Introduction
Aging, as a natural law associated with the
degenerative functional and organic process that occurs in the body
with age, can lead to and aggravate degenerative diseases (1-3).
Studies have shown that aging markedly increases the incidence of
cardiovascular diseases (CVDs) such as myocardial ischemia,
atherosclerosis and cardiac fibrosis (4-7).
Oxidative stress is a key factor in cardiac aging, while the
inflammatory response is an important defense mechanism of the
heart; both can affect the structure of myocardial cells and
cardiac function (8). Cellular
senescence is a core mechanism of organismal aging. It has received
in-depth attention in recent years due to its pathological
importance as a driving factor of chronic diseases (9). Endothelial cell (ECs) senescence
serves a key role in the process of vascular aging. ECs dysfunction
is directly involved in processes such as the formation of
atherosclerotic plaques, the decrease in vascular elasticity and
the pathological remodeling of hypertension (10-12).
These previous studies emphasized that cellular senescence is not
only a sign of organismal aging but also a driving factor for
numerous chronic diseases (13-15).
Senescent ECs exhibit the characteristics of mitochondrial dynamic
imbalance, which directly affects the structure and function of
blood vessels and thereby promotes the development of CVDs
(16). Therefore, seeking
effective strategies to intervene in ECs senescence is of great
importance for delaying blood vessel aging and the progression of
associated diseases.
Probiotic cultures that have undergone inactivation
treatment are collectively referred to as postbiotics (17). In a broad sense, the concept of
postbiotics encompasses both bacteria and their metabolic products,
which possess biological activities similar to or better than those
of live bacteria, such as antibacterial, antioxidant,
anti-inflammatory and immunomodulatory properties (18).
Bifidobacterium is widely present in the
intestines of humans and animals. It exerts regulatory effects on
the brain-gut axis and can be used to treat irritable bowel
syndrome, alleviate depression and improve cognitive impairment.
Its probiotic properties can be used to relieve immune-associated
diseases such as allergies and colitis (19). However, research on
Bifidobacterium and the regulation of cardiovascular health
by its metabolites is still relatively scarce. Previously, a strain
of Bifidobacterium adolescentis Life Age 20 was isolated
from the feces of long-lived cats (20). A mouse experiment further
demonstrated that this strain could markedly extend the lifespan of
mice with accelerated aging (20).
Based on this result, an additional strain of Bifidobacterium
adolescentis, Life Age 20k9 (LA20k9), was isolated preserved in
the China Center for Type Culture Collection (preservation no.
CCTCC-M-025638), from the feces of long-lived dogs. The aim of the
present study was to investigate the anti-aging effects of the
fermentation products of Bifidobacterium adolescentis in
brown algae extract on canine blood vessels and explore its
potential mechanism.
In the present study, an in vitro canine
vascular endothelial cell (CVEC) model was employed to investigate
the anti-vascular aging effects of LA20k9. The effects of LA20k9 on
vascular senescence, endothelial function, inflammation and the
molecular markers of aging were assessed to establish an
association between treatment response and cellular aging status.
Transcriptomics and metabolomics are widely employed
high-throughput omics approaches in biological research (21,22).
Transcriptomic profiling enables the systematic quantification of
gene expression dynamics across experimental conditions, whereas
metabolomic profiling captures condition-dependent alterations in
small-molecule metabolite abundances. Integrating this
complementary molecular information affords a more holistic view of
biological system responses, thereby enhancing mechanistic insight
in biomedical and translational research. Accordingly,
transcriptomic and metabolomic analyses were jointly applied to
comprehensively elucidate the molecular mechanisms underlying the
observed biological activities. The present study aimed to assess
the anti-aging effects of metabolites derived from brown algae
extract fermented by LA20k9 based on a CVEC model and elucidate its
underlying cellular mechanisms. The present findings could provide
both scientific and practical implications for the potential
therapeutic applications of LA20k9, supporting the development of
novel biological interventions targeting post-aging processes.
Materials and methods
Reagents and materials
A Bifidobacterium adolescentis LA20k9 strain
was isolated from the feces of an elderly dog (accession no:
CCTCC-M-2025638). For LA20k9 fermentation and freeze-drying, the
seed culture of Bifidobacterium adolescentis LA20k9,
preserved in 15% glycerol (Oxoid; Thermo Fisher Scientific, Inc.),
was inoculated into De Man-Rogosa-Sharpe medium (Oxoid; Thermo
Fisher Scientific, Inc.) containing 2% (v/v) brown algae extract
(derived from Laminaria japonica; Shanxi Benhe Biotechnology
Engineering Co., Ltd.) and anaerobic culture was carried out at
38˚C for 48 h.
Furthermore, 15% glycerol without Bifidobacterium
adolescentis LA20k9 was fermented under the same conditions,
with blank substrate fermentation metabolite (BMFM) excluding
LA20-k9. The supernatant of the fermentation broth was collected
after room temperature centrifugation for 10 min at 2,000 x g and
freeze-dried. CVECs [Global Pet Cell Resource Center (PETCC)-199
cell line; cat. no. PETCC#199] were obtained from the PETCC
(Tianjin, China). Doxorubicin (DOX) was purchased from Shanghai
Aladdin Biochemical Technology Co., Ltd. Hanks' balanced salt
solution (HBSS), the senescence-associated β-galactosidase
(SA-β-gal) staining kit and SA-β-gal activity assay kit were
purchased from Beijing Solarbio Science & Technology Co., Ltd.
Trypsin, FBS, TRIzol® and M-3 medium were obtained from
Gibco (Thermo Fisher Scientific, Inc.). Canine ELISA kits for MMP-1
(cat. no. ml031816), MMP-3 (cat. no. ml024650), VEGF (cat. no.
ml031966), basic fibroblast growth factor (bFGF; cat. no.
ml031890), granulocyte-macrophage colony stimulating factor
(GM-CSF; cat. no. ml002650) and inflammatory factors IL-1β (cat.
no. ml024767), TNF-α (cat. no. ml031931), IL-6 (cat. no. ml024780),
IL-8 (cat. no. ml024786) and monocyte chemotactic protein-1 (MCP-1;
cat. no. ml002710) were obtained from Shanghai Enzyme-linked
Biotechnology Co., Ltd. Oxidative stress-associated kits for
superoxide dismutase (SOD; cat. no. S0101S), malondialdehyde (MDA;
cat. no. S0131S) and glutathione (GSH; cat. no. S0053).
Analysis of the composition of LA20k9
and BMFM. Detection of monosaccharide and polysaccharide
contents
Disaccharide and monosaccharide contents of LA20k9
and BMFM were determined using precolumn derivatization with
1-phenyl-3-methyl-5-pyrazolone (PMP) (cat. no. A501894-0250; Sangon
Biotech Co., Ltd.) and high-performance liquid chromatography
(HPLC). LA20k9 and BMFM were hydrolyzed in 10 ml 7.2% sulfuric acid
(cat. no. S962529; Shanghai Macklin Biochemical Co., Ltd.) under
nitrogen at 110˚C for 2 h. After hydrolysis, 0.5 ml the sample was
neutralized and diluted with water to 1 ml. Monosaccharide
standards [glucose (cat. no. D823520) and fucose (cat. no. L809666)
were obtained from Shanghai Macklin Biochemical Co., Ltd.] or
hydrolyzed LA20k9 and BMFM were derivatized with 0.2 ml 0.3 M
sodium hydroxide (cat. no. R885161; Shanghai Macklin Biochemical
Co., Ltd.) and 0.4 ml PMP solution. The mixture was incubated at
70˚C for 60 min under nitrogen. Afterward, 0.2 ml 0.3 M
hydrochloric acid (cat. no. C0680110271; Nanjing Chemical Reagent
Co., Ltd.) was added and the volume was adjusted to 2 ml with
water. Chloroform (1.5 ml; cat. no. C1481519267; Nanjing Chemical
Reagent Co., Ltd.) was added, shaken and the organic layer
discarded. The aqueous phase was filtered through a 0.45 µm aqueous
membrane and analyzed on an Agilent 1200 HPLC system (Agilent
Technologies, Inc.) with a C18 (250.0x4.6 mm; 5 µm) column and UV
detection. The method of monosaccharide concentration detection
followed the same procedure as described for polysaccharide content
analysis, excluding the hydrolysis step.
Protein content. LA20k9 and BMFM protein
content was determined using the Pierce™ BCA protein
assay kit (cat. no. A65453; Thermo Fisher Scientific, Inc.). A 20
mg sample was accurately weighed and dissolved in 1 ml ultrapure
water, followed by a 200-fold dilution to prepare the sample
solution. Then, 20 µl solution was mixed with 200 µl BCA working
reagent. The reaction mixture was incubated at 37˚C for 30 min and
the absorbance at 562 nm was measured at the end of the
incubation.
Detection of amino acid content. LA20k9 and
BMFM amino acid content was determined by HPLC coupled with
pre-column derivatization using o-phthaldialdehyde (OPA) (cat. no.
26025; Thermo Fisher Scientific, Inc.). LA20k9 and BMFM were
hydrolyzed in 10 ml 6 M hydrochloric acid under nitrogen at 110˚C
for 24 h to prevent oxidation. After hydrolysis, 100 µl sample
solution or amino acid standard [aspartic acid (cat. no. L708685),
threonine (cat. no. H818497), serine (cat. no. D794545), glutamic
acid (cat. no. D665473), glycine (cat. no. G800880), alanine (cat.
no. D688836), cystine (cat. no. D804429), valine (cat. no.
V828207), methionine (cat. no. L675564), isoleucine (cat. no.
H811900), leucine (cat. no. D688815), tyrosine (cat. no. D818619),
phenylalanine (cat. no. D793640), histidine (cat. no. D810956),
lysine (cat. no. D856989), arginine (cat. no. H677986) and proline
(cat. no. D794397) obtained from Shanghai Macklin Biochemical Co.,
Ltd.] was mixed with 200 µl OPA reagent. The mixture was vortexed
vigorously for 30 sec and allowed to react at room temperature in
the dark for 15-20 min. After reaction, the mixture was filtered
through a 0.22 µm aqueous membrane to remove particulates prior to
HPLC analysis. HPLC separation was performed on an Agilent 1200
HPLC system (Agilent Technologies, Inc.) equipped with a C18
reversed-phase column. The mobile phase consisted of 0.1 M PBS (pH
8.0; solvent A) and a mixture of methanol (cat. no. C0690110124;
Nanjing Chemical Reagent Co., Ltd.) and acetonitrile [cat. no.
C0560116005; Nanjing Chemical Reagent Co., Ltd.; 1:1 (v/v) solvent
B]. A linear gradient elution was employed as follows: 0-10 min,
10-30% B; 10-25 min, 30-50% B; 25-30 min, 50-10% B; and 30-35 min,
10% B for equilibration. The flow rate was set at 1.0 ml/min and
the column temperature was maintained at 30˚C. Amino acids were
emission wavelengths set at 340 and 450 nm, respectively.
Detection of organic acid content. Organic
acid content was determined by HPLC. The analysis was performed on
a reversed-phase C18 column with 0.01 mol/l potassium dihydrogen
phosphate solution (pH 2.5-3.0; cat. no. P837011; Shanghai Macklin
Biochemical Co., Ltd.) as the mobile phase at a flow rate of 1.0
ml/min. The column temperature was maintained at 30˚C and the
detection wavelength was set at 210 nm. The injection volume was 20
µl. Samples or organic acid standard [tartaric acid (cat. no.
D666102), malic acid (cat. no. D689340), lactic acid (cat. no.
D742481), pyruvic acid (cat. no. P796878), oxalic acid (cat. no.
S818087), acetic acid (cat. no. A801299), citric acid (cat. no.
C805019), succinic acid (cat. no. S874398), propionic acid (cat.
no. P757623) and benzenolactic acid (cat. no. L795422) obtained
from Shanghai Macklin Biochemical Co., Ltd.] were dissolved in
water, vortexed to mix, prepared as standard working solutions at
room temperature and filtered through a 0.22 µm aqueous membrane
before injection. Quantification was carried out using the external
standard method (23,24). Calibration curves were constructed
by plotting peak areas against corresponding standard
concentrations. The contents of individual organic acids in samples
were calculated based on the calibration equations.
Cell culture
CVECs were authenticated by the supplier and tested
negative for Mycoplasma contamination. The cells were
cultured in modified M3-DMEM medium (PETCC) supplemented with 12%
FBS. Cultures were maintained in an atmosphere humidified with 5%
CO2 until 90% confluence before use.
Cell viability assay
Cell viability was assessed using a Cell Counting
Kit-8 (CCK-8) assay (Beyotime Biotechnology). CVECs were stained
with trypan blue at room temperature for 5 min, counted and
adjusted to 1x104 cells/ml. The cells were seeded into
96-well plates and incubated for 24 h at 37˚C with numerous
concentrations of LA20k9 (0, 20, 40, 80, 160, 320, 640 and 1,280
µg/ml), BMFM (0, 20, 40, 80, 160, 320, 640 and 1,280 µg/ml) and DOX
(0, 0.2, 0.3, 0.4, 0.5, 0.6, 0.7, 0.8, 0.9 and 1.0 µmol/l). After
removing the drug solutions, 110 µl HBSS containing 10% CCK-8 was
added, mixed for 30 sec and incubated for 2 h. Absorbance was then
measured at 450 nm. HBSS without drugs served as the negative
control (NC). DOX at 60-70% viability was used to induce
senescence.
The following groups were established to assess the
capacity of LA20k9 and BMFM to restore the viability of DOX-induced
vascular ECs: HBSS with only DOX (senescence model group); DOX with
varying concentrations of LA20k9 (LA20k9 treatment groups); DOX
with varying concentrations of BMFM (BMFM treatment groups); and
HBSS alone (NC group). The procedures were the same as those used
to determine the drug concentrations.
SA-β-gal activity assay
CVECs were seeded in 6-well plates at a density of
1x106 cells/well and cultured at 37˚C and 5%
CO2 for 24 h until the cells adhered. Drug treatment
groups were the same as those used for the cell viability assay.
After 24 h, quantitative and qualitative analyses were conducted
using the SA-β-gal activity detection kit and the SA-β-gal staining
kit, respectively, following the manufacturer's protocols.
Evaluation of cellular senescence
markers
CVECs were seeded in 6-well plates at a density of
1x106 cells/well and cultured at 37˚C and 5%
CO2 for 24 h until the cells adhered. Drug treatment
groups were the same as those used in the cell viability assay.
After 24 h, the levels of MMP-1, MMP-3, VEGF, bFGF, IL-1β, TNF-α,
IL-6, IL-8, GM-CSF and MCP-1 were measured using ELISA kits. Canine
ELISA kits for MMP-1 (cat. no. ml031816), MMP-3 (cat. no.
ml024650), VEGF (cat. no. ml031966), basic fibroblast growth factor
(bFGF; cat. no. ml031890), granulocyte-macrophage colony
stimulating factor (GM-CSF; cat. no. ml002650) and inflammatory
factors IL-1β (cat. no. ml024767), TNF-α (cat. no. ml031931), IL-6
(cat. no. ml024780), IL-8 (cat. no. ml024786) and monocyte
chemotactic protein-1 (MCP-1; cat. no. ml002710) were obtained from
Shanghai Enzyme-linked Biotechnology Co., Ltd. Specifically, 50 µl
supernatant from each treatment group was added to wells pre-coated
with primary antibodies specific to either canine protein.
HRP-conjugated secondary antibodies were then added and the plates
were incubated at 37˚C for 60 min. After incubation, the wells were
washed five times with PBS with Tween 20, followed by the addition
of 100 µl 3,3',5,5'-tetramethylbenzidine substrate for color
development. Absorbance was measured at 450 nm.
Evaluation of oxidative stress
markers
CVECs were seeded in 6-well plates at a density of
1x106 cells/well and cultured at 37˚C and 5%
CO2 for 24 h until the cells adhered. The treatment
groups were the same as used for the cell viability assay. After 24
h of action, the SOD content was detected using a total SOD
activity detection kit, following the manufacturer's protocols and
absorbance was measured at 450 nm. MDA content was used to assess
lipid oxidation levels. MDA was measured in cell lysates using a
colorimetric lipid oxidation detection kit, following the
manufacturer's protocols. Absorbance was measured at 532 nm. GSH
content was measured using a GSH detection kit, following the
manufacturer's protocols. GSH levels were measured by mixing 20 µl
sample with 5,5'-dithiobis (2-nitrobenzoic acid) solution, allowing
the reaction to proceed for 5 min and reading the absorbance at 405
nm. An enhanced NAD+ detection kit was used to measure
the NAD+ content through sequential incubation with
ethanol dehydrogenase, followed by the addition of a colorimetric
reagent in strict accordance with the manufacturer's instructions.
Protein concentrations were quantified using a BCA kit (Thermo
Fisher Scientific, Inc.).
RNA sequencing
Cells from different treatment groups were lysed
using TRIzol reagent. Phase separation was achieved by adding
chloroform, followed by RNA precipitation with isopropanol and
washing with ethanol. The isolated RNA was dissolved in RNase-free
water (cat. no. R0022; Beyotime Biotechnology). RNA concentration
and integrity were verified using an Agilent 5400 Bioanalyzer
(Agilent Technologies, Inc.). mRNA was enriched using Oligo(dT)
beads and fragmented in a buffer with divalent cations
[NEBNext® Poly(A) mRNA Magnetic Isolation Module; cat.
no. E7490; New England Biolabs, Inc.]. First-strand cDNA was
synthesized using Moloney Murine Leukemia Virus Reverse
Transcriptase (New England Biolabs, Inc.), followed by
second-strand synthesis with DNA polymerase I and RNase H
(NEBNext® Ultra™ II RNA Library Prep; cat.
no. E7775L; New England Biolabs, Inc.). The double-stranded cDNA
underwent end repair, A-tailing, adaptor ligation and AMPure XP
bead purification, then PCR amplification to construct the cDNA
library (NEBNext® Ultra™ II RNA Library Prep;
cat. no. E7775L; New England Biolabs, Inc.). The sequencing was
carried out using the NovaSeq 6000 platform (Illumina, Inc.)
employing a paired-end 150 bp approach. The final loading
concentration of the library was 1.5 µg/µl.
To generate clean data, sequencing reads were
filtered to remove adapters, ambiguous bases and low-quality
sequences. The filtered reads were aligned to the reference genome
with Hierarchical Indexing for Spliced Alignment of Transcripts
(version 2.2.1; https://daehwankimlab.github.io/hisat2/download/).
Gene expression quantification was performed using featureCounts
(version 2.0.6; https://subread.sourceforge.net/featureCounts.html) to
tally reads mapped to each gene, with results expressed as
fragments per kilobase of transcript per million (FPKM) mapped
reads. Principal component analysis was applied to the FPKM values.
Differential expression was analyzed using the ‘DESeq2’ package
(version 1.42.0; R software; Posit Software, PBC; https://bioconductor.org/packages//release/bioc/html/DESeq2.html).
Raw read count data were normalized to correct for sequencing depth
differences. Statistical models calculated P-values for each gene,
which were adjusted for multiple testing using the
Benjamini-Hochberg method to obtain adjusted P-values.
Differentially expressed genes (DEGs) were identified based on a
fold change of ≥1.2 and adjusted P≤0.05. Normalization and
hierarchical clustering of DEGs' FPKM expression matrices were
performed using the ‘ggplot2’ package in R software (version 3.0.3;
Posit Software, PBC; https://ggplot2.tidyverse.org/), with heatmap
clustering conducted using the ‘pheatmap’ package (https://cran.r-project.org/web/packages/pheatmap/index.html).
To determine impacted signaling pathways and
biological processes, Gene Set Enrichment Analysis (GSEA) was
conducted using Gene Ontology (GO; https://www.geneontology.org/) and Kyoto Encyclopedia
of Genes and Genomes (KEGG). Genes were ranked by differential
expression between groups using ‘Signal2Noise’ as the ranking
metric. Enrichment scores for each gene set were calculated using
permutation tests and normalized enrichment scores were obtained to
account for gene set size. Significance was assessed by computing
P-values and false discovery rates (FDR), with FDR <0.25
indicating significant enrichment. The EnrichmentMap tool (version
3.3.1; https://apps.cytoscape.org/apps/enrichmentmap) was
used to visualize GSEA results.
Non-targeted metabolome analysis
Untargeted metabolomic profiling was performed to
investigate treatment-related metabolic alterations in CVECs. After
treatment, CVECs were digested with trypsin for 2 min at 37˚C and
then neutralized with PBS. Cell pellets were obtained by
centrifuging at 1,000 x g for 5 min (at 4˚C; 5702 R centrifuge;
Eppendorf SE). Samples were sent to Novogene Co., Ltd. for analysis
using a liquid chromatography-tandem mass spectroscopy platform
(Thermo Fisher Scientific, Inc.).
Statistical analysis
All data are expressed as the mean ± SD. Each group
contained ≥4 replicates and CCK-8 assays were performed with a
minimum of 12 replicates per condition. Statistical analyses were
conducted using SPSS software (version 26.0; IBM Corp.). One-way
ANOVA followed by Least Significant Difference or Games-Howell
post-hoc tests was used for the datasets. The linear correlation
between the transcriptome analysis samples was assessed by
Pearson's correlation analysis and the correlation coefficient (r)
was calculated. P-values were calculated using unpaired t-test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Identification of the composition of
LA20k9
Components of LA20k9 and BMFM were analyzed
(Table SI). The most significant
difference between the two was in the organic acid component.
LA20k9 contained 26.63 mg/g of malic acid and 59.17 mg/g of
succinic acid, while these two organic acids were absent in BMFM.
These two metabolites are the main biologically active metabolites
produced by LA20k9. Malic acid and succinic acid are used in
anti-aging studies due to their excellent performance in repairing
oxidative damage and anti-inflammatory activity (25,26).
Therefore, the results suggest that LA20k9 has anti-aging
effects.
Viability of canine vascular
endothelial cells
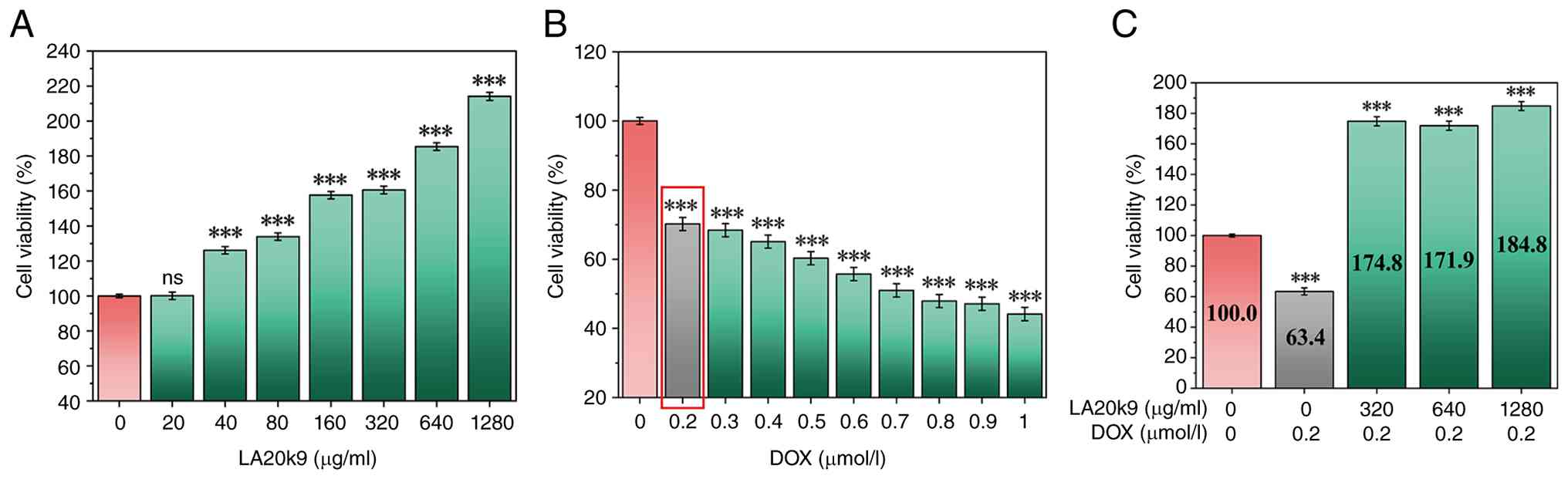
Fig. 1A shows the
cell viability after LA20k9 treatment. The three concentrations
with the highest cell survival rates (320, 640 and 1,280 µg/ml)
were selected for subsequent experiments. As shown in Fig. S1A, for BMFM, concentrations of
160, 320 and 640 µg/ml were utilized for comparative analysis.
The effects of different DOX concentrations (0-1.0
µmol/l) on cell viability were evaluated to determine the optimal
concentration for inducing aging. With increases in DOX
concentration, the viability of CVECs gradually decreased,
demonstrating DOX dose-dependency (Fig. 1B). A DOX concentration of 0.2
µmol/l significantly reduced cell viability to 70.2% (P<0.05)
and was selected as the concentration for DOX-induced CVEC
senescence.
After adding 320, 640 and 1,280 µg/ml LA20k9 to 0.2
µmol/l DOX, the viability of CVECs increased to varying degrees
(Fig. 1C). Among them, 1280 µg/ml
LA20k9 most significantly improved cell viability (P<0.05),
which was be restored to 184.8%, markedly exceeding the level of
the NC group. As shown in Fig.
S1B, after adding 320 µg/ml BMFM to 0.2 µmol/l DOX, the cell
viability of CVEC showed the most significant improvement, reaching
90.7% (P<0.05). Notably, the regenerative capacity of LA20k9
markedly surpassed that of the BMFM. This performance gap suggests
that the bioactive potency was not inherent to the raw algal
substrate but was specifically generated through B.
adolescentis fermentation. Therefore, subsequent experiments
were conducted at 0.2 µmol/l DOX + 1,280 µg/ml LA20k9 and 0.2
µmol/l DOX + 640 µg/ml BMFM.
Effect of LA20k9 on the activity of
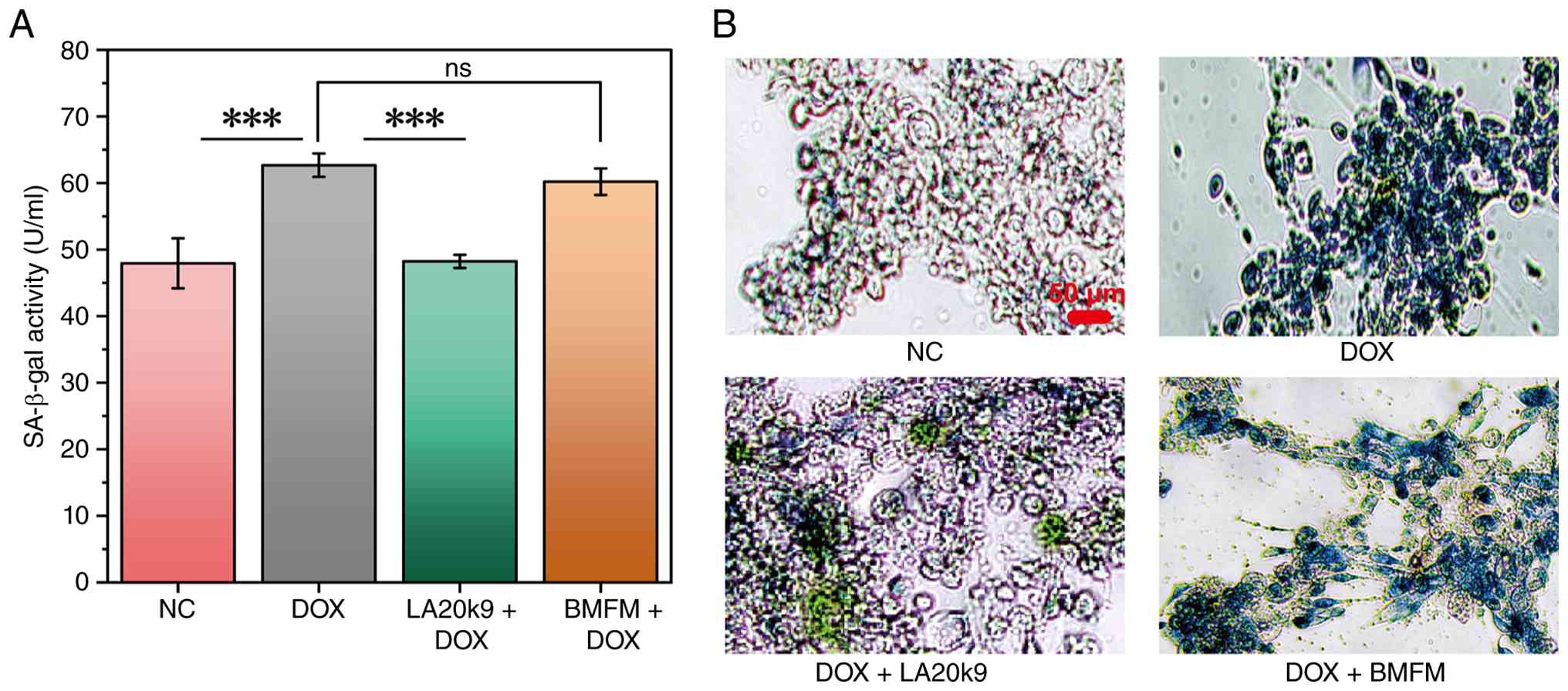
SA-β-gal in CVECs
SA-β-gal accumulates in lysosomes and has become a
commonly used biomarker for identifying senescent cells (27,28).
After treating CVECs with DOX for 24 h, the SA-β-gal activity
significantly increased by 16% compared with the control group
(P<0.05) and the number of blue-infiltrated senescent cells also
increased (Fig. 2A and B). After treatment with 1280 µg/ml
LA20k9, SA-β-gal activity was significantly reduced by 16%
(P<0.05) and the number of blue-infiltrated senescent cells was
also decreased.
By contrast, BMFM failed to mitigate this response,
exhibiting activity levels statistically indistinguishable from the
DOX model group. This quantitative profile was visually
corroborated by microscopic imaging (Fig. 2B). These results indicated that the
anti-aging effect was not derived from the non-fermented algae
extract itself but was specifically produced through the
fermentation process of Bifidobacterium adolescentis.
Effect of LA20k9 on the levels of
senescence-associated secretory phenotype (SASP) markers in
CVECs. SASP markers represent the hallmark features of cellular
senescence, influencing intercellular communication through
paracrine signaling within the tissue microenvironment and thereby
contributing to diverse physiological and pathological outcomes
(29-32).
The expression levels of SASP markers, including MMP-1, MMP-3, VEGF
and bFGF, were evaluated in CVECs to assess the impact of LA20k9 on
DOX-induced senescence.
DOX treatment significantly increased the levels of
MMP-1 and MMP-3 and significantly reduced VEGF and bFGF content
(P<0.05; Fig. 3). However,
MMP-1 and MMP-3 levels significantly decreased after LA20k9
treatment, while VEGF and bFGF content significantly increased
(P<0.001). Specifically, MMP-1 levels increased from 53.4 ng/ml
(NC) to 80.5 ng/ml (DOX) and decreased to 52.8 ng/ml with DOX +
LA20k9 treatment. MMP-3 levels increased from 21.0 ng/ml (NC) to
26.6 ng/ml (DOX) and decreased to 24.8 ng/ml with DOX + LA20k9
treatment. VEGF levels decreased from 132.9 pg/ml (NC) to 102.6
pg/ml (DOX) and increased to 129.6 pg/ml with DOX + LA20k9
treatment. bFGF levels decreased from 23.7 pg/ml (NC) to 20.8 pg/ml
(DOX) and increased to 31.4 pg/ml with DOX + LA20k9 treatment.
These results indicated that LA20k9 protected the structure and
function of vascular ECs under senescence conditions by inhibiting
the expression of MMP-1 and MMP-3 and upregulating the expression
of VEGF and bFGF in CVECs, thereby preventing DOX-induced ECM
degradation and the renewal of senescent cells.
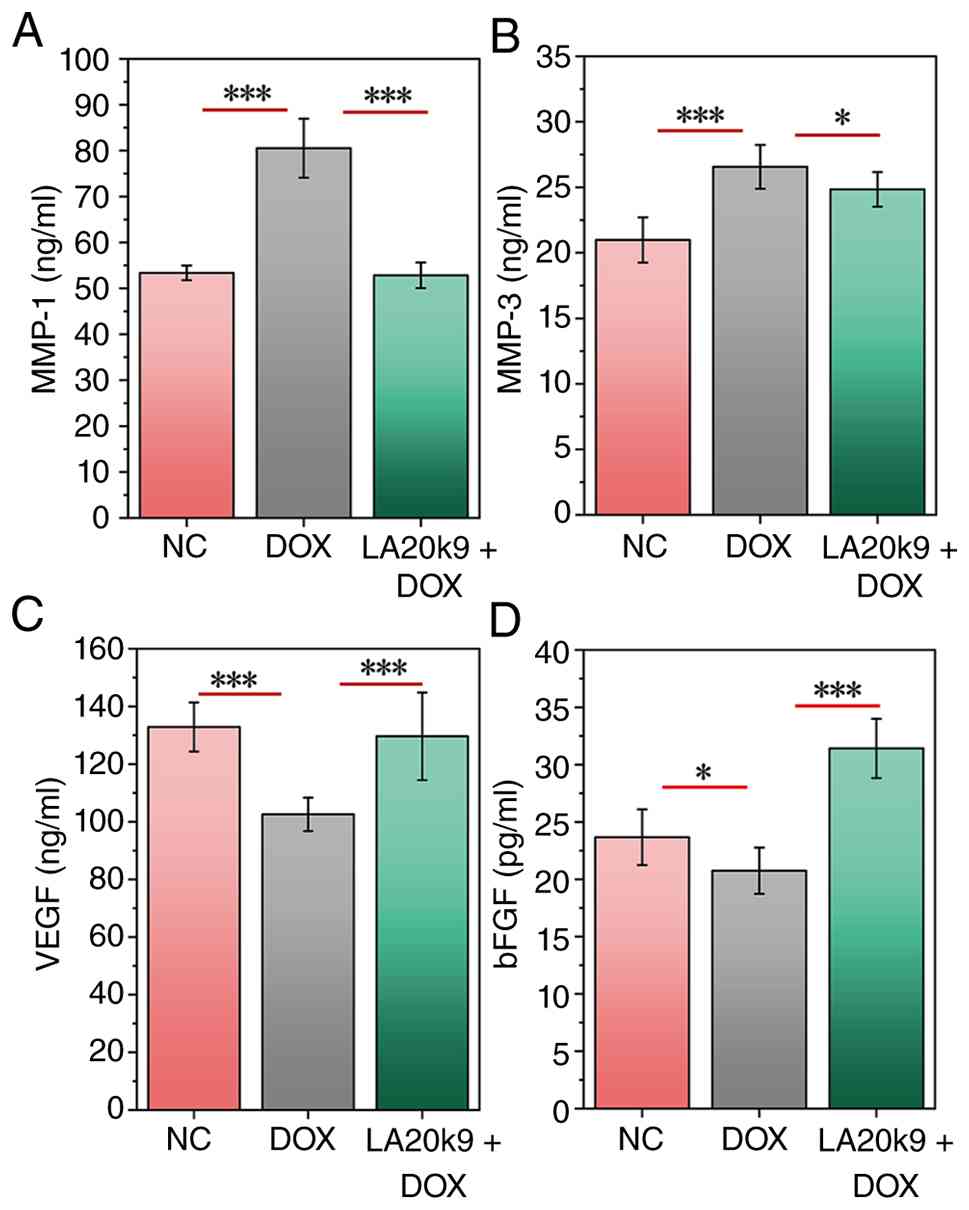 | Figure 3Effects of LA20k9 on SASP in CVECs.
Levels of (A) MMP-1, (B) MMP-3, (C) VEGF and (D) bFGF in CVECs with
DOX-induced senescence. CVECs were treated with 0.2 µmol/l DOX and
1,280 µg/ml LA20k9. SASP assays were conducted with at least four
replicates per group. Data are presented as the mean values with
error bars representing SD. Significance was determined using the
Least Significant Difference method or Games-Howell test.
*P<0.05, ***P<0.001. CVEC, canine
vascular endothelial cells; LA20k9, Life Age 20k9; bFGF, basic
fibroblast growth factor; SASP, senescence-associated secretory
phenotype; DOX, doxorubicin; NC, negative control. |
Effect of LA20k9 on the levels of
inflammatory response and oxidative stress response in CVECs
NAD+ is a central coenzyme in cells that
participates in energy metabolism, redox reactions and signal
transduction. During cellular senescence, a decline in
NAD+ levels leads to mitochondrial dysfunction, reduced
DNA repair capacity and increased inflammation. Supplementation
with NAD+ precursors, such as nicotinamide
mononucleotide, has been shown to delay the aging process (33). Meanwhile, oxidative stress and
inflammatory responses are recognized as the most significant
exogenous factors triggering the senescence response (34).
In CVECs, after DOX treatment, the levels of
inflammatory factors IL-1β, TNF-α, IL-6, IL-8, GM-CSF and MCP-1, as
well as the oxidative stress-associated index MDA, significantly
increased (P<0.05), while the NAD+ content, SOD
activity and GSH concentration significantly decreased (P<0.05;
Figs. 4 and 5). After co-treatment with LA20k9, the
levels of inflammatory factors and MDA decreased significantly
(P<0.05), while the levels of NAD+, SOD and GSH
increased significantly (P<0.05). Specifically, IL-1β levels
increased from 50.4 ng/ml (NC) to 174.5 ng/ml (DOX) and decreased
to 86.8 ng/ml with DOX + LA20k9 treatment. TNF-α levels increased
from 17.5 ng/ml (NC) to 38.3 ng/ml (DOX) and decreased to 22.9
ng/ml with DOX + LA20k9 treatment. IL-6 levels increased from 86.0
ng/ml (NC) to 116.2 ng/ml (DOX) and decreased to 92.9 ng/ml with
DOX + LA20k9 treatment. IL-8 levels increased from 45.5 ng/ml (NC)
to 55.1 ng/ml (DOX) and decreased to 45.6 ng/ml with DOX + LA20k9
treatment. GM-CSF levels increased from 174.0 ng/ml (NC) to 254.5
ng/ml (DOX) and decreased to 174.8 ng/ml with DOX + LA20k9
treatment. MCP-1 levels increased from 84.4 ng/ml (NC) to 95.7
ng/ml (DOX) and decreased to 83.6 ng/ml with DOX + LA20k9
treatment. MDA content increased from 0.151 µmol/mg prot (NC) to
0.162 µmol/mg prot (DOX) and decreased to 0.152 µmol/mg prot with
DOX + LA20k9 treatment. NAD+ content decreased from
0.978 µM (NC) to 0.426 µM (DOX) and increased to 0.719 µM with DOX
+ LA20k9 treatment. SOD activity decreased from 27.2 U/mg prot (NC)
to 12.8 U/mg prot (DOX) and increased to 32.1 U/mg prot with DOX +
LA20k9 treatment. GSH content decreased from 8.1 µM (NC) to 5.3 µM
(DOX) and increased to 7.4 µM with DOX + LA20k9 treatment. These
findings indicated that LA20k9 not only increased cell survival
rates but also restored oxidative balance and reduced inflammatory
responses.
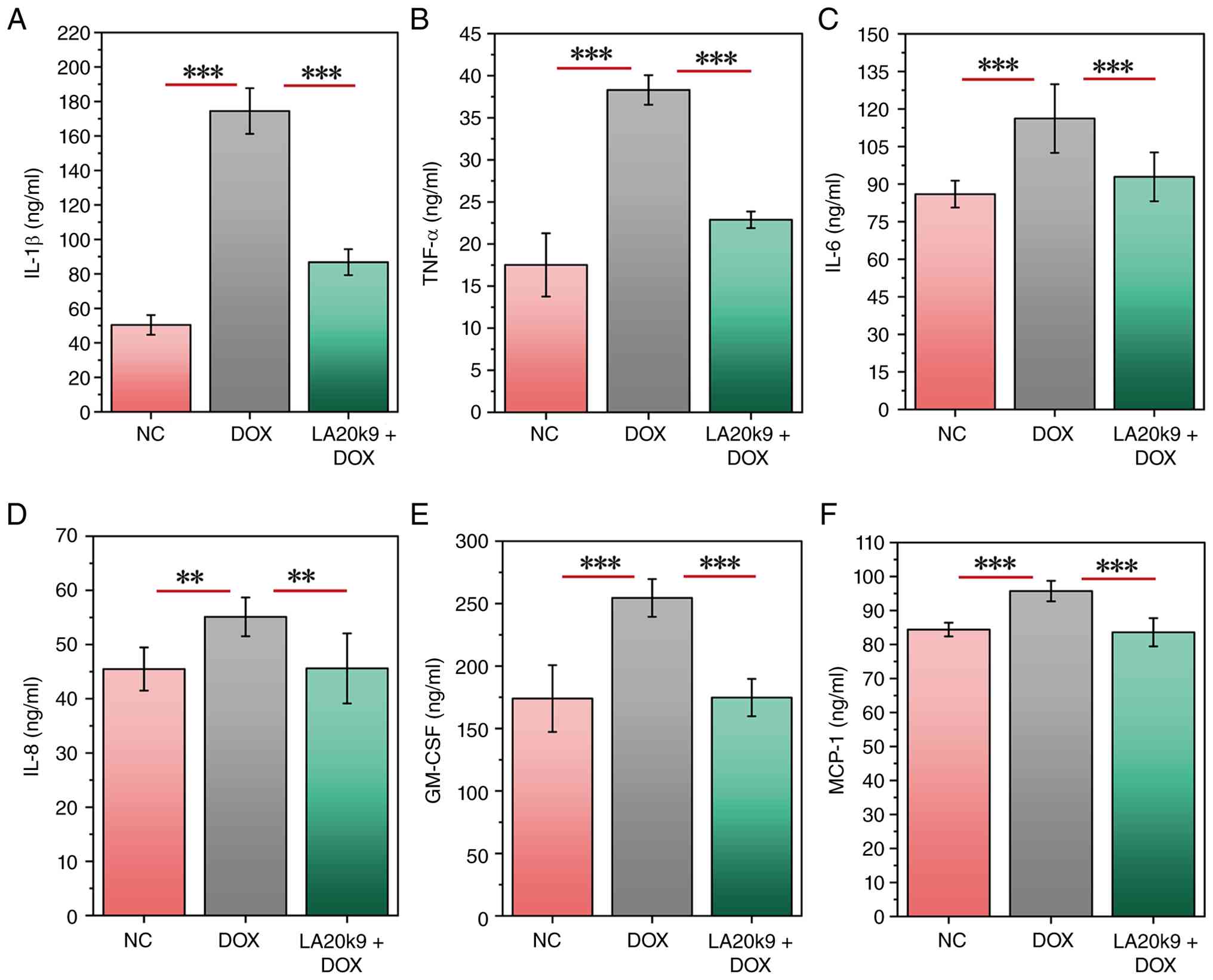 | Figure 4Effects of LA20k9 on inflammatory
cytokine levels in CVECs. Levels of inflammatory cytokines,
including (A) IL-1β, (B) IL-6, (C) IL-8, (D) TNF-α, (E) GM-CSF and
(F) MCP-1 in CVECs. Inflammatory cytokine levels assays were
performed with at least four replicates per group. Data are
presented as the mean values with error bars representing SD.
Significance was determined using the Least Significant Difference
method or Games-Howell test. **P<0.01,
***P<0.001. CVEC, canine vascular endothelial cells;
LA20k9, Life Age 20k9; DOX, doxorubicin; NC, negative control;
MCP-1, chemoattractant protein-1; GM-CSF, granulocyte-macrophage
colony-stimulating factor. |
 | Figure 5Effects of LA20k9 on NAD+
and oxidative stress levels in CVECs. (A) Levels of NAD+
in CVEC with DOX-induced senescence. CVEC were treated with 0.2
µmol/l DOX and 1,280 µg/ml LA20k9. Levels of oxidative stress
markers, including (B) SOD, (C) GSH and (D) MDA in CVEC treated
with 1,280 µg/ml LA20k9 and optimal DOX concentrations, showing
significant improvements compared with the DOX-only group.
NAD+ and oxidative stress levels assays were performed
with at least four replicates per group. Data are presented as the
mean values with error bars representing SD. Significance was
determined using the Least Significant Difference method or
Games-Howell test. P-values indicate specific comparisons between
the indicated groups. *P<0.05,
***P<0.001. CVEC, canine vascular endothelial cells;
LA20k9, Life Age 20k9; DOX, doxorubicin; NC, negative control; SOD,
superoxide dismutase; MDA, malondialdehyde; GSH, glutathione. |
Analysis of RNA-sequencing
differential gene expression
Following transcriptome sequencing, 12 cDNA
libraries were constructed, yielding a total of 35.68 Gb of raw
data. The proportion of Q30 bases exceeded 95.25% across all
samples, indicating high base-calling accuracy. Clean reads from
each sample were aligned to the reference genome, with mapping
efficiencies ranging from 85.03-92.94%. The results demonstrated
that the sequencing quality for all samples was high and suitable
for downstream analysis.
The mechanism of LA20k9 effect on aging was
investigated by conducting a statistical analysis of the DEGs
between the control group and the DOX-induced aging group, as well
as between the DOX-induced aging group and the LA20k9 intervention
group. Reliability was evaluated using Pearson's correlation
coefficient (r). The results showed that the sample r2
within each treatment group exceeded 0.80, indicating a strong
correlation (Fig. 6A). Venn
diagram analysis was used to present the number of DEGs, visually
reflecting the overlapping associations among different DEG sets
and the distribution of genes within each set. This provided clear
and visual data support for subsequent research. Venn analysis
showed that 454 genes were co-expressed in the NC group and the
LA20k9 group (Fig. 6B). The
expression profiles of the NC group and the LA20k9 group were more
similar (Fig. 6C).
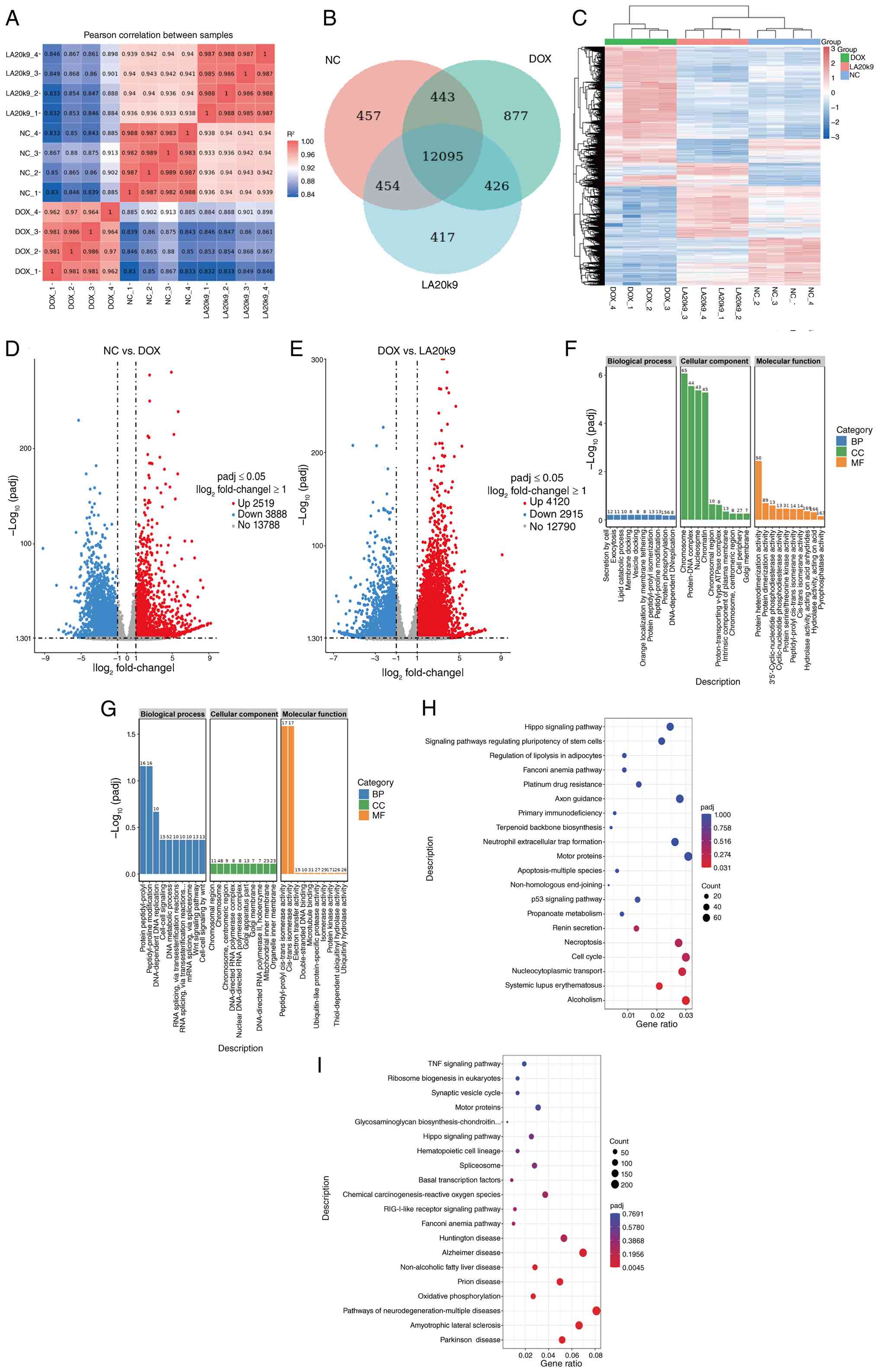 | Figure 6Transcriptomic analysis of in CVECs
upon LA20k9 treatment. (A) Inter-sample correlation heat map, (B)
Venn diagram and (C) and cluster heat map of CVECs treated with
LA20k9. The volcano plot visually displays the distribution of
differential genes in each comparison combination, including (D)
DOX vs. NC and (E) LA20k9 vs. DOX. The abscissa in the figure
represents the fold change of gene expression in the comparison
group (log2 fold change) and the ordinate represents the
significance level of the gene expression difference in the
comparison group (-log10 P-value). Upregulated genes are
represented by red dots and downregulated genes are represented by
green dots. GO functional enrichment histogram for DOX (F) and
LA20k9 (G). The ordinate is the significance level of GO term
enrichment. The value on the column represents the number of
differential genes enriched for this term. Different colors
represent the three GO subcategories of biological processes,
cellular components and molecular functions, respectively. KEGG
pathway enrichment bubble plot of DOX (H) and LA20k9 (I). The
abscissa in the figure is the ratio of the number of differential
genes annotated to the KEGG pathway to the total number of
differential genes. The ordinate is the KEGG pathway. The size of
the dot represents the number of genes annotated to the KEGG
pathway. Transcriptome sequencing was performed with at least four
replicates per group. CVEC, canine vascular endothelial cell;
LA20k9, Life Age 20k9; DOX, doxorubicin; NC, negative control;
padj, adjusted P-value; UP, upregulated; DOWN, downregulated; GO,
Gene Ontology; KEGG, Kyoto Encyclopedia of Genes and Genomes. |
DEGs were screened based on changes in gene
expression levels between the two sample groups and were further
classified into upregulated and downregulated genes according to
their expression trends. The statistical summary of DEGs between NC
and DOX (Fig. 6D and E) showed that a total of 2,519 genes were
upregulated including TOB1 (an anti-proliferative protein that
negatively regulates cell cycle progression), UBR4, TM7SF2, TUBB4B,
PIDD (a p53-induced death domain protein mediating apoptosis and
cell cycle arrest), FOSL1, ESYT3, FAAH, CD70 and CHRD. Conversely,
3,888 genes were downregulated, including WDR75, CFI (a
trypsin-like serine protease involved in complement system
regulation), HSPA13, MSH2, NOP58, CCDC112, SNX2, ZNF25, ETAA1 (an
ATR kinase activator mediating replication stress response and
genome stability maintenance) and NUDT12. The functional
annotations of the DEGs between NC and DOX are described in
Table SII.
The comparison of DOX and LA20k9 DEGs identified
4,120 upregulated genes, including EIF3E, CFI, SULT1C4, IMPACT,
PPP4R2 (a regulatory subunit of protein phosphatase 4 mediating
NF-κB signaling and DNA repair), SERINC1, SLC38A2, YME1L1, SCFD1
and ZC3H15 (a zinc-finger protein involved in mRNA processing and
cellular homeostasis). By contrast, 2,915 genes were downregulated,
including MKI67, INPP5J (an inositol 5-phosphatase regulating
phosphoinositide signaling), PLA2G3 (a secretory phospholipase A2
associated with lipid metabolism and inflammation), CYR61 (a
cellular communication network factor involved in cell adhesion,
migration and stress responses), UBR4 (an E3 ligase associated with
protein ubiquitination and stress signaling), TM7SF2, MVD, DHCR7,
FADS2 and FASN. The functional annotations of the DEGs between DOX
and LA20k9 are described in Table
SIII. These results indicate that gene expression changed
significantly under different treatment conditions and the data
quality was reliable, making it suitable for subsequent in-depth
analysis.
Analysis of differentially expressed
genes: GO and KEGG analyses
GO and KEGG pathway enrichment analyses of DEGs in
the NC vs. the DOX group and the DOX vs. the LA20k9 group were
performed to explore the effect of LA20k9 treatment on the
senescence response of CVECs.
The GO annotations of DEGs included three functional
categories: Biological processes, cellular composition and
molecular function (Fig. 6F and
G). The top terms in the DOX group
were ‘chromosome’, ‘protein-DNA complex’, ‘nucleosome’ and
‘chromatin’. DOX exhibited an upregulatory effect on these
biological functions. DOX induces DNA damage-dependent cellular
senescence by intercalating into double-stranded DNA, triggering
DNA damage and oxidative stress (35,36).
During this process, transcriptomic analysis revealed significant
associations with cellular composition elements, including
chromosomes, chromatin, nucleosomes and DNA-protein complexes.
These changes are closely associated with chromatin remodeling and
structural reinforcement that occur in senescent cells to maintain
genomic stability. The most abundant terms in the LA20k9 group were
‘peptidyl-prolyl cis-trans isomerase activity’, ‘cis-trans
isomerase activity’, ‘protein peptidyl-prolyl isomerization’ and
‘peptidyl-proline modification’. Transcriptome analysis
demonstrated that LA20k9 exhibited a downregulatory effect on these
biological functions. This suggested that LA20k9 may exert the
function associated with peptidyl-prolyl cis-trans isomerase,
reduce the levels of protein peptidyl-prolyl isomerization and
modification, thereby inhabiting the excessive activation of the
p53-centered senescence pathway, which may prevent the sustained
amplification of DNA damage downstream signaling. Ultimately, this
effects may help alleviate cellular senescence and restore the
normal cellular state. Therefore, the downregulation of these
functions is highly consistent with the anti-senescence effect of
LA20k9 against DOX-induced senescence, suggesting that inhibiting
peptidyl-prolyl cis-trans isomerase-associated activities may
represent an important molecular mechanism underlying the
protective effect of LA20k9.
Further KEGG pathway enrichment analysis was
performed to systematically characterize the functional
distribution of DEGs at the signaling pathway level. The top 20
most significantly enriched pathways are displayed in Fig. 6H and I. DOX treatment was associated with the
enrichment of genes involved in the ‘p53 signaling pathway’, a core
regulatory cascade governing the DNA damage response, cell cycle
arrest, apoptosis and cellular senescence. Mechanistically, DOX
upregulated a series of pivotal genes in this pathway, including
ATR, SESN2, GADD45G, PIDD, CCNG1/2, TP73, ZNF385A and GADD45B.
Meanwhile, DOX downregulated CCNE1 and CCNE2. Collectively, these
transcriptomic changes suggest that DOX may trigger the robust
activation of the p53-dependent DNA damage surveillance network,
which, in turn, promotes growth arrest, DNA repair responses,
apoptosis and stress-associated signaling, ultimately leading to
permanent cell cycle exit and establishing cellular senescence in
CVECs.
Further mechanistic insight was obtained by
examining the regulatory effects of LA20k9 on key signaling and
functional pathways. LA20k9 was associated with the enrichment of
genes involved in homologous recombination and cell cycle
regulation, while concurrently suppressing pathways associated with
chemical carcinogenesis and reactive oxygen species (ROS)-driven
genotoxicity. At the molecular level, these protective effects were
associated with the upregulation of a core set of genes key in DNA
double-strand break repair, DNA replication and cell cycle
progression, including RPA1, UIMC1, BRIP1, MRE11A, POLD3, MCM2,
MCM3, RAD21, ORC4, ORC5 and CCNE2. In addition, LA20k9 enhanced the
expression of genes involved in anti-inflammatory and
cytoprotective signaling, such as TANK, ATG5 and CHUK.
Concomitantly, LA20k9 downregulated genes associated with
mitochondrial dysfunction, oxidative stress and ROS-mediated
cellular damage, including UQCRC1, CYP1A1 and SLC25A5.
Collectively, these findings indicate that LA20k9 exerts its
senescence-attenuating effects by restoring homologous
recombination-mediated DNA repair, sustaining normal cell cycle
progression and alleviating ROS-associated oxidative injury. The
present findings suggest that LA20k9 could effectively counteract
the deleterious senescence program triggered by DOX, possibly
through reinforcing genomic stability, reducing oxidative stress
and normalizing cell cycle dynamics, thus potentially protecting
cells against DNA damage-induced growth arrest and functional
decline.
Analysis of non-targeted metabolome
results
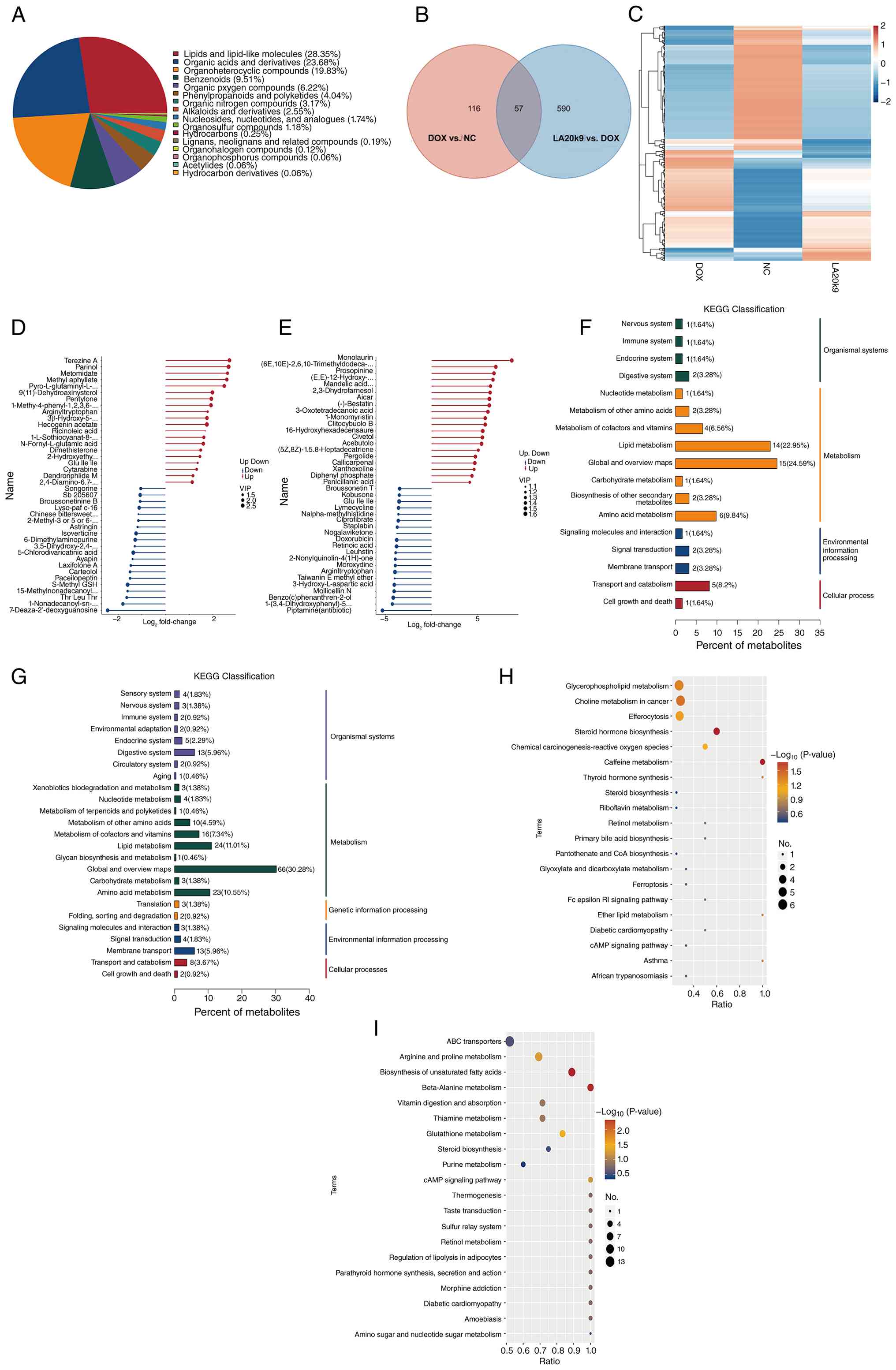
LA20k9 anti-aging mechanism was analyzed using
non-targeted metabolomics technology. Based on the aforementioned
data, systematic pathway annotations were made for all identified
metabolites and differential analysis and KEGG analysis were
conducted for all differentially expressed metabolites. Metabolites
were classified according to their chemical structure, among which,
lipids and lipid-like molecules (29.22%) accounted for a relatively
high proportion (Fig. 7A). Venn
diagrams visually present the overlap and unique metabolites
between different groups, and illustrate the relationships between
multiple groups of differentially expressed metabolites (Fig. 7B). Hierarchical cluster analysis
showed that the metabolic patterns of the NC and LA20k9 groups were
similar (Fig. 7C).
Relative to the NC group, the levels of terezine A,
parinol, metomidate, methylaphyllate,
pyro-L-glutaminyl-L-glutamine, 9(11)-dehydroaxinystero, pentylone and
arginyltryptophan were significantly upregulated in the DOX group
(Tables I and II). The majority of these metabolites
are closely associated with oxidative stress, inflammatory
activation and endothelial dysfunction, which are key pathogenic
characteristics of vascular ECs senescence. By contrast,
monolaurin, (6E,10E)-2,6,10-trimethyldodeca-2,6,10-triene,
clitocybulol B, 3-oxotetradecanoic acid, prosopinine and
(E,E)-12-hydroxy-6,10-dimethyl-6,10-dodecadien-2-one were
significantly upregulated in the LA20k9 group compared with the DOX
group. These metabolites are primarily involved in
anti-inflammatory responses, maintaining membrane stability,
improving lipid metabolism and enhancing endothelial protection,
thereby contributing to the alleviation of vascular endothelial
senescence (37-40).
 | Table IDifferential metabolites of
doxorubicin vs. negative control. |
Table I
Differential metabolites of
doxorubicin vs. negative control.
| Metabolite
name | FC | log2
FC | Up or down | P-value | VIP |
|---|
|
1-methyl-4-phenyl-1,2,3,6-2-tetrahydropyridine
N-oxide | 3.781 | 1.919 | Up | 0.006 | 2.551 |
| Metomidate | 6.127 | 2.615 | Up | 0.044 | 1.928 |
| Pentylone | 3.822 | 1.934 | Up | 0.004 | 2.576 |
|
Pyro-L-glutaminyl-L-glutamine | 5.567 | 2.477 | Up | 0.011 | 2.304 |
| Terezine A | 6.485 | 2.697 | Up | 0.001 | 2.537 |
| Methyl
aphyllate | 5.998 | 2.585 | Up | 0.024 | 2.343 |
| Parinol | 6.321 | 2.660 | Up | 0.004 | 2.759 |
|
9(11)-dehydroaxinysterol | 3.942 | 1.979 | Up | 0.010 | 2.490 |
|
Arginyltryptophan | 3.422 | 1.775 | Up | 0.034 | 1.414 |
|
3-β-hydroxy-5-cholestenoic acid | 3.354 | 1.746 | Up | 0.021 | 2.419 |
| Ayapin | 0.383 | -1.385 | Down | <0.001 | 1.224 |
| Paecilopeptin | 0.355 | -1.492 | Down | 0.010 | 1.503 |
|
7-deaza-2'-deoxyguanosine | 0.184 | -2.440 | Down | <0.001 | 2.305 |
| Carteolol | 0.356 | -1.491 | Down | 0.011 | 1.646 |
|
S-methylglutathione | 0.331 | -1.596 | Down | 0.002 | 2.664 |
|
Threonine-leucine-threonine | 0.322 | -1.634 | Down | 0.005 | 1.659 |
| Laxifolone A | 0.363 | -1.461 | Down | 0.029 | 1.491 |
|
methylnonadecanoyl-carnitine | 0.331 | -1.597 | Down | 0.019 | 1.449 |
|
1-nonadecanoyl-sn-glycero-3-phosphocholine | 0.288 | -1.796 | Down | 0.025 | 2.151 |
|
5-chlorodivaricatinic acid | 0.390 | -1.358 | Down | 0.001 | 2.700 |
| Table IIDifferential metabolites of Life Age
20k9 vs. doxorubicin. |
Table II
Differential metabolites of Life Age
20k9 vs. doxorubicin.
| Metabolite
name | FC | log2
FC | P-value | Up or down | VIP |
|---|
| 1-monomyristin | 56.856 | 5.829 | 0.001 | Up | 1.467 |
| Monolaurin | 418.773 | 8.710 | 0.001 | Up | 1.533 |
| 3-oxotetradecanoic
acid | 69.435 | 6.118 | 0.001 | Up | 1.477 |
|
(6E,10E)-2,6,10-trimethyldodeca-2,6,10-triene | 126.006 | 6.977 | 0.001 | Up | 1.508 |
|
(E,E)-12-hydroxy-6,10-dimethyl-6,10-dodecadien-2-one | 101.728 | 6.669 | 0.001 | Up | 1.455 |
|
2,3-dihydrofarnesol | 81.933 | 6.356 | 0.001 | Up | 1.454 |
| Prosopinine | 109.734 | 6.778 | 0.001 | Up | 1.483 |
|
5-aminoimidazole-4-carboxamide
ribonucleoside | 77.808 | 6.282 | 0.001 | Up | 1.564 |
| (-)-Bestatin | 77.182 | 6.270 | 0.001 | Up | 1.527 |
| Mandelic acid,
methyl ester | 84.517 | 6.401 | 0.001 | Up | 1.386 |
| Piptamine
(antibiotic) | 0.025 | -5.337 | 0.001 | Down | 1.473 |
| Mollicellin N | 0.060 | -4.068 | 0.001 | Down | 1.464 |
| Benzo
(c)phenanthren-2-ol | 0.056 | -4.166 | 0.001 | Down | 1.550 |
| Taiwanin E methyl
ether | 0.062 | -4.011 | 0.002 | Down | 1.095 |
|
3-hydroxy-L-aspartic acid | 0.060 | -4.050 | 0.002 | Down | 1.488 |
|
1-(3,4-dihydroxyphenyl)-5-hydroxy-3-decanone | 0.052 | -4.259 | 0.002 | Down | 1.389 |
|
Arginyltryptophan | 0.064 | -3.974 | 0.002 | Down | 1.562 |
|
2-nonylquinolin-4(1H)-one | 0.065 | -3.935 | 0.002 | Down | 1.321 |
| Leuhistin | 0.067 | -3.900 | 0.002 | Down | 1.242 |
| Moroxydine | 0.065 | -3.940 | 0.002 | Down | 1.340 |
KEGG pathway analysis indicated that ‘global and
overview maps’ and ‘lipid metabolism’ contained the highest number
of differentially expressed metabolites in both the DOX and LA20k9
groups, suggesting that metabolic reprogramming and lipid
homeostasis are key in regulating vascular ECs senescence (Fig. 7D and E). Enrichment analysis revealed that
pathways including ‘caffeine metabolism’, ‘thyroid hormone
synthesis’, ‘ether lipid metabolism’ and ‘asthma’ were
significantly enriched in the DOX group compared with the NC group,
with the largest number of differential metabolites involved in
‘glycerophospholipid metabolism’, ‘choline metabolism in cancer’,
‘efferocytosis’ and ‘steroid hormone biosynthesis’ (Fig. 7F and G). These pathways are closely associated
with oxidative stress injury, mitochondrial dysfunction,
inflammatory responses and endothelial barrier damage, all of which
promote the occurrence and progression of vascular endothelial
senescence induced by DOX (41-44).
Conversely, compared with the DOX group, the LA20k9 group showed
significant enrichment in the ‘cAMP signaling pathway’, ‘amino
sugar and nucleotide sugar metabolism’, ‘parathyroid hormone
synthesis, secretion and action’ and ‘retinol metabolism’ pathways.
The most highly represented pathways included ‘ABC transporters’,
‘arginine and proline metabolism’, ‘biosynthesis of unsaturated
fatty acids’, ‘β-alanine metabolism’ and ‘vitamin digestion and
absorption’. The cAMP pathway protects endothelial function by
mitigating inflammation, oxidative stress and SASP. Retinol
metabolism sustains endothelial integrity and delays senescence.
Arginine/proline metabolism optimizes vascular homeostasis, while
unsaturated fatty acid biosynthesis preserves membrane and
mitochondrial function. ABC transporters alleviate intracellular
metabolic damage (45-49).
This result therefore suggests that LA20k9 improves vascular
endothelial function and resists cellular senescence mainly by
regulating metabolic homeostasis, activating the cAMP signaling
pathway to inhibit inflammation and oxidative stress, enhancing
retinol metabolism and endothelial repair capacity and maintaining
lipid balance and mitochondrial function, thereby exerting
protective effects against DOX-induced vascular ECs senescence.
Discussion
CVDs represent a leading cause of morbidity and
mortality in the elderly population and advancing age is recognized
as a key risk factor for the development and progression of CVDs
(2). Although previous studies
have demonstrated that fermented plant-derived extracts exhibit
promising cardiovascular protective potential due to their
antioxidant, anti-inflammatory, antimicrobial and organ-protective
properties (50-52),
such investigations have generally been limited by the use of
non-species-specific probiotic strains, cross-species cellular
models and insufficient mechanistic validation.
The present study advances this field through three
dimensions. First, a novel canine-specific probiotic strain
(Bifidobacterium adolescentis Life Age 20-k9) isolated from
long-lived dogs was employed, which better matches canine
physiological and pathological features and thereby improves
biological relevance. Second, mechanistic investigations were
performed in a targeted CVECs model rather than relying on
cross-species systems, thus minimizing interspecies variation and
enabling species-specific insights into vascular endothelial
senescence. Third, by integrating functional assays,
transcriptomics and metabolomics, deeper multi-omics mechanistic
evidence that extends beyond preliminary efficacy observations and
reveals key regulatory pathways underlying the anti-senescent
effects was provided, including the p53 signaling pathway, cell
cycle progression, cAMP signaling and retinol metabolism.
In the present study, LA20k9 enhanced cell viability
and reduced the release of SA-β-gal activity associated with aging.
Only an LA20k9 concentration of 1,280 µg/ml enhanced the
NAD+ content in CVECs. In normal cells, the SASP gene is
typically tightly suppressed to prevent the inappropriate
expression of inflammatory signals. However, in senescent cells,
the SASP gene becomes highly upregulated (53). Elevated MMP expression has been
shown to promote the degradation of extracellular matrix
components, such as collagen fibers and hyaluronic acid, thereby
accelerating cellular aging (54,55).
With respect to endothelial function, VEGF exerts pleiotropic
effects on vascular ECs, promoting angiogenesis by stimulating ECs
proliferation and migration, as well as modulating gene expression
profiles. Notably, VEGF has been reported to delay the onset of
senescence in microvascular ECs and potentially reverse
age-associated changes (56).
Furthermore, certain FGF family members, including bFGF, serve key
roles in metabolic regulation and tissue homeostasis, supporting
the regeneration of damaged tissues and facilitating the repair of
aged tissues (57). Treatment with
LA20k9 decreased MMP-1 and MMP-3 content and increased VEGF and
bFGF content in DOX-stimulated aging CVECs. Aging cells produce a
number of inflammatory cytokines and an increase in inflammatory
factors will accelerate cellular senescence (58). Treatment with LA20k9 decreased
IL-1β, IL-6, IL-8, MCP-1, GM-CSF and TNF-α content in
DOX-stimulated aging CVECs. LA20k9 also alleviated the oxidative
damage caused by DOX induction.
The functions of key signaling pathways and their
regulation by LA20k9 were systematically summarized to further
understand the anti-aging mechanisms of LA20k9. The p53 signaling
pathway is a central pathway governing cell survival, DNA damage
repair, cell cycle progression and apoptosis. As a key tumor
suppressor, p53 is activated under stress conditions such as DNA
damage and then stabilized through phosphorylation and acetylation
(59,60). Activated p53 upregulates
p21WAF1/CIP1 and 14-3-3σ, leading to cell cycle arrest in the
G1/S and G2/M phases. In the present study,
DOX significantly increased the mRNA expression of ATR, SESN2,
GADD45G, PIDD and TP73, suggesting that the p53 pathway may be
activated, which may have promoted DNA damage, apoptosis and
cellular senescence in CVECs (61). LA20k9 could counteract these
effects by potentially upregulating homologous recombination and
normal cell cycle progression, while possibly downregulating
chemical carcinogenesis-ROS, which may contribute to the
alleviation of DOX-induced senescence.
The cAMP signaling pathway functions as a central
second-messenger system involved in mitochondrial function,
oxidative stress and aging. cAMP is synthesized from ATP by
adenylate cyclase and acts through downstream effectors, including
cyclic-AMP dependent protein kinase A, exchange protein activated
by cAMP and cyclic nucleotide gated ion channels. This pathway is
widely implicated in age-associated cardiovascular and metabolic
diseases (62-67).
In the present study, non-targeted metabolomics suggested that
LA20k9 may activate the cAMP signaling pathway, which could
contribute to improved mitochondrial function as well as
potentially reduced cellular senescence and inflammation. Retinol
metabolism is another key pathway regulating extracellular matrix
synthesis and anti-aging processes. Retinol promotes the production
of collagen and glycosaminoglycans, inhibits MMP activity and
exerts antioxidant effects (68,69).
The present study suggested that LA20k9 may activate retinol
metabolism, potentially accelerate collagen formation and appear to
suppress matrix degradation, thereby possibly delaying cellular
senescence at the metabolic level. Collectively, LA20k9 may
attenuate endothelial senescence by coordinately regulating the p53
signaling pathway, cAMP signaling pathway and retinol metabolism,
which may exert distinct but complementary functions.
In conclusion, the evidence reported in the present
study demonstrated that LA20k9 exerted a significant anti-aging
protective effect in CVECs. LA20k9 markedly enhanced cell viability
and reduced senescence-associated SA-β-Gal activity. Furthermore,
LA20k9 significantly downregulated the expression of
aging-associated phenotypic markers MMP-1 and MMP-3, while
concomitantly increasing VEGF and bFGF levels, which were important
for vascular integrity and function. Notably, LA20k9 mitigated
DOX-induced stress injury by increasing intracellular levels of
NAD+, SOD and GSH and reducing MDA concentrations.
At the transcriptional level, LA20k9 counteracted
DOX-induced cellular senescence by promoting normal cell-cycle
progression and suppressing chemical carcinogenesis-ROS. At the
metabolic level, it activated the cAMP signaling pathway and
retinol metabolism, thereby restoring mitochondrial function and
suppressing cellular senescence and inflammatory responses. These
results underscore the therapeutic potential of LA20k9 in combating
vascular aging and improving vascular health. The findings of the
present study support the development of post-biological agents as
a novel and promising strategy for ameliorating age-related
cardiovascular decline in animals.
Despite this, the present study has its limitations,
mainly reflected in the following two aspects: First, the present
study was conducted only at the in vitro cellular level to
investigate anti-aging effects, lacking corresponding in
vivo validation using animal models; and second, the underlying
anti-aging mechanisms remain to be further elucidated, especially
through verification in appropriate animal models. In the future,
animal experiments will be carried out in vivo by
establishing a mouse model to systematically explore the anti-aging
effects of metabolites derived from brown algae extract fermented
by canine-origin Bifidobacterium. Despite these limitations,
the present study still provides valuable insights into the
development of postbiotic fermented metabolites with anti-aging
properties and their potential applications in anti-aging
research.
Supplementary Material
Effects of the BMFM on cell viability
in CVECs. (A) Cell viability of CVECs treated with varying
concentrations of BMFM (20-1,280 μg/ml). (B) Cell viability
of CVECs treated with BMFM (160, 320 and 640 μg/ml) in the
presence of DOX (0.2 μmol/l). Cell viability assays were
performed with at least six replicates per group. Data are
presented as the mean values with error bars representing SD.
*P<0.05 and ***P<0.001. BMFM,
non-fermented blank matrix; CVEC, canine vascular endothelial cell;
DOX, doxorubicin.
Composition analysis of LA20k9 and
BMFM.
Differential genes of DOX vs. NC.
Differential genes of DOX vs.
LA20k9.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The RNA-sequencing data generated in the present
study may be found in the NCBI Sequence Read Archive under
accession number PRJNA1451389 or at the following URL: https://www.ncbi.nlm.nih.gov/search/all/?term=PRJNA1451389.
The metabolomics data generated in the present study may be found
in the Open Archive for Miscellaneous Data (China National Center
for Bioinformation/Beijing Institute of Genomics, Chinese Academy
of Sciences) under accession number OMIX016205 or at the following
URL: https://ngdc.cncb.ac.cn/omix/release/OMIX016205.
Authors' contributions
HCJ designed the present research and wrote the
original manuscript draft. HSB, TL and HYW analyzed the
experimental data. YLL and ZZW participated in the conception and
design of the research, provided resources and supervised the work.
HCJ, HSB, TL, HYW, YLL and ZZW confirm the authenticity of all the
raw data. All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
López-Otín C, Blasco MA, Partridge L,
Serrano M and Kroemer G: Hallmarks of aging: An expanding universe.
Cell. 186:243–278. 2023.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Chmielewski PP, Data K, Strzelec B,
Farzaneh M, Anbiyaiee A, Zaheer U, Uddin S, Sheykhi-Sabzehpoush M,
Mozdziak P, Zabel M, et al: Human aging and age-related diseases:
From underlying mechanisms to pro-longevity interventions. Aging
Dis. 16:1853–1877. 2024.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Augustin HG and Koh GY: A systems view of
the vascular endothelium in health and disease. Cell.
187:4833–4858. 2024.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Fajemiroye JO, da Cunha LC,
Saavedra-Rodríguez R, Rodrigues KL, Naves LM, Mourão AA, da Silva
EF, Williams NEE, Martins JLR, Sousa RB, et al: Aging-induced
biological changes and cardiovascular diseases. Biomed Res Int.
2018(7156435)2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Li Z, Zhang Z, Ren Y, Wang Y, Fang J, Yue
H, Ma S and Guan F: Aging and age-related diseases: From mechanisms
to therapeutic strategies. Biogerontology. 22:165–187.
2021.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Xie S, Xu SC, Deng W and Tang Q: Metabolic
landscape in cardiac aging: Insights into molecular biology and
therapeutic implications. Signal Transduct Target Ther.
8(114)2023.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Gude NA, Broughton KM, Firouzi F and
Sussman MA: Cardiac ageing: Extrinsic and intrinsic factors in
cellular renewal and senescence. Nat Rev Cardiol. 15:523–542.
2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Pagan LU, Gomes MJ, Gatto M, Mota GAF,
Okoshi K and Okoshi MP: The role of oxidative stress in the aging
heart. Antioxidants (Basel). 11(336)2022.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Zhang L, Pitcher LE, Yousefzadeh MJ,
Niedernhofer LJ, Robbins PD and Zhu Y: Cellular senescence: A key
therapeutic target in aging and diseases. J Clin Invest.
132(e158450)2022.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Campisi J, Andersen JK, Kapahi P and Melov
S: Cellular senescence: A link between cancer and age-related
degenerative disease? Semin Cancer Biol. 21:354–359.
2011.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Minamino T, Miyauchi H, Yoshida T, Ishida
Y, Yoshida H and Komuro I: Endothelial cell senescence in human
atherosclerosis: Role of telomere in endothelial dysfunction.
Circulation. 105:1541–1544. 2002.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Bu LL, Yuan HH, Xie LL, Guo MH, Liao DF
and Zheng XL: New dawn for atherosclerosis: Vascular endothelial
cell senescence and death. Int J Mol Sci. 24(15160)2023.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Lewis-McDougall FC, Ruchaya PJ,
Domenjo-Vila E, Shin Teoh T, Prata L, Cottle BJ, Clark JE, Punjabi
PP, Awad W, Torella D, et al: Aged-senescent cells contribute to
impaired heart regeneration. Aging cell. 18(e12931)2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Yu S, Kim SR, Jiang K, Ogrodnik M, Zhu XY,
Ferguson CM, Tchkonia T, Lerman A, Kirkland JL and Lerman LO:
Quercetin Reverses cardiac systolic dysfunction in mice fed with a
high-fat diet: Role of angiogenesis. Oxid Med Cell Longev.
2021(8875729)2021.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Wang L, Wang B, Gasek NS, Zhou Y, Cohn RL,
Martin DE, Zuo W, Flynn WF, Guo C, Jellison ER, et al: Targeting
p21Cip1 highly expressing cells in adipose tissue
alleviates insulin resistance in obesity. Cell Metab. 34:75–89.e8.
2022.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Bloom SI, Islam MT, Lesniewski LA and
Donato AJ: Mechanisms and consequences of endothelial cell
senescence. Nat Rev Cardiol. 20:38–51. 2023.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Thorakkattu P, Khanashyam AC, Shah K, Babu
KS, Mundanat AS, Deliephan A, Deokar GS, Santivarangkna C and
Nirmal NP: Postbiotics: Current trends in food and pharmaceutical
industry. Foods. 11(3094)2022.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Wegh CAM, Geerlings SY, Knol J, Roeselers
G and Belzer C: Postbiotics and their potential applications in
early life nutrition and beyond. Int J Mol Sci.
20(4673)2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Leser T and Baker A: Bifidobacterium
adolescentis-a beneficial microbe. Benef Microbes. 14:525–551.
2023.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Li Y, Wang Y, Lu Y, Zhang M, Yan Z, Daun Y
and Ma H: Breeding and application effect evaluation of
Bifidobacterium adolescentis derived from felines with long
life spans. Acta Microbiol Sin. 65:4938–4950. 2025.
|
|
21
|
Zhu M, Hsu CW, Peralta Ogorek LL, Taylor
IW, La Cavera S, Oliveira DM, Verma L, Mehra P, Mijar M, Sadanandom
A, et al: Single-cell transcriptomics reveal how root tissues adapt
to soil stress. Nature. 642:721–729. 2025.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Schrimpe-Rutledge AC, Codreanu SG, Sherrod
SD and McLean JA: Untargeted metabolomics strategies-challenges and
emerging directions. J Am Soc Mass Spectrom. 27:1897–1905.
2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Mohammed Y, Pan J, Zhang S, Han J and
Borchers CH: ExSTA: External standard addition method for accurate
high-throughput quantitation in targeted proteomics experiments.
Proteomics Clin Appl. 12(1600180)2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Moskovets E, Chen HS, Pashkova A, Rejtar
T, Andreev V and Karger BL: Closely spaced external standard: A
universal method of achieving 5 ppm mass accuracy over the entire
MALDI plate in axial matrix-assisted laser desorption/ionization
time-of-flight mass spectrometry. Rapid Commun Mass Spectrom.
17:2177–2187. 2003.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Wu JL, Wu QP, Yang XF, Wei MK, Zhang JM,
Huang Q and Zhou XY: L-malate reverses oxidative stress and
antioxidative defenses in liver and heart of aged rats. Physiol
Res. 57:261–268. 2008.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Fang W, Chen S, Jin X, Liu S, Cao X and
Liu B: Metabolomics in aging research: Aging markers from organs.
Front Cell Dev Biol. 11(1198794)2023.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Chen X, Wu R, Li L, Zeng Y, Chen J, Wei M,
Feng Y, Chen G, Wang Y, Lin L, et al: Pregnancy-induced changes to
the gut microbiota drive macrophage pyroptosis and exacerbate
septic inflammation. Immunity. 56:336–352.e9. 2023.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Zeng Y, Wu R, Wang F, Li S, Li L, Li Y,
Qin P, Wei M, Yang J, Wu J, et al: Liberation of daidzein by gut
microbial β-galactosidase suppresses acetaminophen-induced
hepatotoxicity in mice. Cell Host Microbe. 31:766–780.e7.
2023.PubMed/NCBI View Article : Google Scholar
|
|
29
|
D'Ambrosio M and Gil J: Reshaping of the
tumor microenvironment by cellular senescence: An opportunity for
senotherapies. Dev Cell. 58:1007–1021. 2023.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Hou J and Kim S: Possible role of
ginsenoside Rb1 in skin wound healing via regulating senescent skin
dermal fibroblast. Biochem Biophys Res Commun. 499:381–388.
2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Huang YH, Chen MH, Guo QL, Chen ZX, Chen
QD and Wang XZ: Interleukin-10 induces senescence of activated
hepatic stellate cells via STAT3-p53 pathway to attenuate liver
fibrosis. Cell Signal. 66(109445)2020.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Mosteiro L, Pantoja C, Alcazar N, Marión
RM, Chondronasiou D, Rovira M, Fernandez-Marcos PJ, Muñoz-Martin M,
Blanco-Aparicio C, Pastor J, et al: Tissue damage and senescence
provide critical signals for cellular reprogramming in vivo.
Science. 354(aaf4445)2016.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Ungvari Z, Tarantini S, Kiss T, Wren JD,
Giles CB, Griffin CT, Murfee WL, Pacher P and Csiszar A:
Endothelial dysfunction and angiogenesis impairment in the ageing
vasculature. Nat Rev Cardiol. 15:555–565. 2018.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Huang W, Hickson LJ, Eirin A, Kirkland JL
and Lerman LO: Cellular senescence: The good, the bad and the
unknown. Nat Rev Nephrol. 18:611–627. 2022.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Huang P, Bai L, Liu L, Fu J, Wu K, Liu H,
Liu Y and Qi B and Qi B: Redd1 knockdown prevents
doxorubicin-induced cardiac senescence. Aging (Albany NY).
13:13788–13806. 2021.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Shiheido-Watanabe Y, Sung EA, Ivessa A,
Zhai P, Takada T, Ikeda S, Matsushita M, Zablocki D and Sadoshima
J: Inhibition of PAI-1 shifts cardiomyocyte fate from senescence
toward apoptosis and mitigates doxorubicin-induced cardiotoxicity.
Geromedicine. 2(202521)2026.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Laowansiri M, Suwanchote S, Wannigama DL,
Badavath VN, Hongsing P, Edwards SW, Suratannon N, Chatchatee P,
Lertpichitkul P, Rerknimitr P, et al: Monolaurin inhibits
antibiotic-resistant Staphylococcus aureus in patients with atopic
dermatitis. Sci Rep. 15(23180)2025.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Luo Y, Zhang Z, Zheng W, Zeng Z, Fan L,
Zhao Y, Huang Y, Cao S, Yu S and Shen L: Molecular mechanisms of
plant extracts in protecting aging blood vessels. Nutrients.
16(2357)2024.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Zheng Y, Pang H, Wang J, Shi G and Huang
J: New apoptosis-inducing sesquiterpenoids from the mycelial
culture of Chinese edible fungus Pleurotus cystidiosus. J Agric
Food Chem. 63:545–551. 2015.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Bourrinet P and Quevauviller A:
Prosopinine, an alkaloid from Prosopis africana (Legumineous). Its
effects on the central and autonomic nervous systems C R Seances
Soc Biol Fil. 162:1138–1140. 1968.PubMed/NCBI(In French).
|
|
41
|
Śliwińska S and Jeziorek M: The role of
nutrition in Alzheimer's disease. Rocz Panstw Zakl Hig. 72:29–39.
2021.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Mammen JSR: Thyroid and aging. Endocrinol
Metab Clin North Am. 52:229–243. 2023.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Papsdorf K, Miklas JW, Hosseini A, Cabruja
M, Morrow CS, Savini M, Yu Y, Silva-García CG, Haseley NR, Murphy
LM, et al: Lipid droplets and peroxisomes are co-regulated to drive
lifespan extension in response to mono-unsaturated fatty acids. Nat
Cell Biol. 25:672–684. 2023.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Budde J and Skloot GS: Is aging a
‘comorbidity’ of asthma? Pulm Pharmacol Ther. 52:52–56.
2018.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Aslam M and Ladilov Y: Emerging role of
cAMP/AMPK signaling. Cells. 11(308)2022.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Quan T: Human skin aging and the
anti-aging properties of retinol. Biomolecules.
13(1614)2023.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Rosell-Díaz M, Petit-Gay A, Molas-Prat C,
Gallardo-Nuell L, Ramió-Torrentà L, Garre-Olmo J, Pérez-Brocal V,
Moya A, Jové M, Pamplona R, et al: Metformin-induced changes in the
gut microbiome and plasma metabolome are associated with cognition
in men. Metabolism. 157(155941)2024.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Hampton K, Polski-Delve A, Hellmich C and
Rushworth SA: Linking mitochondria, fatty acids, and hematopoietic
stem cell expansion during infection: Implications for aging and
metabolic diseases. Stem Cells. 43(sxaf053)2025.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Jungwirth H and Kuchler K: Yeast ABC
transporters-a tale of sex, stress, drugs and aging. FEBS Let.
580:1131–1138. 2006.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Jha NK, Sharma C, Hashiesh HM, Arunachalam
S, Meeran MN, Javed H, Patil CR, Goyal SN and Ojha S:
β-Caryophyllene, a natural dietary CB2 receptor selective
cannabinoid can be a candidate to target the trinity of infection,
immunity, and inflammation in COVID-19. Front Pharmacol.
12(590201)2021.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Wu L, He H, Liang T, Du G, Li L, Zhong H,
Li Y, Zhang J, Chen N, Jiang T, et al: Mesaconic acid as a key
metabolite with anti-inflammatory and anti-aging properties
produced by Lactobacillus plantarum 124 from centenarian gut
microbiota. NPJ Biofilms Microbiomes. 11(165)2025.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Liu R and Sun B: Lactic acid bacteria and
aging: Unraveling the interplay for healthy longevity. Aging Dis.
15:1487–1498. 2023.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Keshavarz M, Xie K, Schaaf K, Bano D and
Ehninger D: Targeting the ‘hallmarks of aging’ to slow aging and
treat age-related disease: Fact or fiction? Mol Psychiatry.
28:242–255. 2023.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Kim J, Lee CW, Kim EK, Lee SJ, Park NH,
Kim HS, Kim HK, Char K, Jang YP and Kim JW: Inhibition effect of
Gynura procumbens extract on UV-B-induced matrix-metalloproteinase
expression in human dermal fibroblasts. J Ethnopharmacol.
137:427–433. 2011.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Suganuma K, Nakajima H, Ohtsuki M and
Imokawa G: Astaxanthin attenuates the UVA-induced up-regulation of
matrix-metalloproteinase-1 and skin fibroblast elastase in human
dermal fibroblasts. J Dermatol Sci. 58:136–142. 2010.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Li J, Sun Z, Cui Y, Qin L, Wu F, Li Y, Du
N and Li X: Knockdown of LMNB1 inhibits the proliferation of lung
adenocarcinoma cells by inducing DNA damage and cell senescence.
Front Oncol. 12(913740)2022.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Ma Y, Wang S, Wang H, Chen X, Shuai Y,
Wang H, Mao Y and He F: Mesenchymal stem cells and dental implant
osseointegration during aging: From mechanisms to therapy. Stem
Cell Res Ther. 14(382)2023.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Mohammad S and Thiemermann C: Role of
metabolic endotoxemia in systemic inflammation and potential
interventions. Front Immunol. 11(594150)2021.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Zong Z, Xie F, Wang S, Wu X, Zhang Z, Yang
B and Zhou F: Alanyl-tRNA synthetase, AARS1, is a lactate sensor
and lactyltransferase that lactylates p53 and contributes to
tumorigenesis. Cell. 187:2375–2392.e33. 2024.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Engeland K: Cell cycle regulation:
p53-p21-RB signaling. Cell Death Differ. 29:946–960.
2022.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Zhang L, Pan L, Xiang B, Zhu H, Wu Y, Chen
M, Guan P, Zou X, Valencia CA, Dong B, et al: Potential role of
exosome-associated microRNA panels and in vivo environment to
predict drug resistance for patients with multiple myeloma.
Oncotarget. 7:30876–30891. 2016.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Nwabuo CC and Vasan RS: Pathophysiology of
hypertensive heart disease: Beyond left ventricular hypertrophy.
Curr Hypertens Rep. 22(11)2020.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Dagda RK, Gusdon AM, Pien I, Strack S,
Green S, Li C, Van Houten B, Cherra SJ III and Chu CT:
Mitochondrially localized PKA reverses mitochondrial pathology and
dysfunction in a cellular model of Parkinson's disease. Cell Death
Differ. 18:1914–1923. 2011.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Signorile A, Ferretta A, Pacelli C,
Capitanio N, Tanzarella P, Matrella ML, Valletti A, De Rasmo D and
Cocco T: Resveratrol treatment in human parkin-mutant fibroblasts
modulates cAMP and calcium homeostasis regulating the expression of
mitochondria-associated membranes resident proteins. Biomolecules.
11(1511)2021.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Sanders O and Rajagopal L:
Phosphodiesterase inhibitors for Alzheimer's disease: A systematic
review of clinical trials and epidemiology with a mechanistic
rationale. J Alzheimers Dis Rep. 4:185–215. 2020.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Wiciński M, Socha M, Malinowski B,
Wódkiewicz E, Walczak M, Górski K, Słupski M and Pawlak-Osińska K:
Liraglutide and its neuroprotective properties-focus on possible
biochemical mechanisms in Alzheimer's disease and cerebral ischemic
events. Int J Mol Sci. 20(1050)2019.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Valenti D, De Rasmo D, Signorile A, Rossi
L, de Bari L, Scala I, Granese B, Papa S and Vacca RA:
Epigallocatechin-3-gallate prevents oxidative phosphorylation
deficit and promotes mitochondrial biogenesis in human cells from
subjects with Down's syndrome. Biochim Biophys Acta. 1832:542–552.
2013.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Temova Rakuša Ž, Škufca P, Kristl A and
Roškar R: Retinoid stability and degradation kinetics in commercial
cosmetic products. J Cosmet Dermatol. 20:2350–2358. 2021.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Kim J, Lee SG, Lee J, Choi S, Suk J, Lee
JH, Yang JH, Yang JS and Kim J: Oral supplementation of
low-molecular-weight collagen peptides reduces skin wrinkles and
improves biophysical properties of skin: A randomized,
double-blinded, placebo-controlled study. J Med Food. 25:1146–1154.
2022.PubMed/NCBI View Article : Google Scholar
|