Introduction
Atopic patients are a specific group of individuals
who produce immunoglobulin E (IgE) antibodies in response to
allergens in the environment. These patients often suffer from one
or more atopic diseases, such as atopic dermatitis, asthma or
allergic rhinitis (AR). When atopic patients inhale allergens, they
develop AR, a non-infectious, chronic inflammatory disease of the
nasal mucosa mediated primarily by IgE (1). AR has a high prevalence worldwide,
reaching as high as 50% in some developed countries. It not only
affects adults, but also children and adolescents (1). Although AR is not a serious disease,
it can serve as the basis for several complications and is a major
risk factor for patients with poorly controlled asthma.
Statistically, perennial AR has no less impact on quality of life
than asthma, but has notable negative consequences for the
productivity of society as a whole (2,3). The
prevalence of AR is rising globally due to factors such as
increased urbanization, a decrease in the amount of time spent
outdoors and the misuse of antibiotics. In addition, lifestyle
changes, increased air pollution, fungi, infectious agents,
tobacco, smog and other risk factors have increased the prevalence
of AR, leading to its worsened impact (4). At present, the etiology of AR is not
well understood, and there are no effective strategies to prevent
the disease (5). The existing
therapeutic options cannot completely cure the disease. Therefore,
conducting more in-depth research into its pathogenesis and
identifying new targets for preventing and treating AR are of
notable importance.
Chemically known as 3,3',4',5,7-pentahydroxyflavone,
quercetin (QUE), a naturally occurring flavonoid, is widely found
in vegetables, fruits and medicinal herbs and has notable
therapeutic potential, including anti-inflammatory, anticancer,
antioxidant, neuroprotective, anti-allergic and antibacterial
properties (6,7). QUE may serve a key role in AR
treatment by modulating multiple key targets (8,9). The
T helper 1 (Th1)/Th2 immune cell imbalance can be regulated by QUE,
which reduces IgE and histamine expression and inhibits the
infiltration of eosinophils, macrophages and lymphocytes (10). Toll-like receptors (TLRs) are
closely associated with innate immunity (3). The TLR4 signaling pathway can
activate the downstream inflammatory NF-κB signaling pathway,
leading to the release of inflammatory cytokines and chemokines
(11). The inflammatory response
can be markedly attenuated by inhibiting activation of the
TLR4/NF-κB signaling pathway (12). In the present study, the aim was to
explore the therapeutic effects of QUE on AR via the TLR4/myeloid
differentiation primary response 88 (MyD88)/interleukin-1
receptor-associated kinase 4 (IRAK4) signaling pathway, and to
investigate the effects of QUE on the inflammatory response and
immune regulation in AR using a mouse model of AR.
Materials and methods
Reagents
In the present study, the following reagents were
used: QUE (Puxi), dexamethasone (DEX; MilliporeSigma), ovalbumin
(OVA; MilliporeSigma; grade III), TLR4 and MyD88 (cat. nos. AF8187
and AF7524, respectively; Beyotime Biotechnology), NF-κB p65 (cat.
no. 8242; Cell Signaling Technology, Inc.), IRAK4 (cat. no.
ab119942, Abcam), BCA kit (Thermo Fisher Scientific, Inc.), SYBR
Green qPCR Master Mix and RT Master Mix for quantitative PCR (qPCR;
MedChemExpress), ELISA kit OVA-IgE (cat. no. MOFI01017; Assay
Genie), mouse IL-17 ELISA Kit (cat. no. CSB-E04608m; Cusabio
Technology, LLC), mouse IL-1β ELISA Kit, mouse IL-10 ELISA Kit,
mouse IL-13 ELISA Kit, mouse IL-4 ELISA Kit (cat. nos. 1210122,
1211002, 1211302 and 1210402, respectively; all Dakwei),
Flow-through antibody FITC Rat anti-mouse CD4 antibody, APC Rat
anti-mouse CD25 antibody, PerCP Rat anti-mouse FOXP3 antibody (cat.
nos. 553055, 552880 and 563902; all BD Biosciences) and Cell
Fixation Membrane Breaker (eBioscience; Thermo Fisher Scientific,
Inc.).
Experimental instruments
The following instruments were used in the present
study: SDS-PAGE electrophoresis system, western blot transfer
system, ChemiDoc MP multifunctional chemiluminescence imaging
system (all Bio-Rad Laboratories, Inc.), BD FACSCanto II flow
cytometer (BD Biosciences), Enzyme labeler SpectraMax Paradigm
MultiMode Microplate Reader (Molecular Devices, LLC).
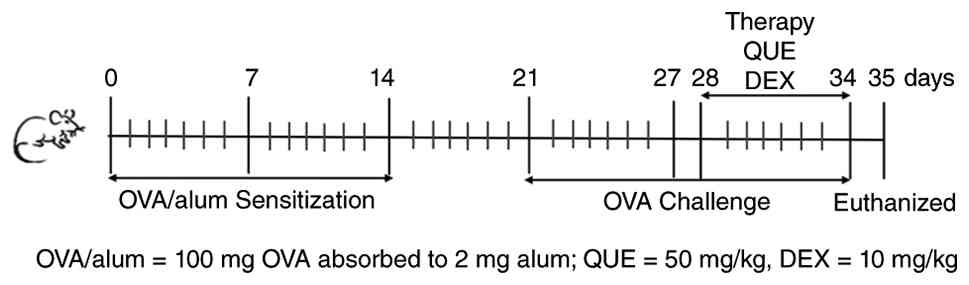
Animal study
In the present study, a total of 72 female BALB/c
mice aged 4-6 weeks old were purchased from the Guangdong
Experimental Animal Center [animal use license no. SCXK (Guangdong)
2022-0002; certificate of conformity no. 44007200114118]. The mice
were housed in an air-conditioned animal room (temperature, 25±1˚C;
humidity, 65±5%) for 1 week to allow them to acclimate. The present
study was performed in accordance with the ARRIVE guidelines for
animal research and complied with all relevant regulations. All
experimental protocols used in the present study complied with
animal ethics standards and were approved by the Experimental
Animal Ethics Committee of Shenzhen Longgang E.N.T Hospital
(approval no. 2022-0426). All efforts were made to minimize animal
suffering. All animals were euthanized in accordance with the
approved protocol at the end of the experiment. No unexpected
deaths were observed, and no premature euthanasia procedures were
carried out during the entire study period. Premature euthanasia
procedures would be initiated if any of the following conditions
occurred: Animals are unable to feed themselves, suffer from severe
dehydration, severe infection, a moribund state, respiratory
distress or cyanosis.
Establishment of the animal model
Previous studies have shown that female sex hormones
increase the likelihood of developing AR and exacerbate
eosinophilic inflammation in the nasal mucosa (13-15).
Therefore, a mouse model of AR was established using female BALB/c
mice. The mice were randomly assigned to one of four groups: Normal
control (NC), OVA, DEX and QUE, with 6 mice per group. A total of
24 mice were used in each independent repeated experiment, and 3
independent repeated animal experiments were conducted. Except for
the NC group, the remaining groups were sensitized with 200 µl
sensitization working solution [containing 2 mg Al(OH)3
and 100 µg OVA] by intraperitoneal injection on days 0, 7 and 14
for basal sensitization, and challenged on day 21 by nasal drip (10
µl per nostril) with OVA dissolved in saline (10 µg/µl) once daily
for 7 days. The treatment of the NC group was replaced with normal
saline, and mice were treated using the same methodology.
Behavioral assessment
The mouse model was expected to show symptoms such
as sneezing, rubbing, face scratching and a runny nose after a
successful challenge; the success of the model was assessed by
observing these symptoms. On the final day of the 7-day challenge,
the behavioral assessment was initiated within 30 min of
completion. Allergy symptoms (such as sneezing, scratching and a
runny nose) were scored after the challenge, and the observation
period lasted 30 min. Scoring was performed based on the criteria
outlined in Table I. The total
score was recorded using the superposition method, and a score of
>5 was considered successful modeling.
 | Table ICriteria for scoring allergic
symptoms in ovalbumin-induced mouse model. |
Table I
Criteria for scoring allergic
symptoms in ovalbumin-induced mouse model.
| Biological
scoring | Scratching | Sneezing | Runny nose |
|---|
| 0 | None | None | None |
| 1 | Occasionally (1-2
times) | 1-3 | Flow to anterior
nostril |
| 2 | Frequently | 4-10 | Beyond the anterior
nostril |
| 3 | Scratching
incessantly | >10 | Runny nose all over
the face |
Treatment of animal models
The treatment groups (DEX and QUE) received 10 mg/kg
DEX or 50 mg/kg QUE by gavage in the afternoon of the day of
challenge, after successful modeling, once daily for 7 days. The NC
and OVA groups were treated by gavage with equal amounts of saline.
Mice in all groups were further sensitized with nasal drops as
before, and their behavioral changes were observed during the 7
days of treatment. Efficacy was evaluated by analyzing symptom
scores in each group during the treatment period. The mice were
anesthetized with isoflurane on day 35 (5% induction and 2%
maintenance). Once the mice completely lost the ability to right
themselves, their respiratory rate markedly decreased or they
showed no response to painful stimuli, they were euthanized by
cervical dislocation. Subsequently, blood from the eye, nose, lungs
and spleen was collected for subsequent analysis (Fig. 1).
ELISA-based detection of OVA-IgE,
IL-4, IL-13, IL-1β, IL-17 and IL-10 in the serum of mice
Blood from the eye was collected, left at room
temperature for 30 min, then centrifuged at 12,880 x g for 10 min
at 4˚C to isolate the upper serum layer, which was stored at -80˚C
until further use. The serum levels of OVA-IgE, IL-4, IL-13, IL-17,
IL-1β and IL-10 in mice were measured using ELISAs according to the
manufacturer's protocol.
Histopathological analysis by
hematoxylin-eosin (HE) staining
The nasal tissues of mice were isolated intact under
aseptic conditions, the surface blood was washed, and the tissues
were stored in EP tubes containing 4% paraformaldehyde for fixation
at room temperature for 24 h. After decalcification, dehydration,
transparency, wax immersion and embedding, the sections were cut
into 5-µm-thick slices. Then the sections were stained with HE
solution at room temperature for 3 min after spreading, baking,
dewaxing and rehydrating. After dehydration, clearing and drying,
the slides were sealed and finally observed under a brightfield
microscope.
Reverse transcription (RT)-qPCR
Total RNA was isolated from mouse lung tissue using
TRIzol®, and RNA concentration and purity were
determined. According to the RT Master Mix for qPCR (gDNA digest
plus) kit instructions, the total RNA was reverse transcribed into
cDNA with a final transcription volume of 20 µl. TLR4, MyD88,
IRAK4, NF-κB p65 and GAPDH primers were synthesized by Sangon
Biotech Co., Ltd. (Table II).
qPCR amplification was performed according to the instructions of
SYBR Green qPCR Master Mix (Low ROX) kit under the following
thermocycling conditions: Initial hold at 95˚C for 5 min, followed
by 40 cycles of denaturation at 95˚C for 10 sec and annealing at
60˚C for 30 sec. The relative mRNA expression levels in each group
were calculated using the 2-ΔΔCq method based on the
reference (16).
| Table IIQuantitative PCR primer
sequences. |
Table II
Quantitative PCR primer
sequences.
| Genes | Primer sequences
(5'-3') |
|---|
| TLR4-F |
ATGGCATGGCTTACACCACC |
| TLR4-R |
GAGGCCAATTTTGTCTCCACA |
| MyD88-F |
TCATGTTCTCCATACCCTTGGT |
| MyD88-R |
AAACTGCGAGTGGGGTCAG |
| IRAK4-F |
CATACGCAACCTTAATGTGGGG |
| IRAK4-R |
GGAACTGATTGTATCTGTCGTCG |
| NF-κB-F |
ATGGCAGACGATGATCCCTAC |
| NF-κB-R |
TGTTGACAGTGGTATTTCTGGTG |
| GAPDH-F |
AGGTCGGTGTGAACGGATTTG |
| GAPDH-R |
TGTAGACCATGTAGTTGAGGTCA |
Western blot
Mice lung tissues were collected under aseptic
conditions, stripped of surrounding fat, blood vessels and
connective tissue and stored at -80˚C. When required, tissues were
thawed to room temperature and washed 3 times with PBS.
Subsequently, the protein lysis solution with protease inhibitor
was added. The lysis solution was prepared by mixing RIPA lysis
buffer (strong; cat. no. P0013B; Beyotime Biotechnology) with PMSF
protease inhibitor (cat. no. 36978; Thermo Fisher Scientific, Inc.)
at a volume ratio of 100:1. The tissues were ground and further
lysed for 30 min at 4˚C. The lysates were then centrifuged at 4˚C
at 12,880 x g for 10 min. The supernatant was collected as a
protein stock solution. Protein concentration was measured using a
BCA kit. After adjusting the protein concentration of each group to
the same level, 30 µg protein was loaded per lane for SDS-PAGE
electrophoresis using a 5% loading gel and a 12% resolving gel, at
80 V during the loading gel and at 120 V during the resolving gel.
Subsequently, the proteins were transferred to PVDF membranes,
blocked with TBST (0.1% Tween-20) containing 5% skim milk powder
for 1 h. The membranes were cut according to the location of the
protein of interest and incubated with primary antibodies overnight
at 4˚C on a shaker at the following dilutions: GAPDH (1:2,000; cat.
no. MA5-15738-A488; Thermo Fisher Scientific, Inc.), TLR4 (1:2,000;
cat. no. AF8187; Beyotime Biotechnology), NF-κB p65 (1:1,000; cat.
no. 8242; Cell Signaling Technology, Inc.), IRAK4 (1:1,000; cat.
no. ab119942; Abcam), and MyD88 (1:1,000; cat. no. AF7524; Beyotime
Biotechnology). The following day, membranes were rewarmed at room
temperature for 1 h, washed three times with TBST (10 min per
wash), incubated with goat anti-rabbit IgG antibody (1:5,000; cat.
no. ab6728; Abcam) for 2 h, washed three times again with TBST (10
min per wash), and developed using the SuperSignal™ West
Femto Chemiluminescent Substrate (cat. no. 34098CN; Thermo Fisher
Scientific, Inc.). The results were analyzed using ImageJ (version
1.54g; National Institutes of Health).
Flow cytometry for detection of Tregs
and Th17 cells in splenocytes
After euthanasia of the mice, the spleen was
aseptically stripped, ground in a flat dish containing PBS,
filtered and washed with PBS, Subsequently, 300 µl erythrocyte
lysate was added, the samples were shaken and mixed, and incubated
at 4˚C for 5 min. Next, the splenocytes were then divided into two
halves; one half was used to assess Tregs, and the other half to
assess Th17 cells. Blank control tubes, control isotype tubes, CD4
and CD25 single-stained tubes, and CD4 and IL-17 single-stained
tubes were set up separately. A total of 1 µl of undiluted CD4
antibody and 1 µl of undiluted CD25 antibody was added to each tube
of the Tregs assay; 1 µl of undiluted CD4 antibody was added to
each tube for the Th17 cells assay, and all the samples were
incubated for 30 min in the dark at 4˚C, and then 400 µl fixation
buffer was added and cell were fixed for 1 h at 4˚C. A total of 1
µl of undiluted FOXP3 antibody and 300 µl permeabilization buffer
were added to each tube of the Treg cells assay; 2 µl of undiluted
IL-17 antibody and 300 µl permeabilization buffer were added to
each tube of the Th17 cells assay. All the samples were shaken
thoroughly, incubated overnight at 4˚C, washed twice with PBS and
then evaluated using a BD FACSCanto II flow cytometer (BD
Biosciences). The number of cells per sample was set to
50,000-100,000 for flow cytometry, and cellular debris and dead
cells were excluded. Data analysis was performed using FlowJo
software (version 7.6.1_Min; FlowJo LLC; BD Biosciences).
Statistical analysis
Data were analyzed and plotted using Graph Pad Prism
version 8.0 (Dotmatics), and data are presented as the mean ±
standard deviation. A one-way ANOVA was used for comparison between
multiple groups, followed by Tukey's post hoc test for multiple
comparisons. P<0.05 was considered to indicate a statistically
significant difference.
Results
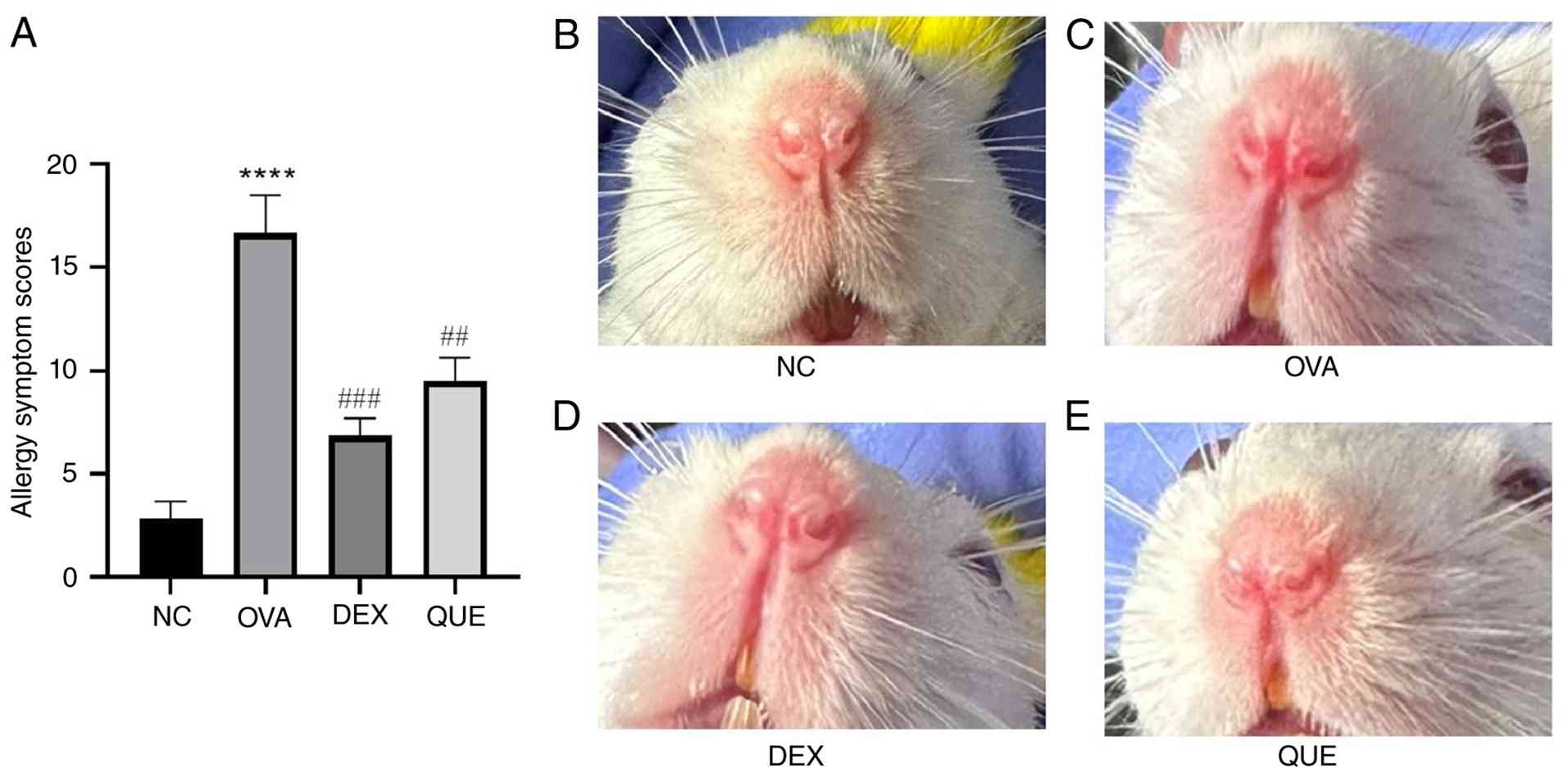
Allergic symptom score in the
OVA-induced mouse model
The mice in the NC group occasionally had scratchy
noses, while the mice in the AR model group had notable symptoms,
including a scratchy nose, sneezing and a runny nose, and all
scores were >10, indicating the successful establishment of the
model. During the treatment of mice after successful modeling, the
nasal drip was continued as before until the day before the mice
were sacrificed (days 28-34); it was noted that the allergic
symptoms persisted in the OVA group during these 7 days, whereas
the scratching, runny nose and rubbing symptoms of the mice in the
treatment groups (DEX and QUE groups) decreased daily in comparison
with the OVA group, as assessed daily according to the scoring
criteria in Table I. Finally, the
results showed that the OVA group allergy symptom scores were
significantly higher than those of the NC group (P<0.0001). The
symptoms were significantly reduced in the DEX (P<0.001) and QUE
(P<0.01) groups compared with those in the OVA group after
treatment, and there was no notable change in the nasal symptoms in
the NC group (Fig. 2A). Under
microscopic examination of HE-stained sections, the nasal mucosa of
the OVA group appeared more congested than that of the NC group,
and after treatment the congestion appeared reduced in the DEX and
QUE groups (representative images shown in Fig. 2B-E, which only partially reflect
the differences observed by the naked eye).
OVA-IgE, IL-4, IL-13, IL-17, IL-1β and
IL-10 expression levels in the serum of mice
OVA-IgE, IL-4, IL-13, IL-17, IL-1β and IL-10 levels
in mouse serum at different time points were assessed using ELISA.
The results showed that OVA-IgE, IL-4, IL-13, IL-17 and IL-1β
levels increased in the OVA group compared with the NC group and
decreased in the DEX and QUE groups compared with the OVA group.
This indicates that OVA increased the serum levels of OVA-IgE,
IL-4, IL-13, IL-17 and IL-1β in mice, and QUE reversed the
OVA-induced changes. For IL-10, its expression was decreased in the
OVA group compared with the NC and increased in the QUE group
compared with the OVA group. This suggests that QUE reversed
OVA-induced changes in IL-10 levels (Fig. 3).
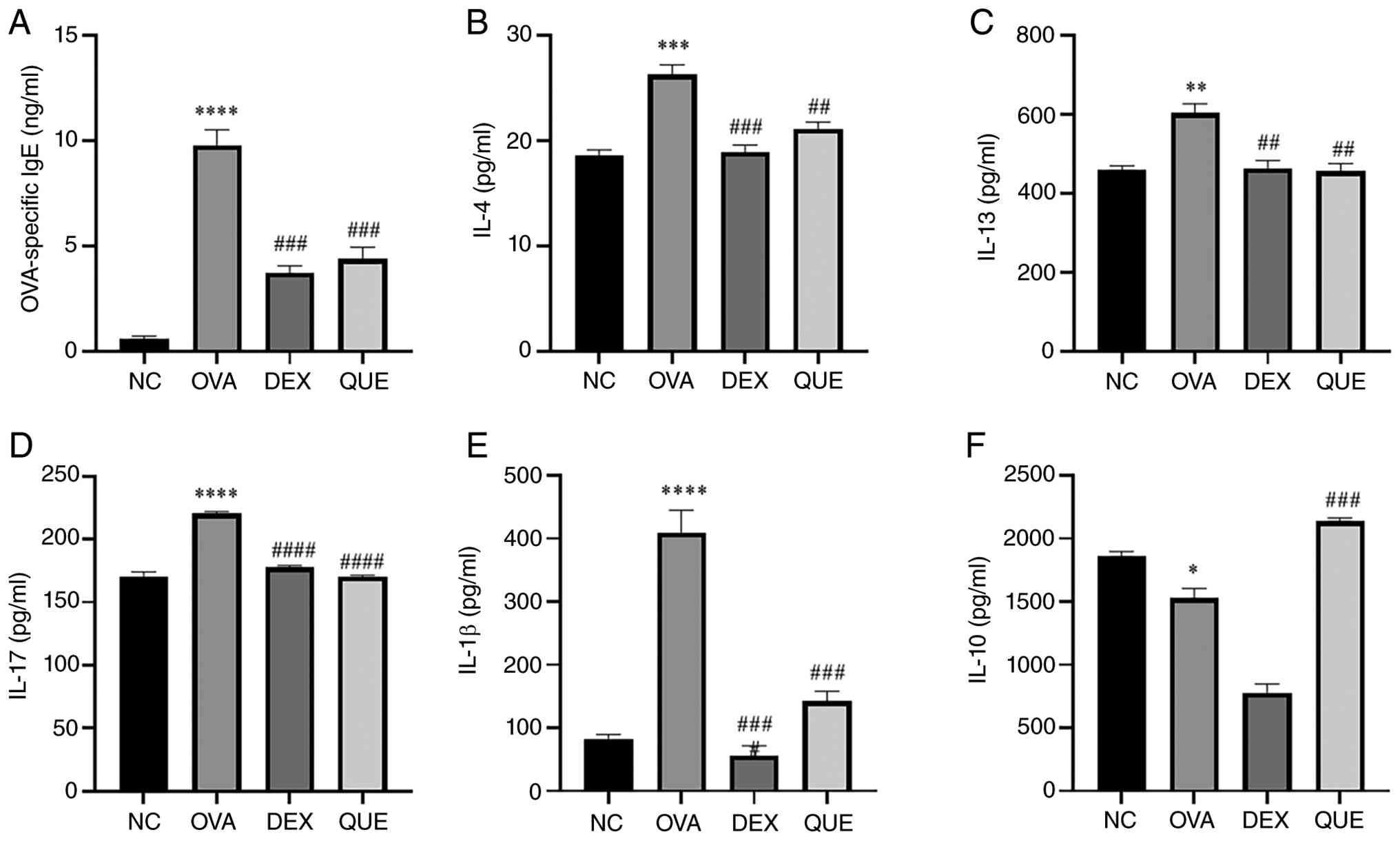 | Figure 3Production of OVA-IgE, IL-4, IL-13,
IL-17, IL-1β and IL-10 in the serum of mice. (A-F) Results of
OVA-IgE, IL-4, IL-13, IL-17, IL-1β and IL-10 relative secretion
levels in NC, OVA, DEX and QUE groups. Data are presented as the
mean ± SEM. *P<0.05, **P<0.01,
***P<0.001, ****P<0.0001 vs. NC;
##P<0.01, ###P<0.001,
####P<0.0001 vs. OVA. NC, normal control; OVA,
ovalbumin; DEX, dexamethasone; QUE, quercetin. |
HE staining of the histopathological
changes of the nasal mucosa in mice
The HE-stained sections of the nasal cavity showed
that the nasal mucosa of the NC group was generally intact, with an
intact epithelial structure; the glandular cell morphology was neat
and orderly, with relatively little inflammatory cell infiltration
and no obvious exudation. In the OVA group, the nasal mucosa was
incomplete, with notable mucosal disruption, shedding, necrosis,
tissue edema, small vessel dilatation, glandular hyperplasia,
disorganized arrangement of glandular cells and obvious
infiltration of inflammatory cells. In the DEX and QUE groups,
mucosal morphology improved, with mucosal damage, shedding and
necrosis all reduced; glandular hyperplasia was reduced, and
glandular cell arrangement tended to be orderly; inflammatory cell
infiltration was reduced; and tissue edema was reduced compared
with that in the OVA group (Fig.
4).
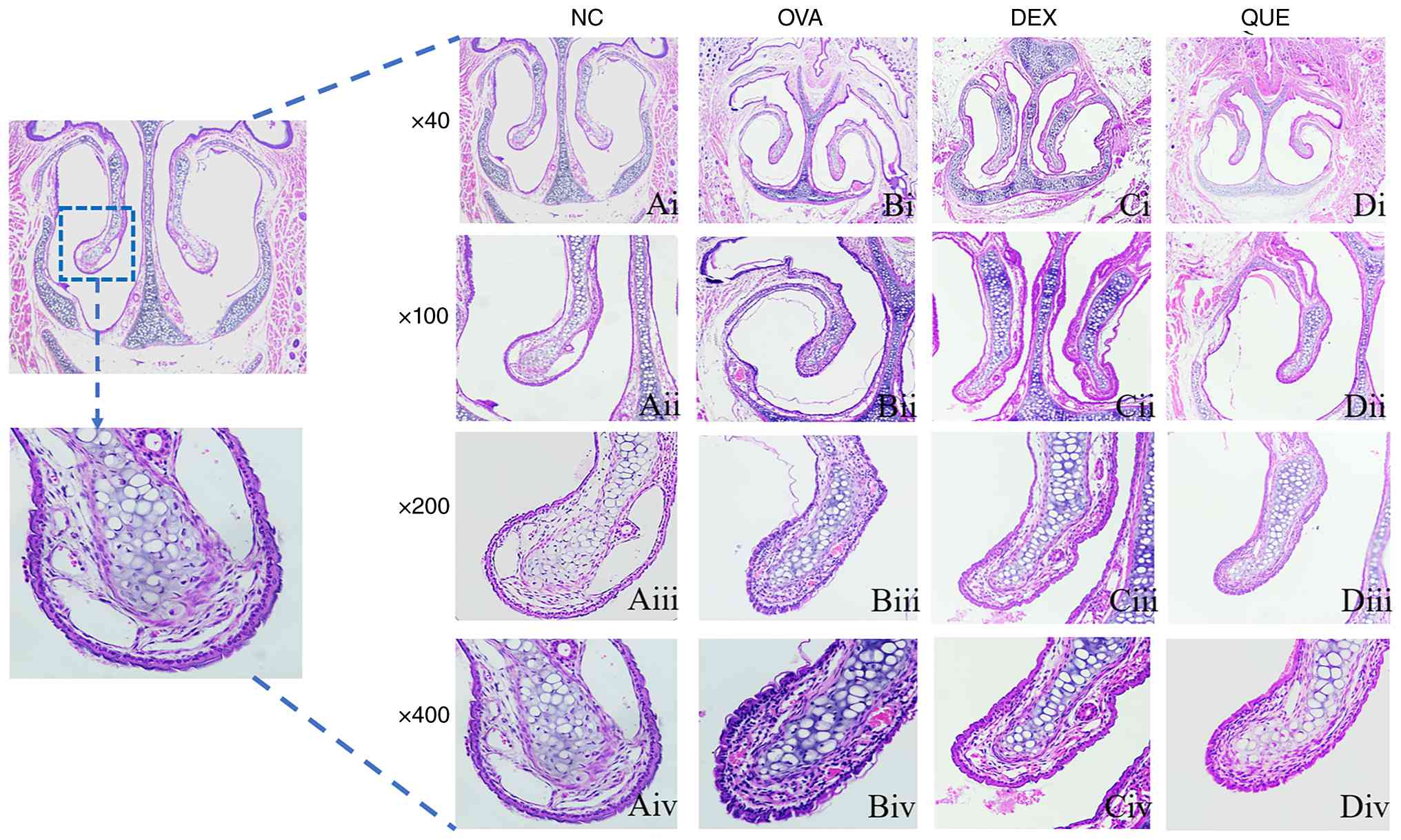 | Figure 4Histopathological
hematoxylin-eosin-stained sections of the mouse nasal mucosa.
Histopathological sections of stained tissues from the (Ai) NC,
(Bi) OVA, (Ci) DEX and (Di) QUE groups; magnification, x40.
Histopathological sections of stained tissues from the (Aii) NC,
(Bii) OVA, (Cii) DEX and (Dii) QUE groups; magnification, x100.
Histopathological sections of stained tissues from the (Aiii) NC,
(Biii) OVA, (Ciii) DEX and (Diii) QUE groups; magnification, x200.
Histopathological sections of stained tissues from the (Aiv) NC,
(Biv) OVA, (Civ) DEX and (Div) QUE groups; magnification, x400. NC,
normal control; OVA, ovalbumin; DEX, dexamethasone; QUE,
quercetin. |
Effect of QUE on the mRNA expression
levels of TLR4, MyD88, IRAK4 and NF-κB
The mRNA expression levels of TLR4, MyD88, IRAK4 and
NF-κB in the lungs of mice were measured using qPCR. The results
showed that the expression of TLR4, MyD88, IRAK4 and NF-κB was
increased in the OVA group compared with the NC group, and
decreased in the DEX and QUE groups compared with the OVA group.
Thus, QUE reversed the OVA-induced effects on the aforementioned
mRNAs (Fig. 5).
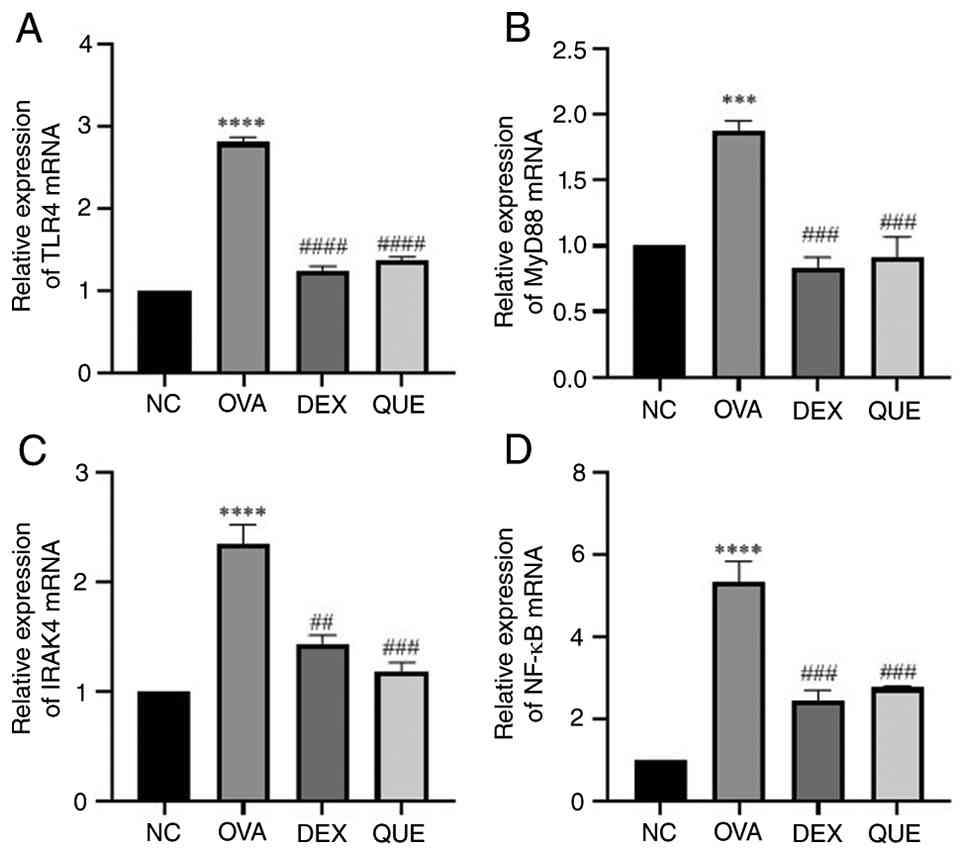 | Figure 5Effect of QUE on mRNA of TLR4, MyD88,
IRAK4 and NF-κB. Results of (A) TLR4, (B) MyD88, (C) IRAK4 and (D)
NF-κB mRNA relative expression levels in NC, OVA, DEX and QUE
groups. Data are presented as the mean ± SEM of three repeats.
***P<0.001, ****P<0.0001 vs. NC;
##P<0.01, ###P<0.001,
####P<0.0001 vs. OVA. NC, normal control; OVA,
ovalbumin; DEX, dexamethasone; QUE, quercetin; TLR4, Toll-like
receptor 4; MyD88, myeloid differentiation primary response 88;
IRAK4, interleukin-1 receptor-associated kinase 4. |
Effect of QUE on the protein
expression levels of TLR4, MyD88, IRAK4 and NF-κB
The relative expression levels of TLR4, MyD88, IRAK4
and NF-κB proteins in the lungs of mice were assessed using western
blotting. The relative expression levels of these proteins were
significantly increased in the OVA group compared with the NC, and
reduced in the DEX and QUE groups compared with the OVA group. This
indicates that QUE inhibited protein expression of members of the
TLR4/MyD88/IRAK4 signaling pathway (Fig. 6).
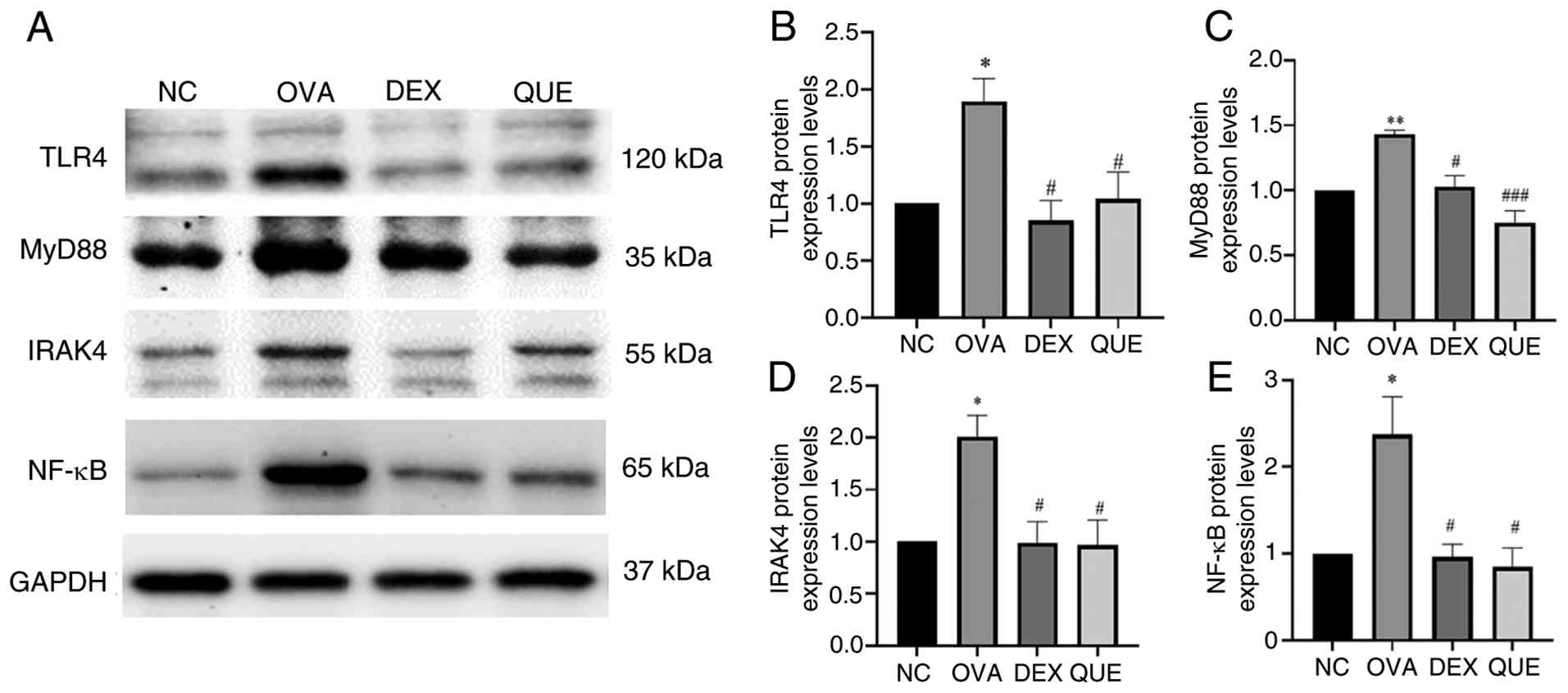 | Figure 6Effects of QUE on TLR4, MyD88, IRAK4
and NF-κB protein levels. (A) A significant increase was observed
in the relative expression of TLR4, MyD88, IRAK4 and NF-κB proteins
in the OVA group compared with the NC group. The relative
expression of these proteins was lower in the DEX and QUE groups
than in the OVA group. Semi-quantification of (B) TLR4, (C) MyD88,
(D) IRAK4 and (E) NF-κB protein expression levels. Data are
presented as the mean ± SEM of three repeats.
*P<0.05, **P<0.01 vs. NC;
#P<0.05, ###P<0.001 vs. OVA. NC, normal
control; OVA, ovalbumin; DEX, dexamethasone; QUE, quercetin; TLR4,
Toll-like receptor 4; MyD88, myeloid differentiation primary
response 88; IRAK4, interleukin-1 receptor-associated kinase 4. |
Effect of QUE on the percentage of
Treg and Th17 cells in splenocytes
The percentages of Tregs
(CD4+/CD25+/FOXP3+) and Th17
(CD4+/IL-17) cells in mouse splenocytes were assessed
using flow cytometry. The results showed that the percentage of
Tregs decreased in the OVA group compared with the NC group, and
increased in the DEX and QUE groups compared with the OVA group
(Fig. 7A-E). This suggests that
OVA reduced the number of Tregs in splenocytes, whereas DEX and QUE
reversed the OVA-induced reduction in Tregs and potentially
promoted Treg differentiation and proliferation. Additionally, the
results showed that the percentage of Th17 cells increased in the
OVA group compared with the NC group, and decreased in the DEX and
QUE groups compared with the OVA group (Fig. 7F-J). This indicates that OVA
increased the percentage of Th17 in splenocytes, whereas DEX and
QUE reversed the OVA-induced increase in the percentage of Th17
cells.
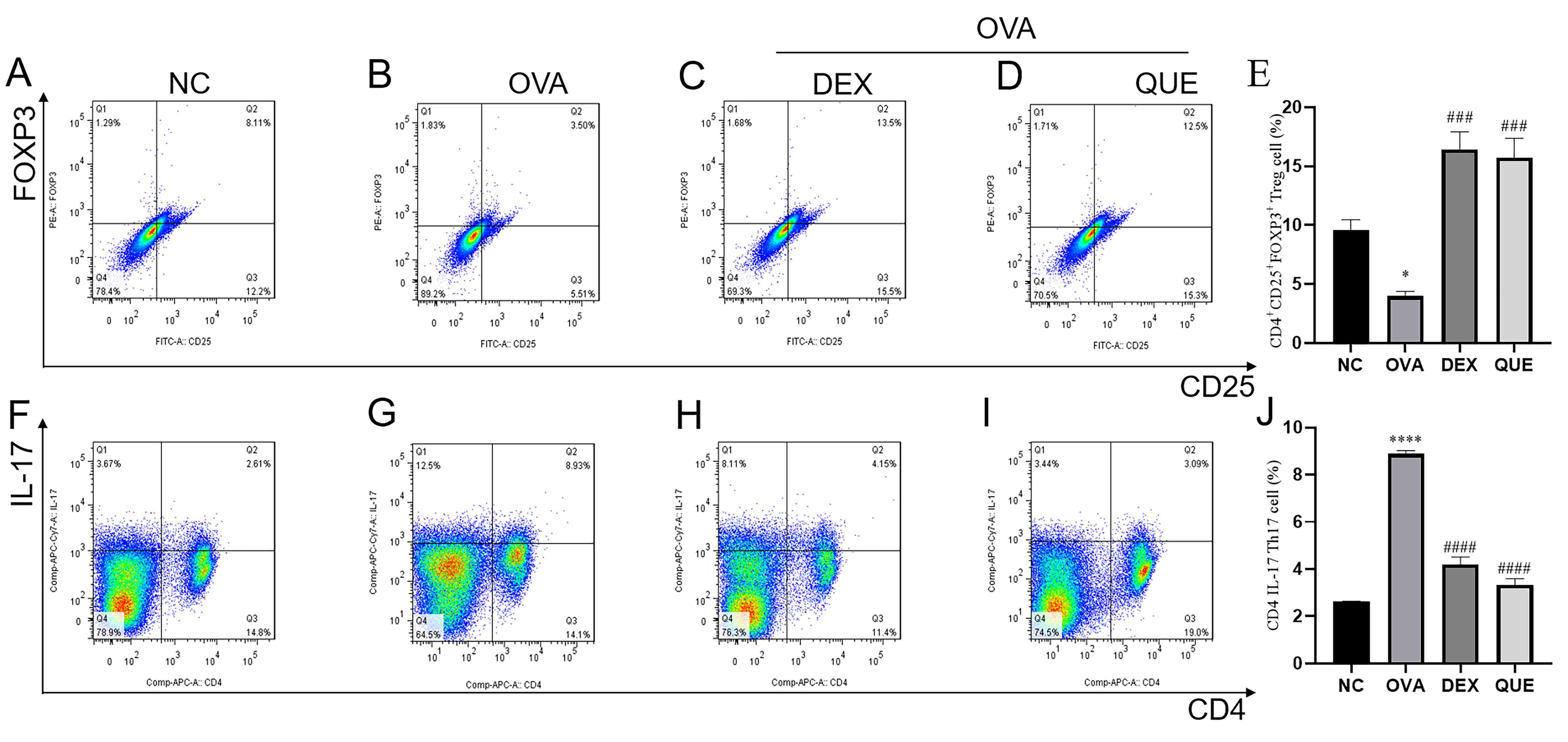 | Figure 7Percentage of Tregs and Th17 cells in
splenocytes of mice. Percentage of Tregs in the splenocytes of mice
in the (A) NC, (B) OVA, (C) DEX and (D) QUE groups. (E) Statistical
analysis of the percentage of Tregs. Percentage of Th17 cells in
splenocytes of mice in (F) NC, (G) OVA, (H) DEX and (I) QUE groups.
(J) Statistical analysis of the percentage of Th17 cells. Data are
presented as the mean ± SEM of three repeats.
*P<0.05, ****P<0.0001 vs. NC;
###P<0.001, ####P<0.0001 vs. OVA. NC,
normal control; OVA, ovalbumin; DEX, dexamethasone; QUE, quercetin;
Th17, T helper 17; Treg, regulatory T cell. |
Discussion
The primary clinical symptoms of AR include an itchy
nose, runny nose and sneezing. Examination can reveal pale edema of
the nasal mucosa, clear water-like secretions in the nasal cavity
and hyperemia of the nasal mucosa in the acute phase. QUE can
alleviate allergic symptoms in the nasal mucosa by reducing the
levels of angiogenic factors and inhibiting oxidative stress
(17,18). In the present study, BALB/c mice
exhibited notable nasal scratching after successful modeling,
suggesting they may have experienced nasal itching. Repeated nose
scratching can lead to whisker loss and thinning of the fur around
the nose in OVA-induced mice, and these manifestations were
observed in the present study post-modeling. At the same time, it
was observed that the allergic symptoms in the QUE and DEX groups
gradually decreased over 7 days of treatment compared with the OVA
group. This indicates that QUE alleviated the OVA-induced allergic
symptoms in mice, and its effects were similar to those of DEX. It
was also observed that the nasal mucosa of the OVA-induced mice
model was more congested than that of the NC group, indicating that
the inflammation was in the acute phase. This also shows that the
modeling was successful. Scratching and rubbing were markedly
reduced in the DEX and QUE groups after treatment, and congestion
of the nasal mucosa was improved compared with that of the OVA
group. This indicates that QUE alleviated nasal itching and nasal
mucosal congestion in mice with AR. From this perspective, the
treatment was effective. For the treatment groups that did not
fully return to normal levels, it was hypothesized that this may be
due to insufficient treatment time or individual differences in
dosage performance, but further research is required. The symptoms
of AR are caused by allergic inflammatory mediators released when
allergens enter the body and stimulate the nasal mucosa.
Re-exposure of sensitized individuals to the same allergen
activates and degranulates mast cells and basophils, which release
histamine, leukotrienes, prostaglandins and other inflammatory
mediators, leading to acute AR symptoms (1,19).
In the present study, a mouse model of AR was induced by intranasal
OVA challenge to evaluate whether treatment with QUE could
alleviate allergic symptoms. Although the levels of histamine,
leukotrienes and prostaglandins were not directly measured in this
study, the reduction in sneezing, scratching and nasal congestion
in the treatment group indirectly suggests that QUE may have
influenced the release of these mediators.
IL-17 is primarily produced by Th17 cells and is
often elevated in AR (20). This
was also demonstrated by the ELISA results, which were consistent
with the flow cytometry findings. IL-1β is a key cytokine that
promotes AR (21). IL-1β activates
the NF-κB pathway, promoting IL-13 production (22). IL-4 and IL-13 contribute to the
production of mucus and IgE, thereby releasing pro-inflammatory
mediators that, in turn, promote the development of allergic
reactions (23). IL-10 serves
multiple roles in immune regulation and inflammation, and its
deficiency is associated with increased IgE in AR (24). The immunomodulatory effect of QUE
was characterized by reductions in IL-4, IL-5, IL-13, IL-1β, TNF-α
and IgE levels (25). The results
of the present study showed that OVA-specific IgE, IL-4 and IL-13
levels were increased and IL-10 levels were decreased in the serum
of the OVA-induced mouse model, and that these indicators were
reversed after QUE and DEX treatment. The increase of OVA-specific
IgE in the OVA group may be associated with the increase of IL-13,
IL-4 and IL-1β and the decrease of IL-10. The mechanism is that
IL-4 and IL-13 promote IgE production, whereas IL-10 suppresses IgE
production, and IL-1β enhances the Th2-type inflammatory response.
Therefore, increased IL-4, IL-13, IL-1β together with decreased
IL-10 result in higher OVA-specific IgE levels. It is necessary to
conduct more in-depth research into their relationship in
subsequent experiments.
QUE, with the anti-allergic function of inhibiting
the production of histamine and pro-inflammatory mediators, can
regulate the stability of the Th1/Th2 balance, reduce
antigen-specific IgE antibodies released by B cells and decrease
IL-4 in serum, thereby reducing allergic airway inflammation and
airway hyperresponsiveness (26).
QUE can also reduce Ca2+ influx induced by allergy and
inhibit chemokine release, similar to the mast cell stabilizer DEX
(27). In allergic conditions, the
activation of innate and adaptive immunity can be attenuated by QUE
(10). In the treatment groups
(DEX and QUE), IL-4 and IL-13 levels were lower than in the OVA
group, suggesting that QUE reduced OVA-induced IL-4 and IL-13
levels and that the reduction of IL-4 and IL-13 may further reduce
IgE. This also suggests that blocking the production of IL-4 and
IL-13 can treat AR and further verifies the anti-inflammatory and
anti-allergic effects of QUE.
The nasal mucosa is rich in small blood vessels.
When allergens trigger an allergic response, inflammatory mediators
cause congestion and edema of the submucosal tissue, smooth muscle
contraction, increased vascular permeability and mucus secretion.
These inflammatory mediators can also increase the number of
eosinophils deep in the nasal mucosa, and excessive accumulation of
eosinophils further releases toxic proteins that can damage the
integrity of respiratory epithelial cells (28,29).
Under the synergistic effect of chemokines, inflammatory cells
infiltrate the nasal mucosal tissue, triggering inflammatory
responses that further damage the nasal mucosal tissue and lead to
clinical symptoms (1). The
infiltration of inflammatory cells in the nasal mucosa tissues of
mice with chronic sinusitis or AR can be alleviated by QUE
(8,30). Ke et al (31) showed that QUE alleviated
OVA-induced thickening of the nasal mucosa and increased
eosinophils in the nasal cavity in an OVA-induced mouse model of
AR. Although the study by Ke et al (31) has demonstrated the anti-allergic
effects of QUE in a similar mouse model, the present study focuses
on the differentiation balance between Th1/Th2 and Treg/Th17 cells.
It systematically reveals the regulatory role of QUE in modulating
the Treg/Th17 balance via the TLR4/MyD88/IRAK4 signaling pathway in
an OVA-induced AR mouse model, thereby expanding the understanding
of the molecular targets of QUE in the treatment of AR. In the
present study, the HE staining results showed that the nasal mucosa
in the OVA group exhibited notable disruption, shedding and
necrosis; dilated submucosal small blood vessels; glandular
hyperplasia; disordered glandular cell arrangement; and
infiltration of inflammatory cells. After treatment, the integrity
of the nasal mucosa was partially restored, tissue edema was
alleviated, the arrangement of glandular cells tended to be more
orderly and the number of infiltrating inflammatory cells was
reduced compared with the OVA group. The number of inflammatory
cells in the nasal mucosa differed markedly. From a morphological
perspective, QUE improved mucosal morphology, reduced the number of
inflammatory cells and restored the integrity of the nasal mucosal
epithelium in the OVA-induced mice model. This further demonstrates
the anti-inflammatory effects of QUE.
TLR4-mediated signaling links innate and adaptive
immunity, and activation of TLR4 drives and promotes inflammation
(32). When TLR4 is activated, it
binds to MyD88, which further activates IRAK4, ultimately leading
to NF-κB activation and the release of inflammatory cytokines and
chemokines, causing inflammation (32,33).
Cheng et al (34) showed
that QUE attenuated LPS-induced inflammatory damage by inducing the
microRNA-21/deleted in malignant brain tumors 1/NF-κB axis.
Therefore, it may serve as a potential compound for the treatment
of AR. Shams and Eissa (35)
showed that QUE ameliorated ethanol-induced gastric ulcers in rats,
involving the nuclear factor erythroid 2-related factor 2/heme
oxygenase-1, high mobility group box 1/TLR4/NF-κB pathways,
suggesting that QUE possesses antioxidant and anti-inflammatory
functions. Gong et al (36)
showed that QUE attenuated LPS-induced cellular damage and
inflammation via the TLR4/NF-κB pathway. The results of the present
study showed that the relative expression of proteins involved in
the TLR4/MyD88/IRAK4 signaling pathway was significantly increased
in the OVA-induced mouse model, suggesting that inflammation in AR
may be associated with this signaling pathway. The expression of
these proteins was reduced in QUE-treated mice, indicating that QUE
can suppress the expression of related proteins in the
TLR4/MyD88/IRAK4 signaling pathway during AR inflammation. The mRNA
expression levels of members of the TLR4/MyD88/IRAK4 signaling
pathway were also assessed, and the changes were consistent with
those obtained using western blotting. This suggests a consistent
effect of QUE on related proteins and mRNAs in the TLR4/MyD88/IRAK4
signaling pathway and further supports the evidence that QUE exerts
a beneficial effect on AR via this pathway.
The immune responses mediated by Th17 cells and
Tregs contribute to the pathogenesis of AR (37). Th17 cells and Tregs represent two
distinct phenotypes of CD4+ T cells with completely
different functions. Th17 cells have pro-inflammatory effects,
while Tregs suppress inflammation; changes in inflammatory cytokine
levels can affect the Th17/Treg balance (38). Th17 cells and Tregs antagonize each
other's function and differentiation, and an imbalance between
Th17/Treg cells can lead to inflammatory responses, which in turn
can induce AR (39,40). QUE has been reported to increase
Tregs and decrease Th17 cells, thereby exerting anti-inflammatory
effects (41), and its regulatory
effect on Tregs may be mediated by FOXP3(10). QUE also inhibits NF-κB activation
and the release of inflammatory mediators, thereby exerting
anti-allergic effects (42). Ke
et al (31) showed that QUE
improved the Treg/Th17 cell imbalance and inhibited the activation
of the NF-κB pathway to treat AR. The results of the present study
showed that in the OVA-induced mouse model, the percentage of Tregs
in splenocytes decreased, while the percentage of Th17 cells
increased, suggesting that OVA can decrease Tregs and increase Th17
cells. The Treg percentage was increased, and Th17 percentage was
decreased in the spleen after QUE treatment, suggesting that QUE
can reduce the number of Th17 cells and reverse the abnormally
downregulated Treg population in splenocytes induced by OVA.
Therefore, it is hypothesized that QUE alleviates AR by restoring
the balance between Treg/Th17 cells, and that AR can be treated by
correcting this imbalance.
In the present study, notable allergic symptoms were
not observed in the NC group, which received only saline solution.
Although QUE is as effective as DEX, long-term use of DEX can lead
to side effects such as immunosuppression and metabolic disorders.
By contrast, QUE is a naturally occurring flavonoid compound with a
good safety profile and low toxicity. This suggests that QUE may
serve as a safer natural alternative for treating AR, though
further clinical validation is needed. Assessing the effect of QUE
in knockout mice may be more convincing for its mechanism of action
in treating AR through the TLR4/MyD88/IRAK4 signaling pathway. This
should be validated through further experiments, which will be
instructive for the development of QUE as a drug to prevent AR.
Furthermore, the present study examined only the effects of QUE on
female mice. The potential deviations in the experimental results
warrant the necessity of future studies that use male mice or
conduct experiments involving both male and female mice
simultaneously.
In conclusion, QUE can effectively alleviate
scratching and sneezing symptoms in the OVA-induced mouse model,
reduce OVA-induced nasal mucosal damage, reduce the expression
levels of inflammatory factors and regulate the imbalance between
Treg/Th17 cells. Its mechanism of action may include reducing the
inflammatory response via the TLR4/MyD88/IRAK4 signaling pathway
and inducing immune tolerance. The present study primarily
evaluated the therapeutic effects of QUE on AR, but its toxicity
and side effects were not explored in additional depth. In
addition, the small sample size used is a limitation. Whether QUE
still has this effect with a larger sample size needs further
verification through additional independent experiments. Therefore,
it is necessary to conduct experiments with larger sample sizes to
observe the therapeutic, toxic and side effects of QUE on AR in
future studies.
Acknowledgements
Not applicable.
Funding
Funding: The present work was supported by grants Natural
Science Foundation of China (grant nos. 81700888, 82260475 and
81860475), Guangdong Basic and Applied Basic Research Foundation
(grant no. 2021A1515010971), Shenzhen Science and Technology
Program for Basic Research (grant no. JCYJ20220531091417040),
Shenzhen Science and Technology Program (grant no.
JCYJ20210324142207019), Science and Technology Development Special
Fund of Shenzhen Longgang District (grant nos. LGKCYLWS2019000864
and LGKCZSYS2019000046), Science and Technology Innovation
Special-Technology Tackling Project of Shenzhen Longgang District
(grant no. LGKCYLWS2022032) and Science and Technology Plan of
Shenzhen Longgang District (grant no. LGKCYLWS2022003).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
CK conceptualized the present study, performed the
experiments, organized the data and prepared the original draft. JL
conducted formal analysis and investigation, and participated in
data analysis and interpretation. SQ participated in
conceptualization and design, supervised the research process,
validated the results and provided funding. XW conceptualized the
study, participated in data interpretation, and critically reviewed
the manuscript for important intellectual content. All authors
reviewed and edited the manuscript and approved the final version
of the manuscript. SQ and JL confirm the authenticity of all the
raw data.
Ethics approval and consent to
participate
The present study was conducted in accordance with
the ARRIVE guidelines for animal research and complied with its
regulations. All experimental protocols used in the study complied
with animal ethics standards and were approved by the Experimental
Animal Ethics Committee of Shenzhen Institute of
Otorhinolaryngology (approval no. 2022-0426). All experimental
methods and steps complied with relevant institutional, regional
and national guidelines and regulations and the suffering to the
animals was minimized as much as possible.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bousquet J, Anto JM, Bachert C, Baiardini
I, Bosnic-Anticevich S, Walter Canonica G, Melén E, Palomares O,
Scadding GK, Togias A and Toppila-Salmi S: Allergic rhinitis. Nat
Rev Dis Primers. 6(95)2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Greiner AN, Hellings PW, Rotiroti G and
Scadding GK: Allergic rhinitis. Lancet. 378:2112–2122.
2011.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Kirtland ME, Tsitoura DC, Durham SR and
Shamji MH: Toll-like receptor agonists as adjuvants for allergen
immunotherapy. Front Immunol. 11(599083)2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Murrison LB, Brandt EB, Myers JB and
Hershey GKK: Environmental exposures and mechanisms in allergy and
asthma development. J Clin Invest. 129:1504–1515. 2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Waage J, Standl M, Curtin JA, Jessen LE,
Thorsen J, Tian C and Schoettler N: 23andMe Research Team; AAGC
collaborators. Flores C, et al: Genome-wide association and HLA
fine-mapping studies identify risk loci and genetic pathways
underlying allergic rhinitis. Nat Genet. 50:1072–1080.
2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Shen P, Lin W, Deng X, Ba X, Han L, Chen
Z, Qin K, Huang Y and Tu S: Potential implications of quercetin in
autoimmune diseases. Front Immunol. 12(689044)2021.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Manzoor MF, Hussain A, Sameen A, Sahar A,
Khan S, Siddique R, Aadil RM and Xu B: Novel extraction, rapid
assessment and bioavailability improvement of quercetin: A review.
Ultrason Sonochem. 78(105686)2021.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Deng Y, Shen L, Zhu H, Zhou Y and Hu X:
Network pharmacology analysis of the Huangqi-Gancao herb pair
reveals quercetin as a therapeutics for allergic rhinitis via the
RELA-regulated IFNG/IRF1 axis response. Naunyn Schmiedebergs Arch
Pharmacol. 398:1597–1612. 2025.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Cheng J, Zhang M, Zheng Y, Wang J and Wang
Q: Integrative analysis of network pharmacology and proteomics to
identify key targets of Tuomin-Zhiti-Decoction for allergic
rhinitis. J Ethnopharmacol. 296(115448)2022.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Lv Z, Pan Z, Huang Y, Yang H and Li X:
Quercetin exhibits multi-target anti-allergic effects in animal
models: A systematic review and meta-analysis of preclinical
studies. Front Pharmacol. 16(1673712)2025.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Yang S, Zhang J, Wang S, Zhao X and Shi J:
SOCS2 overexpression alleviates diabetic nephropathy in rats by
inhibiting the TLR4/NF-κB pathway. Oncotarget. 8:91185–91198.
2017.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Zhang Y and Zeng Y: Curcumin reduces
inflammation in knee osteoarthritis rats through blocking TLR4
/MyD88/NF-κB signal pathway. Drug Dev Res. 80:353–359.
2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Yamatomo T, Okano M, Ono T, Nakayama E,
Yoshino T, Satoskar AR, Harn DA Jr and Nishizaki K: Sex-related
differences in the initiation of allergic rhinitis in mice.
Allergy. 56:525–531. 2001.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Fang S, Li X, Wei X, Zhang Y, Ma Z, Wei Y
and Wang W: Beneficial effects of hydrogen gas inhalation on a
murine model of allergic rhinitis. Exp Ther Med. 16:5178–5184.
2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Ryu G, Bae JS, Kim JH, Kim EH, Chung YJ
and Mo JH: Sneezing and rubbing counts in allergic rhinitis mouse
models are a reliable indicator of type 2 immune response. Clin Exp
Otorhinolaryngol. 13:308–311. 2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Okumo T, Furuta A, Kimura T, Yusa K, Asano
K and Sunagawa M: Inhibition of angiogenic factor productions by
quercetin in vitro and in vivo. Medicines (Basel).
8(22)2021.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Edo Y, Otaki A and Asano K: Quercetin
enhances the thioredoxin production of nasal epithelial cells in
vitro and in vivo. Medicines (Basel). 5(124)2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Cheng L, Chen J, Fu Q, He S, Li H, Liu Z,
Tan G, Tao Z, Wang D, Wen W, et al: Chinese society of allergy
guidelines for diagnosis and treatment of allergic rhinitis.
Allergy Asthma Immunol Res. 10:300–353. 2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Hofmann MA, Fluhr JW, Ruwwe-Glösenkamp C,
Stevanovic K, Bergmann KC and Zuberbier T: Role of IL-17 in atopy-A
systematic review. Clin Transl Allergy. 11(e12047)2021.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Wang HR, Wei SZ, Song XY, Wang Y, Zhang
WB, Ren C, Mou YK and Song XC: IL-1β and allergy: Focusing on Its
role in allergic rhinitis. Mediators Inflamm.
2023(1265449)2023.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Kato A: Group 2 innate lymphoid cells in
airway diseases. Chest. 156:141–149. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Busse WW, Kraft M, Rabe KF, Deniz Y, Rowe
PJ, Ruddy M and Castro M: Understanding the key issues in the
treatment of uncontrolled persistent asthma with type 2
inflammation. Eur Respir J. 58(2003393)2021.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Shao JB, Luo XQ, Wu YJ, Li MG, Hong JY, Mo
LH, Liu ZG, Li HB, Liu DB and Yang PC: Histone deacetylase 11
inhibits interleukin 10 in B cells of subjects with allergic
rhinitis. Int Forum Allergy Rhinol. 8:1274–1283. 2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Marefati N, Ghorani V, Shakeri F,
Boskabady M, Kianian F, Rezaee R and Boskabady MH: A review of
anti-inflammatory, antioxidant, and immunomodulatory effects of
Allium cepa and its main constituents. Pharm Biol. 59:287–302.
2021.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Jafarinia M, Sadat Hosseini M, Kasiri N,
Fazel N, Fathi F, Ganjalikhani Hakemi M and Eskandari N: Quercetin
with the potential effect on allergic diseases. Allergy Asthma Clin
Immunol. 16(36)2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Ding Y, Che D, Li C, Cao J, Wang J, Ma P,
Zhao T, An H and Zhang T: Quercetin inhibits Mrgprx2-induced
pseudo-allergic reaction via PLCγ-IP3R related Ca2+
fluctuations. Int Immunopharmacol. 66:185–197. 2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Liu Y, Sha J, Meng C and Zhu D: Mechanism
of lower airway hyperresponsiveness induced by allergic rhinitis. J
Immunol Res. 2022(4351345)2022.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Okubo K, Kurono Y, Ichimura K, Enomoto T,
Okamoto Y, Kawauchi H, Suzaki H, Fujieda S and Masuyama K: Japanese
Society of Allergology. Japanese guidelines for allergic rhinitis
2020. Allergol Int. 69:331–345. 2020.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Meng L, Qu X, Tao P, Dong J and Guo R:
Quercetin alleviates the progression of chronic rhinosinusitis
without nasal polyps by inhibiting nasal mucosal inflammation and
epithelial apoptosis. Mol Biotechnol. 67:3532–3543. 2025.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Ke X, Chen Z, Wang X, Kang H and Hong S:
Quercetin improves the imbalance of Th1/Th2 cells and Treg/Th17
cells to attenuate allergic rhinitis. Autoimmunity.
56(2189133)2023.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Zamyatina A and Heine H:
Lipopolysaccharide recognition in the crossroads of TLR4 and
caspase-4/11 mediated inflammatory pathways. Front Immunol.
11(585146)2020.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Bruno K, Woller SA, Miller YI, Yaksh TL,
Wallace M, Beaton G and Chakravarthy K: Targeting toll-like
receptor-4 (TLR4)-an emerging therapeutic target for persistent
pain states. Pain. 159:1908–1915. 2018.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Cheng J, Luo XQ and Chen FS: Quercetin
attenuates lipopolysaccharide-mediated inflammatory injury in human
nasal epithelial cells via regulating miR-21/DMBT1/NF-κB axis.
Immunopharmacol Immunotoxicol. 44:7–16. 2022.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Shams SGE and Eissa RG: Amelioration of
ethanol-induced gastric ulcer in rats by quercetin: Implication of
Nrf2/HO1 and HMGB1/TLR4/NF-κB pathways. Heliyon.
8(e11159)2022.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Gong X, Huang Y, Ma Q, Jiang M, Zhan K and
Zhao G: Quercetin alleviates lipopolysaccharide-induced cell damage
and inflammation via regulation of the TLR4/NF-κB pathway in bovine
intestinal epithelial cells. Curr Issues Mol Biol. 44:5234–5246.
2022.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Mu D, Zhou L, Shi L, Liu T, Guo Y, Chen H,
Luo H, Ma J, Zhang H, Xiong P and Tian L: Quercetin-crosslinked
chitosan nanoparticles: A potential treatment for allergic
rhinitis. Sci Rep. 14(4021)2024.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Zhang S, Gang X, Yang S, Cui M, Sun L, Li
Z and Wang G: The alterations in and the role of the Th17/Treg
balance in metabolic diseases. Front Immunol.
12(678355)2021.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Shan J, Jin H and Xu Y: T cell metabolism:
A new perspective on Th17/Treg cell imbalance in systemic lupus
erythematosus. Front Immunol. 11(1027)2020.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Chen Z, Ke X, Wang X, Kang H and Hong S:
LncRNA JPX contributes to Treg/Th17 imbalance in allergic rhinitis
via targeting the miR-378g/CCL5 axis. Immunopharmacol
Immunotoxicol. 44:519–524. 2022.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Zarenezhad E, Abdulabbas HT, Kareem AS,
Kouhpayeh SA, Barbaresi S, Najafipour S, Mazarzaei A, Sotoudeh M
and Ghasemian A: Protective role of flavonoids quercetin and
silymarin in the viral-associated inflammatory bowel disease: An
updated review. Arch Microbiol. 205(252)2023.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Mlcek J, Jurikova T, Skrovankova S and
Sochor J: Quercetin and its anti-allergic immune response.
Molecules. 21(632)2016.PubMed/NCBI View Article : Google Scholar
|