Introduction
Grapes are fruit that botanically belong to berries
and to the plant genus Vitis. They consist of juice, pulp,
skin, seeds and stem. Grapes exert established nutritional and
medicinal properties, as they are an excellent source of water,
carbohydrates, proteins and fat (1). The main properties grapes are
attributed to their antioxidant, anticarcinogenic,
immune-modulatory, antidiabetic, anti-atherogenic, neuroprotective,
anti-obesity, anti-aging and anti-infection activities. Moreover,
grapes represent an excellent source of water (82%), carbohydrates
(12-18%), proteins (0.5-0.6%) and fats (0.3-0.4%). Lipids are
mainly found in the seeds of grapes and include fatty acids,
tocopherols, tocotrienols and phytosterols. Different grape
varieties have different concentrations and profiles of fatty
acids. Apart from the aforementioned properties, grapes are an
essential source of molecules with potent antioxidant properties,
mainly polyphenolic compounds (2).
Polyphenols are abundant in grapes; they contribute to the defense
of the plant against bacteria and environmental factors (3) and are synthesized through the
phenylpropanoid pathway in response to biotic and abiotic stimuli
(4). It has been reported that
polyphenols are beneficial for human health due to their
antioxidant activity, which in some cases is greater than that of
vitamins (5-7).
Grapes and their products are widely consumed as
dietary components worldwide and in particular, as fresh fruit,
dried fruit, wine and juice (8).
Wine is considered the major product of grapes, since it is a
fundamental constituent for social, religious and cultural events
(9). The polyphenolic compounds
which are transferred from grapes to wine are dependent on the
winemaking process (10).
Polyphenolic compounds are major constituents of wine that largely
determine the quality of the wine due to their direct effects on
color, flavor, bitterness, astringency and aroma (11). Following the vinification process,
large amounts of solid organic waste and byproducts are generated,
including grape pomace (62%), wine lees (14%), grape stems (12%)
and dewatered sludge (12%) (12).
Additionally, a large number of polyphenols remain in the
vinification waste, which form >13% of the processed grape
weight (13). Thus, it is of
utmost importance that the scientific community identifies
strategies with which to exploit the byproducts from the winemaking
procedure (i.e., stems), in order to diminish environmental
pollution. The most common method of exploiting grape stems is the
production of compost and energy; however, the polyphenols present
in these are wasted through that process (14). It is noteworthy that the
reintegration of wine byproducts into the food chain is of high
economic, nutritional and environmental interest (15). Indeed, they can improve the
nutritional value of baked, pastry and pasta products (16), whilst they can potentially be
incorporated into dietary supplements (17).
It appears that byproducts of the winemaking
procedure, such as grape stems and pomace are a pollutant when
discarded to the environment due to their high organic load.
Nevertheless, due to their bioactive compounds (i.e., polyphenols),
they possess notable biological properties, mainly acting
protectively against oxidative stress and related pathologies
(18,19). Of note, it has been observed that
grape (by)products improve the redox status of productive (20,21)
and experimental animals (22),
whereas they function as potent antioxidants in vitro
(19,22-24).
The content of polyphenols in grape stems is highly dependent on
the variety and growing conditions (25). However, there is also evidence to
indicate that polyphenols may function as prooxidants, a fact that
is of utmost interest in terms of administration to cancer cells
(26,27).
The present study is part of a greater project; the
first article of this project examined the in vitro
antioxidant and antimutagenic properties of the same grape stem
extracts (19). According to the
methodological practice recently proposed, the examination of the
biological properties of plant extracts in the cellular environment
constitutes the second line of screening for such compounds
(28). To that end, the main
objective of the present study was to determine the effects of the
aforementioned extracts on the redox status [i.e., the levels of
the endogenous reduced form of glutathione (GSH) and reactive
oxygen species (ROS)] of two human cancer cell lines. The results
of the present study are anticipated to provide new insight into
the biological activity of the tested extracts, that may be further
utilized by introducing them into animal and subsequently, the
human diet (i.e., as components of bio-functional foods).
Materials and methods
Collection of plant material,
preparation of plant extracts and determination of their chemical
composition
The plant material was collected from the Northern
Peloponnese (Patras) in Greece. Specifically, the grape stems used
in the present study were collected manually from three grape
varieties, namely Mavrodaphne, Muscat and Rhoditis. Subsequently,
the stem samples were chemically processed according to the
procedure that has been previously described (19). In brief, the samples underwent
repeated extractions in methanol and following liquid
chromatography analysis, the respective extracts were generated and
used for the analyses described herein. The stem extract derived
from the Mavrodaphne variety is rich in gallic acid and caffeic
acid, hydroxybenzoic and hydroxycinnamic acids, respectively,
whereas it contains higher amounts of the flavonol, quercetin, and
quercitrin, a flavonol glycoside, compared to the other two
extracts. The extract generated from the Muscat variety contains
higher concentrations of gallocatechin, a flavanol, polydatin,
which is a stilbene, and hesperidin, a flavanon glycoside, in
comparison to the extracts derived from the varieties Mavrodaphne
and Rhoditis. Finally, the chemical compounds detected in the
extract of the Rhoditis variety in higher amounts compared to the
Mavrodaphne and Muscat varieties are two members of
proanthocyanidins (porcyanidin B1 and B2), the flavanols, catechin
and epicatechin, two hydroxybenzoic acids (i.e., 2,5
dihydroxybenzoic acid, ellagic acid), the flavonol, rutin,
quercitrin-3-b-glucoside, which is a flavonol glycoside, and
trans-resveratrol, a well-known stilbene. The exact chemical
composition of the extracts was determined by high-performance
liquid chromatography (HPLC) and HPLC-electrospray ionization/mass
spectrometry analysis and this has been published in a previous
study (19).
Cell culture conditions
The cervical cancer cell line, HeLa, and the liver
cancer cell line, HepG2, were cultured in Dulbecco's modified
Eagle's medium (DMEM) containing 10% (v/v) fetal bovine serum
(FBS), 2 mM L-glutamine, 100 U/ml penicillin and 100 U/ml
streptomycin (all from Gibco; Thermo Fisher Scientific, Inc.) at
37˚C in 5% CO2. The cell lines used in this experiment,
in accordance with the international guidelines for good cell
culture practice, were examined for mycoplasma using PCR and were
found to be mycoplasma-free (29,30).
The working concentrations of the tested extracts were not found to
be cytotoxic in any cell line (Table
I). Furthermore, a morphological analysis, both at high and low
culture densities using a microscope, was conducted to authenticate
the state of the cells, through their phenotypic characteristics
(data not shown). Finally, the passage number for each cell line
did not exceed 30 population doublings.
 | Table ICytotoxic concentrations of the test
extracts that induced 25% cell death. |
Table I
Cytotoxic concentrations of the test
extracts that induced 25% cell death.
| Cell line | Wine extracts | Cytotoxic
concentration (µg/ml) |
|---|
| HepG2 | Muscat | 25 |
| | Mavrodaphne | 25 |
| | Rhoditis | 50 |
| HeLa | Muscat | 25 |
| | Mavrodaphne | 25 |
| | Rhoditis | 50 |
Cell viability assay
Cell viability was assessed using the XTT assay kit
(Roche Diagnostics GmbH). Briefly, 1x104 HeLa cells/well
were cultured in a 96-well plate in DMEM. Following a 24-h
incubation (37˚C), a wide range of the grape stem
extract concentrations (as shown in the figures) diluted in a
serum-free DMEM were used to treat the cells for 24 h.
Subsequently, 50 µl XTT reagent (50:1) were added to each well.
Following 4 h of incubation (37˚C), the absorbance was monitored at
450 and at 630 nm as a reference wavelength using a BioTek ELx800
microplate reader (BioTek Instruments Inc.). As a negative control,
samples containing serum-free DMEM only were used. In addition, the
absorbance of the extracts alone in serum-free DMEM and XTT test
solution was measured at 450 and 630 nm. The absorbance values of
the extracts alone were subtracted from those derived from the
absorbance of the cells treated with the test compounds. Data were
calculated as the percentage of viability using the following
equation:
Viability(%)=[(ODcontrol-ODsample)/ODcontrol]
x100, where ODcontrol and ODsample indicate
the OD of the negative control and the test compounds,
respectively. The experimental protocols were conducted in
triplicate (three repetitions), at three independent
experiments.
Measurement of endogenous GSH levels
in HeLa and HepG2 cell lines using flow cytometry
The intracellular GSH levels were assessed using the
fluorescent dye, mercury orange (Merck KGaA). The cells were
incubated (37˚C) for 24 h in FBS-free media including the test
extracts. The cells were then used for the measurement of GSH
levels using flow cytometry. According to the experimental
procedure, a 400 µM stock solution of the dye was prepared in
dimethyl sulfoxide (DMSO). The cells were trypsinized and
centrifuged (1,200 x g, 5 min, 4˚C). Subsequently, the cell pellet
was resuspended in phosphate-buffered saline (PBS) at the
concentration of 2x105 cells/ml and incubated in the
presence of mercury orange (40 µΜ) in 37˚C for 30 min. Following
centrifugation (1,200 x g, 5 min, 4˚C), the supernatant was
removed, and the pellet was resuspended in 350 µl PBS. The cells
were then subjected to flow cytometric analysis using a FACSCalibur
flow cytometer (BD Biosciences) with excitation and emission
wavelengths at 488 and 580 nm. The analysis was performed on 10,000
cells per sample, and the fluorescence intensities were measured on
a logarithmic scale. Data were analyzed using BD Cell Quest
software (BD Biosciences, version 6). Each experiment was repeated
at least three times.
Measurement of endogenous ROS levels
in HeLa and HepG2 cell lines using flow cytometry
The intracellular ROS levels were assessed using
2,7-dichlorofluorescein diacetate (DCF-DA), which is deacetylated
within cells by esterases and is further converted into fluorescent
DCF by the oxidative action of ROS. The cells were incubated (37˚C)
for 24 h in FBS-free media including the test extracts. The cells
were then used for the measurement of ROS using flow cytometry. A
20 mM stock solution of DCF-DA (Merck KGaA) was prepared in DMSO
and a 10 µM solution of DCF (Merck KGaA) was added in each well and
the cells were incubated at 37˚C for 45 min. The cells were then
trypsinized and centrifuged (1,200 x g, 5 min, 4˚C). The
supernatant was removed, and the pellet was resuspended in 350 µl
PBS. The cells were then subjected to flow cytometric analysis
using a FACSCalibur flow cytometer (BD Biosciences) with excitation
and emission wavelengths at 488 and 530 nm. The analysis was
performed on 10,000 cells per sample, and the fluorescence
intensities were measured on a logarithmic scale. Data were
analyzed using BD Cell Quest software (BD Biosciences, version 6).
Each experiment was repeated at least three independent times.
Statistical analysis
Data were analyzed using one-way ANOVA followed by
Tukey's test for multiple pair wise comparisons and are presented
as the mean ± standard error of the mean (SEM). A value of
P<0.05 was considered to indicate a statistically significant
difference. For the analyses, the statistical package for social
sciences (version 21.0; IBM Corp.) was used.
Results
Effects of the extracts on cell
viability
The extracts induced 25% cell death in the
concentrations shown in Table I.
Concentrations below these levels were selected for further
analysis to determine their effects on the GSH and ROS levels in
the human cancer cell lines.
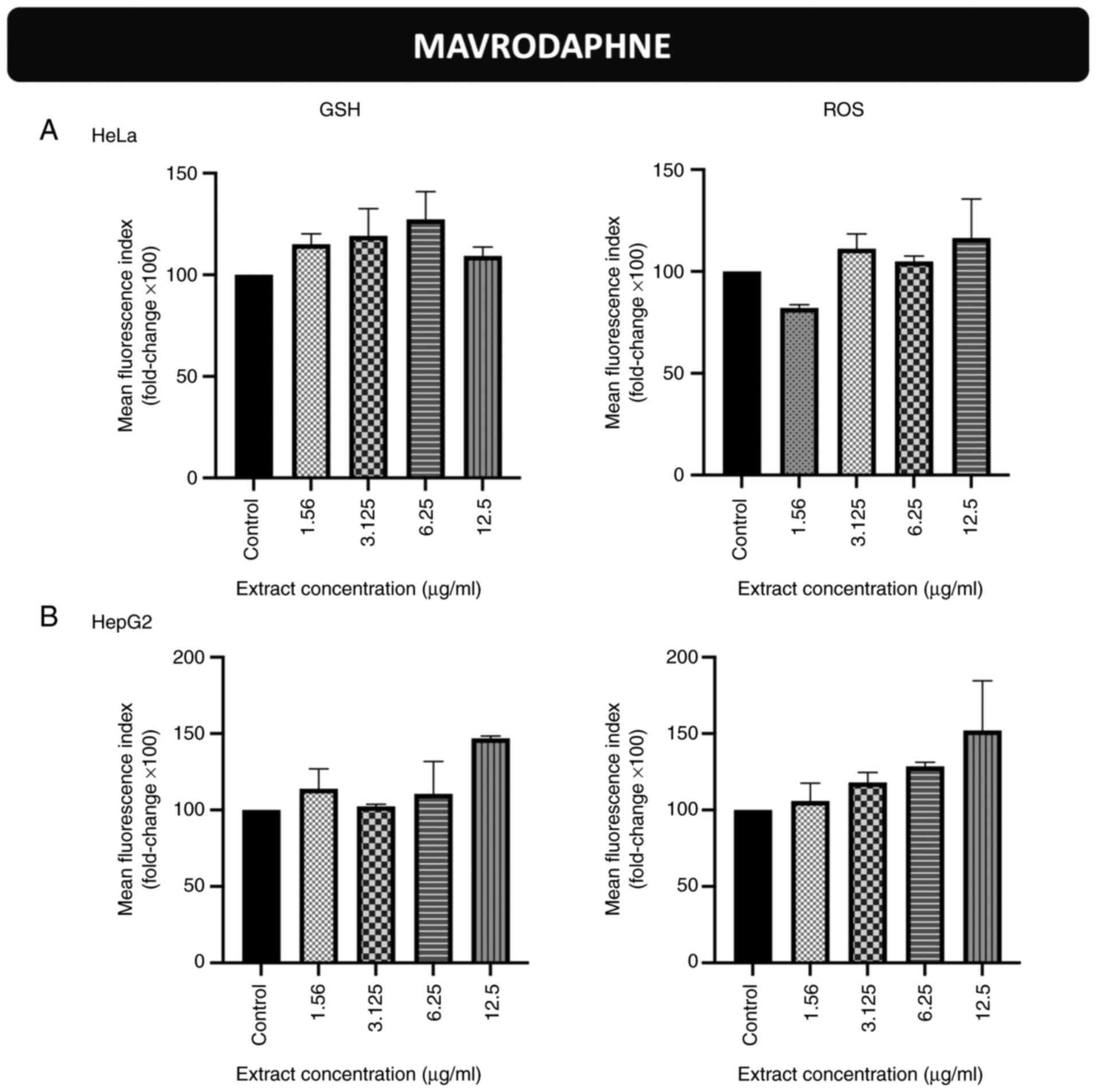
Effects of incubation of the cells
with the stem extract from the Mavrodaphne grape variety on
endogenous GSH and ROS levels in HeLa and HepG2 cells
The GSH and ROS levels in HeLa (Figs. 1A, S1 and S2) and HepG2 (Figs. 1B, S3 and S4) cells were not significantly altered
following incubation with the grape stem extract derived from the
Mavrodaphne variety.
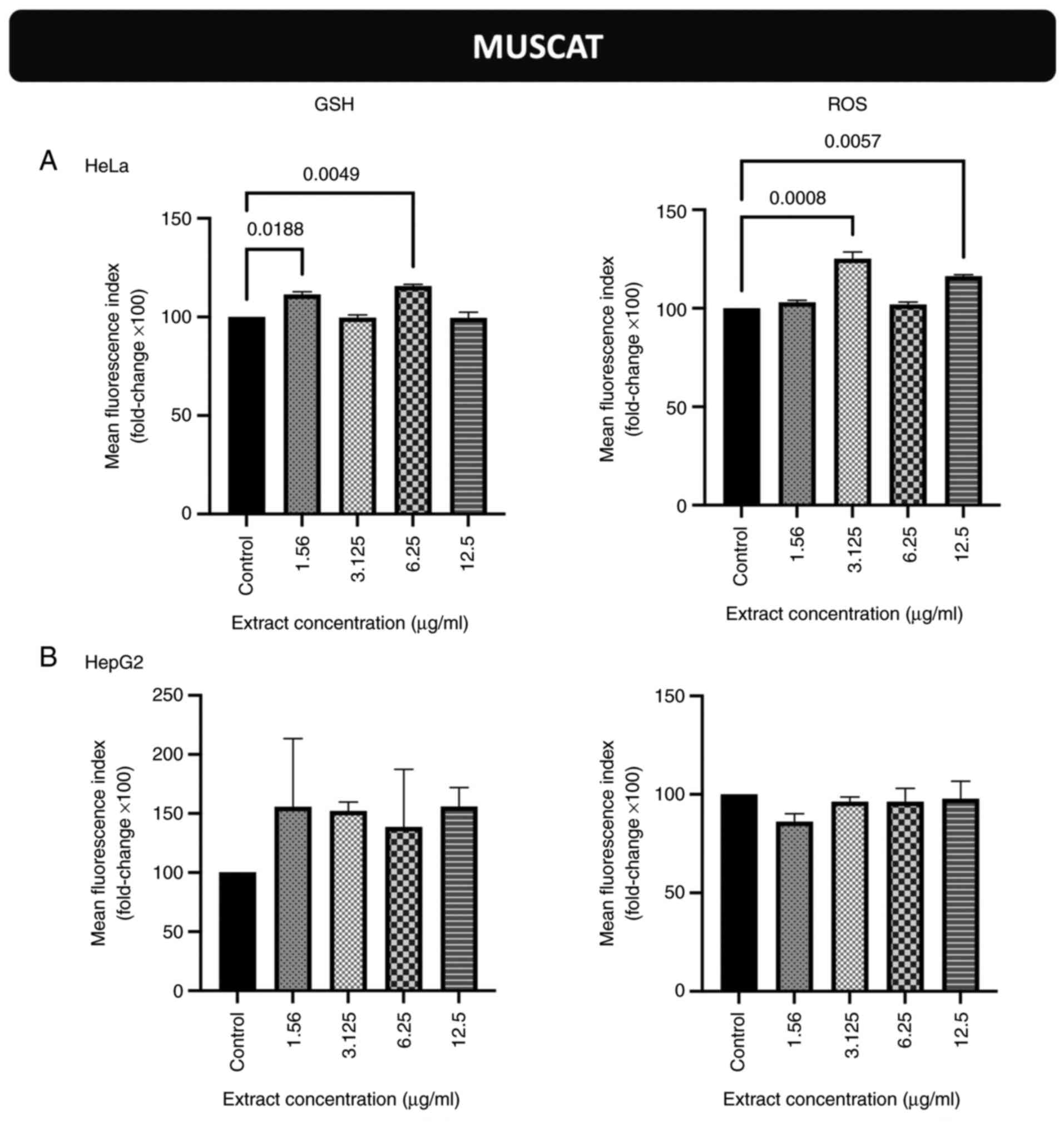
Effects of incubation of the cells
with the stem extract from the Muscat grape variety on endogenous
GSH and ROS levels in HeLa and HepG2 cells
The GSH levels in HeLa cells were increased compared
to those in the untreated cells (control) following incubation with
the extract concentrations equal to 1.56 and 6.25 µg/ml (Figs. 2A and S5). The ROS levels in HeLa cells were
also increased compared to those in the control cells following
incubation with the extract concentrations equal to 3.125 and 12.5
µg/ml (Figs. 2A and S6). As regards the HepG2 cells, the GSH
and ROS levels were not significantly altered (Figs. 2B, S7 and S8).
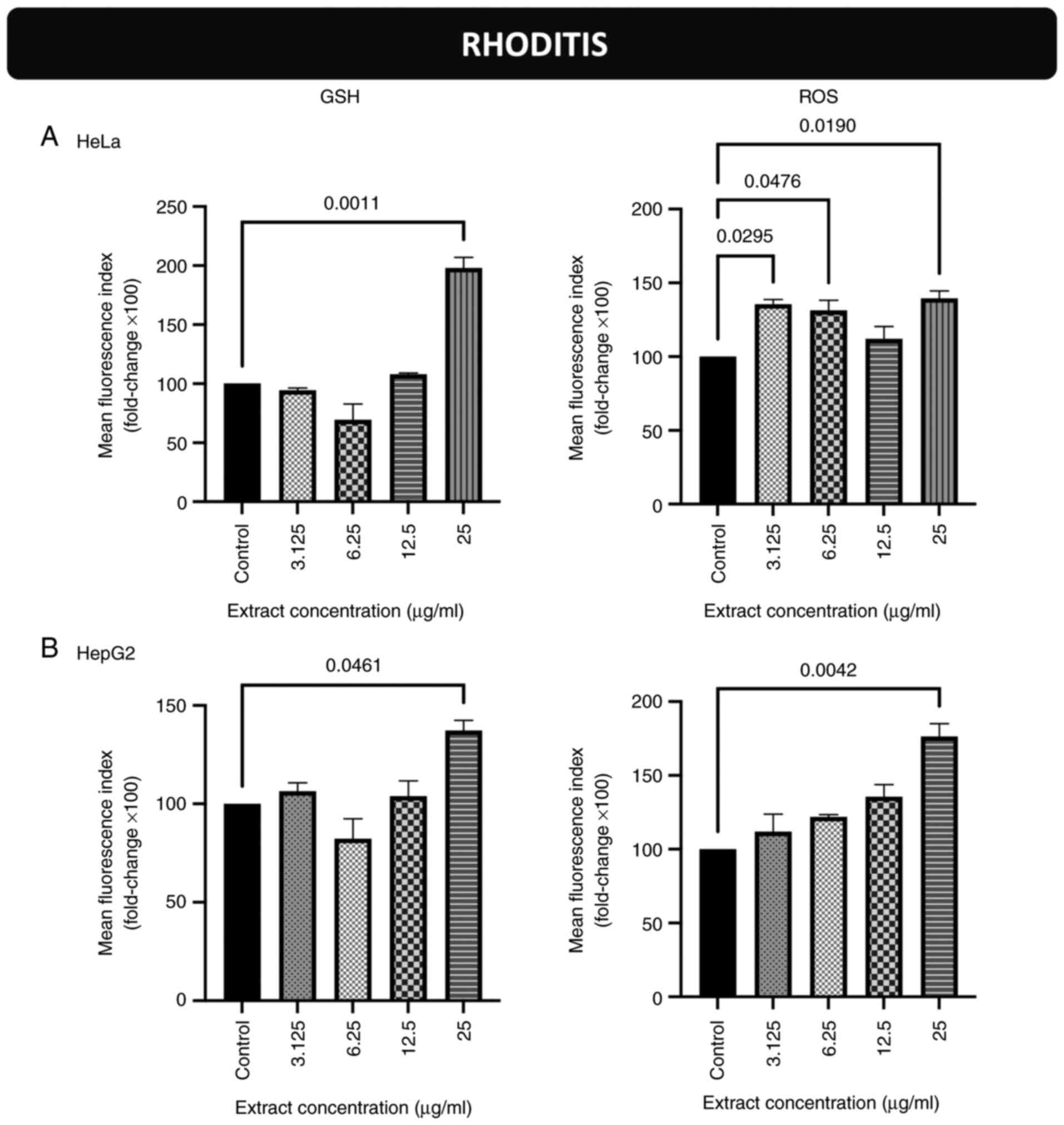
Effects of incubation of the cells
with the stem extract from the Rhoditis grape variety on endogenous
GSH and ROS levels in HeLa and HepG2 cells
The GSH levels of the HeLa cells were increased
compared to those of the control cells following incubation with
extract concentrations equal to 25 µg/ml (Figs. 3A and S9). The ROS levels in HeLa cells were
also increased compared to the controls following incubation with
extract concentrations equal to 3.125, 6.25 and 25 µg/ml (Figs. 3A and S10). As regards the HepG2 cells, the
GSH and ROS levels were increased compared to those in the control
cells following incubation with extract concentrations equal to 25
µg/ml for both biomarkers (Figs.
3B, S11 and S12).
Discussion
The present study examined the effects of three
grape stem extracts derived from Mavrodaphne, Muscat and Rhoditis,
which are native Greek vine varieties, on the redox status of
cancer cells. In particular, the endogenous levels of GSH and ROS
were measured in two human cancer cell lines (HeLa and HepG2)
following incubation with the extracts. Overall, the extract
derived from the grapes of the variety Rhoditis exerted prominent
prooxidant effects on both cell lines, as indicated by the
increased levels of ROS. The extract from Muscat only exerted
prooxidant effects on HeLa cells, whereas the examined parameters
remained unaltered following incubation of both cell lines with the
extract from Mavrodaphne. Thus, it is evident that the stem
extracts derived from Rhoditis and Muscat appear to be promising
compounds against cancer progression in the specific cellular
environment.
Grape stems are byproducts of the winemaking
procedure and studies have demonstrated that, due to their high
polyphenolic content, they could potentially be considered as
anticancer agents (31,32), whereas their antioxidant and
antimutagenic properties have also been observed (33). Concomitantly, research evidence has
demonstrated the prooxidant role of grape stems in the cellular
level by increasing ROS levels in epithelial cells (34). Although the literature regarding
stems is limited, there is sufficient evidence to indicate the
prooxidant role of polyphenols in cancer cell lines. Numerous in
vitro and in vivo studies have provided evidence towards
the direction of adopting polyphenolic schemes for cancer
prevention (35). For instance, it
has been demonstrated that gallic acid exerts pro-oxidant effects
on prostate cancer cells (36,37).
Moreover, other studies have reported that quercetin can function
either as an antioxidant or as a prooxidant depending on the cancer
cell line (38-40).
The anticancer properties of polyphenols have been attributed to
their potent antioxidant activity (41,42).
These results are in line with the findings presented herein.
Indeed, the present study demonstrated that the extracts derived
from the grape varieties Rhoditis and Muscat increased the levels
of both ROS and GSH. Thus, it is suggested that these extracts
function as prooxidant agents in the tested cancer cell lines,
whereas GSH appears to be activated by the cells in order to
overcome the potential oxidative modifications that are induced by
the increase in ROS levels.
It has been found that increased levels of ROS
activate the nuclear erythroid 2-related factor 2-antioxidant
response element signaling pathway that regulates a plethora of
genes whose products play crucial roles in the reinforcement of the
antioxidant arsenal of cells, ensuring their survival (43). This may also be associated with the
apoptosis of cancer cells and thus, with the prevention against
cancer. In the apoptotic process, the role of GSH is controversial
and is dependent on cell types and pro-apoptotic stimuli. Low
intracellular GSH levels have been found to prevent apoptosis by
compromising caspase activation in mouse hepatocytes (44). In addition, the depletion of
intracellular GSH appears to prevent CD95-triggered apoptosis
upstream of caspase-8 activation in T- and B-cells (45). Furthermore, cells undergoing
apoptosis also appear to export GSH into the extracellular space
(46,47). The production of ROS plays crucial
role in the proapoptotic effects of polyphenols against cancer cell
lines (48). ROS mediate the
release of cytochrome c from the mitochondria, which in turn
leads to caspase activation and apoptosis (49). Therefore, the increased levels of
GSH accompanied or induced by the elevated ROS levels observed in
the present study may contribute to the apoptosis of cancer cells.
Of note, the capacity of the test extracts to modulate the cell
redox status does not correspond to their content in polyphenolic
compounds. The stem extract from the variety Mavrodaphne has a
higher total polyphenolic content in comparison to the two other
extracts, as has been previously reported (19). However, it is the only extract that
did not affect the redox status of the cells. This finding may be
associated with the different capacities of the individual
phenolics to penetrate the cell membrane, based on their polarity
and size (50,51).
The present study demonstrated that grape stem
extracts rich in polyphenolic compounds with potent antioxidant
properties, do not appear to exert potent effects on the redox
status of cancer cells. As aforementioned, polyphenols can function
as prooxidants due to their antioxidant content (52,53).
This is an interesting biological aspect of polyphenols, and it
could be capitalized by researchers in the fight against cancer.
Previous studies have demonstrated that antioxidants are
detrimental against specific cancer types in animal models
(54,55). The proposed mechanism is related to
the fact that antioxidants scavenge free radicals, which are
usually harmful to cancer cells. Indeed, free radicals can kill
cancer cells as they have very weak antioxidant mechanisms compared
to healthy cells (56). Therefore,
free radicals appear to be ‘allies’ with anticancer therapies. The
use of cancer cell lines is valuable to provide a repeatable source
of biological activity estimation for experimental purposes.
Therefore, the establishment of a novel cell line or normal primary
cells would be more suitable, although a very complex process that
is still not well understood, a limitation of the present study. In
conclusion, the role of the tested extracts appears to be
promising; however, this is a research topic that warrants further
investigation.
Supplementary Material
Scatter plots and histograms from flow
cytometric analysis for the determination of GSH levels in the HeLa
cell line treated with grape stem extract derived from the
Mavrodaphne variety. GSH, glutathione.
Scatter plots and histograms from flow
cytometric analysis for the determination of ROS levels in the HeLa
cell line treated with grape stem extract derived from the
Mavrodaphne variety. ROS, reactive oxygen species.
Scatter plots and histograms from flow
cytometric analysis for the determination of GSH levels in the
HepG2 cell line treated with grape stem extract derived from the
Mavrodaphne variety. GSH, glutathione.
Scatter plots and histograms from flow
cytometric analysis for the determination of ROS levels in the
HepG2 cell line treated with grape stem extract derived from the
Mavrodaphne variety. ROS, reactive oxygen species.
Scatter plots and histograms from flow
cytometric analysis for the determination of GSH levels in the HeLa
cell line treated with grape stem extract derived from the Muscat
variety. GSH, glutathione.
Scatter plots and histograms from flow
cytometric analysis for the determination of ROS levels in the HeLa
cell line treated with grape stem extract derived from the Muscat
variety. ROS, reactive oxygen species.
Scatter plots and histograms from flow
cytometric analysis for the determination of GSH levels in the
HepG2 cell line treated with grape stem extract derived from the
Muscat variety. GSH, glutathione.
Scatter plots and histograms from flow
cytometric analysis for the determination of ROS levels in the
HepG2 cell line treated with grape stem extract derived from the
Muscat variety. ROS, reactive oxygen species.
Scatter plots and histograms from flow
cytometric analysis for the determination of GSH levels in the HeLa
cell line treated with grape stem extract derived from the Rhoditis
variety. GSH, glutathione.
Scatter plots and histograms from flow
cytometric analysis for the determination of ROS levels in the HeLa
cell line treated with grape stem extract derived from the Rhoditis
variety. ROS, reactive oxygen species.
Scatter plots and histograms from flow
cytometric analysis for the determination of GSH levels in the
HepG2 cell line treated with grape stem extract derived from the
Rhoditis variety. GSH, glutathione.
Scatter plots and histograms from flow
cytometric analysis for the determination of ROS levels in the
HepG2 cell line treated with grape stem extract derived from the
Rhoditis variety. ROS, reactive oxygen species.
Acknowledgements
Not applicable.
Funding
Funding: The present study was funded by the Action ‘RESEARCH-
CREATE-INNOVATE’ supported by the Operational Program
Competitiveness, Entrepreneurship and Innovation 2014-2020 (EPAnEK)
and the European Union (Τ1ΕΔΚ-01889).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
EV was involved in data curation, methodology and in
the writing of the original draft. ASV was involved in the
conceptualization of the study, as well as in data curation, in the
writing of the original draft, and in the writing, review and
editing of the manuscript. FT was involved in data curation, and in
the writing, review and editing of the manuscript. ZS and KP were
involved in the conceptualization of the study. SH was involved in
the conceptualization and methodology of the study. DK was involved
in the conceptualization of the study, as well as in funding
acquisition, project administration, study supervision, and in the
writing, review and editing of the manuscript. All authors
contributed to the interpretation of the data. All authors have
read and agreed to the published version of the manuscript. EV and
DK confirm the authenticity of all the raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
SH and DK are Editors of the journal, but had no
personal involvement in the reviewing process, or any influence in
terms of adjudicating on the final decision, for this article. The
other authors declare that they have no competing interests.
References
|
1
|
Yadav M, Jain S, Bhardwaj A, Nagpal R,
Puniya M, Tomar R, Singh V, Parkash O, Prasad GB, Marotta F and
Yadav H: Biological and medicinal properties of grapes and their
bioactive constituents: An update. J Med Food. 12:473–484.
2009.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Pandey KB and Rizvi SI: Role of red grape
polyphenols as antidiabetic agents. Integr Med Res. 3:119–125.
2014.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Renaud S and de Lorgeril M: Wine, alcohol,
platelets, and the French paradox for coronary heart disease.
Lancet. 339:1523–1526. 1992.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Vogt T: Phenylpropanoid biosynthesis. Mol
Plant. 3:2–20. 2010.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Graf BA, Milbury PE and Blumberg JB:
Flavonols, flavones, flavanones, and human health: Epidemiological
evidence. J Med Food. 8:281–290. 2005.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Fresco P, Borges F, Marques MPM and Diniz
C: The anticancer properties of dietary polyphenols and its
relation with apoptosis. Curr Pharm Des. 16:114–134.
2010.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Hendric AB: Flavonoid-membrane
interactions: Possible consequences for biological effects of some
polyphenolic compounds. Acta Pharmacol Sin. 27:27–40.
2006.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Ali K, Maltese F, Choi YH and Verpoorte R:
Metabolic constituents of grapevine and grape-derived products.
Phytochem Rev. 9:357–378. 2010.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Riedel H, Thaw Saw NMM, Akumo DN, Kütük O
and Smetanska I: Wine as food and medicine. In: Scientific, Health
and Social Aspects of the Food Industry. Valdez B (ed.) IntechOpen:
399-418, 2012.
|
|
10
|
Alonso AM, Guillén DA, Barroso CG, Puertas
B and García A: Determination of antioxidant activity of wine
byproducts and its correlation with polyphenolic content. J Agric
Food Chem. 50:5832–5836. 2002.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Somers TC and Vérette E: Phenolic
composition of natural wine types. In: Wine analysis. Linskens HF
and Jackson JF (eds.). 1st edition. Springer, Berlin, Heidelberg,
pp219-257, 1988.
|
|
12
|
Ruggieri L, Cadena E, Martínez-Blanco J,
Gasol CM, Rieradevall J, Gabarrell X, Gea T, Sort X and Sánchez A:
Recovery of organic wastes in the Spanish wine industry. Technical,
economic and environmental analyses of the composting process. J
Clean Prod. 17:830–838. 2009.
|
|
13
|
Torres JL, Varela B, García MT, Carilla J,
Matito C, Centelles JJ, Cascante M, Sort X and Bobet R:
Valorization of grape (Vitis vinifera) byproducts. Antioxidant and
biological properties of polyphenolic fractions differing in
procyanidin composition and flavonol content. J Agric Food Chem.
50:7548–7555. 2002.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Villaescusa I, Fiol N, Martínez M,
Miralles N, Poch J and Serarols J: Removal of copper and nickel
ions from aqueous solutions by grape stalks wastes. Water Res.
38:992–1002. 2004.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Kosińska-Cagnazzo A, Heeger A, Udrisard I,
Mathieu M, Bach B and Andlauer W: Phenolic compounds of grape stems
and their capacity to precipitate proteins from model wine. J Food
Sci Technol. 57:435–443. 2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
luga M and Mironeasa S: Potential of grape
byproducts as functional ingredients in baked goods and pasta.
Compr Rev Food Sci Food Saf. 19:2473–2505. 2020.PubMed/NCBI View Article : Google Scholar
|
|
17
|
García Lomillo J and González-SanJosé ML:
Applications of wine pomace in the food industry: Approaches and
functions. Compr Rev Food Sci Food Saf. 16:3–22. 2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Barros A, Gironés-Vilaplana A and Texeira
A: Grape stems as a source of bioactive compounds: Application
towards added-value commodities and significance for human health.
Phytochem Rev. 14:921–931. 2015.
|
|
19
|
Veskoukis AS, Vassi E, Poulas K,
Kokkinakis M, Asprodini E, Haroutounian S and Kouretas D: Grape
stem extracts from three native greek vine varieties exhibit strong
antioxidant and antimutagenic properties. Anticancer Res.
40:2025–2032. 2020.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Kerasioti E, Terzopoulou Z, Komini O,
Kafantaris I, Makri S, Stagos D, Gerasopoulos K, Anisimov NY,
Tsatsakis AM and Kouretas D: Tissue specific effects of feeds
supplemented with grape pomace or olive oil mill wastewater on
detoxification enzymes in sheep. Toxicol Rep. 4:364–372.
2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Kafantaris I, Stagos D, Kotsampasi B,
Hatzis A, Kypriotakis A, Gerasopoulos K, Makri S, Goutzourelas N,
Mitsagga C, Giavasis I, et al: Grape pomace improves performance,
antioxidant status, fecal microbiota and meat quality of piglets.
Animal. 12:246–255. 2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Veskoukis AS, Kyparos A, Nikolaidis MG,
Stagos D, Aligiannis N, Halabalaki M, Chronis K, Goutzourelas N,
Skaltsounis L and Kouretas D: The antioxidant effects of a
polyphenol-rich grape pomace extract in vitro do not correspond in
vivo using exercise as an oxidant stimulus. Oxid Med Cell Longev.
2012(185867)2012.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Spanou C, Veskoukis AS, Stagos D, Liadaki
K, Anastasiadi M, Haroutounian SA, Tsouka M, Tzanakouli E and
Kouretas D: Effects of grape extracts on the in vitro activity of
enzymes involved in oxidative stress regulation. In Vivo.
25:657–662. 2011.PubMed/NCBI
|
|
24
|
Goutzourelas N, Stagos D, Spanidis Y,
Liosi M, Apostolou A, Priftis A, Haroutounian S, Spandidos DA,
Tsatsakis AM and Kouretas D: Polyphenolic composition of grape stem
extracts affects antioxidant activity in endothelial and muscle
cells. Mol Med Rep. 12:5846–5856. 2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Anastasiadi M, Pratsinis H and Kletsas D:
Grape stem extracts: Polyphenolic content and assessment of their
in vitro antioxidant properties. LWT-Food Sci Technol. 48:316–322.
2012.
|
|
26
|
León-González AJ, Auger C and Schini-Kerth
VB: Pro-oxidant activity of polyphenols and its implication on
cancer chemoprevention and chemotherapy. Biochem Pharmacol.
98:371–380. 2015.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Eghbaliferiz S and Iranshahi M: Prooxidant
activity of polyphenols, flavonoids, anthocyanins and carotenoids:
Updated review of mechanisms and catalyzing metals. Phytother Res.
30:1379–1391. 2016.PubMed/NCBI View
Article : Google Scholar
|
|
28
|
Veskoukis AS, Kerasioti E, Priftis A,
Kouka P, Spanidis Y, Makri S and Kouretas D: A battery of
translational biomarkers for the assessment of the in vitro and in
vivo antioxidant action of plant polyphenolic compounds: The
biomarker issue. Curr Opin Toxicol. 13:99–109. 2019.
|
|
29
|
Bal-Price A and Coecke S: Guidance on good
cell culture practice (GCCP). Neuromethods. 56:1–25. 2011.
|
|
30
|
Kouka P, Tekos F, Valta K, Mavros P,
Veskoukis AS, Angelis A, Skaltsounis AL and Kouretas D: Οlive tree
blossom polyphenolic extracts exert antioxidant and antimutagenic
activities in vitro and in various cell lines. Oncol Rep.
42:2814–2825. 2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Pasini F, Chinnici F, Caboni MF and
Verardo V: Recovery of oligomeric proanthocyanidins and other
phenolic compounds with established bioactivity from grape seed
by-products. Molecules. 24(677)2019.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Ferri M, Bin S, Vallini V, Fava F,
Michelini E, Roda A, Minnucci G, Bucchi G and Tassoni A: Recovery
of polyphenols from red grape pomace and assessment of their
antioxidant and anti-cholesterol activities. N Biotechnol.
33:338–344. 2016.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Apostolou A, Stagos D, Galitsiou E, Spyrou
A, Haroutounian S, Portesis N, Trizoglou I, Wallace Hayes A,
Tsatsakis AM and Kouretas D: Assessment of polyphenolic content,
antioxidant activity, protection against ROS-induced DNA damage and
anticancer activity of Vitis vinifera stem extracts. Food Chem
Toxicol. 61:60–68. 2013.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Quero J, Jiménez-Moreno N, Esparza I,
Osada J, Cerrada E, Ancín-Azpilicueta C and Rodríguez-Yoldi MJ:
Grape stem extracts with potential anticancer and antioxidant
properties. Antioxidants (Basel). 10(243)2021.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Niedzwiecki A, Roomi MW, Kalinovsky T and
Rath M: Anticancer efficacy of polyphenols and their combinations.
Nutrients. 8(552)2016.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Chen HM, Wu YC, Chang FR, Hsu HK, Hsieh
YC, Chen CC and Yuan SS: Gallic acid, a major component of Toona
sinensis leaf extracts, contains a ROS mediated anticancer activity
in human prostate cancer cells. Cancer Lett. 286:161–171.
2009.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Russell LH Jr, Mazzio E, Badisa RB, Zhu
ZP, Agharahimi M, Oriaku ET and Goodman CB: Autoxidation of gallic
acid induces ROS-dependent death in human prostate cancer LNCaP
cells. Anticancer Res. 32:1595–1602. 2012.PubMed/NCBI
|
|
38
|
Ward AB, Mir H, Kapur N, Gales DN,
Carriere PP and Singh S: Quercetin inhibits prostate cancer by
attenuating cell survival and inhibiting anti-apoptotic pathways.
World J Surg Oncol. 16(108)2018.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Costea T, Nagy P, Ganea C, Szöllősi J and
Mocanu MM: Molecular mechanisms and bioavailability of polyphenols
in prostate cancer. Int J Mol Sci. 20(1062)2019.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Hashemzaei M, Delarami Far A, Yari A,
Heravi RE, Tabrizian K, Taghdisi SM, Sadegh SE, Tsarouhas K,
Kouretas D, Tzanakakis G, et al: Anticancer and apoptosis inducing
effects of quercetin in vitro and in vivo. Oncol Rep.
38:819–828. 2017.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Li F, Li S, Li HB, Deng GF, Ling WH, Wu S,
Xu XR and Chen F: Antiproliferative activity of peels, pulps and
seeds of 61 fruits. J Funct Foods. 5:1298–1309. 2013.
|
|
42
|
Li F, Li S, Li HB, Deng GF, Ling WH and Xu
XR: Antiproliferative activities of tea and herbal infusions. Food
Funct. 4:530–538. 2013.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Shaw P and Chattopadhyay A: Nrf2-ARE
signaling in cellular protection: Mechanism of action and the
regulatory mechanisms. J Cell Physiol. 235:3119–3130.
2020.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Hentze H, Künstle G, Volbracht C, Ertel W
and Wendel A: CD95-Mediated murine hepatic apoptosis requires an
intact glutathione status. Hepatology. 30:177–185. 1999.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Hentze H, Schmitz I, Latta M, Krueger A,
Krammer PH and Wendel A: Glutathione dependence of caspase-8
activation at the death-inducing signaling complex. J Biol Chem.
277:5588–5595. 2002.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Ghibelli L, Fanelli C, Rotilio G, Lafavia
E, Coppola S, Colussi C, Civitareale P and Ciriolo MR: Rescue of
cells from apoptosis by inhibition of active GSH extrusion. FASEB
J. 12:479–486. 1998.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Pullar JM and Hampton MB:
Diphenyleneiodonium triggers the efflux of glutathione from
cultured cells. J Biol Chem. 277:19402–19407. 2002.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Morales P and Haza AI: Selective apoptotic
effects of piceatannol and myricetin in human cancer cells. J Appl
Toxicol. 32:986–993. 2012.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Udenigwe CC, Ramprasath VR, Aluko RE and
Jones PJ: Potential of resveratrol in anticancer and
anti-inflammatory therapy. Nutr Rev. 66:445–454. 2008.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Ramos-Escudero F, Muñoz AM, Alvarado-Ortíz
C, Alvarado Á and Yáñez JA: Purple corn (Zea mays L.) phenolic
compounds profile and its assessment as an agent against oxidative
stress in isolated mouse organs. J Med Food. 15:206–215.
2012.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Sarega N, Imam MU, Ooi DJ, Chan KW, Md Esa
N, Zawawi N and Ismail M: Phenolic rich extract from clinacanthus
nutans attenuates hyperlipidemia-associated oxidative stress in
rats. Oxid Med Cell Longev. 2016(4137908)2016.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Galadari S, Rahman A, Pallichankandy S and
Thayyullathil F: Reactive oxygen species and cancer paradox: To
promote or to suppress? Free Radic Biol Med. 104:144–164.
2017.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Zou ZV, Le Gal K, El Zowalaty AE,
Pehlivanoglu LE, Garellick V, Gul N, Ibrahim MX, Bergh PO,
Henricsson M, Wiel C, et al: Antioxidants promote intestinal tumor
progression in mice. Antioxidants (Basel). 10(241)2021.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Le Gal K, Ibrahim MX, Wiel C, Sayin VI,
Akula MK, Karlsson C, Dalin MG, Akyürek LM, Lindahl P, Nilsson J
and Bergo MO: Antioxidants can increase melanoma metastasis in
mice. Sci Transl Med. 7(308re8)2015.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Sayin VI, Ibrahim MX, Larsson E, Nilsson
JA, Lindahl P and Bergo MO: Antioxidants accelerate lung cancer
progression in mice. Sci Transl Med. 6(221ra15)2014.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Glasauer A and Chandel NS: Targeting
antioxidants for cancer therapy. Biochem Pharmacol. 92:90–101.
2014.PubMed/NCBI View Article : Google Scholar
|