Introduction
Acute kidney injury (AKI) is a clinical syndrome
characterised by a rapid decline in kidney function over a short
period of time. AKI remains a major global health burden, with
recent evidence showing that it affects millions of individuals
worldwide each year and is associated with high mortality and an
increased risk of progression to chronic kidney disease. Therefore,
AKI has become an important public health problem requiring early
prevention and effective therapeutic strategies (1). Pathogenic factors for AKI include
hypovolemia, septic shock, nephrotoxic drug use and urinary tract
obstruction (2), among which 12.2%
of AKI cases are associated with the use of nephrotoxic
therapeutics (3).
Cisplatin (Cis) is widely used to treat a number of
solid cancer types, including lung, ovarian, testicular, bladder,
and head and neck cancer. However, owing to its side effects,
particularly nephrotoxicity, its clinical application is limited.
Cis-induced acute kidney injury (Cis-AKI), which is characterised
by renal tubular injury, inflammation and impaired renal function,
occurs in 20-30% of treated patients despite hydration-based
preventive measures. Identifying effective drugs for treating
kidney injury and elucidating their mechanisms of action therefore
remains a challenge (4-6).
However, owing to its side effects, including nephrotoxicity,
ototoxicity, hepatotoxicity, gastrointestinal toxicity and
neurotoxicity, its clinical application is markedly limited,
especially due to Cis nephrotoxicity, as patients with tumours
often suffer from Cis-induced AKI (Cis-AKI), which is characterised
by acute tubular necrosis and inflammation, with an incidence rate
as high as 20-30% (7). Identifying
effective drugs for treating kidney injury and elucidating their
mechanisms of action therefore remains a challenge (8).
BaiYangJie is the dry whole grass of Arundina
graminifolia (D. Don) Hochr. (Laeli, Arundina
Blume), whose primary active chemical components are stilbenes,
phenanthrene and flavonoids, which exhibit anti-inflammatory,
antioxidant, antibacterial and antitumour activities (9). BaiYangJie is a characteristic
medicine of the Dai nationality and is used to treat urinary system
diseases, including urinary tract infections and chemical,
pharmaceutical and food-induced poisoning, including liver and
kidney detoxification (9).
Previously, it has been found that the methanolic extract of
BaiYangJie (MEAG) can improve Cis-AKI (10). Despite this discovery, the
mechanism by which MEAG improves Cis-AKI remains unclear, hindering
its clinical application. Therefore, it is necessary to further
explore its mechanisms of action.
Metabolomics has the unique potential to identify
biomarkers that predict the incidence, severity and progression of
diseases, including disease-related endogenous metabolites and
metabolic signatures, and to reveal perturbed metabolic pathways,
thereby providing novel clues to mechanistic abnormalities
(11). Therefore, metabolomics is
often used to study botanical drugs in complex situations (12). Both tools account for the
complexity of biological systems and can complement each other,
which is beneficial for the systematic study of diseases with
complex mechanisms. Consequently, network pharmacology was combined
with serum metabolomics to comprehensively examine how MEAG affects
Cis-AKI in mice, providing a theoretical basis for the future
development of anti-renal injury drugs.
Materials and methods
Drugs and reagents
Arundina graminifolia (D. Don) Hochr samples
used in the present study were collected from the Yunnan Branch of
the Institute of Medicinal Plant Development (Chinese Academy of
Medical Sciences, Jinghong, China). It was identified as
Arundina graminifolia (D. Don) Hochr. by Research Professor
Lixia Zhang (Yunnan Branch of the Institute of Medicinal Plant
Development, Chinese Academy of Medical Sciences, Jinghong, China).
The appraisal report is shown in Fig.
S1.
The following reagents were used in the present
study: Resveratrol (cat. no. 111535; National Institutes for Food
and Drug Control), pterostilbene (cat. no. 455753; J&K Chemical
Ltd.), acetonitrile [American Chemical Society/high-performance
liquid chromatography (HPLC) grade; cat. no. AH015-4HC; Honeywell
International Inc.], methanol (HPLC grade; cat. no. 01104351;
Adamas-Beta® Ltd.), formic acid (HPLC grade; cat. no.
F112034; Shanghai Aladdin Biochemical Technology Co., Ltd.),
distilled water [Watsons Water; AS Watson Group (HK) Ltd.], Cis
(cat. no. 275688; J&K Chemical Ltd.), H&E (cat. no. G1120)
and Masson staining kit (cat. no. G1346; both from Beijing Solarbio
Science & Technology Co., Ltd.), kidney injury molecule 1
(KIM-1)/hepatitis A virus cellular receptor 1 rabbit polyclonal
antibody (cat. no. 30948-1-AP) and lipocalin-2/neutrophil
gelatinase-associated lipocalin (NGAL) rabbit polyclonal antibody
(cat. no. 30700-1-AP; discontinued; both from ProteinTech Group,
Inc.), mouse IL-6 (cat. no. YJ063159), IL-1β (cat. no. YJ35346) and
TNF-α (cat. no. YJ002095) ELISA kits (all from Shanghai Yuanju
Bio-Technology Co., Ltd.) migration inhibitory factor (MIF)
recombinant rabbit monoclonal antibody (cat. no. ET1703-89;
HUABIO), MIF rabbit monoclonal antibody (cat. no. A22623; ABclonal,
Inc.), phosphorylated (p)-NF-κB rabbit polyclonal antibody (cat.
no. WL02169), IL-6 (cat. no. WL02841), IL-1β (cat. no. WL02257) and
TNF-α rabbit polyclonal antibody (cat. no. WL01581; all from
Wanleibio Co., Ltd.), S-vision polymer secondary antibody for
immunohistochemistry (IHC; goat anti-rabbit; cat. no. G1302; Wuhan
Servicebio Technology Co., Ltd.), sodium citrate antigen retrieval
buffer (cat. no. G1219-1L; Wuhan Servicebio Technology Co., Ltd.),
anti-NF-κB p65 rabbit polyclonal antibody (cat. no. GB11997-100;
Wuhan Servicebio Technology Co., Ltd.), HRP-conjugated β-actin
recombinant rabbit monoclonal antibody (cat. no. PSH03-63; HUABIO)
and HRP-conjugated goat anti-rabbit IgG polyclonal antibody (cat.
no. HA1001; HUABIO).
Preparation of extract. Preparation of
MEAG freeze-dried powder
A total of 100 g BaiYangJie powder was added to
1,000 ml methanol and treated ultrasonically (800 W; 40 kHz;
25±3˚C) for 30 min, filtered and concentrated under reduced
pressure. The concentrated solution was stored overnight at -80˚C,
then freeze-dried to a powder (pro-4055/4085 freeze-dryer; Nanjing
Genscience Instrument and Equipment Co., Ltd.) and stored at 4˚C. A
total of 20 mg freeze-dried methanol extract powder was added to a
100 ml volumetric flask and methanol was added to make up the
volume to 100 ml. Then, the mixture was sonicated and finally
filtered through a 0.22 µm microporous membrane for further
analyses.
Preparation of the reference substance. A
total of ~1 mg resveratrol and pterostilbene were added to 100 ml
volumetric flasks. Methanol was added to dilute to scale and make
up a total volume of 100 ml to obtain a 10 µg/ml standard solution.
This solution was filtered through a 0.22 µm microporous membrane
for further analysis.
Establishment of Cis-AKI in a mouse
model
All animal experiments were approved by the
Institutional Ethics Review Committee of the Yunnan Branch of the
Institute of Medicinal Plant Development, Chinese Academy of
Medical Sciences (Jinghong, China; approval no. 20240213001). The
present study was strictly performed in accordance with the
guidelines of the National Research Council Guide for the Care and
Use of Laboratory Animals (13).
Male Kunming (KM) mice [weight: 20-22 g; aged 8 weeks; Sibefu
(Beijing) Biotechnology Co., Ltd.] were housed in SPF environments
with a controlled temperature (22±2˚C), relative humidity (50±5%),
atmospheric pressure (101±2 kPa) and a 12-h light/dark cycle,
before being allowed to acclimatise for 7 days, during which
animals were provided with ad libitum access to water and
standardised feed. A total of 30 KM mice were randomly allocated
into three experimental cohorts (n=10/group): i) Nor [0.5% sodium
carboxymethyl cellulose (CMC-Na), intragastrically], ii) Cis (Cis
13 mg/kg, intraperitoneally) and iii) MEAG (800 mg/kg,
intragastrically). The MEAG cohort received daily oral gavage for
10 consecutive days, while the mice in the Nor group were given
equivalent volumes of 0.5% CMC-Na solution (0.1 ml/10 g). On day 7,
Cis-induced nephropathy was established in the Cis through a single
intraperitoneal dose (13 mg/kg). Biological specimens (blood and
kidney tissues) were collected on day 10. Mice were anaesthetised
with 4% isoflurane in an induction chamber until loss of
consciousness and then maintained at a concentration of 1-3% using
a mask. Once the toe pinch reflex disappeared for >60 sec,
breathing became deep and slow and there was no limb contraction or
respiratory change upon strong pinching, the head was fixed and the
jugular vein was gently compressed to engorge. A capillary tube was
inserted into the inner canthus of the eye to collect blood, with
the volume reaching 20-30% of the total blood volume (0.5-0.8 ml
for 25 g mice). Immediately after blood collection, euthanasia was
performed (under deep anaesthesia, rapid cervical dislocation), To
clarify, cervical dislocation rather than exsanguination (blood
loss) was used for euthanasia as waiting for natural mortality due
to blood loss was prohibited. Mortality was determined when
breathing and heartbeat stopped for ≥2 min and the pupils dilated
and fixed. The left kidney was collected and fixed in 4%
paraformaldehyde at 24˚C for 24 h for histopathological
examination, while the right kidney was rapidly frozen in liquid
nitrogen and stored at -80˚C for molecular biological
experiments.
Metabolomics sample preparation
Serum aliquots (80 µl) were mixed with 320 µl
methanol/acetonitrile (ACN) solution (1:1; v/v) followed by 30 sec
vortex-mixing. The mixture underwent cryoprecipitation at -20˚C for
60 min. Subsequent centrifugation (4˚C; 16,100 x g; 15 min) yielded
supernatant that was lyophilised using the Bionoon-VAC1 vacuum
concentrator (Shanghai Bionoon Biotechnology Co., Ltd.) at 4˚C. The
lyophilised products were reconstituted in 160 µl ACN/water (1:1;
v/v) solution. Quality control (QC) samples were prepared by
pooling 10 µl aliquots from each reconstituted sample. Prior to
analysis, all specimens were filtered through 0.22 µm membrane
filters to remove particulate matter.
Ultra-performance liquid
chromatography-quadrupole time-of-flight mass spectrometry
(UPLC-Q-TOF-MS) analysis
Chromatographic separation was achieved using an
Acquity HSS T3 column (2.1x100.0 mm; 1.8 µm; Waters China Ltd.) on
a Shimadzu LC400 system (Shimadzu Scientific Instruments) coupled
with AB SCIEX X500B Q-TOF mass spectrometer (SCIEX) operating in
dual electrospray ionization mode. The eluent system comprised of
(A) 0.1% aqueous formic acid and (B) 0.1% formic acid in
acetonitrile. The following parameters were used: Flow rate, 0.3
ml/min; column oven, 35˚C; injection volume, 5 µl; nitrogen gas
temperature, 350˚C; and nitrogen nebuliser flow rate, 10 l/min.
Mass detection employed information-dependent acquisition mode with
the following settings: Primary MS spectrometry scan, m/z
100-1,200; and secondary tandem MS scan, m/z 50-1,200. The top 15
intense ions were selected for fragmentation.
For the MEAG samples, the mobile phase gradient was
0-2 min, 5% B; 2-5 min, 5-20% B; 5-8 min, 20-40% B; 8-10 min,
40-60% B; 10-15 min, 60-100% B; 15-19 min, 100% B; 19-20 min,
100-5% B; and 20-23 min, 5% B. Nitrogen gas was used in each gas
path and ionisation voltage was 5,500 V. Sprayer pressure was 60
psi, the auxiliary heating gas was 60 psi, the air curtain gas was
35 psi, the cone hole voltage was 100 V and the collision energy
was 10 V.
For the metabolomics samples, the mobile phase
gradient was 0-2 min, 5% B; 2-8 min, 5-50% B; 8-12 min, 50-80% B;
12-14 min, 80-95% B; 14-17 min, 95% B; 17-18 min, 95-5% B; and
18-21 min, 5% B. Nitrogen gas was used in each gas path. The
ionisation voltage was 4,500 V, the sprayer pressure was 60 psi,
the auxiliary heating gas was 60 psi, the air curtain gas was 35
psi, the cone hole voltage was 80 V and the collision energy was 20
V.
Network pharmacology and molecular
docking
Active components were initially screened using
SwissADME (http://www.swissadme.ch) with dual
criteria: Oral bioavailability ≥30% and drug-likeness. Potential
therapeutic targets were identified through SwissTargetPrediction
(swisstargetprediction.ch). Cis-AKI
targets were systematically retrieved from five major databases:
Online Mendelian Inheritance in Man (OMIM; https://omim.org/), DrugBank (version 5.1.10;
https://go.drugbank.com/), GeneCards (version
5.22; https://www.genecards.org/), Therapeutic
Target Database (TTD; https://db.idrblab.net/ttd/; accessed on October 31,
2023) and DisGeNET (version 24.0; https://www.disgenet.org/). The targets shared by MEAG
components and Cis-AKI were identified using the Venn analysis
module in SRplot (bioinformatics.com.cn).
Protein-protein interaction (PPI) networks were
constructed using Search Tool for the Retrieval of Interacting
Genes/Proteins (STRING; version 11.5; https://cn.string-db.org/), with species restricted to
Homo sapiens and a confidence cutoff >0.4. Following
network visualisation in Cytoscape (14) (version 3.7.1), the ‘CytoNCA’ plugin
(version 2.4.6) was used to prioritise nodes based on
multidimensional centrality metrics: Degree, betweenness and
closeness. A network of 12 metabolites was constructed using the
Search Tool For Interactions Of Chemicals (https://stitch-db.org/) and Cytoscape to represent the
metabolic module regulated by MEAG. The consensus targets underwent
comprehensive functional annotation using Kyoto Encyclopedia of
Genes and Genomes (KEGG) (15,16)
pathways and Gene Ontology (GO) enrichment analyses in Metascape
(metascape.org), with organism specification
restricted to Homo sapiens. Based on the number of gene
enrichments, diagrams were drawn using SRplot.
Metabolomics data analyses
Original data for each sample were processed to
obtain peak alignment, peak extraction, peak comparison and
standardisation using MS DIAL software (version 4.9.221218) as
previously described (17). The
online MetaboAnalyst (version 6.0; https://www.metaboanalyst.ca/) database was used for
principal component analysis (PCA) and orthogonal partial least
squares discriminant analysis (OPLS-DA). By analysing differences
between the Nor group and Cis group and between the Cis group and
MEAG group, differential metabolites [variable importance in the
projection (VIP)>1; P<0.05] were identified and imported into
MetaboAnalyst for enrichment and metabolic pathway analyses.
Renal function tests and histological
examination
Serum creatinine (Scr) and blood urea nitrogen (BUN)
concentrations were quantified using the SMT-120VP clinical
chemistry system (Chengdu Seamaty Technology Co., Ltd.) with a
seven-parameter renal function assay kit (cat. no. AW03076; Chengdu
Seamaty Technology Co., Ltd.), according to the manufacturer's
protocol.
Kidney specimens were fixed in 4% paraformaldehyde
at 24˚C for 24 h, followed by sequential dehydration at room
temperature and paraffin embedding at 60˚C for 2 h. Subsequently, 4
µm microtome sections were prepared. A total of three tissue slices
per sample were deparaffinized with xylene at room temperature for
15 min twice, rehydrated through a graded ethanol series at room
temperature and rinsed with distilled water. For H&E staining,
the sections were stained with haematoxylin at room temperature for
5 min, rinsed with running tap water, differentiated, blued and
then counterstained with eosin at room temperature for 2 min. For
Masson's trichrome staining, the sections were stained according to
the manufacturer's protocol. Briefly, the sections were stained
with haematoxylin at room temperature for 5 min, stained with
Masson's ponceau acid fuchsin solution at room temperature for 5
min, differentiated with phosphomolybdic acid solution at room
temperature for 2 min and counterstained with aniline blue solution
at room temperature for 2 min. After staining, all sections were
dehydrated, cleared and mounted with neutral resin. Renal
morphological alterations were visualised using an Olympus BX53
bright-field microscope at x200 and x400 magnification (Olympus
Corporation). The glomerular architecture, proximal convoluted
tubule morphology, basement membrane characteristics of glomeruli
and collagen deposition patterns were observed.
ELISA
Circulating concentrations of TNF-α, IL-1β and IL-6
in mouse serum were quantified using commercially available ELISA
kits for mice according to the manufacturer's protocols.
IHC
Following dewaxing, the sections were rehydrated
through a graded ethanol series and rinsed with PBS. Antigen
retrieval was performed by boiling samples in sodium citrate
antigen retrieval buffer at 100˚C for 10 min. Subsequently, tissues
were treated with a quenching reagent (3% hydrogen peroxide
solution) at room temperature for 5 min to block endogenous
peroxidase activity, followed by blocking with 10% goat serum
(Wuhan Servicebio Technology Co., Ltd.) for 60 min at room
temperature. The goat serum was discarded and KIM-1 rabbit
polyclonal antibody (1:300), NGAL rabbit polyclonal antibody
(1:200) and MIF rabbit monoclonal antibody (1:800) were applied to
the sections and incubated overnight (~12 h) at 4˚C. Next,
secondary antibody conjugation was performed using S-vision polymer
secondary antibody for IHC at room temperature for 60 min before
the sections were washed three times with PBS. Signals were
visualised using diaminobenzidine at room temperature for 3 min,
counterstained with haematoxylin at room temperature for 1 min,
dehydrated through a graded ethanol series, cleared with xylene and
mounted. Tissues were viewed using standard bright-field
illumination.
Western blotting
Renal tissues were homogenised in lysis buffer (cat.
no. CW2333; CoWin Biosciences, Inc.) supplemented with 1%
protease/phosphatase inhibitors, followed by a 1 h incubation to
ensure complete lysis and subsequent centrifugation (16,100 x g; 20
min; 4˚C). Protein quantification was performed using a BCA protein
detection kit (cat. no. CW0014; CoWin Biosciences, Inc.). To
resolve proteins, a 10% SDS-PAGE gel was used and 30 µg of protein
was loaded per lane before electrophoresis; the resolved proteins
were transferred to PVDF membranes. Membranes were blocked with 5%
non-fat milk/TBST (at 25˚C for 2 h; 0.05% Tween-20) prior to
overnight incubation (4˚C) with anti-MIF (1:1,000), anti-p-NF-κB
(1:2,000), NF-κB (1:1,000) anti-IL-6 (1:2,000), anti-IL-1β
(1:2,000) and anti-TNF-α (1:2,000). After three TBST washes,
membranes were incubated with an HRP-conjugated secondary antibody
(25˚C for 2 h; 1:50,000). with β-actin as the internal reference.
Signals were visualised with ECL reagents (New Cell & Molecular
Biotech Co., Ltd.) and band intensities were quantified using an
imaging system from Tanon Science and Technology Co., Ltd. with
Image ProPlus (version 5.0; Media Cybernetics, Inc.; https://www.mediacy.com/).
Statistical analysis
Statistical analysis was performed utilising
GraphPad Prism (version 9.0; Dotmatics) software. Data were
compared using a one-way ANOVA followed by Tukey's HSD post hoc
test. A total of three independent biological replicates were
performed for each experiment. Results are presented as the mean ±
SD. P<0.05 was considered to indicate a statistically
significant difference.
Results
Characterisation of chemical
constituents of MEAG
Based on UPLC-Q-TOF-MS analysis, combined with
database searches, reference compounds and related literature, 33
chemical components were identified in MEAG using multilevel MS
information (18-24).
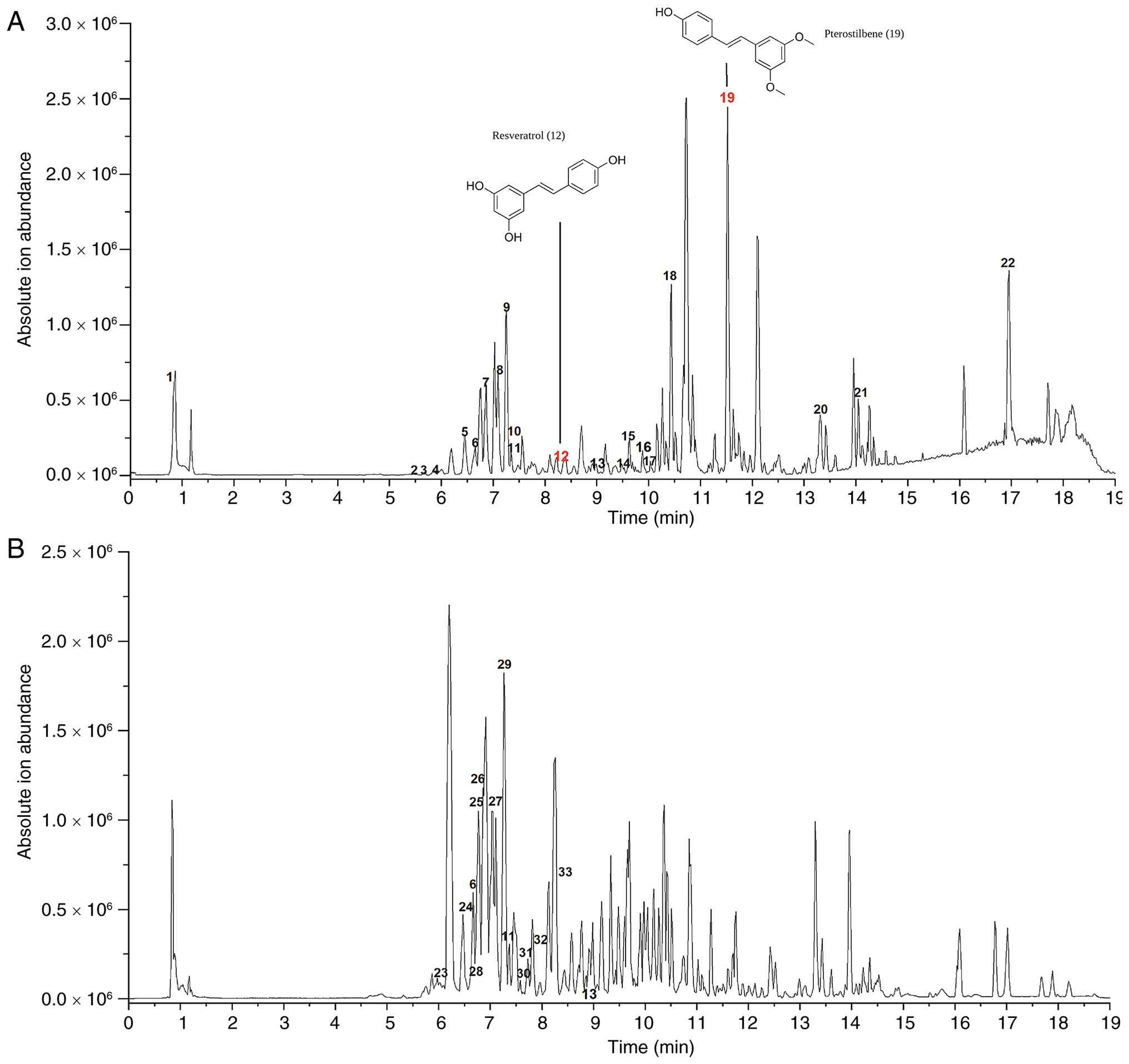
The chemical components of the MEAG are listed in Table I and the base peak chromatograms of
the positive and negative ion modes are shown in Fig. 1A and B.
 | Table ICharacterisation of main components
of MEAG in positive and negative ion modes. |
Table I
Characterisation of main components
of MEAG in positive and negative ion modes.
| No. | Compound name | RT, min | Measured value,
m/z | Molecular
formula | MS/MS | Ion species | Error, ppm |
|---|
| 1 | Betaine | 0.826 | 118.0860 |
C5H11NO2 | 58.0649,
118.0862 | (M+
H)+ | -2.2 |
| 2 | Naringin | 5.411 | 603.1691 |
C27H32O14 | 603.1686, 441.1173,
409.0927, 277.0471 | (M+
Na)+ | 1.1 |
| 3 | Verbenalin | 5.502 | 411.1266 |
C17H24O10 | 411.1272, 249.0751,
231.0647, 187.0754, 181.0507 | (M+
Na)+ | 1.0 |
| 4 | Riboflavin | 5.934 | 377.1459 |
C17H20N4O6 | 377.1465, 359.1345,
243.0879, 198.0663, 172.0869 | (M+
H)+ | 0.9 |
| 5 | Mauritianin | 6.439 | 763.2061 |
C33H40O19 | 763.2036, 617.1505,
477.1587, 331.1008 | (M+
Na)+ | 0.7 |
| 6 | Rutin | 6.653/6.665 |
611.1614/609.1451 |
C27H30O16 | 611.1623, 303.0502,
287.0548, 257.0437, 229.0945/609.1424, 300.0253, 271.0233,
255.0288, 178.9980, 151.0031 | (M+
H)+/(M- H)- | 1.2/-1.7 |
| 7 | Vicenin 2 | 6.899 | 617.1489 |
C27H30O15 | 617.1476, 471.0946,
455.1299, 331.1005 | (M+
Na)+ | 2.0 |
| 8 | Isoorientin | 7.111 | 471.0909 |
C21H20O11 | 471.0905, 309.0374,
185.0425, 147.0453 | (M+
Na)+ | 2.4 |
| 9 |
Kaempferol-4'-glucoside | 7.253 | 471.0908 |
C21H20O11 | 471.0894, 405.0513,
365.1428, 309.0380, 202.0515, 185.0427 | (M+
Na)+ | 2.2 |
| 10 | Nepitrin | 7.340 | 479.1193 |
C22H22O12 | 479.1555, 317.0659,
302.0427, 385.0400, 165.0197, 153.0188 | (M+
H)+ | 1.9 |
| 11 |
Isorhamnetin-3-O-glucoside | 7.340/7.366 |
501.1009/477.1029 |
C22H22O12 | 501.1001, 435.0493,
357.0457, 3338.0415, 261.0492, 185.0422/477.1020, 314.0415,
299.0191, 285.0395, 271.0236 | (M+
Na)+/(M- H)- | 1.1/-2.0 |
| 12 | Resveratrol | 8.255 | 229.0859 |
C14H12O3 | 229.0876, 211.0768,
183.0811, 193.0643, 109.0279 | (M+
H)+ | -0.1 |
| 13 | Quercetin | 9.074/9.086 |
303.0501/301.0350 |
C15H10O7 | 303.0506, 285.0398,
257.0450, 247.0602, 229.0500, 153.0191/301.0345, 273.0405,
245.0459, 178.9982, 151.0035, 121.0292 | (M+
H)+/(M- H)- | 0.6/-1.3 |
| 14 | Benzaldehyde | 9.505 | 107.0489 |
C7H6O | 107.0495,
77.0385 | (M+
H)+ | -2.2 |
| 15 | Lusianthridin | 9.631 | 243.1013 |
C15H14O3 | 243.1016, 228.0783,
215.1074, 211.0756, 197.0967, 181.0651 | (M+
H)+ | -1.1 |
| 16 | Kaempferol | 9.883 | 287.0547 |
C15H10O6 | 287.0550, 258.0535,
241.0505, 213.0556, 165.0191, 153.0189 | (M+
H)+ | -1.1 |
| 17 | Isorhamnetin | 10.028 | 317.0661 |
C16H12O7 | 317.0661, 302.0428,
285.0401, 229.0500, 153.0183 | (M+
H)+ | 1.6 |
| 18 | Batatasin III | 10.437 | 245.1163 |
C15H16O3 | 245.1169, 151.0749,
137.0595, 121.0646, 107.0490, 91.0540 | (M+
H)+ | -3.8 |
| 19 |
Trans-pterostilbene | 11.524 | 257.1160 |
C16H16O3 | 257.1169, 242.0931,
226.0979, 211.0748, 197.0955, 181.0643 | (M+
H)+ | -4.7 |
| 20 | Linoleic acid | 13.289 | 281.2471 |
C18H32O2 | 281.2490, 155.1437,
109.1016, 95.0858, 81.0703, 67.0545 | (M+
H)+ | -1.5 |
| 21 | E-resveratrol
trimethyl ether | 14.038 | 271.1320 |
C17H18O3 | 271.0324, 255.1024,
240.1141, 225.0909, 209.0964, 165.0698 | (M+
H)+ | -3.2 |
| 22 | Oleanonic acid | 16.950 | 455.3531 |
C30H46O3 | 455.3499, 381.3399,
269.2257, 238.1526, 81.0695 | (M+
H)+ | 2.5 |
| 23 |
4-hydroxybenzaldehyde | 6.082 | 121.0291 |
C7H6O2 | 121.0290, 92.0263,
75.0232 | (M-
H)- | -3.3 |
| 24 | p-Coumaric
acid | 6.630 | 163.0398 |
C9H8O3 | 163.0392, 119.0503,
93.0344 | (M-
H)- | -1.7 |
| 25 | Vitexin | 6.752 | 431.0987 |
C21H20O10 | 431.0973, 341.0667,
311.0553, 283.0606, 269.0455 | (M-
H)- | 0.8 |
| 26 | Quercetin
3-galactoside | 6.857 | 463.0870 |
C21H20O12 | 463.0866, 300.0261,
271.0235, 255.0283, 243.0290, 151.0033 | (M-
H)- | -2.6 |
| 27 |
Kaempferol-3-O-rutinoside | 7.049 | 593.1497 |
C27H30O15 | 593.1489, 447.0949,
327.0507, 285.0384, 255.0284, 151.0035 | (M-
H)- | -2.5 |
| 28 | Pinoresinol | 7.208 | 357.1343 |
C20H22O6 | 357.1247, 160.0532,
151.0405, 136.0167, 122.0383, 92.0261 | (M-
H)- | -0.2 |
| 29 | Kaempferol
3-O-glucoside | 7.261 | 447.0919 |
C21H20O11 | 447.0923, 284.0316,
255.0285, 227.0336, 163.0612 | (M-
H)- | -3.1 |
| 30 |
Kaempferol-3-O-α-L-arabinoside | 7.543 | 417.0824 |
C20H18O10 | 417.0818, 284.0327,
255.0300, 227.0352 | (M-
H)- | -0.8 |
| 31 | Azelaic acid | 7.723 | 187.0970 |
C9H16O4 | 187.0962, 169.0864,
143.1062, 125.0970, 97.0657, 57.0345 | (M-
H)- | -3.1 |
| 32 |
Kaempferol-3-O-α-L-rhamnoside | 8.044 | 431.0980 |
C21H20O10 | 431.0978, 284.0319,
269.0460, 255.0293, 227.0342 | (M-
H)- | -0.9 |
| 33 | Coelonin | 8.290 | 241.0873 |
C15H14O3 | 431.0978, 284.0319,
269.0460, 255.0293, 227.0342 | (M-
H)- | 1.2 |
Network pharmacology
A total of 14 active ingredients (Table SI) and 348 active ingredient
targets were screened using the Swiss ADME and Swiss Target
Prediction databases. A total of 2,514 non-redundant disease
targets were retrieved from OMIM, DrugBank, GeneCards, TTD and
DisGeNET. A total of 210 intersecting targets were obtained by
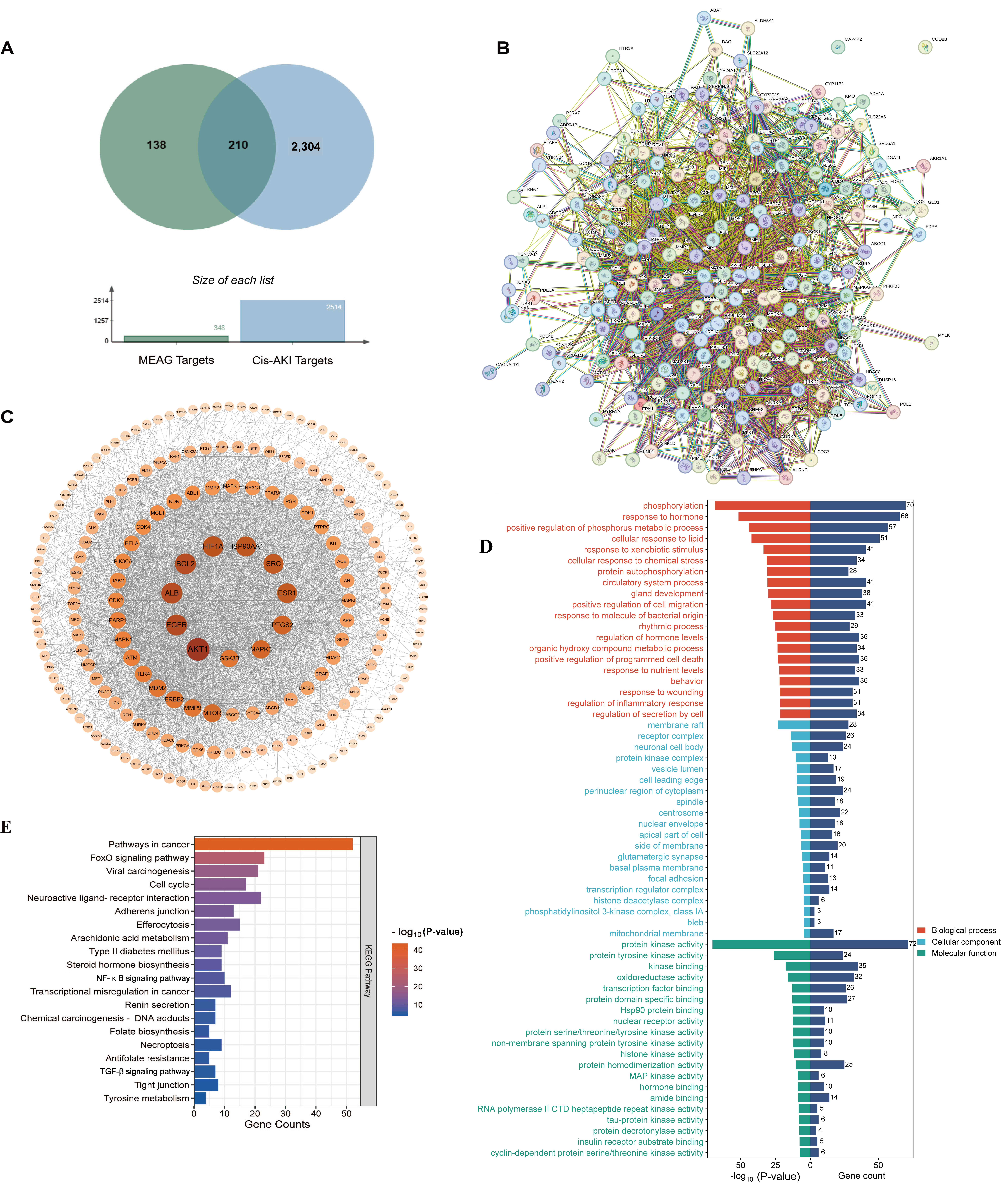
intersecting the drug and disease targets (Fig. 2A).
The PPI network reflects protein interactions; 210
intersecting targets formed a network with 208 nodes and 2,861
edges. Subsequently, the PPI network was visualised using Cytoscape
and the topology analysis was performed using the ‘CytoNCA’
plug-in. The top 15 nodes of the network were sorted by betweenness
centrality, closeness centrality and degree centrality values and
the hub targets were the intersection of these three values. These
hub targets were AKT1, EGFR, ALB, BCL2, HIF1A, HSP90AA1, SRC, ESR1,
PTGS2, MAPK3 and GSK3B (Fig. 2B
and C; Table SII).
Based on GO functional enrichment, biological
process enrichment involved ‘cellular response to lipid’ and
‘cellular response to chemical stress’, ‘regulation of inflammatory
response’ and ‘positive regulation of programmed cell death’.
Cellular component enrichment included ‘perinuclear region of
cytoplasm’, ‘centrosome’ and ‘basal plasma membrane’. Molecular
function enrichment involved ‘protein kinase activity’ and ‘protein
tyrosine kinase activity’ (Fig.
2D). Collectively, MEAG may have contributed to the treatment
of Cis-AKI through the regulation of numerous components, targets
and pathways.
The target set was imported into the Metascape
database for KEGG pathway and biological process enrichment
analysis. There were 184 KEGG pathways associated with Cis-AKI
therapy in mice. In addition to the cancer pathway with the largest
number of annotation genes, the target set was also closely
associated with ‘FoxO signalling pathway’, arachidonic acid
metabolism’, ‘NF-κB signalling pathway’ and ‘TGF-β signalling
pathway’ (Fig. 2E).
Serum metabolomics. Multivariate
statistical analyses
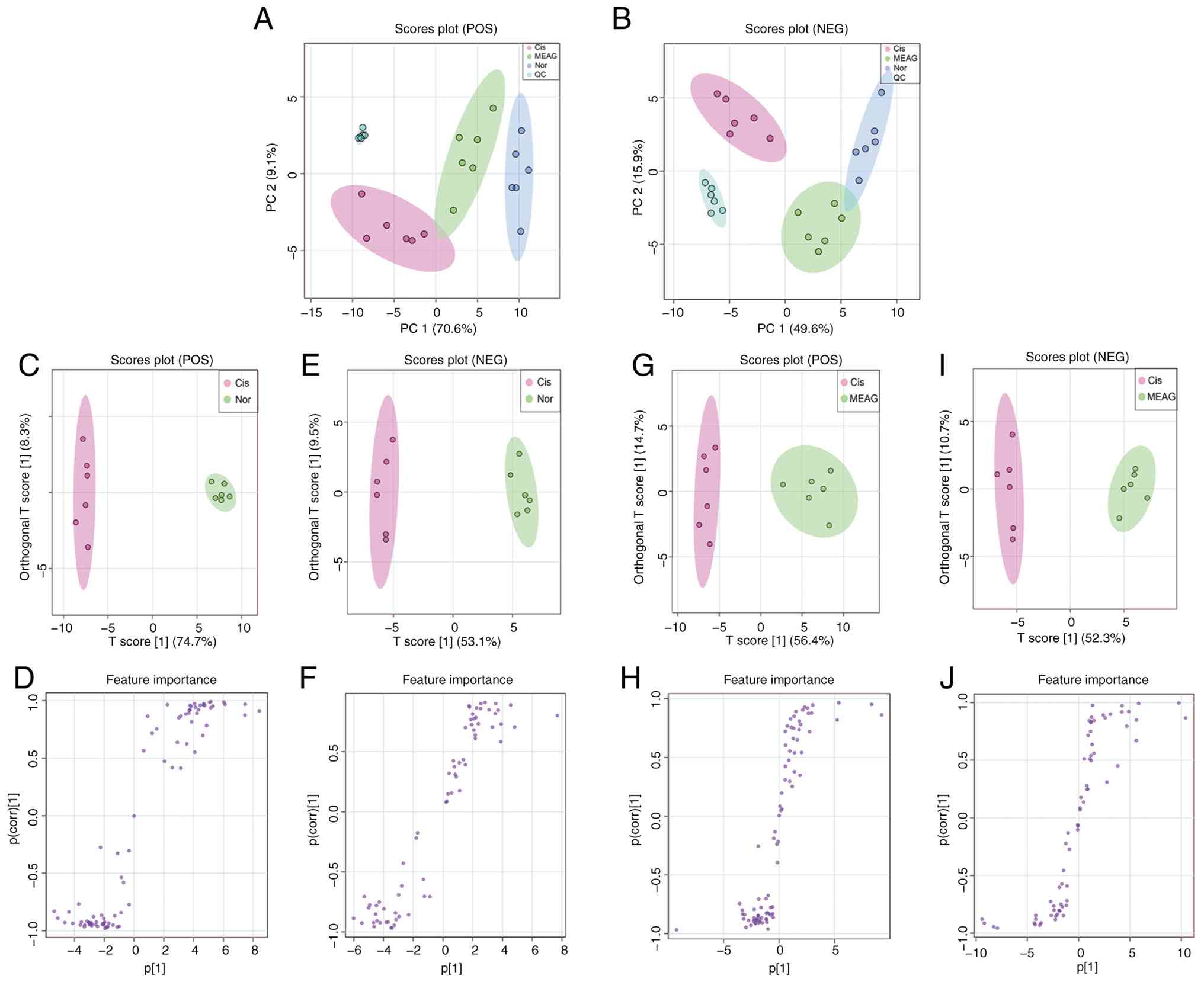
General distribution of samples was examined by PCA,
with results showing the QC samples clustered together, indicating
that the instrument was stable (Fig.
3A and B). The three groups of
test samples were clustered, which indicated similarities within
the data. There were notable metabolic differences among the three
groups. The Nor group was clearly separated from the Cis group and
the MEAG group exhibited a downward trend after treatment,
indicating that MEAG exerted an ameliorative effect on metabolic
disorders in Cis-AKI model mice (Fig.
3A and B).
To establish an association model between the
expression of metabolites and sample categories and to screen the
differential metabolites, OPLS-DA in positive and negative ion mode
was used (Fig. 3C, E, G and
I) and the samples in the Nor and
Cis groups and Cis and MEAG groups were all significantly
separated.
The S-plot diagram of positive and negative ions
showed differences in metabolites between the Nor and Cis groups
and between the Cis and MEAG groups (Fig. 3D, F, H and
J). VIP, fold change (FC) and
P-values were then used to identify characteristic differentially
expressed metabolites among the three groups. Thus, substances with
log2 (FC) >1, VIP >1 and P<0.05 were
potentially upregulated differential metabolites and substances
with log2 (FC)<-1, VIP>1 and P<0.05 were
potential downregulated differential metabolites. In the positive
ion mode, six endogenous differential metabolites were identified
between Nor and Cis groups, and Cis and MEAG groups, including
L-phenylalanine, L-carnitine, creatine, phosphocholine,
lysophosphatidylethanolamine (LysoPE 16:0/0:0, the notation
16:0/0:0 indicates the fatty acyl chain composition, with a
16-carbon saturated fatty acyl chain and no second fatty acyl
chain) and stearoyl-L-carnitine. In negative ion mode, eight
endogenous differential metabolites were identified between the Nor
and Cis groups and the Cis and MEAG groups, including L-tryptophan,
citric acid, ascorbic acid, L-phenylalanine, hyocholic acid,
docosahexaenoic acid, L-tyrosine and L-pyroglutamic acid. All
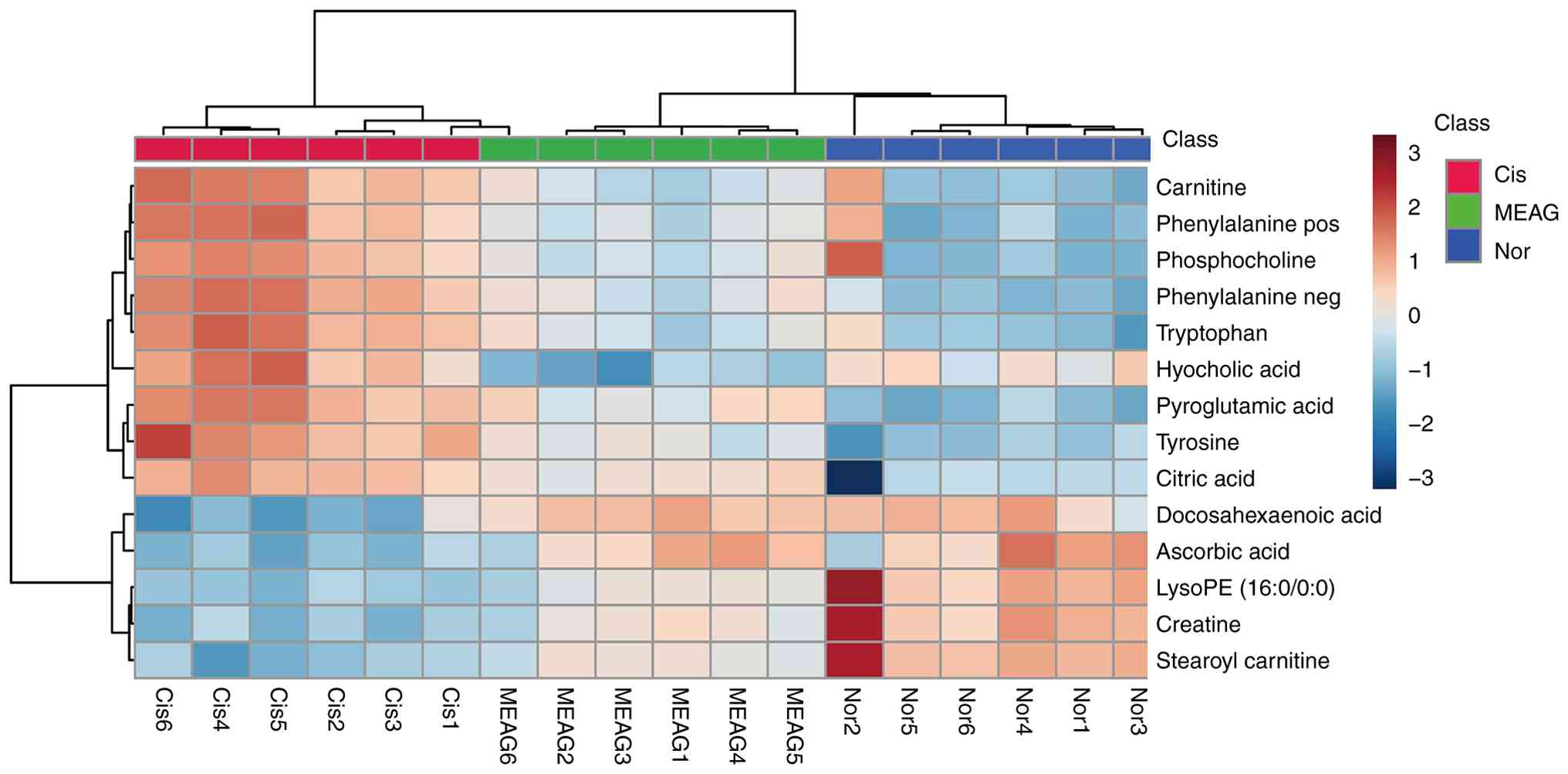
differential metabolites in different groups were visualised as a
heatmap, in which blue to red indicated an increase in the
abundance of differential metabolites (Fig. 4). Table SII and Fig. 5 provide detailed information
regarding each differential metabolite and the changes in the
relative contents of potentially different metabolites.
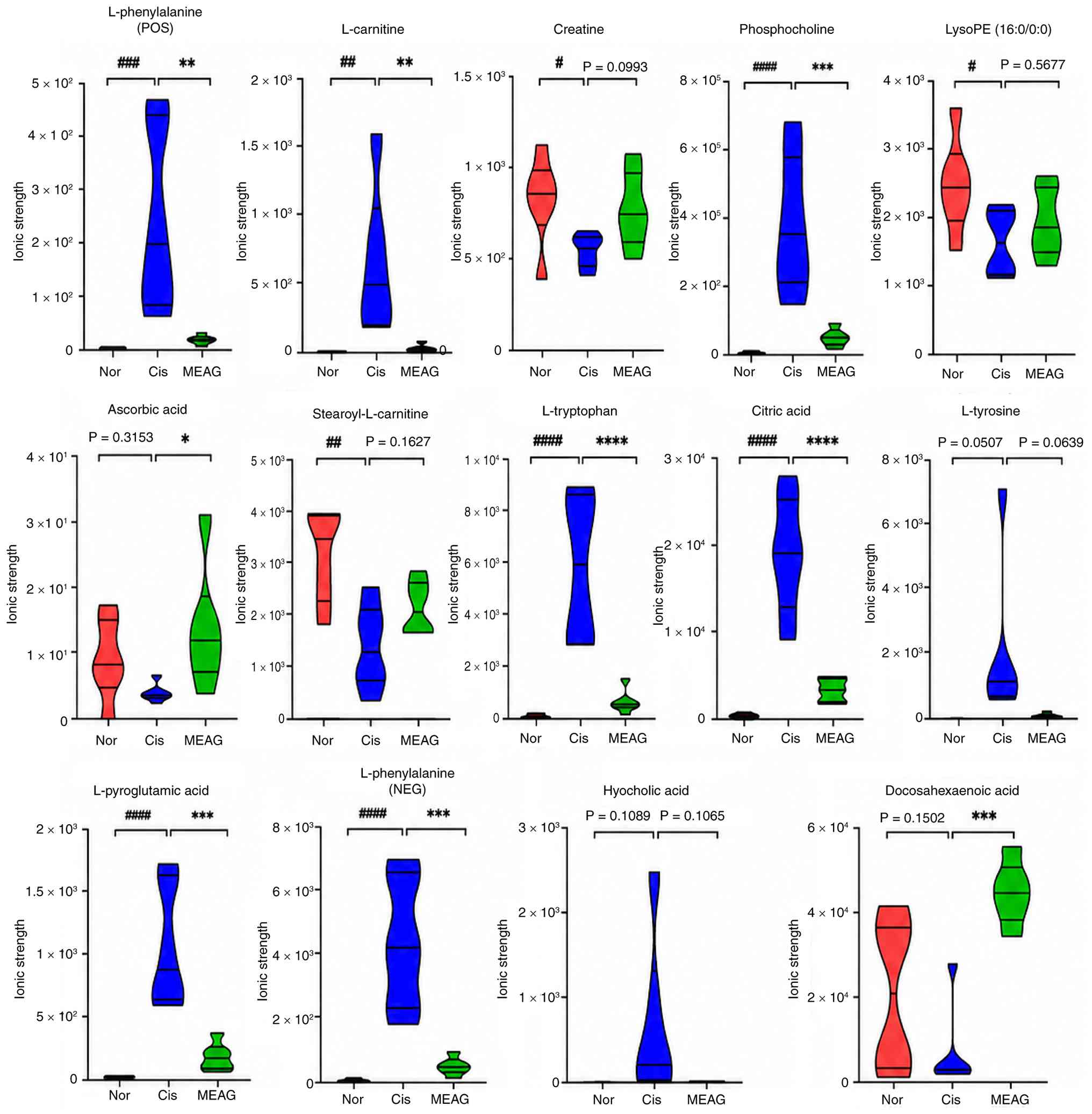 | Figure 5Relative content changes of potential
metabolites. ####P<0.0001, ##P<0.01,
#P<0.05; compared with the Nor group,
****P<0.0001, ***P<0.001,
**P<0.01 and *P<0.05 compared with the
Cis group. LysoPE, lysophosphatidylethanolamine; POS, positive;
NEG, negative; Cis, cisplatin; MEAG, methanolic extract of
BaiYangJie; Nor, normal control group. |
Construction and analysis of metabolic
module
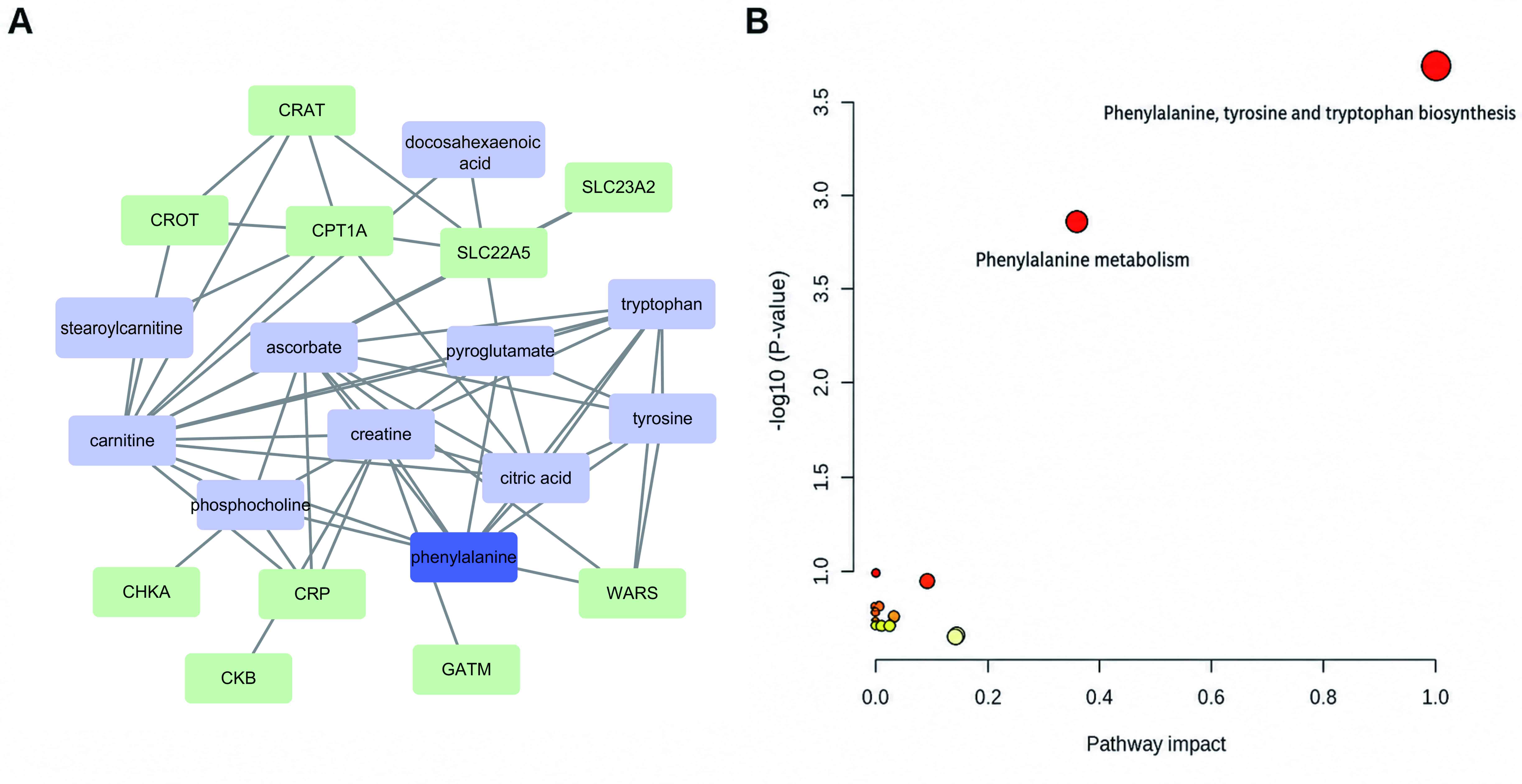
Metabolite and gene regulatory network is shown in
Fig. 6A. Through topological
analysis of metabolic modules, phenylalanine was identified as the
hub with the highest centrality. Then, MetaboAnalyst was used to
analyse the metabolic pathway enrichment of the 12 metabolites. As
shown in Fig. 6B, 14 pathways
affecting the metabolism of mice with Cis-AKI were obtained from 13
different metabolites. Among them, the potential metabolic pathways
with path influence >0.1 were ‘phenylalanine metabolism’ and
‘phenylalanine, tyrosine and tryptophan biosynthesis’. Based on the
results of metabolite network and pathway analyses, phenylalanine
metabolism may be a key metabolic pathway regulated by MEAG in the
treatment of Cis-AKI.
Comprehensive analysis of network
pharmacology and serum metabolomics
Associations between metabolic pathways and their
targets were next assessed. To comprehensively explore the
mechanism of action of MEAG in treating Cis nephrotoxicity, the
targets of phenylalanine metabolism were assessed using KEGG
(25) and intersected with drug
targets and disease targets, integrated the results of serum
metabolomics and network pharmacology analysis and subsequently
identified an intersection target (MIF).
Among the aforementioned key signalling pathways,
NF-κB serves a central role in cellular inflammatory and immune
responses (26). A previous study
suggested that, in a Cis-AKI mouse model, injured renal tubular
epithelial cells released inflammatory cytokines, activated the
MIF/NF-κB pathway and triggered a cascade of inflammatory
responses, thereby exacerbating renal tubular injury (27). In addition, phenylalanine
metabolism is also associated with inflammation. Therefore, the
ability of MEAG to regulate the MIF/NF-κB signalling pathway and
associated inflammatory factors was evaluated. A schematic diagram
of the regulatory pathway is shown in Fig. 7.
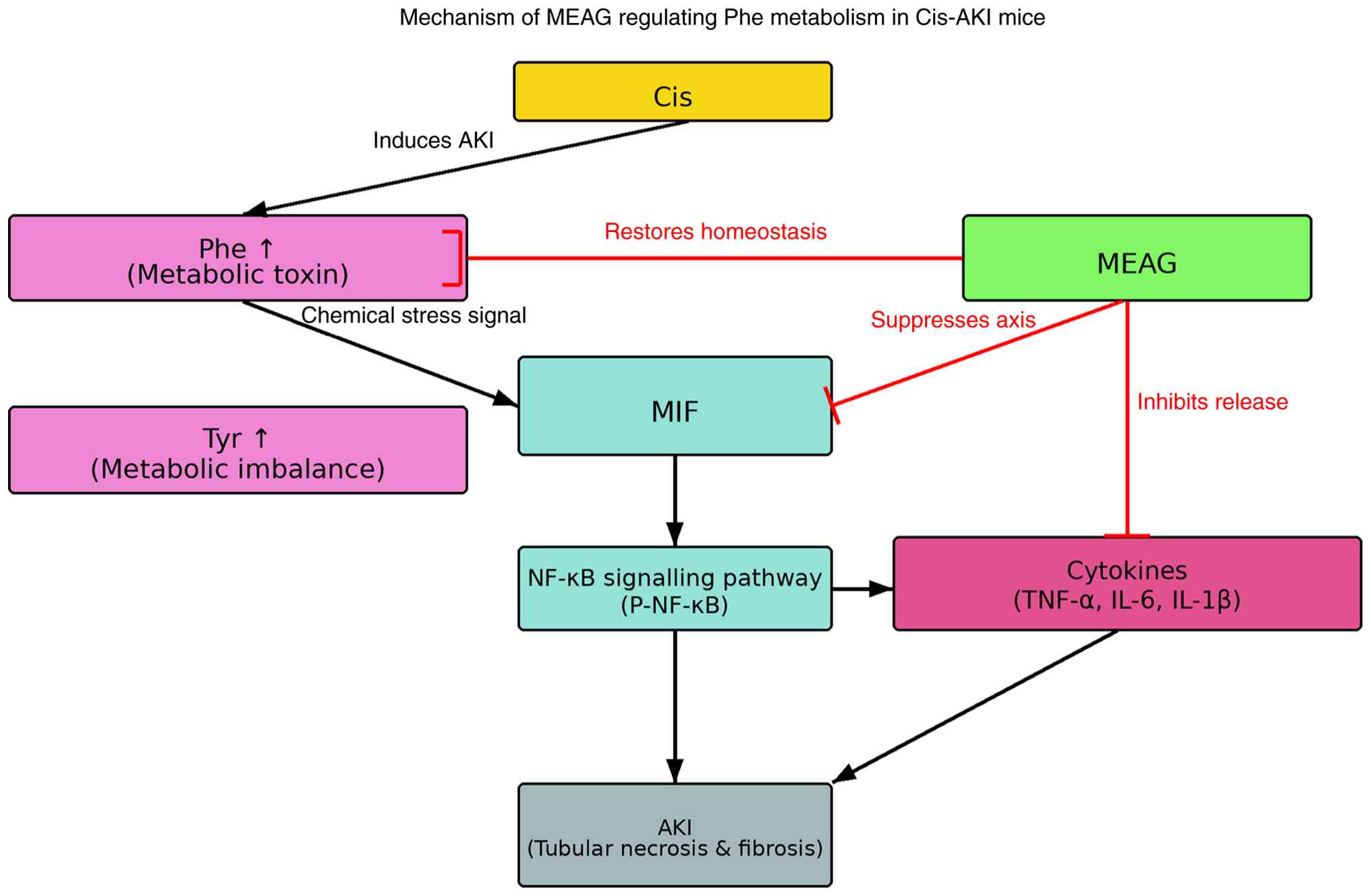 | Figure 7Schematic diagram of MEAG regulating
phenylalanine metabolic flux. MEAG, methanolic extract of
BaiYangJie; Cis, cisplatin; Cis-AKI, cisplatin-induced acute kidney
injury; Phe, phenylalanine; Tyr, tyrosine; MIF, macrophage
migration inhibitory factor; NF-κB, nuclear factor κB; P-NF-κB,
phosphorylated NF-κB; TNF-α, tumor necrosis factor α; IL-6,
interleukin-6; IL-1β, interleukin-1β; AKI, acute kidney injury. |
MEAG ameliorates Cis-AKI in mice
After intraperitoneal injection of Cis, mice
exhibited clear nephrotoxicity, including weight loss and a
significantly increased renal index (P<0.0001; Fig. 8A and B). The levels of Scr (P<0.0001) and
BUN (P<0.0001) also increased significantly (Fig. 8C and D). H&E staining of renal tissue
showed that intraperitoneal injection of Cis caused serious damage
to renal tubules and glomeruli, with renal tubular dilatation,
renal tubular epithelial cell shedding, gelatinous casts,
glomerular atrophy and basement membrane thickening (Fig. 8E). Quantitative analysis further
showed that the tubular injury score was significantly increased in
the Cis group compared with that in the Nor group (P<0.0001;
Fig. 8F). Masson's trichrome
staining revealed severe fibrosis in renal tissue (Fig. 8G). Consistently, the collagen
volume fraction was significantly increased in the Cis group
compared with that in the Nor group (P<0.0001; Fig. 8H). MEAG treatment significantly
ameliorated the loss of renal function of mice and significantly
reduced the kidney index (P<0.001; Fig. 8B) and levels of Scr (P<0.0001)
and BUN (P<0.0001), which were increased by Cis injection
(Fig. 8C and D). Simultaneously, MEAG ameliorated renal
tubular damage and reduced the degree of renal tissue fibrosis
(Fig. 8E-H).
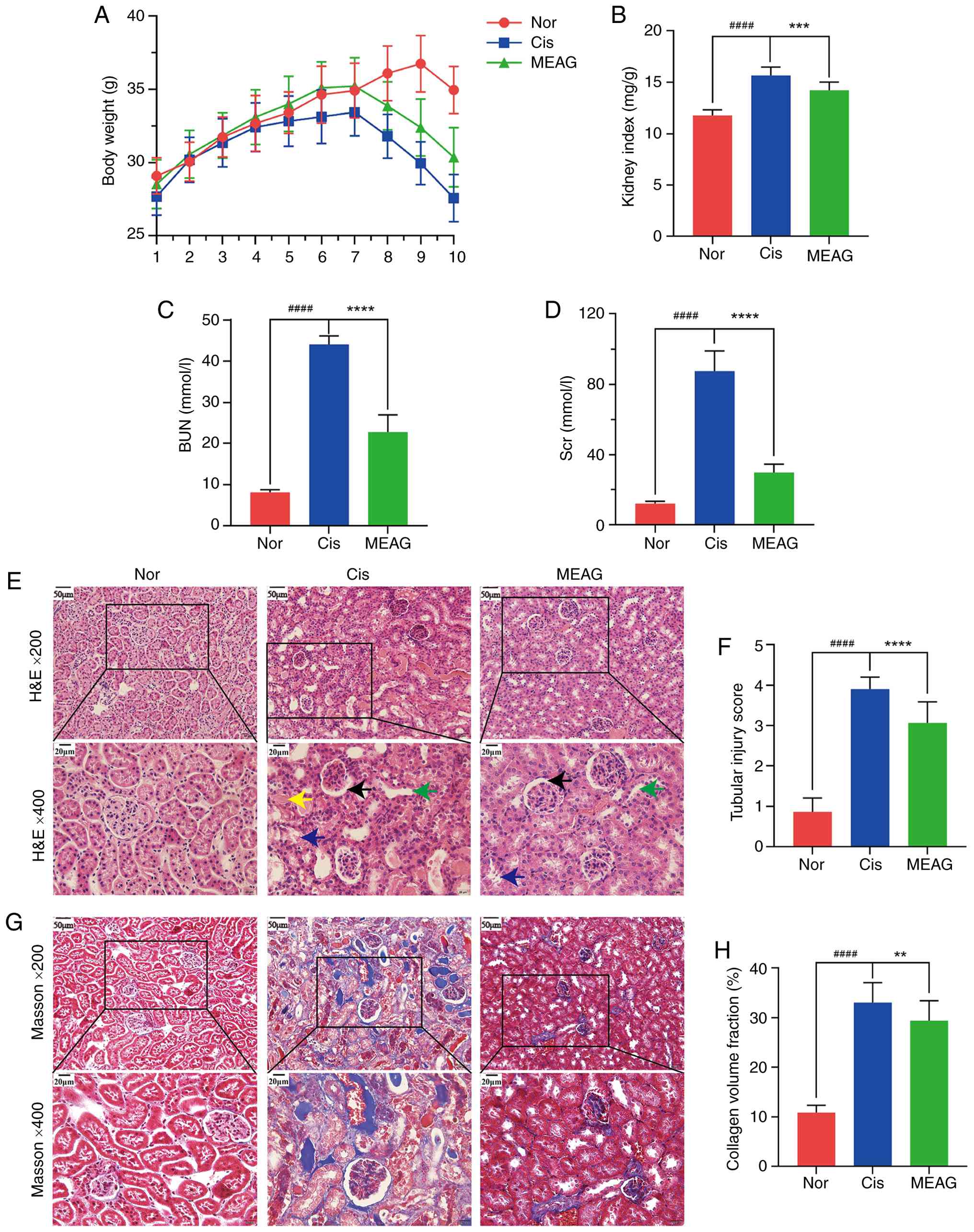 | Figure 8MEAG treatment ameliorated Cis-AKI in
mice. (A) Effect of MEAG on body weight change in mice with
Cis-AKI. (B) Effect of MEAG on kidney index in mice with Cis-AKI.
(C) Effect of MEAG on BUN levels in mice with Cis-AKI. (D) Effect
of MEAG on Scr levels in mice with Cis-AKI. (E) Representative
H&E-stained kidney sections from the Nor, Cis, and MEAG groups,
shown at x200 magnification with corresponding higher-magnification
views at x400. Yellow arrows indicate rubber-like tubular casts,
blue arrows indicate shedding of renal tubular epithelial cells,
green arrows indicate renal tubular dilatation, and black arrows
indicate glomerular atrophy and basement membrane thickening. (F)
Tubular injury score in each group. (G) Representative
Masson-stained kidney sections from the Nor, Cis and MEAG groups,
shown at x200 magnification with corresponding higher-magnification
views at x400. (H) Collagen volume fraction in each group. Data in
panels (A-D), (F) and (H) are presented as mean ± SD (n=6 mice per
group). ####P<0.0001 compared with the Nor group;
**P<0.01, ***P<0.001 and
****P<0.0001 compared with the Cis group. Scale bars,
50 µm for x200 images and 20 µm for x400 images. Nor, normal
control group; Cis, cisplatin; MEAG, methanolic extract of
BaiYangJie; AKI, acute kidney injury; Scr, serum creatinine; BUN,
blood urea nitrogen. |
KIM-1 and NGAL are biomarkers of AKI (28). The expression and distribution of
KIM-1 and NGAL in renal tissue were examined. IHC staining showed
that the expression of KIM-1 (P<0.0001) and NGAL (P<0.0001)
in mouse renal tissue significantly increased after Cis injection
and were significantly reduced by MEAG treatment (P<0.0001;
Fig. 9A-D).
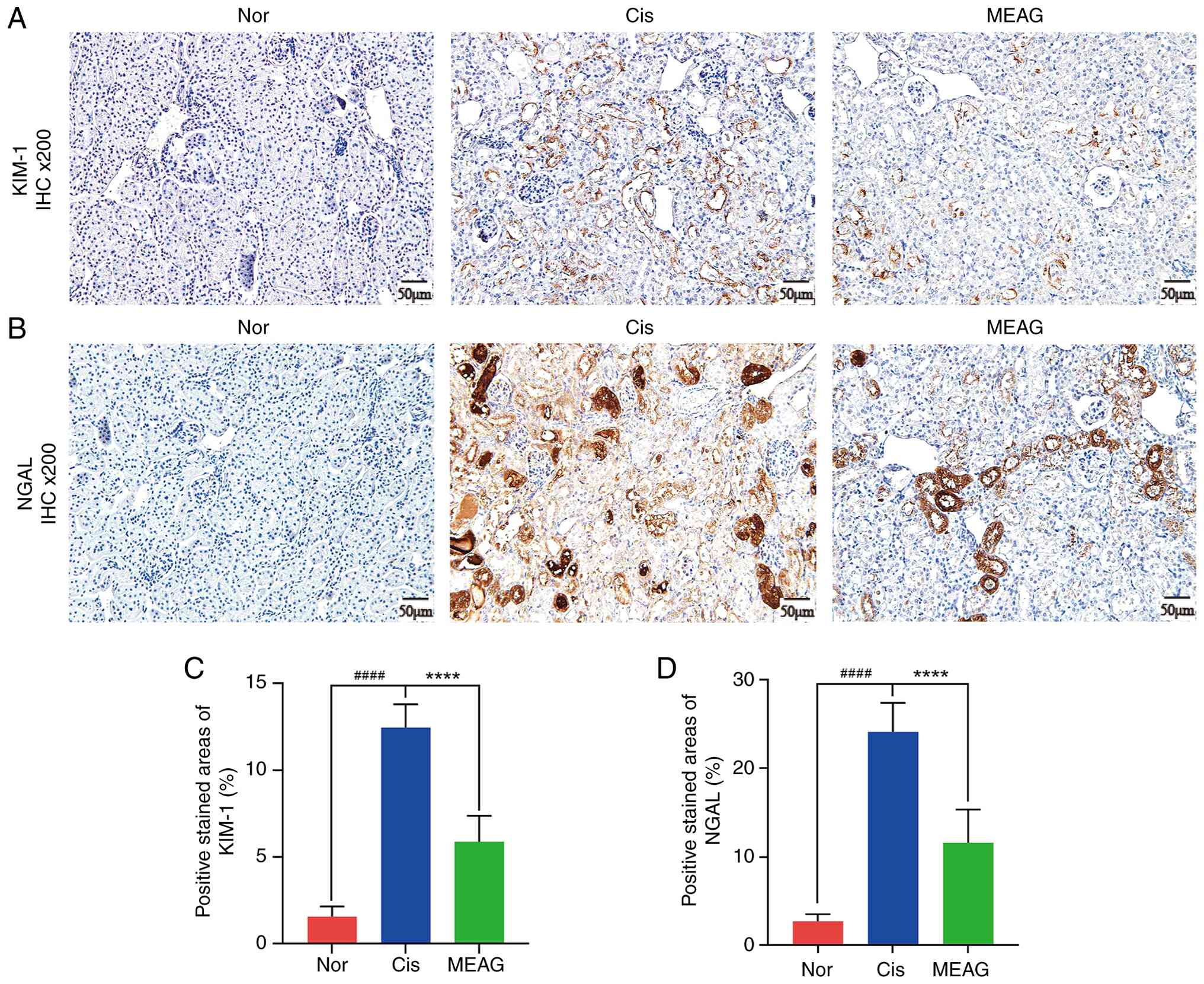 | Figure 9Upregulation of KIM-1 and NGAL
expression induced by cisplatin was reduced by MEAG. (A)
Representative IHC images of KIM-1 expression in kidney tissues
from the Nor, Cis and MEAG groups. (B) Representative IHC images of
NGAL expression in kidney tissues from the Nor, Cis and MEAG
groups. (C) Quantitative analysis of the positive stained area of
KIM-1 in kidney tissues. (D) Quantitative analysis of the positive
stained area of NGAL in kidney tissues. For panels (A) and (B),
magnification x200 and scale bar, 50 µm. For panels (C) and (D),
data are presented as mean ± SD (n=3 per group).
####P<0.0001, compared with the Nor group and
****P<0.0001, compared with the Cis group. Nor,
normal control group; Cis, cisplatin; MEAG, methanolic extract of
BaiYangJie; IHC, immunohistochemistry; NGAL, neutrophil
gelatinase-associated lipocalin; KIM-1, kidney injury molecule
1. |
MEAG ameliorates inflammatory
responses in mice treated with Cis
Proinflammatory cytokine expression in mice after
MEAG treatment was analysed. ELISA data showed that MEAG
significantly reduced the increase in IL-1β (P<0.0001), IL-6
(P<0.0001) and TNFα (P<0.0001) levels in the serum of mice
initially induced by Cis (Fig.
10A-C). Furthermore, western blotting showed that Cis
significantly induced the expression of the inflammatory factors
Il-1β (P<0.0001), IL-6 (P<0.0001) and TNF-α (P<0.0001) and
that this induction was attenuated by MEAG treatment (Fig. 10F and H-J). These findings indicated that MEAG
reduced the inflammatory response of mice following Cis
treatment.
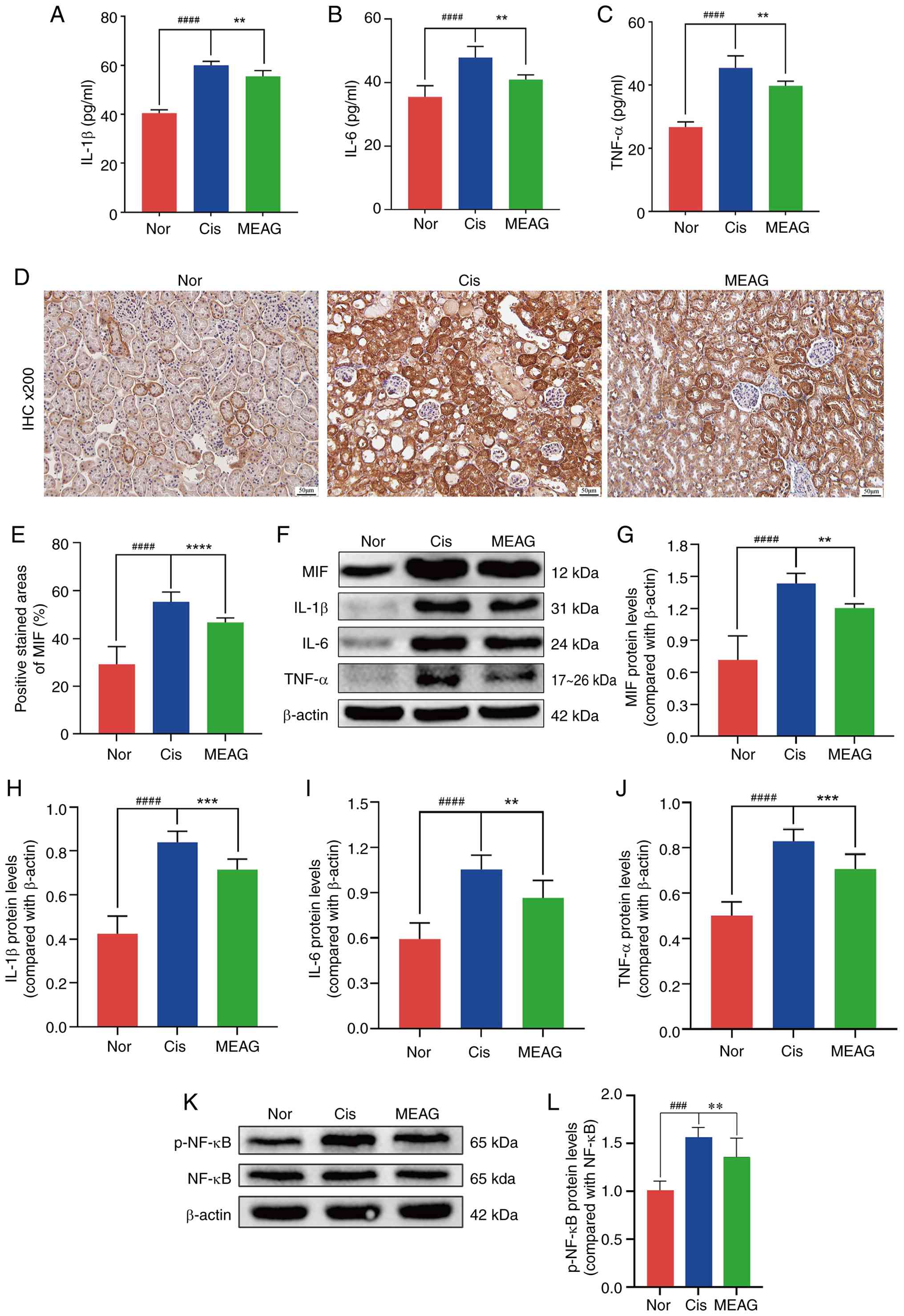 | Figure 10MEAG alleviated Cis-AKI in mice by
regulating the expression of MIF/NF-κB. (A-C) Serum concentrations
of pro-inflammatory cytokines (A) IL-1β, (B) IL-6 and (C) TNF-α
were quantified through ELISA across experimental cohorts
(n=3/group). (D) Renal tissue MIF distribution patterns visualized
through IHC staining (n=3/group). (E) Semiquantitative evaluation
of MIF immunoreactivity intensity. (F) Western blotting profiles
demonstrating protein expression alterations in MIF, p-NF-κB,
IL-1β, IL-6 and TNF-α protein levels. (G-J) Densitometric analysis
of (G) MIF, (H) IL-1β, (I) IL-6 and (J) TNF-α (n=3/group). (K)
Western blotting profiles demonstrating protein expression
alterations in p-NF-κB and NF-κB. (L) Densitometric analysis of
p-NF-κB/NF-κB (n=3/group). ####P<0.0001,
###P<0.001, compared with the Nor group;
****P<0.0001, ***P<0.001 and
**P<0.01, compared with the Cis group. Magnification,
x200 and scale bar, 50 µm. Nor, normal control group; Cis,
cisplatin; MEAG, methanolic extract of BaiYangJie; AKI, acute
kidney injury; IHC, immunohistochemistry; MIF, migration inhibitory
factor; p-, phosphorylated. |
MEAG alleviates Cis-AKI in mice by
regulating MIF/NF-κB expression
MIF expression profiles were quantitatively assessed
by IHC and western blotting. Histopathological examination
demonstrated a significant elevation of MIF immunoreactivity in
Cis-challenged murine models compared with the Nor group
(P<0.0001), which was attenuated following MEAG intervention
(P<0.0001; Fig. 10D and
E). Western blotting
quantification further demonstrated a significant reduction in
renal MIF protein content following MEAG intervention compared with
the Cis group (P<0.01; Fig.
10F and G), corroborating the
histopathological observations. In addition, a comparative analysis
between experimental groups revealed significantly enhanced
p-NF-κB/NF-κB levels in the Cis-treated group in comparison with
the Nor group (P<0.001), whereas MEAG administration
significantly suppressed this phosphorylation event (P<0.01;
Fig. 10K and L). These collective findings demonstrate
that MEAG treatment exerts regulatory effects on the pathological
activation of the MIF/NF-κB signalling cascade in Cis-induced AKI
models.
Discussion
Cis-induced nephrotoxicity manifests through a
number of pathological mechanisms, with acute tubular necrosis and
acute inflammatory responses being the most prevalent and notable
(29,30). As Cis is primarily excreted through
the kidneys, it is dependent on glomerular filtration and secretion
through the proximal renal tubules (31). Cis accumulates in the proximal
renal tubule fluid and spreads to high-permeability renal tubular
epithelial cells (19). After
renal tubular epithelial cells ingest Cis, a large number of cells
are damaged, eventually leading to acute renal tubular necrosis. In
the present study, the histological examination revealed evidence
of renal tubular necrosis following MEAG treatment and the results
showed that MEAG significantly ameliorated Cis-induced renal
tubular epithelial cell injury and necrosis.
Renal tubular epithelial cells secrete a number of
inflammatory cytokines and chemokines that induce inflammation
following damage (32). Among
these inflammatory factors, TNF-α is an important regulatory
component (33). Following
Cis-induced nephrotoxicity in mice, large quantities of TNF-α may
be produced. By inducing the expression of endothelial adhesion
molecules and chemokines, inflammatory leukocytes migrate to the
site of renal injury and stimulate the production of additional
inflammatory cytokines and chemokines, including IL-6 and IL-1β,
thereby promoting the local accumulation of immune effector cells
and further amplifying TNF-α-mediated inflammation (34). Consistent with previous studies,
the present study showed that Cis injection induced a severe
inflammatory reaction and significantly increased the expression of
pro-inflammatory cytokines (35,36).
To more comprehensively investigate the
physiological changes associated with AKI, metabolomics was used to
characterise metabolic network changes during AKI and to analyse
differential metabolites in the treatment of Cis-AKI. The results
showed that the phenylalanine metabolic pathway was the most
notable. Phenylalanine is a key amino acid, with previous studies
having shown that phenylalanine levels increase in patients with
AKI (37-39).
When phenylalanine levels are high, it acts as a metabolic toxin,
resulting in adverse health effects, which are common in chronic
inflammation and septic shock (40,41).
Phenylalanine hydroxylase converts L-phenylalanine into L-tyrosine
(42) and L-tyrosine accumulation
may lead to changes in energy metabolism, inflammation and systemic
immunity (43). The results of the
present study showed that Cis disrupted amino acid metabolism in
normal kidney tissue, leading to increased levels of phenylalanine
and tyrosine and resulting in inflammatory reactions in the kidney.
The metabolic levels of phenylalanine and tyrosine were reduced
following MEAG treatment, demonstrating that MEAG effectively
ameliorated the inflammatory response.
A total of 13 endogenous metabolites were identified
as differentially expressed in the present study. In addition to
phenylalanine, the other metabolites were primarily involved in
amino acids, lipid and energy metabolisms and the antioxidant
system, all of which are closely associated with the
pathophysiological processes and inflammatory responses associated
with Cis-AKI. Among amino acid metabolites, L-tryptophan is a
precursor in tryptophan metabolism and its metabolic disorder leads
to an imbalance in the kynurenine pathway, thereby exacerbating
oxidative stress and inflammatory responses in renal tissue
(44). L-pyroglutamic acid is a
key intermediate of glutamine metabolism, and Cis-induced
deviations in its level from that of the Nor group are associated
with energy metabolism disorders and renal tubular epithelial
apoptosis in renal injury (45).
MEAG reversed these Cis-induced abnormal changes in L-tryptophan
and L-pyroglutamic acid levels, alleviating the overall imbalance
in the amino acid metabolic network. Among lipid metabolism-related
substances, L-carnitine and stearoyl-L-carnitine were involved in
the β-oxidation of fatty acids, their Cis-induced deviations from
the Nor group levels reflected mitochondrial lipid metabolism
disorders in the kidney during Cis-AKI. Impaired mitochondrial
function can further trigger oxidative stress and the release of
inflammatory factors (46).
LysoPE (16:0/0:0) and docosahexaenoic acid (DHA) are
important components of phospholipid metabolism. Compared with the
Nor group, LysoPE (16:0/0:0) was increased in the Cis group, which
may impair the integrity of renal tubular cell membranes, whereas
DHA was decreased in the Cis group, weakens its anti-inflammatory
and antioxidant effects (47). The
regulation of the aforementioned lipid metabolites by MEAG restored
the physiological state of renal lipid metabolism and alleviated
cell membrane damage and inflammatory responses. Among energy
metabolism-associated substances, citric acid is a core
intermediate of the tricarboxylic acid cycle and creatine and
phosphocholine are involved in cellular energy storage and
transport; Cis-induced deviations in the levels of citric acid,
creatine and phosphocholine from those of the Nor group indicated
disruption of tricarboxylic acid cycle activity and cellular energy
storage and transport, and renal tubular epithelial cells undergo
injury and necrosis due to energy deficiency (48). However, MEAG restored these
metabolites to normal levels, improved energy metabolism in renal
tissue and provided energy support for cell repair. The levels of
the antioxidant ascorbic acid (vitamin C) were decreased in
Cis-AKI, reflecting high consumption by the renal antioxidant
system and thus, a cycle between oxidative stress and inflammatory
responses was formed (49,50). Furthermore, MEAG upregulated
vitamin C levels, enhanced the antioxidant capacity of the kidney,
reduced reactive oxygen species production and thereby inhibited
the release of inflammatory factors. The aforementioned
differential metabolites jointly constitute a network of renal
metabolic disorders in Cis-AKI. MEAG can regulate key nodes of this
network through numerous targets, thereby synergistically restoring
metabolic homeostasis and alleviating renal injury and inflammatory
responses. This regulatory effect synergises with the regulation of
phenylalanine metabolism and together mediates the protective
effect of MEAG on Cis-AKI.
A total of 210 potential MEAG targets against
Cis-AKI were identified across all disease databases and potential
drug targets. By analysing the DC, BC and CC values of the PPI
network, 11 core targets were identified, including AKT1, EGFR,
ALB, BCL2, HIF1A, HSP90AA1, SRC, ESR1, PTGS2, MAPK3 and GSK3B,
which may serve key roles in the MEAG interaction network targeting
Cis-AKI. Inhibiting AKT1 expression reduces oxidative stress in
cells and the levels of inflammatory factors such as IL-1β and
IL-6(51). MAPK3 is the final
component of the MAPK phosphorylation cascade and an important
member of the MAPK signalling pathway (52), where it serves central roles in
responses to numerous forms of stress and injury, such as oxidative
stress, inflammatory stimulation and metabolic stress, as well as
in the pathophysiology of a number of diseases, including renal
tubular inflammation and diabetic nephropathy (53). SRC family kinases are non-receptor
tyrosine kinases and their abnormalities can lead to inflammation,
autoimmune diseases and cancer, including colorectal, breast and
lung cancer (54).
Macrophage MIF is a pro-inflammatory cytokine and an
important upstream mediator of innate immunity, adaptive immunity
and survival pathways, including MAPK/ERK and PI3K/AKT signalling
pathways (55), which can activate
macrophages and T lymphocytes, regulating pathological processes,
including leukocyte recruitment, inflammation, immune responses,
cell proliferation, tumourigenesis and the reverse regulation of
glucocorticoids (56). Under
physiological circumstances, the expression of MIF in renal tissue
is low (57). However, following
AKI, severe renal inflammation occurs, including macrophage and T
cell infiltration. Thus, the expression of MIF is markedly
increased (58). In the present
study, it was determined that MIF expression in the renal tissue of
mice in the Cis group was significantly increased and associated
with the severity of renal tubular necrosis. MIF is associated with
the NF-κB pathway. Once released, MIF binds CD74 to initiate the
membrane recruitment of CD44, resulting in the activation of the
NF-κB signalling pathway, the expression of TNF-α, IL-1β and IL-6
increases and macrophages, neutrophils and T cells are recruited
and activated, exacerbating AKI (59).
The results of the present study showed that MEAG
significantly reduced the expression levels of MIF and the NF-κB
signalling pathway and reduced MIF-mediated TNF-α, IL-6 and IL-1β
levels, effectively inhibiting the inflammatory reaction and
improving Cis-AKI. In particular, NF-κB does not exist as an
independent signal pathway and it may directly or indirectly
regulate other molecules in addition to MIF, including previously
reported (60). A previous study
showed that NF-κB inhibited TNF-induced apoptosis of rat
hepatocytes by downregulating JNK and c-Jun/activator
protein-1(61). There appears to
be an association between STAT3 and NF-κB. IL-6 is a gene product
regulated by the NF-κB signal and an important STAT3 activator
(62). In renal diseases, NF-κB
not only participates in the transcription of pro-inflammatory
genes in T helper (Th)-1 and Th17 cells, but also promotes their
differentiation into CD4 T cells through interaction with innate
immune cells (63). In addition,
mesangial epithelial cells and renal tubular epithelial cells can
also promote inflammation through toll-like receptor/NF-κB axes
(64). The discovery of these
synergistic regulatory effects may highlight a novel direction for
further exploring the mechanism of MEAG in treating Cis-AKI.
Notably, integrated analysis suggested that MIF
served as a pivotal molecular bridge associating phenylalanine
metabolic dysfunction with NF-κB-mediated inflammatory responses.
In the context of Cis-AKI, renal dysfunction leads to the marked
accumulation of phenylalanine and its downstream metabolite,
tyrosine (65). These metabolites
act as ‘metabolic toxins’ that can directly trigger or exacerbate
chemical stress and inflammatory reactions within the renal
microenvironment (66). Database
intersection analysis identified MIF as a key target within the
phenylalanine metabolic pathway. Thus, it was hypothesised that
stress signals arising from metabolic imbalances (specifically,
accumulation of phenylalanine) may upregulate MIF expression. Once
upregulated, MIF binds to its receptor complex, activating
NF-κB-mediated signalling cascade and leading to the release of
pro-inflammatory cytokines such as TNF-α, IL-6 and IL-1β, thereby
forming a cycle between metabolic disorder and inflammatory injury.
The therapeutic efficacy of MEAG is characterised by a
‘dual-regulatory’ strategy; it not only restores phenylalanine
metabolic homeostasis, thereby reducing the production of metabolic
toxins, but also directly suppresses the pathological activation of
the MIF/NF-κB axis. This synergistic modulation of metabolic and
immune pathways provides a systematic pharmacological basis for the
renoprotective effects of MEAG against chemotherapy-induced
nephrotoxicity.
Collectively, the results of the present study
showed that MEAG may serve a role in the treatment of Cis-AKI by
regulating numerous targets and signalling pathways. However, there
were also a number of limitations, such as the lack of a positive
drug control group, incomplete data searches, insufficient coverage
of Traditional Chinese Medicine components and dependence on
calculation and prediction. A positive control group was not
established in the present study, as to the best of our knowledge,
there are currently no specific drugs for Cis-AKI in clinical
practice. Furthermore, network pharmacology analysis in the present
study failed to clearly illustrate the direct binding association
between specific chemical components in MEAG and the targets
associated with the MIF/NF-κB signalling pathway or phenylalanine
metabolism. The potential bioactive components and core targets
were only screened through component-target-pathway analysis,
without conducting molecular docking simulation or in vitro
binding verification, meaning it was not possible to clarify the
direct interaction mode and binding affinity between single
components or component combinations and core targets and also
hindered the accurate interpretation of the structure-activity
association between the material basis of the renoprotective effect
of MEAG and target binding.
To address these shortcomings, the present study
combined analysis of serum metabolomics data with verification by
western blotting and IHC analysis to improve the accuracy of the
results. Based on the aforementioned limitations, future research
should aim to conduct molecular docking experiments using the
identified active components in MEAG (such as resveratrol and
pterostilbene), screen out active monomers that can directly bind
to core targets such as MIF and NF-κB and further verify the
binding affinity and binding sites between components and targets
through in vitro technologies, such as surface plasmon
resonance and microscale thermophoresis. In addition, in
vitro cell experiments should be used to clarify the
dose-effect association and mechanism of action of single active
components or component combinations in regulating the MIF/NF-κB
signalling pathway and phenylalanine metabolism, so as to provide
further direct experimental evidence for understanding the material
basis and structure-activity association of the therapeutic effects
of MEAG. Follow-up studies should also aim to construct a positive
drug control group with clinically commonly used renoprotective
drugs to further determine the therapeutic effect of MEAG on
Cis-AKI. In addition, the aforementioned hub targets and key
signalling pathways may also serve important roles in Cis-AKI and
other kidney diseases, warranting further exploration.
In summary, the present study combined network
pharmacology with metabolomics to demonstrate that MEAG improves
Cis-AKI by reducing inflammatory reactions and regulating MIF/NF-κB
signalling pathway. In addition, serum phenylalanine levels were a
key differential metabolite that may be used to evaluate
preventative treatment and therapeutic effects. This comprehensive
strategy offers a potential method for identifying the
multi-target, multi-pathway pharmacodynamic mechanism of botanical
drugs and provided a theoretical basis for the future development
of BaiYangJie in clinical practice.
Supplementary Material
Plant identification report.
Differentially expressed serum
endogenous metabolites in Nor, Cis and MEAG groups.
Topological properties of core
targets.
Acknowledgements
Not applicable.
Funding
Funding: The present study was financially supported by the
China Academy of Medical Sciences, Medical and Health Science and
Technology Innovation Project (grant no. 2021-I2M-1-031), Yunnan
Province Wang Jinhui Expert Workstation (grant no. 202405AF140073),
Yunnan Science and Technology Talents and Platform Plan (grant no.
202105 AF070011), Yunnan Fundamental Research Projects (grant no.
202201AT070286), the Major Science and Technology Special Plan of
Yunnan Province (grant no. 202402AA310041) and the ‘Rainforest
Talent Support Program’ for Young Talents in Xishuangbanna
Prefecture (grant no. 2023006).
Availability of data and materials
The data generated in the present study may be found
in the National Genomics Data Center Open Archive for Miscellaneous
Data database under accession number OMIX016002 or at the following
URL: (persistent, https://ngdc.cncb.ac.cn/omix/release/OMIX016002).
Authors' contributions
BX and GL contributed to the conception and design
of the study and drafted the manuscript. LZ and XD contributed to
study design, data interpretation and critical revision of the
manuscript for important intellectual content. JLC, JS, YL, SL, XZ
and TW performed the experiments and contributed to data
acquisition and interpretation. JW, DL, XZ and TW performed the
network pharmacology and serum metabolomics analyses and
contributed to data interpretation. LZ contributed to resources,
project administration and funding acquisition. GL and BX confirm
the authenticity of all the raw data. All authors revised the
manuscript, read and approved the final version, and agree to be
accountable for all aspects of the work. GL and BX confirm the
authenticity of all the raw data.
Ethics approval and consent to
participate
All animals were obtained from Sibefu (Beijing)
Biotechnology Co., Ltd. The animal research protocol was approved
by the Institutional Ethics Review Committee of the Yunnan Branch
of the Institute of Medicinal Plant Development, Chinese Academy of
Medical Sciences (Jinghong, China; approval no. 20240213001).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Use of artificial intelligence tools
During the preparation of this work, artificial
intelligence tools were used to improve the readability and
language of the manuscript or to generate images, and subsequently,
the authors revised and edited the content produced by the
artificial intelligence tools as necessary, taking full
responsibility for the ultimate content of the present
manuscript.
References
|
1
|
Ostermann M, Lumlertgul N, Jeong R, See E,
Joannidis M and James M: Acute kidney injury. Lancet. 405:241–256.
2025.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Scholz H, Boivin FJ, Schmidt-Ott KM,
Bachmann S, Eckardt KU, Scholl UI and Persson PB: Kidney physiology
and susceptibility to acute kidney injury: Implications for
renoprotection. Nat Rev Nephrol. 17:335–349. 2021.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Susantitaphong P, Cruz DN, Cerda J,
Abulfaraj M, Alqahtani F, Koulouridis I and Jaber BL: Acute Kidney
Injury Advisory Group of the American Society of Nephrology. World
incidence of AKI: A meta-Analysis. Clin J Am Soc Nephro.
8:1482–1493. 2013.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Dasari S and Tchounwou PB: Cisplatin in
cancer therapy: Molecular mechanisms of action. Eur J Pharmacol.
740:364–378. 2014.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Motwani SS, Kaur SS and Kitchlu A:
Cisplatin nephrotoxicity: Novel insights into mechanisms and
preventative strategies. Semin Nephrol. 42(151341)2022.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Ambe K, Aoki Y, Murashima M, Wachino C,
Deki Y, Ieda M, Kondo M, Furukawa-Hibi Y, Kimura K, Hamano T and
Tohkin M: Prediction of cisplatin-induced acute kidney injury using
an interpretable machine learning model and electronic medical
record information. Clin Transl Sci. 18(e70115)2025.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Ghosh S: Cisplatin: The first metal based
anticancer drug. Bioorg Chem. 88(102925)2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Fang CY, Lou DY, Zhou LQ, Wang JC, Yang B,
He QJ, Wang JJ and Weng QJ: Natural products: Potential treatments
for cisplatin-induced nephrotoxicity. Acta Pharmacol Sin.
42:1951–1969. 2021.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Zhang X, Chen W, Du Y, Su P, Qiu Y, Ning J
and Liu M: Phytochemistry and pharmacological activities of
Arundina graminifolia (D.Don) Hochr. And other common
Orchidaceae medicinal plants. J Ethnopharmacol.
276(114143)2021.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Li J, Zhou L, Wang J, Zhang L, Xia B, Li
G, Ren J and Li J: Evaluation of the methanol extract of
BaiYangJie: Toxicology and protective effect against acute kidney
injury. J Adv Pharm Technol Res. 15:185–193. 2024.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Newgard CB: Metabolomics and metabolic
diseases: Where do we stand? Cell Metab. 25:43–56. 2017.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Cao H, Zhang A, Zhang H, Sun H and Wang X:
The application of metabolomics in traditional Chinese medicine
opens up a dialogue between Chinese and Western medicine. Phytother
Res. 29:159–166. 2015.PubMed/NCBI View Article : Google Scholar
|
|
13
|
National Research Council, Division on
Earth and Life Studies, Institute for Laboratory Animal Research,
Committee for the Update of the Guide for the Guide for the Care
and Use of Laboratory Animals: Eighth edition. Guide for the care
and use of laboratory animals. National Academies Press, 2011.
|
|
14
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The gene
ontology consortium. Nat Genet. 25:25–29. 2000.PubMed/NCBI View Article : Google Scholar
|
|
16
|
The Gene Ontology Consortium. The gene
ontology resource: 20 years and still GOing strong. Nucleic Acids
Res. 47 (D1):D330–D338. 2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Tsugawa H, Cajka T, Kind T, Ma Y, Higgins
B, Ikeda K, Kanazawa M, VanderGheynst J, Fiehn O and Arita M:
MS-DIAL: Data-independent MS/MS deconvolution for comprehensive
metabolome analysis. Nat Methods. 12:523–526. 2015.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Yan ZH, Li WS, Liang BY, et al: Comparison
of chemical constituents between Isatidis Radix and Isatidis Folium
based on UPLC-Q-TOF-MS/MS technology. Chin Tradit Herbal Drugs.
55:3956–3965. 2024.
|
|
19
|
Meng XW, Cai DJ, Zhu Q, et al: Rapid
identification of chemical constituents in Dalbergia
cochinchinensis heartwood based on UPLC-Q-TOF-MS/MS technology.
Chin J Exp Tradit Med Formula. 26:143–156. 2020.
|
|
20
|
Wang YC, Han B, Li ZJ, et al: Study on
chemical constituents of Dendrobium pendulum based on
UPLC-Q-TOF-MS. Chin Pharm J. 56:708–714. 2021.
|
|
21
|
Mou Y, Liu B, Zhang X, et al: Component
analysis and anti-inflammatory activity evaluation of different
parts of Forsythia suspensa based on UPLC-Q-TOF-MS. China J Chin
Mater Med. 49:968–980. 2024.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Huo JH, Sun GD, Wei WF, et al: Analysis of
chemical constituents in Niuhuang Qinggan Capsules by
UPLC-Q-TOF/MS. Chin Tradit Pat Med. 40:2340–2348. 2018.
|
|
23
|
Lei JC, Zhang ST, Hu XR, et al:
Investigation of the pharmacodynamic material basis and mechanism
of Jinbei Oral Liquid against idiopathic pulmonary fibrosis by
UHPLC-Q-TOF-MS/MS combined with network pharmacology. China J Chin
Mater Med. 1–16. 2025.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Wu CH, Liu YY, Xing SP, et al: Screening
of potential anti-inflammatory active components of Gonglaomu based
on UPLC-Q-TOF-MS metabolomics technology. J Chin Med Mater.
48:389–395. 2025.
|
|
25
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Hayden MS, West AP and Ghosh S: NF-kappaB
and the immune response. Oncogene. 25:6758–6780. 2006.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Zhang Y, Tang PM, Niu Y, García Córdoba
CA, Huang XR, Yu C and Lan HY: Long non-coding RNA LRNA9884
promotes acute kidney injury via regulating NF-kB-mediated
transcriptional activation of MIF. Front Physiol.
11(590027)2020.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Chen J, Xu X, Cai Y, Zhang Q, Wang X and
Zhang D: Acute kidney injury over the past decade: From definition
evolution to pathogenesis insights and innovative therapeutic
strategies. Cell Mol Life Sci. 83(156)2026.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Gao J, Deng Q, Yu J, Wang C and Wei W:
Role of renal tubular epithelial cells and macrophages in
cisplatin-induced acute renal injury. Life Sci. 339:122450.
2024.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Megyesi J, Safirstein RL and Price PM:
Induction of p21WAF1/CIP1/SDI1 in kidney tubule cells affects the
course of cisplatin-induced acute renal failure. J Clin Invest.
101:777–782. 1998.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Holditch SJ, Brown CN, Lombardi AM, Nguyen
KN and Edelstein CL: Recent advances in models, mechanisms,
biomarkers, and interventions in cisplatin-induced acute kidney
injury. Int J Mol Sci. 20(3011)2019.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Salei N, Rambichler S, Salvermoser J,
Papaioannou NE, Schuchert R, Pakalniškytė D, Li N, Marschner JA,
Lichtnekert J, Stremmel C, et al: The kidney contains
ontogenetically distinct dendritic cell and macrophage subtypes
throughout development that differ in their inflammatory
properties. J Am Soc Nephrol. 31:257–278. 2020.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Ramesh G and Reeves WB: TNF-alpha mediates
chemokine and cytokine expression and renal injury in cisplatin
nephrotoxicity. J Clin Invest. 110:835–842. 2002.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Pober JS: Endothelial activation:
Intracellular signaling pathways. Arthritis Res. 4 (Suppl
3):S109–S116. 2002.PubMed/NCBI View
Article : Google Scholar
|
|
35
|
Zhang J, Luan ZL, Huo XK, Zhang M,
Morisseau C, Sun CP, Hammock BD and Ma XC: Direct targeting of sEH
with alisol B alleviated the apoptosis, inflammation, and oxidative
stress in cisplatin-induced acute kidney injury. Int J Biol Sci.
19:294–310. 2023.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Zhou X, Jiang K, Luo H, Wu C, Yu W and
Cheng F: Novel lncRNA XLOC_032768 alleviates cisplatin-induced
apoptosis and inflammatory response of renal tubular epithelial
cells through TNF-α. Int Immunopharmacol. 83(106472)2020.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Qu X, Gao H, Sun J, Tao L, Zhang Y, Zhai
J, Song Y, Hu T and Li Z: Identification of key metabolites during
cisplatin-induced acute kidney injury using an HPLC-TOF/MS-based
non-targeted urine and kidney metabolomics approach in rats.
Toxicology. 431(152366)2020.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Mahmod II, Ismail IS, Alitheen NB, Normi
YM, Abas F, Khatib A, Rudiyanto and Latip J: NMR and LCMS
analytical platforms exhibited the nephroprotective effect of
Clinacanthus nutans in cisplatin-induced nephrotoxicity in the in
vitro condition. BMC Complement Med Ther. 20(320)2020.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Sun J, Shannon M, Ando Y, Schnackenberg
LK, Khan NA, Portilla D and Beger RD: Serum metabolomic profiles
from patients with acute kidney injury: A pilot study. J Chromatogr
B Analyt Technol Biomed Life Sci. 893-894:107–113. 2012.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Huang SS, Lin JY, Chen WS, Liu MH, Cheng
CW, Cheng ML and Wang CH: Phenylalanine- and leucine-defined
metabolic types identify high mortality risk in patients with
severe infection. Int J Infect Dis. 85:143–149. 2019.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Song Y, Hu T, Gao H, Zhai J, Gong J, Zhang
Y, Tao L, Sun J, Li Z and Qu X: Altered metabolic profiles and
biomarkers associated with astragaloside IV-mediated protection
against cisplatin-induced acute kidney injury in rats: An
HPLC-TOF/MS-based untargeted metabolomics study. Biochem Pharmacol.
183(114299)2021.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Ashe K, Kelso W, Farrand S, Panetta J,
Fazio T, De Jong G and Walterfang M: Psychiatric and cognitive
aspects of phenylketonuria: The limitations of diet and promise of
new treatments. Front Psychiatry. 10(561)2019.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Zheng T, Su S, Dai X, Zhang L, Duan JA and
Ou-Yang Z: Metabolomic analysis of biochemical changes in the serum
and urine of freund's Adjuvant-Induced arthritis in rats after
treatment with silkworm excrement. Molecules.
23(1490)2018.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Tokuno M, Commey KL, Yamamoto A, Tokushige
M, Tsukigawa K, Nishi K, Otagiri M, Ekino K and Yamasaki K:
Insights into the renal protective effects of 7-phenylheptanoic
acid in chronic kidney disease mice: Modulation of indoxyl sulfate
production and gut microbiome homeostasis. Drug Metab Dispos.
53(100149)2025.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Wang SY and Nesheim MC: Kidney clearance
of D- and L-tryptophan by rats. Comp Biochem Physiol A Comp
Physiol. 85:451–453. 1986.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Gao J, Gu Z, Li M, Xu Y, Gao Y, Wei J,
Liang B and Na Y: L-carnitine ameliorates the decrease of aquaporin
2 levels in rats with cisplatin-induced kidney injury. Nephron.
135:315–325. 2017.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Chisty TTE, Sarif S, Jahan I, Ismail IN,
Chowdhury FI, Siddiqua S, Yasmin T, Islam MN, Khan F, Subhan N and
Alam MA: Protective effects of l-carnitine on isoprenaline-induced
heart and kidney dysfunctions: Modulation of inflammation and
oxidative stress-related gene expression in rats. Heliyon.
10(e25057)2024.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Li AS, Baker PR II, Park S, Budnick I, He
Z, Okamura K, Gil HW, Miyazaki M, Anderson CC, Reisz JA and Faubel
S: Metabolomic assessment reveals depletion of amino acids and
energy metabolites in skeletal muscle after ischemic acute kidney
injury in mice. Sci Rep. 16(8823)2026.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Okamoto K, Kitaichi F, Saito Y, Ueda H,
Narumi K, Furugen A and Kobayashi M: Antioxidant effect of ascorbic
acid against cisplatin-induced nephrotoxicity and P-glycoprotein
expression in rats. Eur J Pharmacol. 909(174395)2021.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Wei T, Liu B, Chen Y and Li C: Protective
effect of ascorbic acid against renal injury induced by
3-chloropropane-1,2-diol-dipalmitate in rats. Renal Fail.
46(2429694)2024.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Kim IY, Song SH, Seong EY, Lee DW, Bae SS
and Lee SB: Akt1 is involved in renal fibrosis and tubular
apoptosis in a murine model of acute kidney injury-to-chronic
kidney disease transition. Exp Cell Res. 424(113509)2023.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Lucas RM, Luo L and Stow JL: ERK1/2 in
immune signalling. Biochem Soc Trans. 50:1341–1352. 2022.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Capolongo G, Suzumoto Y, D'Acierno M,
Simeoni M, Capasso G and Zacchia M: ERK1,2 signalling pathway along
the nephron and its role in acid-base and electrolytes balance. Int
J Mol Sci. 20(4153)2019.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Poh AR, Love CG, Chisanga D, Steer JH,
Baloyan D, Chopin M, Nutt S, Rautela J, Huntington ND, Etemadi N,
et al: Therapeutic inhibition of the SRC-kinase HCK facilitates T
cell tumor infiltration and improves response to immunotherapy. Sci
Adv. 8(eabl7882)2022.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Sumaiya K, Langford D, Natarajaseenivasan
K and Shanmughapriya S: Macrophage migration inhibitory factor
(MIF): A multifaceted cytokine regulated by genetic and
physiological strategies. Pharmacol Ther.
233(108024)2022.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Bloom BR and Bennett B: Mechanism of a
reaction in vitro associated with delayed-type hypersensitivity.
Science. 153:80–82. 1966.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Lan HY: Role of macrophage migration
inhibition factor in kidney disease. Nephron Exp Nephrol.
109:e79–e83. 2008.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Kong YZ, Chen Q and Lan HY: Macrophage
migration inhibitory factor (MIF) as a stress molecule in renal
inflammation. Int J Mol Sci. 23(4908)2022.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Pohl J, Hendgen-Cotta UB, Stock P, Luedike
P and Rassaf T: Elevated MIF-2 levels predict mortality in
critically ill patients. J Crit Care. 40:52–57. 2017.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Hoffmann A: Immune response signaling:
Combinatorial and dynamic control. Trends Immunol. 37:570–572.
2016.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Liu H, Lo CR and Czaja MJ: NF-kappaB
inhibition sensitizes hepatocytes to TNF-induced apoptosis through
a sustained activation of JNK and c-Jun. Hepatology. 35:772–778.
2002.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Grivennikov S, Karin E, Terzic J, Mucida
D, Yu GY, Vallabhapurapu S, Scheller J, Rose-John S, Cheroutre H,
Eckmann L and Karin M: IL-6 and Stat3 are required for survival of
intestinal epithelial cells and development of colitis-associated
cancer. Cancer Cell. 15:103–113. 2009.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Zhang H and Sun SC: NF-κB in inflammation
and renal diseases. Cell Biosci. 5(63)2015.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Liu H, Xiong J, He T, Xiao T, Li Y, Yu Y,
Huang Y, Xu X, Huang Y, Zhang J, et al: High uric acid-induced
epithelial-mesenchymal transition of renal tubular epithelial cells
via the TLR4/NF-kB signaling pathway. Am J Nephrol. 46:333–342.
2017.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Zhang B, Zeng M, Wang Y, Li M, Wu Y, Xu R,
Zhang Q, Jia J, Huang Y, Zheng X and Feng W: Oleic acid alleviates
LPS-induced acute kidney injury by restraining inflammation and
oxidative stress via the Ras/MAPKs/PPAR-γ signaling pathway.
Phytomedicine. 94(153818)2022.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Hu JC, Song JY, Feng R, Ye ML, Xu H, Lu
JY, Zuo HT, Zhao Y, Wang JY, Jin JY, et al: Integrated gut
microbiota-drug interaction analysis and network pharmacology for
the investigation of renal-protective effect of Polygala tenuifolia
Willd. Int J Mol Sci. 26(10889)2025.PubMed/NCBI View Article : Google Scholar
|