Introduction
The skin serves as the outermost barrier of the
human body and is particularly susceptible to various external
stressors. Among these, harmful environmental factors such as
ultraviolet (UV) radiation, air pollution and environmental toxins
are major contributors that can disturb the cutaneous redox
balance, resulting in oxidative stress and subsequent inflammation.
This inflammatory response may accelerate the progression of
various skin disorders, including photoaging, psoriasis, pigmentary
abnormalities and skin tumors (1-4).
Inflammation is a natural response of the body to infections,
tissue injuries, chemicals, endotoxins and other factors. An
adequately regulated inflammatory response is beneficial as it
helps protect the body from harmful stimuli; however, when
inflammation becomes uncontrolled, it can damage tissue structures
and functions, leading to further deterioration in various
diseases, such as psoriasis and atopic dermatitis (5-7).
Cyclooxygenase-2 (COX-2) is typically expressed at
low or undetectable levels in normal tissues under physiological
conditions; however, it can be induced by pro-inflammatory
cytokines, growth factors, oncogenes, carcinogens and tumor
promoters, indicating a role for COX-2 in inflammation and the
regulation of cell proliferation (8-10).
Compounds that inhibit COX-2 activity or expression have been
widely investigated and utilized as therapeutic targets,
particularly in cancer prevention and anti-inflammation strategies
(11).
Inducible nitric oxide synthase (iNOS) serves a role
in the development of various inflammatory conditions (12). iNOS is minimally expressed or
virtually undetectable in normal resting cells but is induced by
inflammatory cytokines or a combination of these cytokines and
bacterial polysaccharides. Often, a synergistic effect is observed
between different external stimuli, such as pro-inflammatory
cytokines, bacterial components (e.g., lipopolysaccharide), and
environmental stressors, which is mediated by the nuclear
factor-kappa B (NF-κB) signaling pathways (13).
Signal transducers and activators of transcription 3
(STAT3) serves a crucial role in regulating cell survival and
inflammation. STAT3 influences gene expression related to survival,
proliferation and angiogenesis by interacting with other
transcription factors, such as NF-κB. (14,15)
(NF-κB plays a crucial role in host defense and the inflammatory
response to microbial and viral infections (16); upon exposure to external signals
such as bacterial lipopolysaccharide (LPS), tumor necrosis factor-α
or other inflammatory mediators, NF-κB is activated. Once
activated, this transcription factor promotes the expression of
various genes involved in inflammation, including COX-2,
iNOS and specific cytokines such as tumor necrosis factor-α
(TNF-α), interleukin-1β (IL-1β), and interleukin-6 (IL-6) (17,18).
STAT3 and NF-κB are transcription factors that can increase the
expression of pro-inflammatory enzymes and mediators; consequently,
the STAT3 and NF-κB pathways have been suggested as potential
targets for controlling inflammation (19,20).
Belamcanda chinensis (L.) DC. (BC) is a
traditional Chinese medicinal herb, and its dried rhizome is widely
used in China to treat inflammation, asthma and throat disorders
(21-24).
It is also effective against several bacterial, fungal and viral
organisms, and has been used as a folk medicine in China (25). Thus, it can be inferred that the
presence of essential chemical compounds with known
anti-inflammatory properties has played a notable role in the
prevalent use of this herb in traditional medicine (26).
In recent years, increasing attention has been
directed toward integrating natural phytochemicals with
conventional pharmaceuticals to enhance therapeutic efficacy and
reduce adverse effects (27,28).
In this context, BC extract was selected for testing in the present
study, due to its well-documented antioxidant and anti-inflammatory
activities, as well as its potential applicability in the
development of combination therapies (29,30).
Notably, Belamcanda chinensis has been traditionally used as
a key component of multi-herb formulations such as Shegan Mahuang
Tang and Qing Yan Li Ge Wan, which are prescribed for the treatment
of respiratory and inflammatory conditions including asthma, cough,
pharyngitis, and throat inflammation. Unlike previous studies
(31,32) that primarily focused on evaluating
the efficacy of individual standard compounds, the present study
provided a distinctive approach by identifying the specific
bioactive constituents within the BC extract that predominantly
contribute to its molecular interactions. This enabled the
exploration of potential competitive or synergistic relationships
among the components, thereby offering a more comprehensive
understanding of the overall pharmacological potential of the
extract and supporting its relevance for future drug
development.
Materials and methods
Plant extract preparation
BC is native to Cheongju, Chungcheongbuk-do, Korea,
and the plant material was obtained from the Herbmaul e-shop
(https://aromainherb.net/shopinfo/company.html, plant
code, 1211.90-1999). The purchased rhizomes were harvested, washed
with water, cut into pieces and air dried at 60-70˚C for 72 h. They
were then sealed in polyethylene bags with silica gel and stored at
-20˚C until further use.
Extraction and purification of BC
phenolic compounds
The plant polyphenols were extracted using a
modified method (33). First, 200
g rhizomes of BC was steeped in 10 l of 70% ethanol for 4 days at
room temperature, and filter paper was used to remove contaminants
from the extract (Whatman Qualitative No. 6). Afterwards, the
extract was concentrated at 45˚C using a rotary evaporator (N-1110;
Eyela; Tokyo Rikakikai Co., Ltd.) until the volume was reduced to
500 ml. Following this, 500 ml hexane was used three times to wash
the concentrated extract to remove fatty particles. Then, 250 ml of
ethyl acetate was used three times to extract the remaining
extract, yielding the ethyl acetate fraction. The residue was
eluted using silica gel solvent (40x2.5 cm) and ethyl acetate,
followed by dehydration with MgSO4 to remove the
residual water. The extract was concentrated at a pressure between
100 to 130 mbar and stored at -80˚C to produce a mixed polyphenol
powder, which was 6.88% of the raw material, or 48.2 g.
High-performance liquid chromatography
(HPLC) and LC-mass spectrometry (MS/MS) analysis
A 10 µl sample of the extract powder was diluted in
70% ethanol to a concentration of 1,000 µg/ml was used for compound
identification. The Ultra Quadrupole Time of Flight LC-MS/MS System
(X500R; SCIEX, Inc.) and Nexera Lite HPLC system (Shimadzu
Corporation) were used for HPLC and LC-MS/MS. Both systems operated
in the positive ion mode with the voltage set at -4.5 kV. Using
electrospray ionization and multi-scan in the m/z 100-2,000 range,
mass spectrometry settings were carried out in positive ion mode.
500˚C was the desolvation temperature, 5,500 V was the spray
voltage, 50 psi was the ion source gas pressure, 30 psi was the
curtain gas pressure and 80 V was the declustering potential. For
MS/MS spectra, the collision energy was fixed at 35±15 V. SCIEX OS
software (3.0.0) was used to generate the obtained ion chromatogram
data. The solvents used were, distilled water (solvent A) and
acetonitrile with 0.1% formic acid (solvent B). A Prontosil C18
column (length: 250 mm, inner diameter: 4.6 mm, particle size: 5
µm; manufactured by Bischoff Chromatography and distributed by
Phenomenex Inc. was used, and a gradient system was employed for
analysis at a flow rate of 0.5 ml/min. The solvent B conditions for
the mobile phases were as follows: 10-15% for 0-10 min, 20% for
20-30 min, 40% for 30-40 min, 70% for 40-50 min and 95% for 50-60
min. A wavelength of 284 nm and a temperature of 35˚C were used for
the analysis.
Antioxidant activity using
2,2-Diphenyl-1-picrylhydrazyl (DPPH) binding HPLC technology
A 1:1 (v:v) mixture of 0.2 mg/ml DPPH
(MilliporeSigma) and 1 mg/ml (1,000 ppm) BC sample was allowed to
react for 15 min at room temperature. A 0.45 µm syringe filter was
used to filter the mixture before HPLC analysis, and methanol was
used as a control in place of the DPPH reagent. Identification of
the reacting chemical composition was made by analyzing the
chromatographic peak values and standard curve results from the
samples and controls that underwent the DPPH reaction. This enabled
the identification of the key antioxidant elements found in the
phenolic compounds of BC.
Cell culture
The human keratinocyte HaCaT cell line, which is
widely used as an in vitro model to study skin inflammation
and oxidative stress due to its stable phenotype and
well-characterized inflammatory signaling responses, was purchased
from AddexBio Technologies (cat. no. T0020001). The cell line
identity was verified by matching the supplier-provided STR profile
with the HaCaT reference profile from Ubigene (https://www.ubigene.us/). The cells were grown in
Dulbecco's modified Eagle's medium (Gibco; Thermo Fisher
Scientific, Inc.) with 10% heat-inactivated fetal bovine serum
(Gibco; Thermo Fisher Scientific, Inc.) with 100 U/ml penicillin
and 100 g/ml streptomycin. The cells were incubated at 37˚C in a
humidified environment with 5% CO2.
Cell viability assay
HaCaT cells (5x103) were seeded into
96-well plates and treated with BC extract dissolved in dimethyl
sulfoxide (DMSO) until fully solubilized at various concentrations
(0, 0.5, 1, 2.5, 5, 7.5, 10, 12.5, 25, 50, 75 and 100 µg/ml) for 24
h, either with or without 1 µg/ml lipopolysaccharide (LPS;
MilliporeSigma), which was prepared by dissolving LPS in sterile
phosphate-buffered saline (PBS) to ensure complete solubility, with
0.01% of DMSO alone used as the vehicle control. MTT reagent (0.5
mg/ml, dissolved in PBS) was added into each well for 2 h at 37˚C
and then the optical density was measured at 450 nm using a
microplate reader (Synergy HTX; BioTek; Agilent Technologies,
Inc.).
Western blotting
HaCaT cells (5x105) were seeded into
6-well plates and treated for 24 h with BC extract (0.5, 1 and 2
µg/ml), which was dissolved in DMSO, either with or without 1 µg/ml
LPS at 37˚C. DMSO was used as the vehicle control. Cell pellets
were lysed in RIPA buffer (50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 1
mM EDTA, 1% Triton X-100, 1% sodium deoxycholate, 0.1% SDS) and
then centrifuged at 11,000 x g for 10 min at 4˚C to remove debris.
Equal amounts of protein (10 µg), as determined by a bicinchoninic
acid protein assay, were separated via Mini-PROTEAN TGX 4-20% gels
(Bio-Rad Laboratories, Inc.) and transferred onto PVDF membranes in
iBlot™ 3 Transfer Stacks (Thermo Scientific; Thermo
Fisher Scientific, Inc.) using the iBlot 3 Western Blot Transfer
System (Thermo Scientific; Thermo Fisher Scientific, Inc.). After
blocking with EzBlockChemi (ATTO Corporation), the membranes were
incubated overnight at 4˚C with the following primary antibodies,
each diluted to 1:1,000: COX-2 (cat. no. ab15191; Abcam), iNOS
(cat. no. ab283655; Abcam), phospho-STAT3 (cat. no. 9131; Cell
Signaling Technology, Inc.), STAT3 (cat. no. 9139; Cell Signaling
Technology, Inc.), phospho-p65 (cat. no. 3031; Cell Signaling
Technology, Inc.), p65 (cat. no. 8242; Cell Signaling Technology,
Inc.), phospho-IκB-α (cat. no. 2859; Cell Signaling Technology,
Inc.), IκB-α (cat. no. 9242; Cell Signaling Technology, Inc.) and
β-actin (cat. no. A5441; MilliporeSigma). The membranes were
incubated 2 h at room temperature with the following second
antibodies, each diluted to 1:5,000: Horseradish peroxidase
(HRP)-conjugated secondary antibodies to anti-rabbit (cat. no.
A120-101P; Bethyl Laboratories, Inc.) and anti-mouse (cat. no.
A90-116P; Bethyl Laboratories, Inc.). The blots were visualized
using SuperSignal™ West Pico PLUS Chemiluminescent Substrate
(Thermo Scientific; Thermo Fisher Scientific, Inc.) and
semi-quantified by densitometry using ImageJ software (version
1.54p) (National Institutes of Health) with β-actin as the loading
control.
Molecular docking
The protein structure was obtained from the Protein
Data Bank (PDB; https://www.wwpdb.org/) using the search ID 4Q3J
(NF-κB). The 3D compound structures of tectoridin (ID: 5281810),
iridin (ID: 5281777), genistein (ID: 5280961), tectorigenin (ID:
5281811) and irisflorentin (ID: 170569) were downloaded from
PubChem (https://pubchem.ncbi.nlm.nih.gov/). Docking analysis
was performed using the default settings of UCSF Chimera (version
1.18; https://www.cgl.ucsf.edu/chimera/) and AutoDock Vina
(https://vina.scripps.edu/). To ensure
reproducibility, detailed grid box parameters were applied for each
receptor-ligand pair. NF-κB was docked with tectoridin (center:
8.98975, 30.8691, 54.2354; size: 36.5078, 53.8813, 36.2329), iridin
(center: 8.99029, 28.355, 52.8342; size: 36.5068, 48.8537,
39.0972), genistein (center: 9.20401, 31.1588, 53.1349; size:
35.0665, 54.4613, 39.7049), tectorigenin (center: 8.99103, 30.7641,
53.8333; size: 36.506, 53.6719, 38.3141) and irisflorentin (center:
8.9912, 29.8613, 53.1376; size: 36.506, 50.9934, 39.7055). These
parameters were carefully selected to comprehensively cover the
active binding region and ensure consistent and reproducible
docking conditions across all ligands. The docking results,
including the interactions of amino acids, were displayed using
Discovery Studio 2021 Client (https://www.3ds.com/products/biovia/discovery-studio).
The binding affinity was calculated based on the total
intermolecular energy and the predicted free energy of binding. All
experiments were conducted in triplicate, with a root-mean-square
deviation (RMSD) of ≤2 Å.
Statistical analysis
Western blot experimental data were analyzed using
GraphPad Prism software (version 5.0; Dotmatics). The data are
expressed as the means ± standard deviation of triplicate samples.
Statistical significance was determined using one-way analysis of
variance followed by Dunnett's multiple comparison test, comparing
each treatment group with the control. Screening of antioxidant
potential data are presented as the mean ± standard deviation of
triplicate samples. Two-way ANOVA was used for statistical
analysis, followed by Tukey's HSD test. Means with different
superscripts (A-E) within the same column and different
superscripts (a-c) within the same row indicate significant
differences at P<0.05. P<0.05 was considered to indicate a
statistically significant difference.
Results and Discussion
Separation and characterization of
polyphenol compounds in BC
HPLC-MS/MS was used to qualitatively analyze the
compounds present in the BC extract. UV-visible spectroscopy and
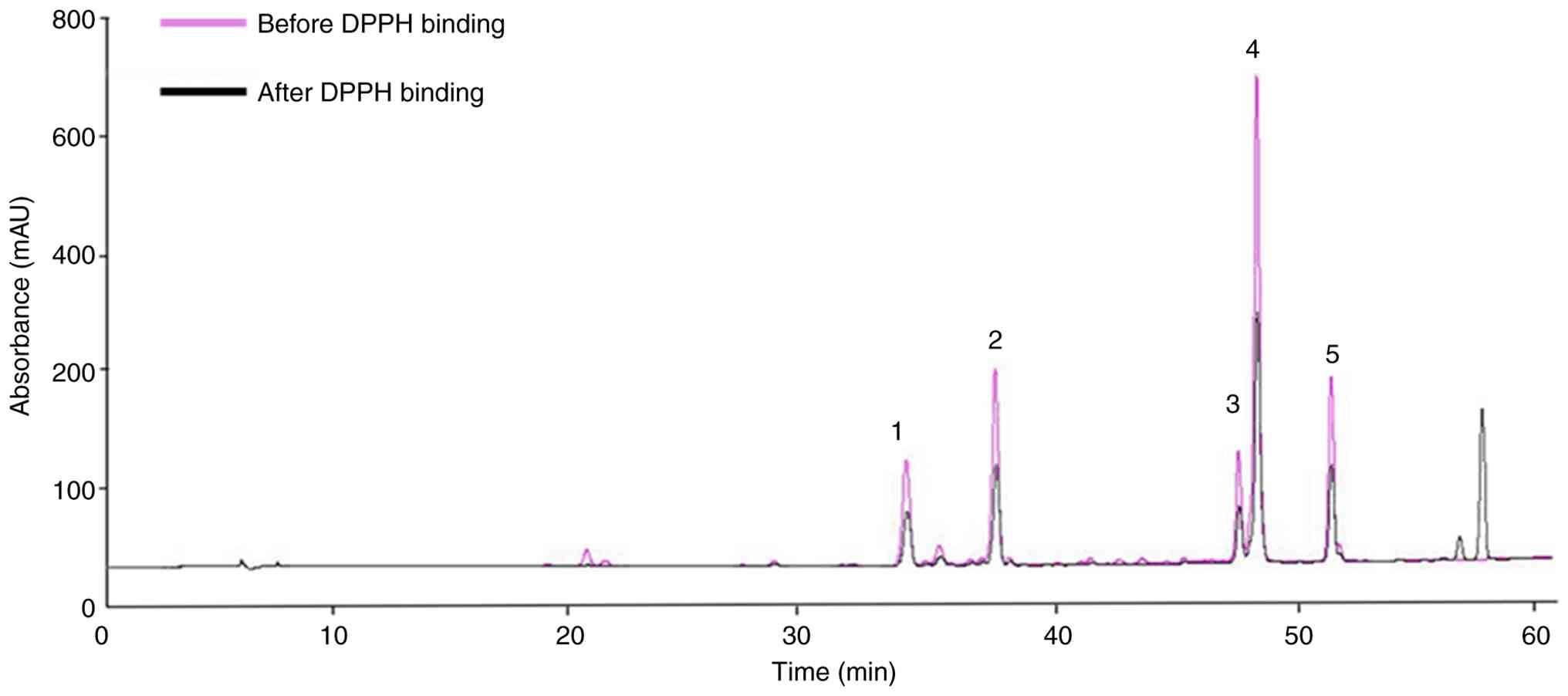
HPLC retention time yielded a total of five major peaks (Fig. 1). HPLC was used to identify the
five compounds contained in the extract at a wavelength of 284 nm.
Although the present study did not use authentic reference
standards, the identified compounds were confirmed by matching the
observed MS/MS spectra with SCIEX OS 3.0.0 system library data.
Additionally, the fragmentation patterns of the precursor ions were
carefully analyzed and compared with previously published
literature to ensure accurate compound identification. Previous
reports have demonstrated biological functions of these compounds
and, notably, several have been investigated for antitumor
potential. For example, tectoridin and tectorigenin have been
reported to exhibit antiproliferative effects in breast and
hepatocellular carcinoma models, while genistein is a well-studied
isoflavone with known chemopreventive activity through modulation
of estrogen signaling and induction of apoptosis. Iridin and
irisflorentin have also shown anti-inflammatory and
anticancer-related bioactivities in vitro, suggesting that
the presence of these metabolites in BC may contribute to its
potential therapeutic properties (34,35).
Incorporating quantitative methodologies such as high performance
LC-based quantification using authenticated reference standards
would allow more precise determination of metabolite concentrations
and could complement the qualitative approach used in the present
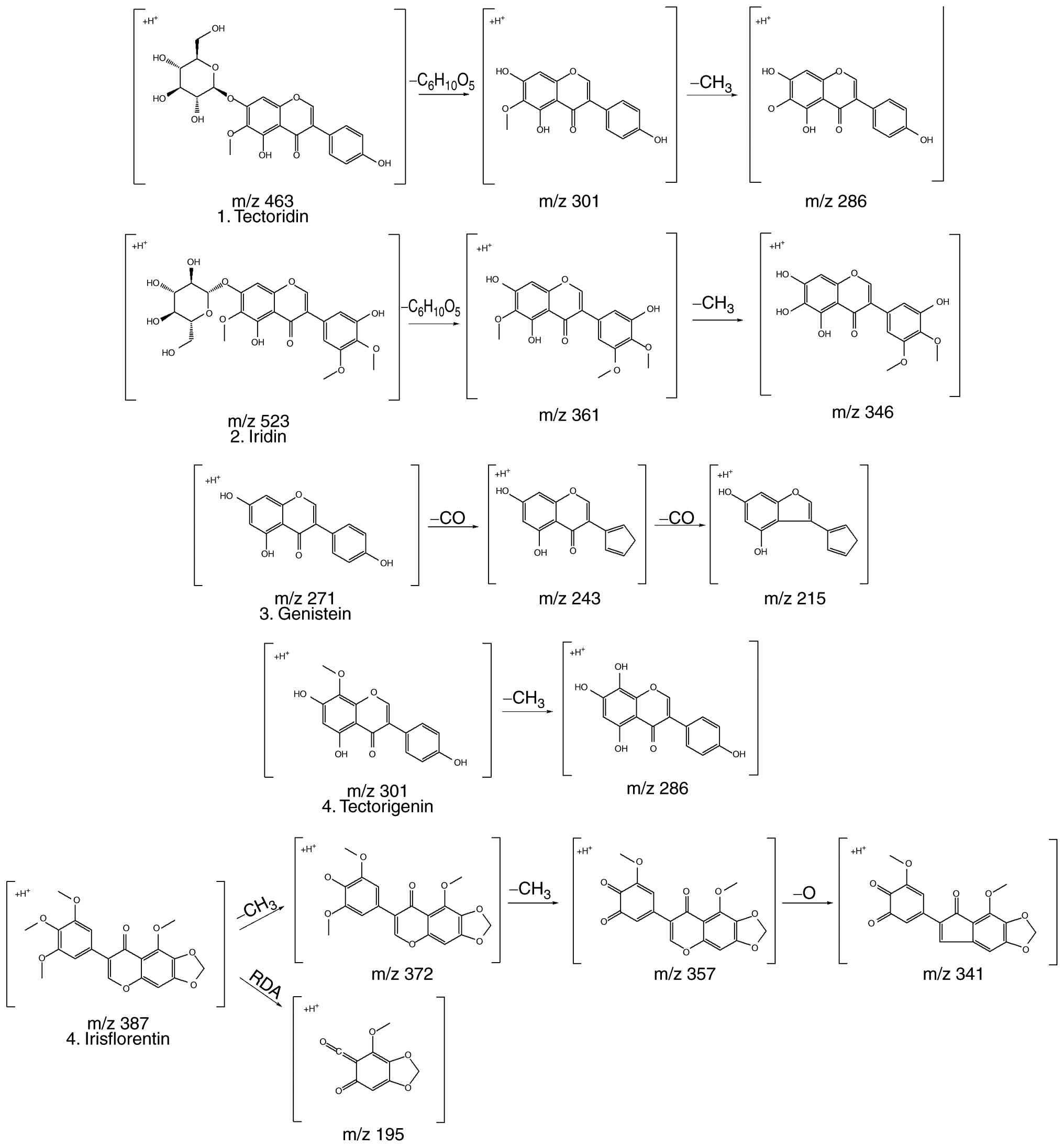
study. The five phenolic compounds identified by MS, referred to in
published sources, are shown in Table
I. The five compounds identified, included tectoridin (36-38),
iridin (36,37,39),
genistein (40,41), tectorigenin (37,38)
and irisflorentin (42,43). The purpose of this
conceptualization was to characterize the compounds found and
analyze the data using the molecular ions and mass patterns found
in the LC-MS/MS data. Fig. 2 shows
the results of predicting the fragmentation of the compounds based
on these results; this fragmentation pattern was the basis for the
compound formation. Therefore, the structural properties of the
phenolic compounds found in this extract were investigated.
 | Table IHigh-performance liquid
chromatography-MS/MS data of phenolic compounds from Belamcanda
chinensis (L.) DC. |
Table I
High-performance liquid
chromatography-MS/MS data of phenolic compounds from Belamcanda
chinensis (L.) DC.
| Peak No. | Rt, min | Formula | Compound | UV max (nm) | Precursor ion
[M+H]+ | MS/MS | (Refs.) |
|---|
| 1 | 34.20 |
C22H22O11 | Tectoridin | 263, 331 | 463 | 301
(C16H12O6)
[M+H-C6H12O5]+, 286
(C15H10O6)
[M+H-CH2]+ | (36-38) |
| 2 | 38.14 |
C24H26O13 | Iridin | 264 | 523 | 361
(C18H16O8)
[M+H-C6H12O5]+, 346
(C17H14O8)
[M+H-CH2]+ | (36,37,39) |
| 3 | 48.40 |
C15H10O5 | Genistein | 327, 259 | 271 | 243
(C14H10O4) [M+H-CO], 215
(C13H10O3) [M+H-CO] | (40,41) |
| 4 | 48.74 |
C16H12O6 | Tectorigenin | 338, 269 | 301 | 286
(C15H10O6)
[M+H-CH2]+ | (37,38) |
| 5 | 52.81 |
C20H18O8 | Irisflorentin | 321, 266 | 387 | 372
(C19H15O8)
[M+H-CH3]+, 357
(C18H12O8)
[M+H-CH3]+, 341
(C18H12O7) [M+H-O]+,
195 (C9H6O5)
[M+H-RDA]+ | (42,43) |
Screening of antioxidant polyphenolic
compounds in BC extract
The DPPH reagent is mainly used to analyze radical
scavenging or antioxidant effects of natural products. Antioxidants
provide electrons to free radicals, causing a color change of the
reagent from purple to yellow, which indicates its antioxidant
capacity (44). Screening
candidates for potential antioxidant effects among polyphenol
compounds from BC extract was conducted by combining DPPH and HPLC
methods. HPLC peak area values were compared and analyzed before
and after the reaction between the polyphenol components and DPPH.
According to the HPLC-MS/MS results after reacting DPPH with the BC
extract, all selected polyphenol compounds in the extract displayed
changes in peak area values, indicating their potential
contribution to the antioxidant effect. Table II shows that the decrease in peak
area after the reaction demonstrates the antioxidant effect of the
polyphenol compounds, which reacted competitively with DPPH. The
difference in area values (%) indicates a notable radical
scavenging ability, and the difference between the reactive area of
DPPH for each compound was as follows: Tectoridin (1,804.71 mAU),
iridin (2,725.72 mAU), genistein (1,217.06 mAU), tectorigenin
(6,182.29 mAU) and irisflorentin (2,027.54 mAU). Furthermore,
post-hoc tests revealed statistically significant differences among
the five compounds. The compounds that reacted notably with DPPH
were tectorigenin and iridin; taken together, these initial
screening results suggest that tectorigenin and iridin may
represent particularly promising antioxidant candidates, although
further validation is required to substantiate their potential. The
present analytical approach has inherent limitations, as DPPH-based
assays do not fully recapitulate physiological redox conditions
(45). To address this limitation,
additional experiments evaluating the inhibition of induced
intracellular reactive oxygen species production may provide more
biologically relevant evidence and could be incorporated to
strengthen future investigations.
| Table IIScreening of antioxidant potential of
Belamcanda chinensis (L.) DC. compounds. |
Table II
Screening of antioxidant potential of
Belamcanda chinensis (L.) DC. compounds.
| Peak no. | Compound | Before DPPH area
absorbance (mAU) | After DPPH binding
area absorbance (mAU) | Reactive area
absorbance (mAU) |
|---|
| 1 | Tectoridin |
3,720.58±6.03Bc |
1,915.86±1.40Bb |
1,804.71±7.33Ba |
| 2 | Iridin |
5,710±2.18Dc |
2,984.28±1.27Dd |
2,725.72±2.26Da |
| 3 | Genistein |
2,484.44±3.48Ac |
1,267.38±2.37Ab |
1,217.06±4.36Aa |
| 4 | Tectorigenin |
13,145.37±3.92Ec |
6,963.09±2.15Eb |
6,182.29±6.02Ea |
| 5 | Irisflorentin |
4,425.03±2.45Cc |
2,396.49±2.46Cb |
2,027.54±2.03Ca |
Effects of BC extract on the viability
of HaCaT cells
The cytotoxicity of the extract was evaluated using
a MTT assay on the HaCaT cells (Fig.
3). HaCaT cells were treated with BC extract at various
concentrations: 0, 0.5, 1, 2.5, 5, 7.5, 10, 12.5, 25, 50, 75 and
100 µg/ml, both with and without 1 µg/ml LPS. LPS was included to
establish an inflammation-stimulated cellular model and to assess
whether BC extract exerted cytotoxic effects under inflammatory
conditions. The results indicated that the BC extract was non-toxic
at ~75 µg/ml. Consequently, the concentrations of 0.5, 1 and 2
µg/ml were selected for further experiments.
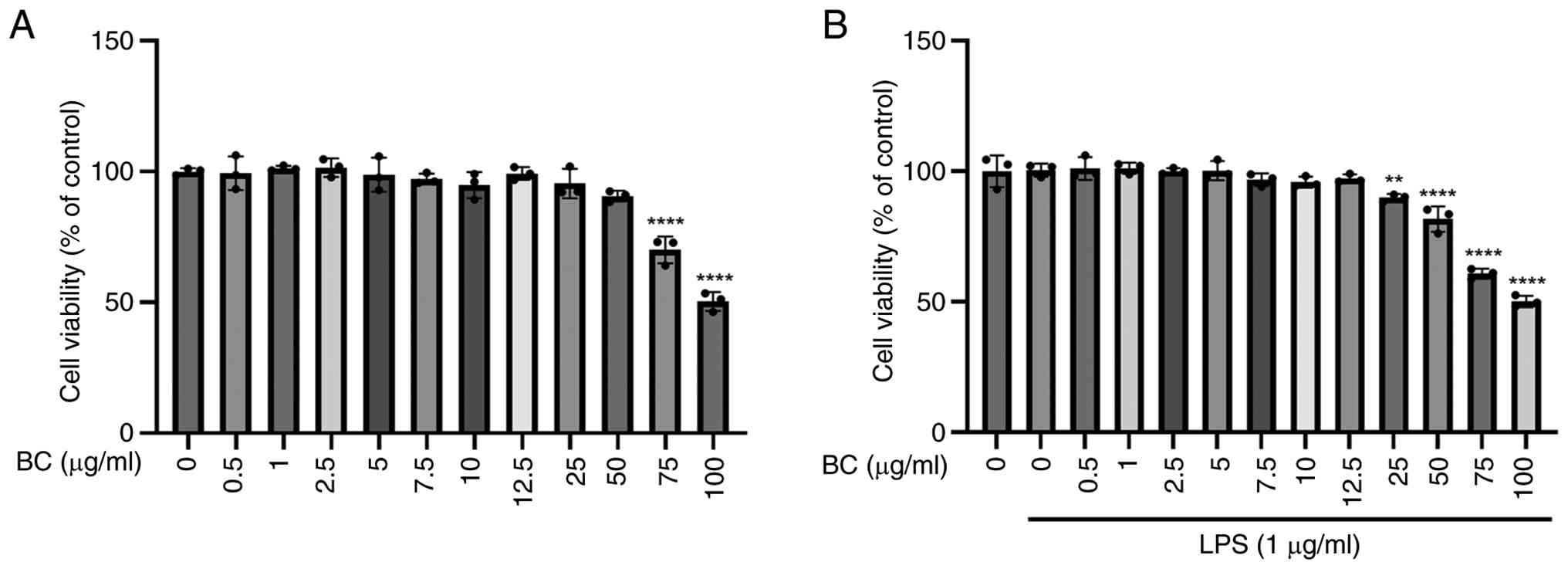 | Figure 3Effects of BC extract on HaCaT cell
viability. (A) HaCaT cells were treated with BC (0, 0.5, 1, 2.5, 5,
7.5, 10, 12.5, 25, 50, 75 and 100 µg/ml) for 24 h.
****P<0.0001 vs. untreated group, respectively. (B)
HaCaT cells were treated with BC (0, 0.5, 1, 2.5, 5, 7.5, 10, 12.5,
25, 50, 75 and 100 µg/ml) with or without LPS (1 µg/ml) for 24 h.
**P<0.01; ****P<0.0001 vs.
LPS-alone-treated group, respectively. MTT assay showed that BC
extract was non-toxic to HaCaT cells at concentrations up to ~75
µg/ml. Each experiment was independently repeated at least three
times with consistent results. BC, Belamcanda chinensis (L.)
DC.; LPS, lipopolysaccharide. |
BC extract inhibits iNOS and COX-2
expression on LPS-induced HaCaT Cells
Increased levels of COX-2 or iNOS are linked to
certain inflammatory skin disorders in humans, such as atopic
dermatitis and psoriasis, where up-regulation of COX-2 contributes
to inflammatory responses in atopic skin lesions and iNOS
expression is elevated in psoriatic epidermal keratinocytes,
implicating these enzymes in chronic skin inflammation and
pathological changes. Since inflammation is closely associated with
tumor promotion, substances with strong anti-inflammatory
properties are anticipated to have chemopreventive effects on
carcinogenesis, particularly during the promotion stage (46-48).
Therefore, the ability of BC extract to suppress COX-2 and iNOS
expression suggests its potential as an anti-inflammatory and
chemopreventive agent against inflammation-associated skin
disorders. To determine the effects of BC extract on iNOS and
COX-2, the BC extract was administered at concentrations of 0.5, 1
and 2 µg/ml with LPS treatment. The results showed that iNOS and
COX-2 protein expression decreased in a dose-dependent manner
(Fig. 4). Although the current
design provides initial insight, including a positive inhibitory
control, future studies could further validate assay sensitivity
and strengthen the findings by incorporating techniques such as
reverse transcription-quantitative PCR to assess mRNA levels,
enzyme activity assays for iNOS and COX-2, and the use of
additional inflammatory stimuli or cytokine profiling to confirm
anti-inflammatory effects.
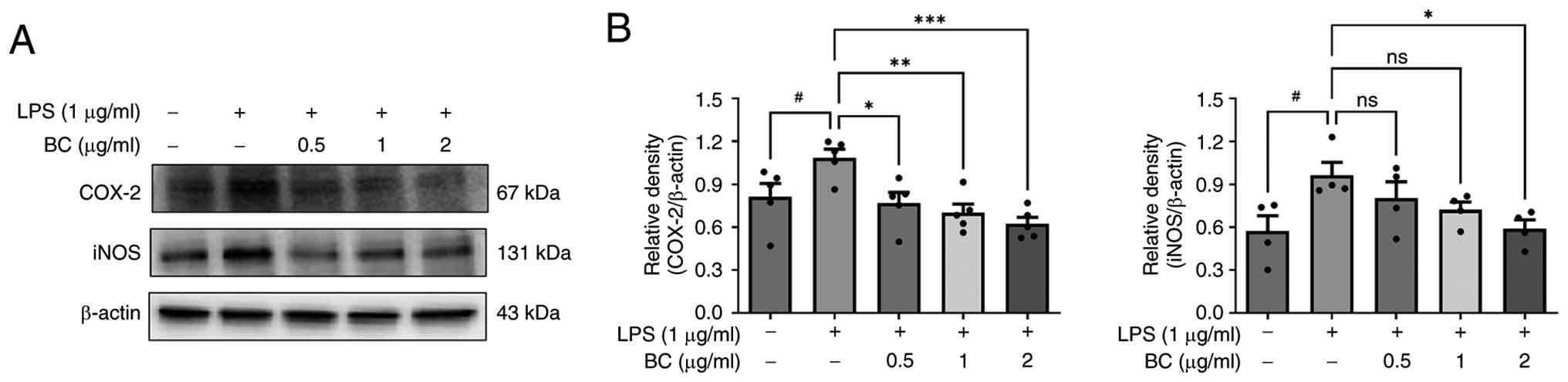 | Figure 4Effect of BC extract on the
expression of COX-2 and iNOS. (A) HaCaT cells were treated with BC
(0, 5, 1 and 2 µg/ml), with or without LPS (1 µg/ml), for 24 h. The
expression levels of COX-2 and iNOS were assessed using western
blotting. BC treatment decreased COX-2 and iNOS levels in a
dose-dependent manner, indicating its anti-inflammatory effects.
(B) The results are presented as the mean ± standard deviation of
three independent experiments. #P<0.05 vs. untreated
group; *P<0.05, **P<0.01,
***P<0.001 vs. LPS alone-treated group. BC,
Belamcanda chinensis (L.) DC.; LPS, lipopolysaccharide;
COX-2, cyclooxygenase-2; iNOS, inducible nitric oxide synthase; ns,
not significant. |
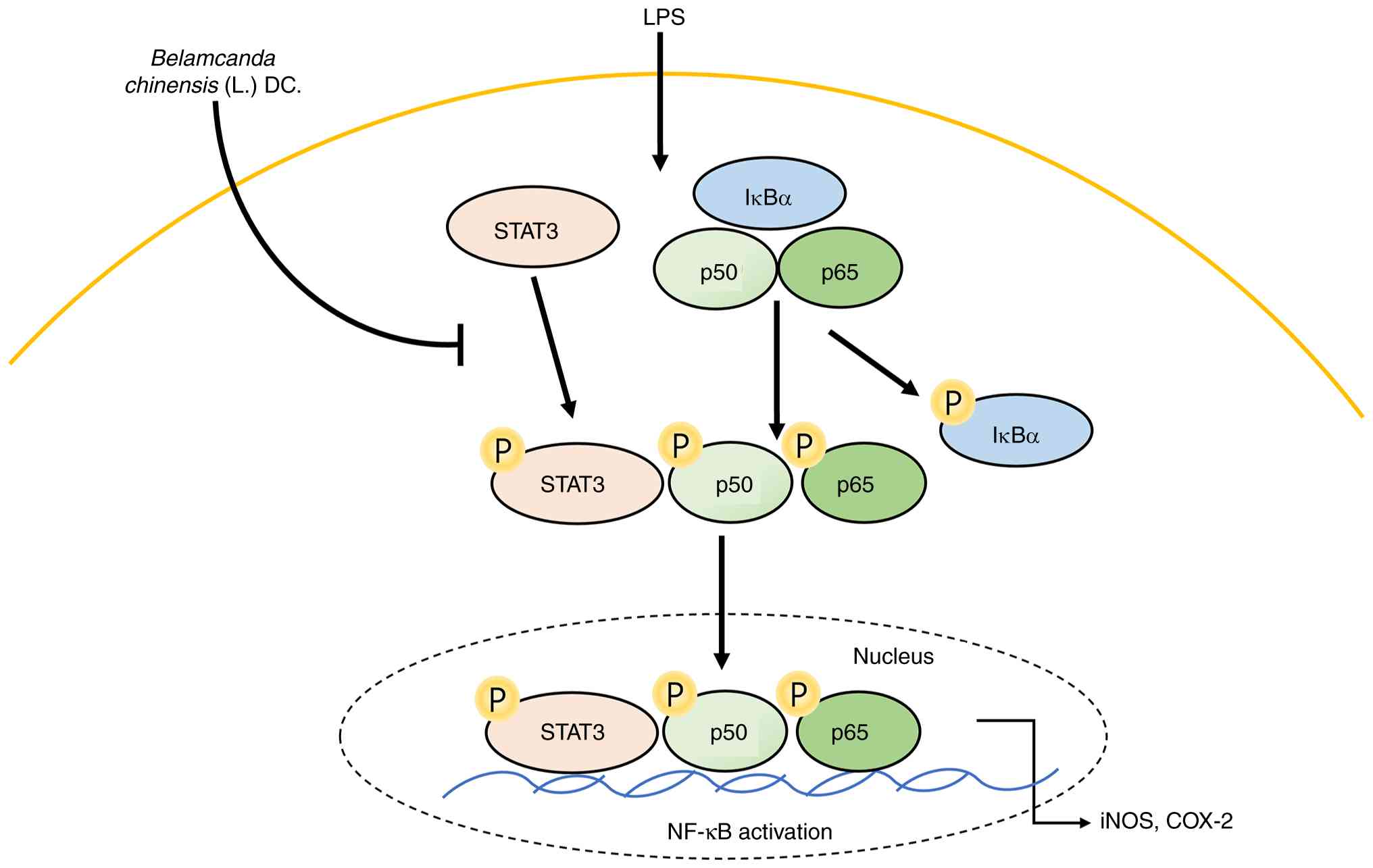
BC extract inhibits STAT3 and NF-кB
expression on LPS-induced HaCaT Cells
Understanding the molecular mechanisms that underlie
the cooperation between the interaction of STAT3 and NF-κB in
inflammation may lead to the development of new chemopreventive and
chemotherapeutic strategies (49).
It was confirmed that the STAT3 and NF-кB activation were decreased
when LPS-induced HaCaT cells were treated with BC extract (Fig. 5). Treatment with BC at
concentrations of 1 and 2 µg/ml with LPS treatment significantly
reduced the levels of p-STAT3, p-p65 and p-IкB-α; in addition,
p-p65 and p-IкB-α levels were also decreased with 0.5 µg/ml BC.
While these data suggest a potential inhibitory effect of BC on
NF-κB signaling, incorporating functional assays such as NF-κB
luciferase reporter activity or p65 nuclear translocation analyses
would provide a more direct assessment of pathway inhibition and
further reinforce the mechanistic interpretation in future studies.
Based on these results, molecular docking between NF-κB and the
five polyphenolic compounds contained in the BC extract was also
conducted.
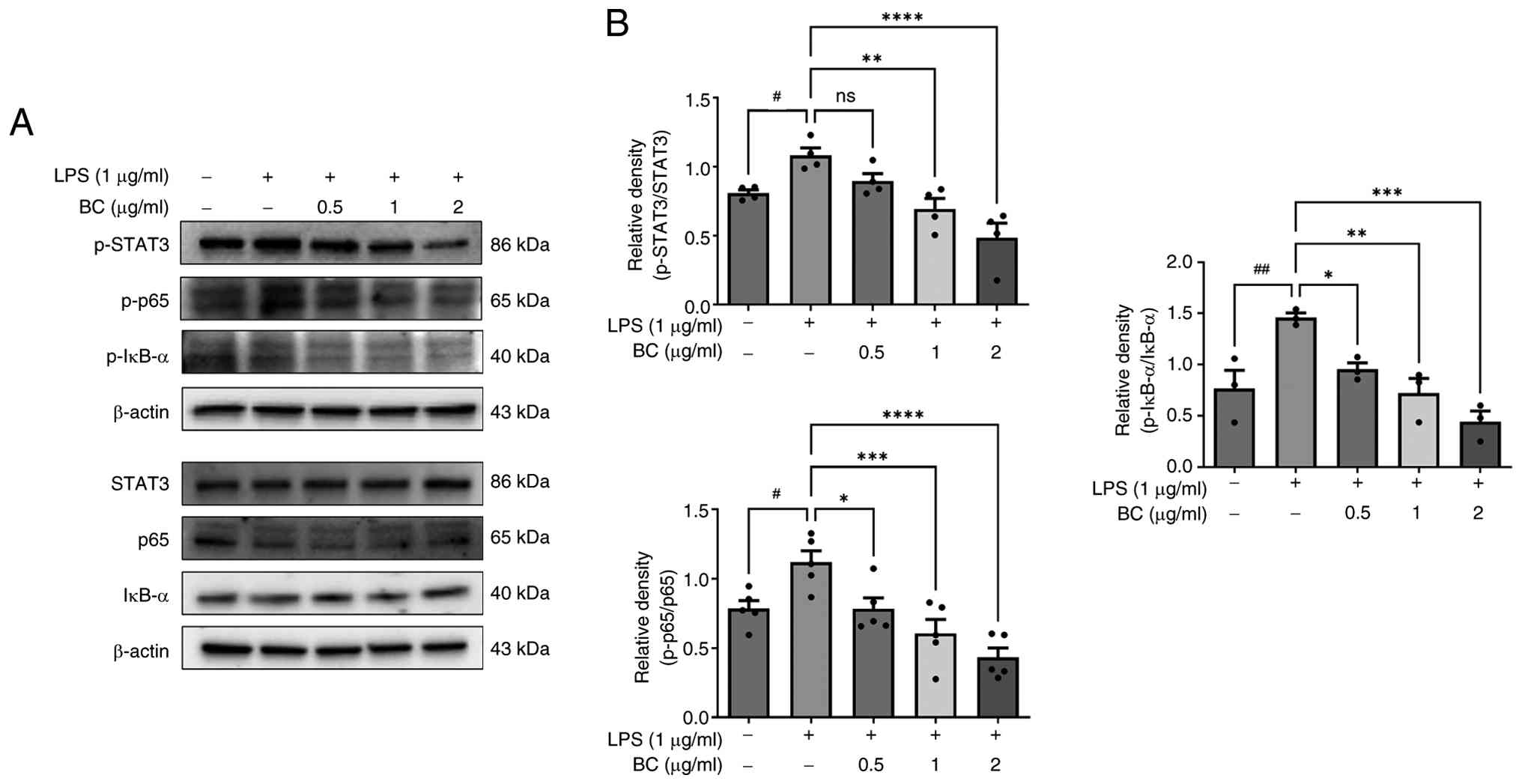 | Figure 5Effect of BC on the expression of
STAT3 and NF-κB. (A) HaCaT cells were treated with BC (0, 5, 1 and
2 µg/ml), with or without LPS (1 µg/ml), for 24 h. The expression
of STAT3, p65 and IκB-α was determined by western blotting. BC
treatment dose-dependently reduced the levels of p-STAT3, p-p65 and
p-IκB-α in LPS-stimulated HaCaT cells. (B) The results are
presented as the mean ± standard deviation of three independent
experiments. #P<0.05 and ##P<0.01 vs.
untreated group; *P<0.05, **P<0.01,
***P<0.001 and ****P<0.0001 vs. LPS
alone-treated group. BC, Belamcanda chinensis (L.) DC.; LPS,
lipopolysaccharide; ns, not significant; STAT3, signal transducers
and activators of transcription 3; NF-κB, nuclear factor-κB. |
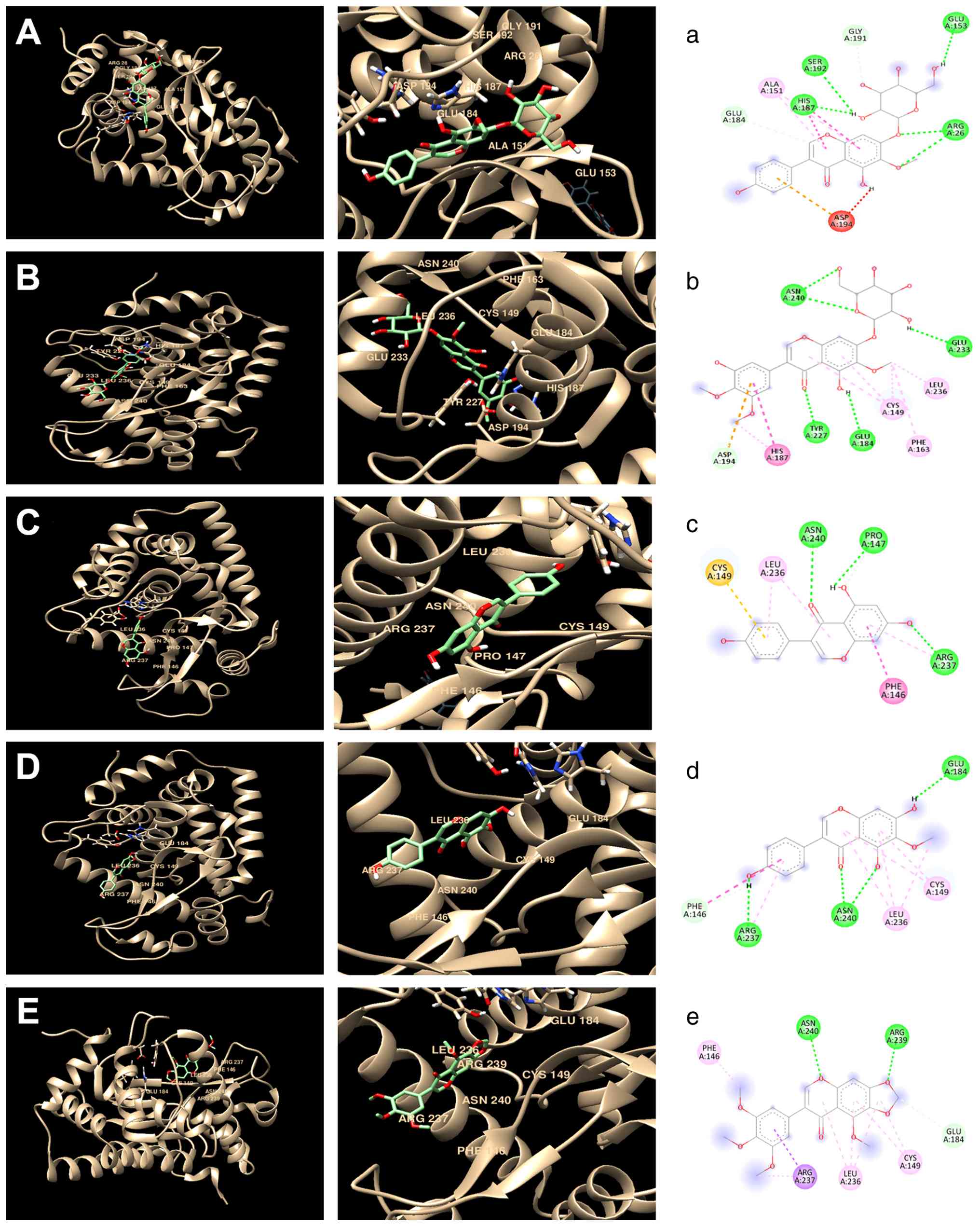
Molecular docking analysis of selected
polyphenol compounds with NF-кB from extracts
The hyperactivation of NF-κB in a cellular system is
linked to inflammation, cancer and various other human diseases;
key components of the NF-kB cascade may regulate the translocation
and activation of the NF-κB transcription factor (50). In the present study, the focus was
on the NF-κB signaling pathway, examining the active sites of its
key components and conducting molecular docking analysis to
understand their interactions and functions. Molecular docking
studies were performed with NF-κB protein complex and the five
polyphenol compounds: Tectoridin, iridin, genistein, tectorigenin
and irisflorentin. Notably, the PDB entry 4Q3J used in the present
study indicated that these compounds primarily bind to the NF-κB
subunit p65 (RelA), with minimal interaction with p50 or IκB-α,
suggesting that the modulatory effects observed are mainly mediated
through p65. The interactions between each protein and ligand with
their complementary surfaces are depicted in Fig. 6. The ligand-protein docking was
performed using the UCSF Chimera program (Fig. 6A-E), whereas the analysis of amino
acid interactions was performed using the Discovery Studio program
(Fig. 6A-E). Table III summarizes the amino acid
residues involved in these interactions and their corresponding
binding energy scores.
| Table IIIMolecular docking of tectoridin,
iridin, genistein, tectorigenin and irisflorentin with the NF-кB
complex and their binding energies. |
Table III
Molecular docking of tectoridin,
iridin, genistein, tectorigenin and irisflorentin with the NF-кB
complex and their binding energies.
| Binding ligand | Amino acid residue
that interacts with NF-κB | Docking score,
kcal/mol |
|---|
| Tectoridin | ASP194, ALA151,
ARG26, GLU184, HIS187, SER192, GLU153 and GLY191 | -8.0 |
| Iridin | ASN240, ASP194,
CYS149, GLU184, GLU233, HIS187, LEU236, PHE163 and TYR227 | -8.3 |
| Genistein | ASN240, ARG237,
CYS149, LEU236, PRO147 and PHE146 | -7.4 |
| Tectorigenin | ASN240, ARG237,
CYS149, GLU184, LEU236 and PHE146 | -7.4 |
| Irisflorentin | ASN240, ARG237,
ARG239, CYS149, GLU184, LEU236 and PHE146 | -6.9 |
The docking analysis identified several active sites
for tectoridin: ASP194, ALA151, ARG26, GLU184, HIS187, SER192,
GLU153 and GLY191; this compound achieved a binding energy score of
-8.0 kcal/mol. In the case of iridin, docking results showed active
sites at ASN240, ASP194, CYS149, GLU184, GLU233, HIS187, LEU236,
PHE163 and TYR227, and had the highest recorded binding energy
score of -8.3 kcal/mol. For genistein, the docking analysis
identified the following active sites: ARG237, ASN240, CYS149,
GLU184, LEU236, PHE146, PRO147 and TYR227, leading to a binding
energy score of -6.9 kcal/mol. Molecular docking of tectorigenin
with NF-κB revealed active sites at ARG232, ALA228, CYS149, LEU236
and TYR227; this compound achieved a binding energy score of -7.4
kcal/mol. Finally, for irisflorentin, the identified active sites
interacting with NF-κB included ARG211, ASP223, ILE208 and LEU203,
with a binding energy score of -6.8 kcal/mol, which was the lowest
among the compounds analyzed. Furthermore, all five compounds
exhibited an RMSD value of 0 Å, indicating that their docked
conformations were identical to the crystallographic reference
structures. This high degree of structural congruence validates the
reliability of the docking simulations and suggests that the
observed interactions are not computational artifacts but rather
reflect true binding potential. Given that RMSD values <2 Å are
typically considered indicative of successful docking poses
(51,52), these results demonstrate the
precision and stability of the predicted binding modes. Taken
together, these findings highlight iridin as the most promising
candidate for further investigation as an NF-κB inhibitor. Its high
binding affinity and favorable interaction profile suggest
potential anti-inflammatory effect and, based on the central role
of NF-κB in cancer-related signaling pathways, possible antitumor
properties through suppression of NF-κB-mediated transcriptional
activity, including the downregulation of pro-survival and
proliferation genes. However, it should be noted that the present
study was performed on skin keratinocytes rather than a cancer cell
line, and thus the antitumor potential of BC extract remains to be
experimentally validated. Future studies could explore the effects
of BC extract and iridin in relevant cancer cell models and in
vivo systems to evaluate its chemopreventive or therapeutic
potential, investigate the underlying molecular mechanisms, and
assess pharmacokinetics and safety profiles (53,54).
However, while in silico docking provides an initial
mechanistic rationale, in vitro and in vivo
validation, such as reporter gene assays, electrophoretic mobility
shift assays or cytokine profiling, will be essential to confirm
its biological efficacy and specificity. Ultimately, the present
study supports the hypothesis that naturally occurring isoflavones,
particularly iridin and tectoridin, may serve as potential lead
compounds for the development of novel NF-κB-targeting therapeutics
aimed at treating inflammation-related disorders or cancers.
In the present study, the aim was to investigate the
anti-inflammatory effects of BC extract on LPS-induced HaCaT cells
(Fig. 7). The present results
showed a decrease in the expression of anti-inflammatory markers,
specifically iNOS and COX-2, along with a significant reduction in
the levels of proteins associated with the STAT3-NF-κB pathway.
Based on these findings, NF-κB was selected as a candidate for the
molecular docking analysis.
In conclusion, the present study confirmed the
mechanism of action of the compounds present in BC through the
simultaneous application of DPPH-HPLC. Through LC-MS/MS analysis,
five compounds: Tectoridin, iridin, genistein, tectorigenin and
irisflorentin were identified and quantified in the polyphenol
extract of BC. The results showed that these compounds effectively
contributed to antioxidant and anti-inflammatory activities by
demonstrating marked reaction amounts and rates (%) as ligands for
DPPH. Furthermore, the BC extract significantly reduced the
expression of iNOS, COX-2, p-STAT3 and NF-κB proteins in
LPS-induced HaCaT cells. The five selected BC compounds showed
marked binding scores during molecular docking with NF-κB, a key
inflammatory transcriptional regulator.
The antioxidant and anti-inflammatory properties of
the compounds found in BC were evaluated to determine which
exhibited the most notable potential. These findings serve as a
valuable resource for identifying potential bioactive compounds
with antioxidant and anti-inflammatory properties from natural
sources. They also provide valuable data for the future application
of BC in the food and pharmaceutical sectors. However, the present
study has certain limitations, as it was conducted solely using the
HaCaT cell line, which may not fully recapitulate the complex
microenvironment of human skin in vivo or reflect the
responses of different cell types present in actual tissue. To
overcome this limitation, it is the aim to include experiments
using normal primary keratinocytes in future studies to further
validate and strengthen the physiological relevance of the present
findings. In addition, the concentration range tested in the
present study was relatively limited, and a more comprehensive dose
response evaluation would provide a clearer understanding of
concentration-dependent effects and further enhance the robustness
of the present conclusions.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the Korea Institute
of Toxicology, Republic of Korea (grant no. 2710086927).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
SEH participated in all aspects of the experimental
design, performed the analysis and wrote the manuscript. HHK
conducted the data analysis investigation, confirms the
authenticity of all the raw data and reviewed and edited the
manuscript. SHJ performed data curation and editing of the
manuscript. GSK confirms the authenticity of all the raw data and
participated in experimental design and project administration. KHH
was responsible for funding acquisition, acquisition of data and
supervision. All authors have read and approved the final version
of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Use of artificial intelligence tools
During the preparation of the present manuscript,
artificial intelligence tools were used to improve the readability
and language of the manuscript, and subsequently, the authors
revised and edited the content produced by the artificial
intelligence tools as necessary, taking full responsibility for the
ultimate content of the present manuscript.
References
|
1
|
Meisser SS, Altunbulakli C, Bandier J,
Opstrup MS, Castro-Giner F, Akdis M, Bonefeld CM, Johansen JD and
Akdis CA: Skin barrier damage after exposure to
paraphenylenediamine. J Allergy Clin Immunol. 145:619–631.e2.
2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Hong M, Xiao K, Lin P and Lin J: Five
Rutaceae family ethanol extracts alleviate
H2O2 and LPS-induced inflammation via NF-κB
and JAK-STAT3 pathway in HaCaT cells. Chin J Nat Med. 20:937–947.
2022.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Slominski RM, Chen JY, Raman C and
Slominski AT: Photo-neuro-immuno-endocrinology: How the ultraviolet
radiation regulates the body, brain, and immune system. Proc Natl
Acad Sci USA. 121(e2308374121)2024.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Slominski AT, Zmijewski MA, Plonka PM,
Szaflarski JP and Paus R: How UV light touches the brain and
endocrine system through skin, and why. Endocrinology.
159:1992–2007. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Medzhitov R: Origin and physiological
roles of inflammation. Nature. 454:428–435. 2008.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Nestle Frank O..Kaplan D and Barker J:
Mechanisms of disease: psoriasis. New England Journal of Medicine.
361.5:496–509. 2009.
|
|
7
|
Weidinger , Stephan and Novak
N: Atopic dermatitis. The Lancet. 387:1109–1122. 2016.
|
|
8
|
Kujubu DA, Fletcher BS, Varnum BC, Lim RW
and Herschman HR: TIS10, a phorbol ester tumor promoter-inducible
mRNA from Swiss 3T3 cells, encodes a novel prostaglandin
synthase/cyclooxygenase homologue. J Biol Chem. 266:12866–12872.
1991.PubMed/NCBI
|
|
9
|
Oshima M, Dinchuk JE, Kargman SL, Oshima
H, Hancock B, Kwong E, Trzaskos JM, Evans JF and Taketo MM:
Suppression of intestinal polyposis in APC716 knockout mice by
inhibition of prostaglandin endoperoxide synthase-2 (COX-2). Cell.
87:803–809. 1996.
|
|
10
|
Subbaramaiah K, Telang N, Ramonetti JT,
Araki R, DeVito B, Weksler BB and Dannenberg AJ: Transcription of
cyclooxygenase-2 is enhanced in transformed mammary epithelial
cells. Cancer Res. 56:4424–4429. 1996.PubMed/NCBI
|
|
11
|
Hong CH, Hur SK, Oh OJ, Kim SS, Nam KA and
Lee SK: Evaluation of natural products on inhibition of inducible
cyclooxygenase (COX-2) and nitric oxide synthase (iNOS) in cultured
mouse macrophage cells. J Ethnopharmacol. 83:153–159.
2002.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Chi C, Ozawa T and Anzai K: In vivo nitric
oxide production and iNOS expression in X-ray irradiated mouse
skin. Biol Pharm Bull. 29:348–353. 2006.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Ganster RW and Geller DA: Molecular
regulation of inducible nitric oxide synthase. In: Nitric Oxide
Elsevier, pp129-156, 2000.
|
|
14
|
Andrés RM, Montesinos MC, Navalón P, Payá
M and Terencio MC: NF-κB and STAT3 inhibition as a therapeutic
strategy in psoriasis: In vitro and in vivo effects of BTH. J
Invest Dermatol. 133:2362–2371. 2013.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Miao X, Xiang Y, Mao W, Chen Y, Li Q and
Fan B: TRIM27 promotes IL-6-induced proliferation and inflammation
factor production by activating STAT3 signaling in HaCaT cells. Am
J Physiol Cell Physiol. 318:C272–C281. 2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Li Q and Verma IM: NF-kappaB regulation in
the immune system. Nat Rev Immunol. 2:725–734. 2002.PubMed/NCBI View
Article : Google Scholar
|
|
17
|
Chiang YM, Lo CP, Chen YP, Wang SY, Yang
NS, Kuo YH and Shyur LF: Ethyl caffeate suppresses NF-kappaB
activation and its downstream inflammatory mediators, iNOS, COX-2,
and PGE2 in vitro or in mouse skin. Br J Pharmacol. 146:352–363.
2005.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Bayon Y, Ortiz MA, Lopez-Hernandez FJ, Gao
F, Karin M, Pfahl M and Piedrafita FJ: Inhibition of IkappaB kinase
by a new class of retinoid-related anticancer agents that induce
apoptosis. Mol Cell Biol. 23:1061–1074. 2003.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Baker RG, Hayden MS and Ghosh S: NF-κB,
inflammation, and metabolic disease. Cell Metab. 13:11–22.
2011.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Carpenter RL and Lo HW: STAT3 target genes
relevant to human cancers. Cancers (Basel). 6:897–925.
2014.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Moriyasu M, Igi Y, Ichimaru M, Iwasa K,
Kobayakawa J, Sato-Nishimori F, Matsukawa Y and Nagase C: New
isoflavones from Belamcandae Rhizoma. J Nat Med. 61:329–333.
2007.
|
|
22
|
Xin RH, Zheng JF, Cheng L, Peng WJ and Luo
YJ: Belamcanda chinensis (L.) DC: Ethnopharmacology and
pharmacology review, including anti-inflammatory activities.
African Journal of Traditional, Complementary and Alternative
Medicines, 2015.
|
|
23
|
Xin RH, et al: Belamcanda chinensis is
used in the clinical treatment of respiratory diseases including
bronchial asthma in traditional Chinese medicine practices. Afr J
Tradit Complement Altern Med, 2015.
|
|
24
|
Zhang et al: Traditional uses of
Belamcanda chinensis include soothing the throat and treating cough
and pharyngitis in Chinese medicine; rhizome extracts exhibit
anti-inflammatory effects relevant to these conditions. Plants
(Basel), 2024.
|
|
25
|
Chen JJ, Wu ML, Hwang TL, Wu WJ, Tsai YH
and Chen YJ: Anti-inflammatory natural products from Belamcanda
chinensis. Planta Med. 79(PI26)2013.
|
|
26
|
Liu KD, Yang WQ, Dai MZ, Xu Y, Qin YP,
Dong YY, Fu J and Qu J: Phenolic constituents with
anti-inflammatory and cytotoxic activities from the rhizomes of
Iris domestica. Phytochemistry. 203(113370)2022.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Chihomvu P, Ganesan A, Gibbons S, Woollard
K and Hayes MA: Phytochemicals in drug discovery-a confluence of
tradition and innovation. Int J Mol Sci. 25(8792)2024.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Boța M, Vlaia L, Jîjie AR, Marcovici I,
Crişan F, Oancea C, Dehelean CA, Mateescu T and Moacă EA: Exploring
synergistic interactions between natural compounds and conventional
chemotherapeutic drugs in preclinical models of lung cancer.
Pharmaceuticals (Basel). 17(598)2024.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Xin RH, Zheng JF, Cheng L, et al:
Belamcanda chinensis (L.) DC.: Ethnopharmacology, phytochemistry,
and pharmacology. African Journal of Traditional, Complementary and
Alternative Medicines. 12:135–145. 2015.
|
|
30
|
Zhang Y, Wang X, Chen J, et al:
Traditional uses, phytochemistry, and pharmacological.
|
|
31
|
Li J..Zhao Y..Wu X..Chen L..Li X..Zhao J:
Preparative isolation and purification of seven isoflavones from
Belamcanda chinensis by high-speed counter-current chromatography.
Journal of Chromatography A,. 1218(42):7448–7453. 2011.PubMed/NCBI View
Article : Google Scholar
|
|
32
|
Kim YS, Jung JH, Kim NJ and Lee SH:
Anti-angiogenic and anti-tumor activities of isoflavonoids isolated
from the rhizomes of Belamcanda chinensis. Planta Medica,.
69:617–622. 2003.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Kim HH, Ha SE, Vetrivel P, Bhosale PB, Kim
SM and Kim GS: Potential antioxidant and anti-inflammatory function
of Gynura procumbens polyphenols ligand. Int J Mol Sci.
22(8716)2021.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Peng F, Xie X and Cheng P: Chinese herbal
medicine-based cancer therapy: Novel anticancer agents targeting
MicroRNAs to regulate tumor growth and metastasis. The American
Journal of Chinese Medicine. 47.08:1711–1735. 2019.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Spagnuolo Carmela, et al: Genistein and
cancer: current status, challenges, and future directions. Advances
in nutrition. 6.4:408–419. 2015.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Li J, Li WZM, Huang W, Cheung AWH, Bi CWC,
Duan R, Guo AJY, Dong TTX and Tsim KWK: Quality evaluation of
Rhizoma Belamcandae (Belamcanda chinensis (L.) DC.) by using
high-performance liquid chromatography coupled with diode array
detector and mass spectrometry. J Chromatogr A. 1216:2071–2078.
2009.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Zhou H, Zhang Y, Liang H, Song H, Zhao J,
Liu L, Zeng J, Sun L, Ma S and Meng D: A novel multidimensional
strategy to evaluate Belamcanda chinensis (L) DC and Iris
tectorum Maxim based on plant metabolomics, digital reference
standard analyzer and biological activities evaluation. Chin Med.
16(85)2021.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Chen YJ, Liang ZT, Zhu Y, Xie GY, Tian M,
Zhao ZZ and Qin MJ: Tissue-specific metabolites profiling and
quantitative analyses of flavonoids in the rhizome of Belamcanda
chinensis by combining laser-microdissection with
UHPLC-Q/TOF-MS and UHPLC-QqQ-MS. Talanta. 130:585–597.
2014.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Zhang YY, Wang Q, Qi LW, Qin XY and Qin
MJ: Characterization and determination of the major constituents in
Belamcandae Rhizoma by HPLC-DAD-ESI-MS(n). J Pharm Biomed Anal.
56:304–314. 2011.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Rostagno MA, Manchón N, Guillamón E,
García-Lafuente A, Villares A and Martínez JA: Methods and
techniques for the analysis of isoflavones in foods. Chromatography
types, techniques and methods. Nova Science Publishers Inc,
Hauppauge, New York, pp157-198, 2010.
|
|
41
|
Coldham NG, Howells LC, Santi A,
Montesissa C, Langlais C, King LJ, Macpherson DD and Sauer MJ:
Biotransformation of genistein in the rat: Elucidation of
metabolite structure by product ion mass fragmentologyn. J Steroid
Biochem Mol Biol. 70:169–184. 1999.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Zhang X, Qiao GX, Zhao GF and Zhao SF:
Characterization of the metabolites of irisflorentin by using
ultra-high performance liquid chromatography combined with
quadrupole/orbitrap tandem mass spectrometry. J Pharm Biomed Anal.
203(114222)2021.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Liu Y and Guo MQ: Irisflorentin: Advances
on resources, metabolism, and pharmacological activities. In:
Handbook of Dietary Flavonoids Springer, pp1-15, 2023.
|
|
44
|
Kedare SB and Singh RP: Genesis and
development of DPPH method of antioxidant assay. J Food Sci
Technol. 48:412–422. 2011.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Sharma OP and Tej KB: DPPH antioxidant
assay revisited. Food chemistry. 113:1202–1205. 2009.
|
|
46
|
Surh YJ, Chun KS, Cha HH, Han SS, Keum YS,
Park KK and Lee SS: Molecular mechanisms underlying chemopreventive
activities of anti-inflammatory phytochemicals: Down-regulation of
COX-2 and iNOS through suppression of NF-kappa B activation. Mutat
Res. 480-481:243–268. 2001.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Bruch-Gerharz D, et al: A proinflammatory
activity of interleukin 8 in human skin: expression of the
inducible nitric oxide synthase in psoriatic lesions and cultured
keratinocytes. The Journal of Experimental Medicine. 184:2012.
|
|
48
|
Suleman M, et al: Phytocompounds in
Precision Dermatology: COX-2 Inhibitors as a Therapeutic Target in
Atopic-Prone Skin. Biomolecules. 15(998)2025.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Fan Y, Mao R and Yang J: NF-κB and STAT3
signaling pathways collaboratively link inflammation to cancer.
Protein Cell. 4:176–185. 2013.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Cheemanapalli S, Chinthakunta N, Shaikh
NM, Shivaranjani V, Pamuru RR and Chitta SK: Comparative binding
studies of curcumin and tangeretin on up-stream elements of NF-kB
cascade: A combined molecular docking approach. Netw Model Anal
Health Inform Bioinform. 8(15)2019.
|
|
51
|
Wang S, et al: Docking-based virtual
screening of TβR1 inhibitors: evaluation of pose prediction and
scoring functions. BMC chemistry. 14(52)2020.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Wang Z, et al: Comprehensive evaluation of
ten docking programs on a diverse set of protein-ligand complexes:
the prediction accuracy of sampling power and scoring power.
Physical Chemistry Chemical Physics. 18:12964–12975.
2016.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Ma Q, et al: Versatile function of NF-ĸB
in inflammation and cancer. Experimental Hematology & Oncology.
13(68)2024.PubMed/NCBI View Article : Google Scholar
|
|
54
|
DiDonato JA, Frank M and Michael K: NF‐κB
and the link between inflammation and cancer. Immunological
reviews. 246:379–400. 2012.PubMed/NCBI View Article : Google Scholar
|