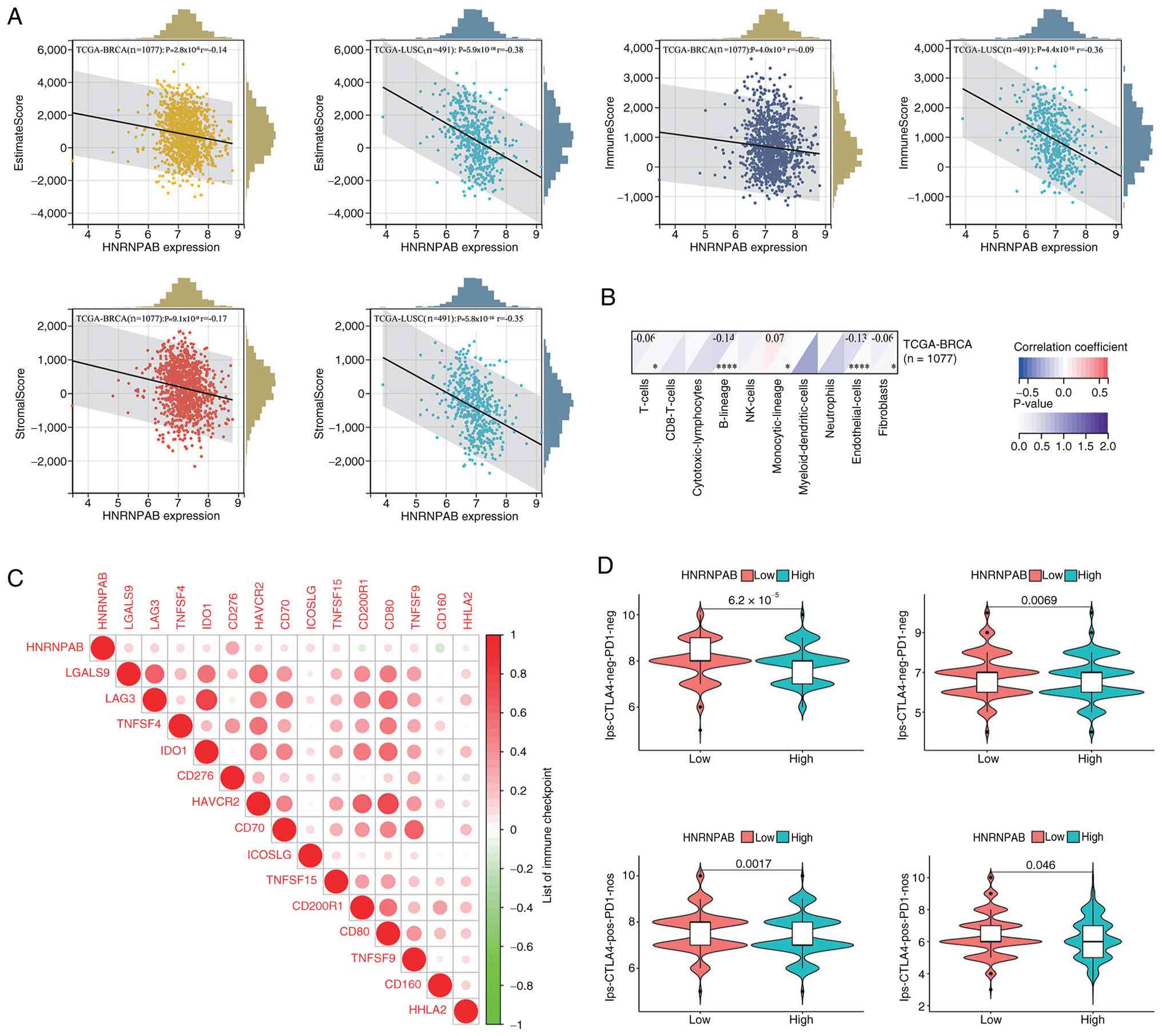
HNRNPAB association with the tumor
microenvironment and tumor immunotherapy
ESTIMATE algorithm analysis results showed that the
StromalScore, ImmuneScore and ESTIMATEScore of the low HNRNPAB
expression group was significantly higher compared with those of
the high expression group (Fig.
5A). Although reduced immune cell infiltration was observed,
the present findings revealed a phenomenon whereby the abundance of
‘Monocytic_lineage’ cells increased with elevated HNRNPAB
expression (Fig. 5B). Correlation
analysis between HNRNPAB and immune checkpoint gene expression
levels indicated that HNRNPAB was significantly positively
correlated with 11 immune checkpoint genes and negatively
correlated with 3 immune checkpoint genes (Fig. 5C). IPS data of patients with BRCA
from TCIA database demonstrated that the low HNRNPAB expression
group exhibited a higher IPS and improved responsiveness to immune
checkpoint inhibitors (Fig.
5D).
This result suggests that HNRNPAB downregulation is
associated with enhanced immunotherapeutic potential, highlighting
an important prognostic and predictive implication for BRCA
immunotherapy.
Correlation between HNRNPAB and breast
cancer stem cells
Based on the aforementioned enrichment results, it
was hypothesized that HNRNPAB participates in biological processes
such as cell cycle regulation, DNA replication and metabolic
control, all of which are associated with stem cell functions.
Therefore, the standardized TCGA pan-cancer dataset was retrieved
from the UCSC Xena browser. Specifically, HNRNPAB expression data
was extracted from each sample and the RNA stemness score for each
tumor was calculated based on mRNA signatures. Pearson correlation
coefficients were computed for each tumor type, revealing
significant correlations in 26 tumors. Notably, 24 tumor types
(including breast cancer) exhibited a positive correlation, while 2
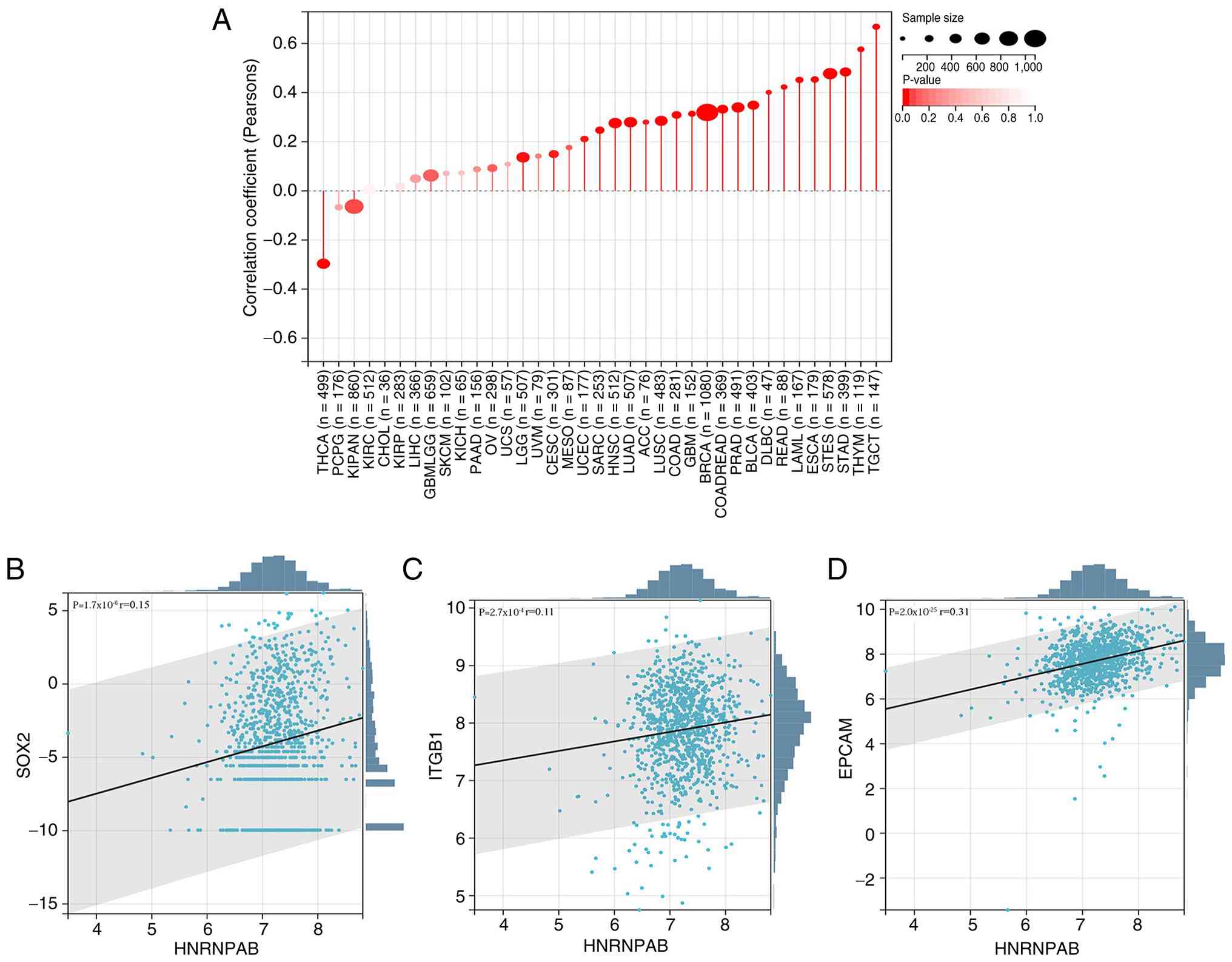
tumor types exhibited a negative correlation (Fig. 6A). In addition, the correlation
between HNRNPAB and common breast cancer stem cell markers was
explored, identifying significant positive correlations with SOX2,
integrin subunit β-1 (ITGB1), epithelial cell adhesion molecule
(EPCAM) (Fig. 6B-D). Collectively,
these preliminary results suggested that HNRNPAB may positively
regulate breast cancer stem cells.
Discussion
In summary, the present study utilized online
databases to elucidate the variations in HNRNPAB mRNA expression
and its associations with clinical prognosis, cancer stem cell
biomarkers, genetic mutation profiles and HNRNPAB-associated
signaling pathway alterations, with a particular focus on immune
infiltration during breast cancer progression. Furthermore, in
vitro experiments were performed to validate the impact of
HNRNPAB on the proliferative, migratory and invasive capacities of
breast cancer cells.
Through analyses of public repositories including
TCGA and HPA, it was determined that HNRNPAB is markedly
upregulated in breast cancer cells and tissues relative to normal
breast cells and adjacent non-tumor tissues. Concurrently,
significant upregulation of HNRNPAB in 24 tumor types (including
endometrial carcinoma) and downregulation in 2 tumor types
(including renal chromophobe carcinoma) was observed. This pattern
aligns with the upregulation of HNRNPAB reported in a number of
malignancies by previous research (33-35),
indicating its potential utility as a diagnostic biomarker for
tumors, particularly breast cancer. CCK-8 and Transwell assays
further demonstrated that HNRNPAB facilitates the proliferation,
migration and invasion of breast cancer cells, suggesting it acts
as an oncogenic risk factor promoting breast cancer initiation and
progression. HNRNPAB serves key roles in both normal biological
processes and cancer development (36-39).
Beyond its involvement in Harvey rat sarcoma viral oncogene homolog
oncogene inactivation, observed elevated HNRNPAB levels in solid
tumor metastases have demonstrated its pivotal function in tumor
progression (40,41). Prognostic analyses in the current
study have shown that high HNRNPAB expression impacts the survival
and clinical outcomes of patients with breast cancer. The present
study revealed that the median HNRNPAB expression level was
significantly higher in breast cancer patients aged 21-40 years
compared with normal tissues, indicating close associations with
age and disease classification.
As well-established cancer hallmarks, ~75% of solid
tumors exhibit aneuploidy and chromosomal instability, resulting in
complex and heterogeneous karyotypic profiles (42,43).
Assessing tumor ploidy provides valuable insights into cancer
genome evolution and tumor heterogeneity. In the present study,
HNRNPAB expression exhibited a positive correlation with tumor
ploidy in breast cancer, indicating its close association with
polyploidy and chromosomal instability in this malignancy, thus
deepening the understanding of its role in disease progression. In
addition, it was found that the majority of HNRNPAB alterations
during breast tumorigenesis stem from mRNA expression amplification
or mutation, a novel finding that, to the best of our knowledge,
has not been previously reported.
However, it has been documented that HNRNPAB can
enhance stemness characteristics in human stem cells and reduce
their sensitivity to colorectal cancer chemotherapeutics (44). Yet, research regarding the
associations and regulatory mechanisms of HNRNPAB in breast cancer
stem cell modulation remains limited. Previous studies have
indicated a regulatory association between HNRNPAB and the
Wnt/β-catenin pathway: Decreased expression of the upstream factor
microRNA-8063 attenuates its inhibitory effect on HNRNPAB, leading
to activation of the Wnt/β-catenin signaling cascade and subsequent
suppression of tumor metastasis (17,45).
Based on these bioinformatics findings, the present study
hypothesized that HNRNPAB may be involved in numerous stem
cell-associated biological processes, such as cell cycle
progression, DNA replication and metabolic regulation.
In KEGG and other databases, it was identified that
the primary active pathways in the upregulated HNRNPAB group were
associated with cell cycle signaling transduction pathways, which
are aberrant pathways common to all malignancies (46). A number of components of cell cycle
signaling pathways trigger uncontrolled cell division when mutated
and these are closely associated with the mutation frequencies of
numerous genes. These enrichment analysis findings indicate that
HNRNPAB may precisely regulate the metabolic reprogramming process
of tumor cells, break the normal metabolic homeostasis of tumor
cells, reshape the energy supply mode and material anabolism
network of tumor cells, before then regulating the proliferation
activity, invasion and metastasis ability as well as the
anti-apoptosis potential of breast cancer cells; thus, ultimately
participating in and regulating the occurrence, development and
malignant progression of breast cancer. This hypothesis may be
further determined and improved in future research through targeted
metabolomics detection techniques, combined with multidimensional
research methods such as in vitro cell function experiments,
in vivo animal model validation and clinical sample
correlation analysis.
A number of studies have reported that HNRNP family
proteins may promote PI3K/AKT/FOXO1-mediated bladder cancer cell
proliferation (47,48). Previous research has also
demonstrated an association between the PI3K/AKT/mTOR pathway and
immune responses (49). The
present HALLMARK-based GSEA analyses demonstrated a strong linear
association between HNRNPAB and the PI3K/AKT/mTOR pathway,
indicating that HNRNPAB may promote breast cancer development by
activating this pathway. Building upon this finding, further
immune-associated bioinformatics investigations of HNRNPAB were
conducted. From the perspective of clinical practice, although the
current immunotherapy for breast cancer has brought survival
benefits to a number of patients. The immunocheckpoint inhibitor
pabolizumab combined with chemotherapy has been approved by the FDA
for the treatment of PD-L1-positive metastatic and early
triple-negative breast cancer, while the ongoing clinical trial
aims to expand the current treatment pattern of immunocheckpoint
inhibitors in hormone receptor-positive and HER2-positive breast
cancer. Antibody-conjugated drugs have been approved by the FDA for
triple-negative and HER2-positive diseases, and their combination
with immune checkpoint inhibitors is currently being studied.
Vaccines and bispecific antibodies are active research areas. There
are still problems such as limited response rate and marked
individual differences (50).
HNRNPAB, as a potential marker for predicting the responsiveness of
immunotherapy, if its specificity and sensitivity can be further
determined through large sample clinical verification, may serve as
an important reference for clinicians to develop individualized
immunotherapy strategies for breast cancer and also provide new
research targets and ideas for further exploring the immune escape
mechanism of breast cancer and developing new combined
immunotherapy programs.
In conclusion, while dedicated mechanistic studies
are lacking and limited in-depth explorations of how HNRNPAB
modulates immune cell infiltration and immune checkpoint regulation
have been translated into clinical settings, the unique expression
patterns of HNRNPAB, as well as it's transcriptional profile,
interactions with cancer stem cells and associations with lipid
metabolism and tumor immunity, provide a novel perspective for
HNRNPAB-targeted molecular therapy in patients with breast cancer.
These findings also contribute to a deeper understanding of the
molecular mechanisms driving breast cancer tumorigenesis. However,
further experiments are required to elucidate the specific
mechanisms by which HNRNPAB regulates tumor immune infiltration and
its role in immune checkpoint blockade responses. This will be the
aim of future in-depth investigations conducted on this topic.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the Ningbo Health
Science and Technology Plan Project (grant no. 2024Y07), the Ningbo
Public Welfare Science and Technology Program (grant no. 2024S144)
and the Health Commission of Zhejiang Province (grant no.
2024KY1546).
Availability of data and materials
The data generated in the present study are included
in the figures and/or tables of this article.
Authors' contributions
JX and SZ designed the research and confirmed the
authenticity of all the raw data. QH performed experiments. LC, MY
and JH performed statistical analysis. JX and SZ wrote the
manuscript. JX and QH prepared figures. LC, MY, JH and SL
interpreted the data. All authors have read and approved the final
manuscript. JX and QH confirm the authenticity of all the raw
data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Barzaman K, Karami J, Zarei Z,
Hosseinzadeh A, Kazemi MH, Moradi-Kalbolandi S, Safari E and
Farahmand L: Breast cancer: Biology, biomarkers, and treatments.
Int Immunopharmacol. 84(106535)2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Xu W, Zhang T, Zhu Z and Yang Y: The
association between immune cells and breast cancer: Insights from
Mendelian randomization and meta-analysis. Int J Surg. 111:230–241.
2025.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Nafissi N, Saghafinia M, Motamedi MH and
Akbari ME: A survey of breast cancer knowledge and attitude in
Iranian women. J Cancer Res Ther. 8:46–49. 2012.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Lüönd F, Tiede S and Christofori G: Breast
cancer as an example of tumour heterogeneity and tumour cell
plasticity during malignant progression. Br J Cancer. 125:164–175.
2021.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Onkar SS, Carleton NM, Lucas PC, Bruno TC,
Lee AV, Vignali DAA and Oesterreich S: The great immune escape:
Understanding the divergent immune response in breast cancer
subtypes. Cancer Discov. 13:23–40. 2023.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Li JJ, Tsang JY and Tse GM: Tumor
microenvironment in breast cancer-updates on therapeutic
implications and pathologic assessment. Cancers (Basel).
13(4233)2021.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Zhao H, Yin X and Wang L, Liu K, Liu W, Bo
L and Wang L: Identifying tumour microenvironment-related signature
that correlates with prognosis and immunotherapy response in breast
cancer. Sci Data. 10(119)2023.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Bergman PJ: Cancer immunotherapy. Vet Clin
North Am Small Anim Pract. 54:441–468. 2024.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Esteva FJ, Hubbard-Lucey VM, Tang J and
Pusztai L: Immunotherapy and targeted therapy combinations in
metastatic breast cancer. Lancet Oncol. 20:e175–e186.
2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Lu Y, Wang X, Gu Q, Wang J, Sui Y, Wu J
and Feng J: Heterogeneous nuclear ribonucleoprotein A/B: An
emerging group of cancer biomarkers and therapeutic targets. Cell
Death Discov. 8(337)2022.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Geuens T, Bouhy D and Timmerman V: The
hnRNP family: Insights into their role in health and disease. Hum
Genet. 135:851–867. 2016.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Sudhakaran M and Doseff AI: Role of
heterogeneous nuclear ribonucleoproteins in the cancer-immune
landscape. Int J Mol Sci. 24(5086)2023.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Neriec N and Percipalle P: Sorting mRNA
molecules for cytoplasmic transport and localization. Front Genet.
9(510)2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Thibault PA, Ganesan A, Kalyaanamoorthy S,
Clarke JWE, Salapa HE and Levin MC: hnRNP A/B Proteins: An
encyclopedic assessment of their roles in homeostasis and disease.
Biology (Basel). 10(712)2021.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Zhou ZJ, Dai Z, Zhou SL, Hu ZQ, Chen Q,
Zhao YM, Shi YH, Gao Q, Wu WZ, Qiu SJ, et al: HNRNPAB induces
epithelial-mesenchymal transition and promotes metastasis of
hepatocellular carcinoma by transcriptionally activating SNAIL.
Cancer Res. 74:2750–2762. 2014.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Pan T, Yu Z, Jin Z, Wu X, Wu A, Hou J,
Chang X, Fan Z, Li J, Yu B, et al: Tumor suppressor lnc-CTSLP4
inhibits EMT and metastasis of gastric cancer by attenuating
HNRNPAB-dependent Snail transcription. Mol Ther Nucleic Acids.
23:1288–1303. 2021.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Zhou J, Chen S, Liu J, Du J and Li J:
Knockdown of hnRNPAB reduces the stem cell properties and enhances
the chemosensitivity of human colorectal cancer stem cells. Oncol
Rep. 49(129)2023.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Livak KJ and Schmittgen TD: . Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Blum A, Wang P and Zenklusen JC: SnapShot:
TCGA-Analyzed tumors. Cell. 173(530)2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
GTEx Consortium: The genotype-tissue
expression (GTEx) project. Nat Genet. 45:580–585. 2013.PubMed/NCBI View
Article : Google Scholar
|
|
21
|
Tang Z, Kang B, Li C, Chen T and Zhang Z:
GEPIA2: An enhanced web server for large-scale expression profiling
and interactive analysis. Nucleic Acids Res. 47(W1):W556–W560.
2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Zhao P, Zhen H, Zhao H, Huang Y and Cao B:
Identification of hub genes and potential molecular mechanisms
related to radiotherapy sensitivity in rectal cancer based on
multiple datasets. J Transl Med. 21(176)2023.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Ranstam J and Cook JA: Kaplan-Meier curve.
Br J Surg. 104(442)2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Cerami E, Gao J, Dogrusoz U, Gross BE,
Sumer SO, Aksoy BA, Jacobsen A, Byrne CJ, Heuer ML, Larsson E, et
al: The cBio cancer genomics portal: an open platform for exploring
multidimensional cancer genomics data. Cancer Discov. 2:401–404.
2012.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Tsujimoto H and Osafune K: Current status
and future directions of clinical applications using iPS
cells-focus on Japan. FEBS J. 289:7274–7291. 2022.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Sepulveda JL: Using R and bioconductor in
clinical genomics and transcriptomics. J Mol Diagn. 22:3–20.
2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Chen L, Zhang YH, Wang S, Zhang Y, Huang T
and Cai YD: Prediction and analysis of essential genes using the
enrichments of gene ontology and KEGG pathways. PLoS One.
12(e0184129)2017.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Wu B, Fu L, Guo X, Hu H, Li Y, Shi Y,
Zhang Y, Han S, Lv C and Tian Y: Multi-omics profiling and digital
image analysis reveal the potential prognostic and
immunotherapeutic properties of CD93 in stomach adenocarcinoma.
Front Immunol. 14(984816)2023.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Rappsilber J, Mann M and Ishihama Y:
Protocol for micro-purification, enrichment, pre-fractionation and
storage of peptides for proteomics using StageTips. Nat Protoc.
2:1896–1906. 2007.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Lei L, Bai YH, Jiang HY, He T, Li M and
Wang JP: A bioinformatics analysis of the contribution of m6A
methylation to the occurrence of diabetes mellitus. Endocr Connect.
10:1253–1265. 2021.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Zhao X, Zhang L, Wang J, Zhang M, Song Z,
Ni B and You Y: Identification of key biomarkers and immune
infiltration in systemic lupus erythematosus by integrated
bioinformatics analysis. J Transl Med. 19(35)2021.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Huang R, Liao X and Li Q: Identification
of key pathways and genes in TP53 mutation acute myeloid leukemia:
Evidence from bioinformatics analysis. Onco Targets Ther.
11:163–173. 2017.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Yang Y, Chen Q, Piao HY, Wang B, Zhu GQ,
Chen EB, Xiao K, Zhou ZJ, Shi GM, Shi YH, et al: HNRNPAB-regulated
lncRNA-ELF209 inhibits the malignancy of hepatocellular carcinoma.
Int J Cancer. 146:169–180. 2020.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Xu C, Li B, Yu N, Yao B, Wang F and Mei Y:
The c-Myc targeting hnRNPAB promotes lung adenocarcinoma cell
proliferation via stabilization of CDK4 mRNA. Int J Biochem Cell
Biol. 156(106372)2023.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Xin R, Shen B, Jiang YJ, Liu JB, Li S, Hou
LK, Wu W, Jia CY, Wu CY, Fu D, et al: Comprehensive analysis to
identify a novel PTEN-associated ceRNA regulatory network as a
prognostic biomarker for lung adenocarcinoma. Front Oncol.
12(923026)2022.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Han SP, Tang YH and Smith R: Functional
diversity of the hnRNPs: Past, present and perspectives. Biochem J.
430:379–392. 2010.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Carpenter B, MacKay C, Alnabulsi A, MacKay
M, Telfer C, Melvin WT and Murray GI: The roles of heterogeneous
nuclear ribonucleoproteins in tumour development and progression.
Biochim Biophys Acta. 1765:85–100. 2006.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Matta A, Tripathi SC, DeSouza LV, Grigull
J, Kaur J, Chauhan SS, Srivastava A, Thakar A, Shukla NK, Duggal R,
et al: Heterogeneous ribonucleoprotein K is a marker of oral
leukoplakia and correlates with poor prognosis of squamous cell
carcinoma. Int J Cancer. 125:1398–1406. 2009.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Ma YL, Peng JY, Zhang P, Huang L, Liu WJ,
Shen TY, Chen HQ, Zhou YK, Zhang M, Chu ZX and Qin HL:
Heterogeneous nuclear ribonucleoprotein A1 is identified as a
potential biomarker for colorectal cancer based on differential
proteomics technology. J Proteome Res. 8:4525–4535. 2009.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Mikheev AM, Mikheev SA, Zhang Y, Aebersold
R and Zarbl H: CArG binding factor A (CBF-A) is involved in
transcriptional regulation of the rat Ha-ras promoter. Nucleic
Acids Res. 28:3762–3770. 2000.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Ramaswamy S, Ross KN, Lander ES and Golub
TR: A molecular signature of metastasis in primary solid tumors.
Nat Genet. 33:49–54. 2003.PubMed/NCBI View
Article : Google Scholar
|
|
42
|
Kuang X and Li J: Chromosome instability
and aneuploidy as context-dependent activators or inhibitors of
antitumor immunity. Front Immunol. 13(895961)2022.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Castellanos G, Camargo-Herrera LV, Rangel
N, Jiménez-Tobón GA, Martínez-Agüero M and Rondón-Lagos M:
Exploring chromosomal instability and clonal heterogeneity in
breast cancer. Endocr Relat Cancer. 31(e240096)2024.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Tauler J, Zudaire E, Liu H, Shih J and
Mulshine JL: hnRNP A2/B1 modulates epithelial-mesenchymal
transition in lung cancer cell lines. Cancer Res. 70:7137–7147.
2010.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Chen ZQ, Yuan T, Jiang H, Yang YY, Wang L,
Fu RM, Luo SQ, Zhang T, Wu ZY and Wen KM: MicroRNA-8063 targets
heterogeneous nuclear ribonucleoprotein AB to inhibit the
self-renewal of colorectal cancer stem cells via the Wnt/β-catenin
pathway. Oncol Rep. 46(219)2021.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Glaviano A, Singh SK, Lee EHC, Okina E,
Lam HY, Carbone D, Reddy EP, O'Connor MJ, Koff A, Singh G, et al:
Cell cycle dysregulation in cancer. Pharmacol Rev.
77(100030)2025.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Li F, Xie W, Fang Y, Xie K, Liu W, Hou L
and Tan W: HnRNP-F promotes the proliferation of bladder cancer
cells mediated by PI3K/AKT/FOXO1. J Cancer. 12:281–291.
2021.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Shi X, Ran L, Liu Y, Zhong SH, Zhou PP,
Liao MX and Fang W: Knockdown of hnRNP A2/B1 inhibits cell
proliferation, invasion and cell cycle triggering apoptosis in
cervical cancer via PI3K/AKT signaling pathway. Oncol Rep.
39:939–950. 2018.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Passacantilli I, Frisone P, De Paola E,
Fidaleo M and Paronetto MP: hnRNPM guides an alternative splicing
program in response to inhibition of the PI3K/AKT/mTOR pathway in
Ewing sarcoma cells. Nucleic Acids Res. 45:12270–12284.
2017.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Guo JN, Chen D, Deng SH, Huang JR, Song
JX, Li XY, Cui BB and Liu YL: Identification and quantification of
immune infiltration landscape on therapy and prognosis in left- and
right-sided colon cancer. Cancer Immunol Immunother. 71:1313–1330.
2022.PubMed/NCBI View Article : Google Scholar
|