Introduction
Treatment of tissue defects and injury due to
congenital causes, trauma and infection has been one of the
challenges for plastic surgeons and trauma surgeons. The rapid
development of tissue-engineering techniques provides strategies
and potential sources of materials for the repair and regeneration
following tissue injury.
The possible use of mesenchymal stem cells (MSCs)
for tissue engineering therapies has been increasingly explored.
Ideally, MSCs should be autologous, available in sufficient
quantity, and easily obtained, with the ability to proliferate and
differentiate in vitro (1).
Studies have confirmed the multipotentiality of
adipose-derived stem cells (ADSCs) (2,3),
which have advantages as seed cells for tissue engineering. For
example, collection of tissue for ADSC isolation is relatively
simple as compared to other sources; human adipose tissue, a rich
source of ADSCs, can be obtained by suction-assisted lipectomy
(i.e., liposuction) (4,5). This technique is minimally invasive,
increasing the likelihood of patient acceptance. However, the
chondrogenic potential of ADSCs has come into question (6–8).
For example, the maturity and homogeneity of cartilage constructed
using ADSCs was lower as compared to that constructed using bone
marrow stem cells (BMSCs) (8).
This may be attributed to the low proportion of MSCs and the
presence of contaminant cells that are likely terminally
differentiated.
ADSCs, in fact adipose-derived cells (ADCs), consist
of a heterogeneous cell population, including fat precursor cells,
hematopoietic cells, endothelial cells, vascular pericytes, and
fibroblasts (9). Because some
ADCs have directed differentiation and terminally differentiated
cells have no chondrogenic potential, the maturity and homogeneity
of cartilage is affected. Therefore, separation of ADCs prior to
their application is necessary.
The traditional methods employed for separating stem
cells include density gradient centrifugation, differential
adhesion, and culturing in serum. However, because these methods
are largely nonspecific, the resulting cultures are often of low
purity. In addition, these methods are complex, and the
experimental period is relatively long. An alternative to the
aforementioned methods includes separating the stem cells using
stem cell-specific antigens with immunomagnetic beads or flow
cytometry. These methods require a short experimental period and
result in an increased purity in the stem cell cultures. However,
these techniques have yet to be applied to ADSCs due to the lack of
a specific surface marker.
To identify a possible ADSC-specific surface
antigen, ADCs were analyzed for the expression of commonly used
MSC-related antigens, including endoglin (CD105), CD166 (activated
leukocyte cell adhesion molecule, ALCAM) and STRO-1. In the present
study, CD105, a relatively specific antigen for MSCs, was applied
for the separation of ADSCs. CD105+ and
CD105− ADC growth, colony formation, and differentiation
potentials were assessed. These findings provide theoretical and
practical evidence for the separation and purification of
ADSCs.
Materials and methods
Isolation and culture of ADCs
Adipose tissues were collected from patients
receiving suction lipectomy in the Department of Plastic Surgery,
Ninth People’s Hospital, Shanghai Jiao Tong University School of
Medicine. Of the 5 patients, 4 were female; the mean age was 28.5
years (range, 22–45 years), and the lipectomy sites were abdomen
(n=2) and thigh (n=3).
The adipose tissues from five patients were
separately washed twice in PBS and then digested in 0.075%
collagenase NB4 (Serva, Heidelberg, Germany) for 1 h followed by
centrifugation at 1,380 rpm for 10 min. The cell sediments were
collected and resuspended in DMEM (Gibco, Grand Island, NY, USA)
supplemented with 10% fetal bovine serum (FBS; Hyclone Labs, Thermo
Scientific, Rockford, IL, USA). The cell density was adjusted to
4x105 cells/cm2, and the ADCs were seeded
into disks followed by culture and passaging. ADCs in the second
passage were used for subsequent experiments.
Identification of surface antigens on the
ADCs
Analysis of surface antigen expression was
undertaken in ADCs of the first and second passage using flow
cytometry. Cells were digested in 0.25% trypsin and 0.02% EDTA
(Sigma, St. Louis, MO, USA), collected, and isolated by
centrifugation at 1,500 rpm for 5 min. The cell density was
adjusted to 0.5x106 cells/ml, and 1 ml of the cell
suspension was added into a 1.5-ml Eppendorf tube followed by
centrifugation at 1,500 rpm for 5 min. After the supernatant was
removed, the ADCs were resuspended in 100 μl of 1% BSA after
which they were treated with the following primary antibodies
diluted 1:50 at 4°C: integrin β1/CD29 (sc8978; Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA), integrin α4/CD49d
(9431-09A; Southern Biotech, Birmingham, AL, USA), ALCAM/CD166
(MCA1926F; AbD Serotec), PECAM-1/CD31 (F-8402; Sigma),
hematopoietic/CD34 (M0824; Dako, Carpinteria, CA, USA), CD14
(T5647; Sigma), endoglin/CD105 (MCA1557PE; AbD Serotec, Raleigh,
NC, USA), VCAM-1/CD106 (sc13160) and VEGFR-2/FLK-1 (sc505) (Santa
Cruz Biotechnology, Inc.), and STRO-1 (MAB1038; R&D Systems,
Minneapolis, MN, USA). After 30 min, ADCs were separated by
centrifugation, washed in PBS, and treated with goat anti-mouse
IgM-FITC secondary antibody (AMI3608; Biosource, Carlsbad, CA, USA)
diluted 1:200 at 4°C for 30 min. After the cells were fixed in 10%
formalin, they were subjected to flow cytometry. Control groups
consisted of an isotype control group and blank control group. Each
experiment was performed 5 times and the averages were
obtained.
Separation of ADCs using anti-CD105
magnetic beads and MACS
ADCs in their second passage were digested in 0.25%
trypsin + 0.02% EDTA and centrifuged at 1,500 rpm for 5 min. The
cell density was adjusted to 0.5x106 cells/ml, and 10 ml
was added to a 15-ml centrifuge tube followed by centrifugation at
1,500 rpm for 5 min. The cells were resuspended in 1 ml of 1% BSA
and treated with CD105 conjugated to phycoerythrin (PE; AbD
Serotec, Oxford, UK) diluted 1:50 at 4°C for 30 min followed by
centrifugation at 1,500 rpm for 5 min. After washing in PBS, the
cells were centrifuged at 1,500 rpm for 5 min, and resuspended in 1
ml of 1% BSA, and incubated with anti-PE magnetic beads (Miltenyi
Biotec, Auburn, CA, USA) diluted 1:50 at 4°C for 30 min. After
centrifugation (1,500 rpm, 5 min), the cells were washed in PBS,
centrifuged, at 1,500 rpm for 5 min, and resuspended in 1 ml of 1%
BSA. The remaining cells were prepared for separation by MACS.
CD105-conjugated ADCs were separated using autoMACS
(Miltenyi Biotec). CD105+ and CD105− ADCs
were collected into aseptic 15-ml centrifuge tubes. The
double-positive and double-negative modes were employed to achieve
pure CD105+ and CD105− ADCs, respectively.
The isolated cells were seeded onto five 100-mm disks at a density
of 1x104 cells/cm2 for the following
experiments. Flow cytometry was used to measure the purities of
CD105+ and CD105− ADCs; 0.5x106
cells (CD105+, CD105−, or unseparated ADCs)
were suspended in 100 μl of 1% BSA and subjected to flow
cytometry using the methods described above.
Analysis of cell growth and colony
formation
CD105+ and CD105− ADC growth
and colony formation was determined by cell counting and Giemsa
staining (Shanghai Chemical Reagents Co., Ltd., Shanghai, China),
respectively. The growth kinetics of CD105+ ADCs were
compared with that of CD105− ADCs.
Analysis of ADC differentiation
potential
The differentiation potentials of the following cell
groups were determined: induced CD105+ ADCs, non-induced
CD105+ ADCs, induced CD105− ADCs, and
non-induced CD105− ADCs. Each experiment was
independently repeated 6 times.
For chondrogenetic induction cells
(2.5x105) were induced with 5 ml of chondrogenetic
induction medium containing 10 μg transformation growth
factor-β1 (TGF-β1) (R&D Systems), 50 μg insulin growth
factor-1 (IGF-1) (R&D Systems), and 2 mg/ml dexamethasone
(Sigma) followed by centrifugation at 1380 rpm for 5 min. The cell
pellets were maintained in the chondrogenetic induction medium for
14 and 21 days. In the non-induced groups, the medium was replaced
with DMEM alone.
For osteogenic induction the cells were digested and
seeded onto a 24-well plate at a density of 1x104
cells/well. For induction, cells were maintained in osteogenic
induction medium containing 10 nM vitamin D3 (Sigma) and 10 mM
β-phosphoglycerol (ICN Biomedicals, Solon, OH, USA) and 0.1
μM dexamethasone for 14 and 21 days, and those in the
non-induced groups were cultured in traditional DMEM.
For adipogenic induction the cells were digested and
seeded onto a 24-well plate at a density of 1x104
cells/well. When cell confluence reached 80%, cells in the induced
groups were maintained in the adipogenic induction medium
containing 0.5 mM 3-isobutyl-1-methylxanthine (IBMX), 200 μM
indomethacin, 10 μM insulin and 1 μM dexamethasone
(all from Sigma) for 14 and 21 days; those in the non-induced
groups were cultured in traditional DMEM.
Immunohistochemistry analysis
As previously described in Liu et al
(10), the cells were fixed in
10% formalin, embedded in paraffin, and sectioned into 5-μm
slices. The sections were stained with hematoxylin and eosin
(H&E) to evaluate histological structure. Analysis of collagen
II expression was performed by 1% BSA blocked sections using 1:100
mouse anti-human type II collagen monoclonal primary antibody
(Dako) at 4°C overnight, followed by 1:200 horseradish peroxidase
(HRP)-conjugated goat anti-mouse secondary antibody (Dako) at 37°C
for 30 min. DAB was used as a substrate for HRP.
Alizarn Red and Oil Red staining was performed
respectively to evaluate the formation of calcium deposits and
lipid droplet.
Reverse-transcriptase polymerase chain
reaction (RT-PCR) analysis
Total-RNA was extracted from the cells after
multi-lineage induction using TRIzol (Invitrogen, CA). cDNA was
obtained using previously described methods (11). RT-PCR was performed with different
primers respectively. β-actin mRNA expression was quantified as an
internal control. The primer sequences for each gene analyzed are
listed in Table I.
 | Table IReverse transcriptase-polymerase
chain reaction (RT-PCR) primers. |
Table I
Reverse transcriptase-polymerase
chain reaction (RT-PCR) primers.
| Gene (product
size) | Sense | Antisense |
|---|
| PPARγ (493 bp) |
5′-GATCCAGTGGTTGCAGATTA-3′ |
5′-GGTCAGCGGGAAGGACTTTA-3′ |
| COLII (510 bp) |
5′-TCCCCGGCACTCCTGGCACTGAT-3′ |
5′-CTTGGGCACCTCGGGCTCCTTTAG-3′ |
| AKP (467 bp) |
5′-CTGGTAGGCGATGTCCTTA-3′ |
5′-ACGTGGCTAAGAATGTCATC-3′ |
| Leptin (481
bp) |
5′-CAAGCTGTGCCCATCCAAAA-3′ |
5′-GCCAGAGTTCCTTCCCTTAA-3′ |
| β-actin (318
bp) |
5′-ATCATGTTTGAGACCTTCAA-3′ |
5′-CATCTCTTGCTCGAAGTCCA-3′ |
Statistical analysis
Cell growth is presented as mean ± standard error
(SE). Mean cell number over time were compared through repeated
ANOVA measurements. Two-sample t-test at each time-point was
performed to compare the differences between CD105− ADCs
and CD105+ ADCs. Colony formation rates are presented as
bar graphs, representing the mean ± SE and compared using
two-sample t-tests. Collagen II, alkaline phosphatase (AKP),
leptin, and PPARγ2 mRNA expression was presented as bar graphs,
representing mean ± SE. Data among four conditions
(CD105+ induced and non-induced and CD105−
induced and non-induced) were compared using one-way ANOVA with
Bonferroni adjustment. Data within-conditions (change from Day
14–21) were compared using paired t-tests. All statistical
assessments were two-tailed, and P<0.05 was considered
significant. Statistical analyses were performed using the SPSS
15.0 statistics software (SPSS Inc., Chicago, IL, USA).
Results
ADC surface antigen expression
To determine which surface antigen may be useful for
isolation of ADSCs from a mixed ADC population, ADCs in the first
and second passage were analyzed for various stem cell-associated
surface antigens. First passage ADCs were largely negative for
CD14, CD106, STRO-1, and Flk-1 but positive for CD29, CD31, CD34,
CD49d, CD105 and CD166 (Table
II). ADCs in the second passage had significantly decreased
CD34 expression and were negative for CD31 and CD49d but positive
for CD14, STRO-1 and Flk-1. In the second passage, the expressions
of CD14, CD105, CD166, STRO-1, and Flk-1 were markedly higher than
observed in the first passage (Table
II).
| Table IIStem-cell antigen expression in
ADCs. |
Table II
Stem-cell antigen expression in
ADCs.
| Antigen | First generation
(%) | Second generation
(%) | P-valuea |
|---|
| CD14 | 2.99±0.4 | 7.98±0.78 | 0.009 |
| CD29 | 82.03±3.4 | 64.26±2. 49 | 0.025 |
| CD31 | 15.12±1.24 | 2.69±0.5 | 0.001 |
| CD34 | 20.99±1.84 | 6.02±0.56 | 0.001 |
| CD49d | 14.8±0.84 | 1.29±0.23 | <0.001 |
| CD105 | 20.71±2.08 | 35.33±1.6 | 0.002 |
| CD106 | 4.49±0.34 | 0.42±0.09 | 0.001 |
| CD166 | 18.98±0.93 | 62.41±2.35 | <0.001 |
| Flk-1 | 2.24±0.21 | 9.42±0.87 | 0.001 |
| STRO-1 | 3.41±0.18 | 10.55±0.99 | 0.002 |
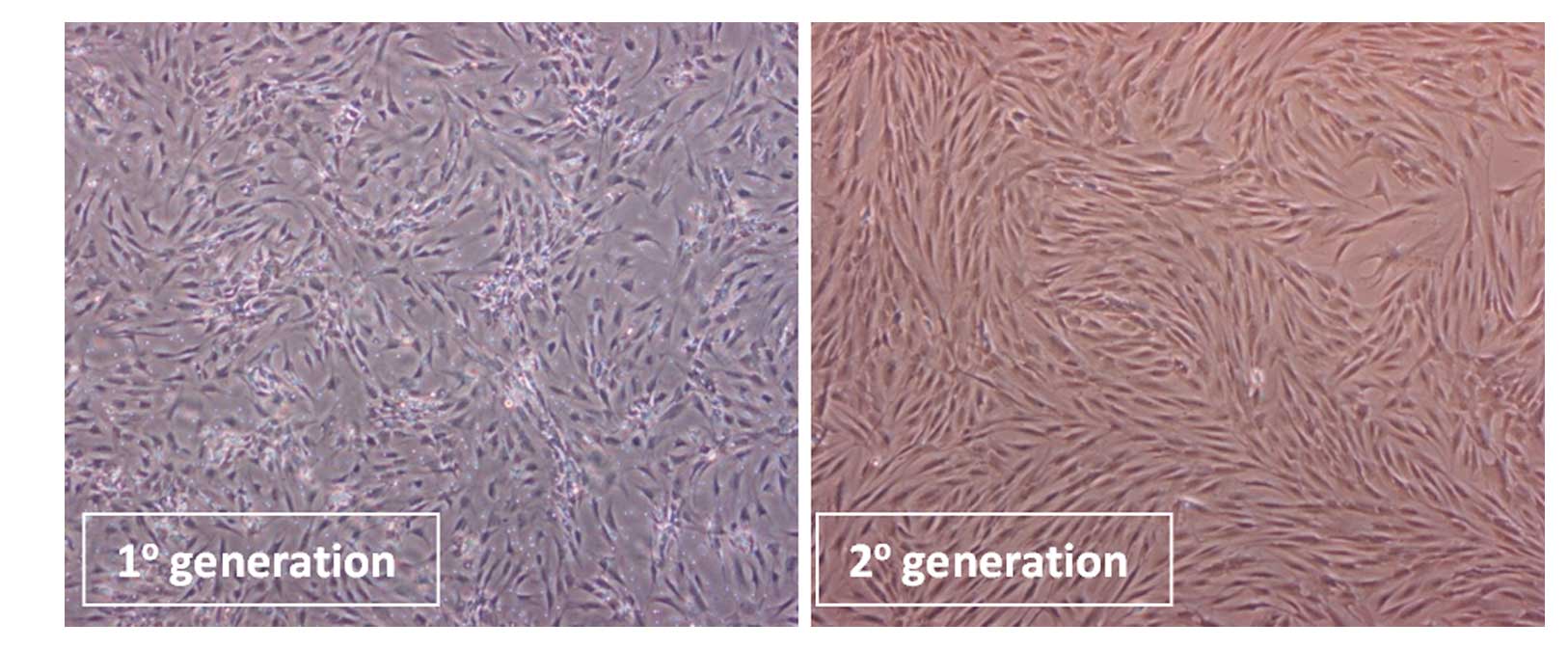
Inverted phase contrast microscopy was employed to
observe the morphology of ADCs in the first and second passage as
well as after separation (Fig.
1). The first passage ADCs consisted of different types of
cells with diverse morphologies, including short-spindle shape,
spindle shape, and flat shaped cells (Fig. 1, left panel). The second passage
cells were largely spindle shaped (Fig. 1, right panel).
Cell growth and morphology of ADCs
separated by CD105
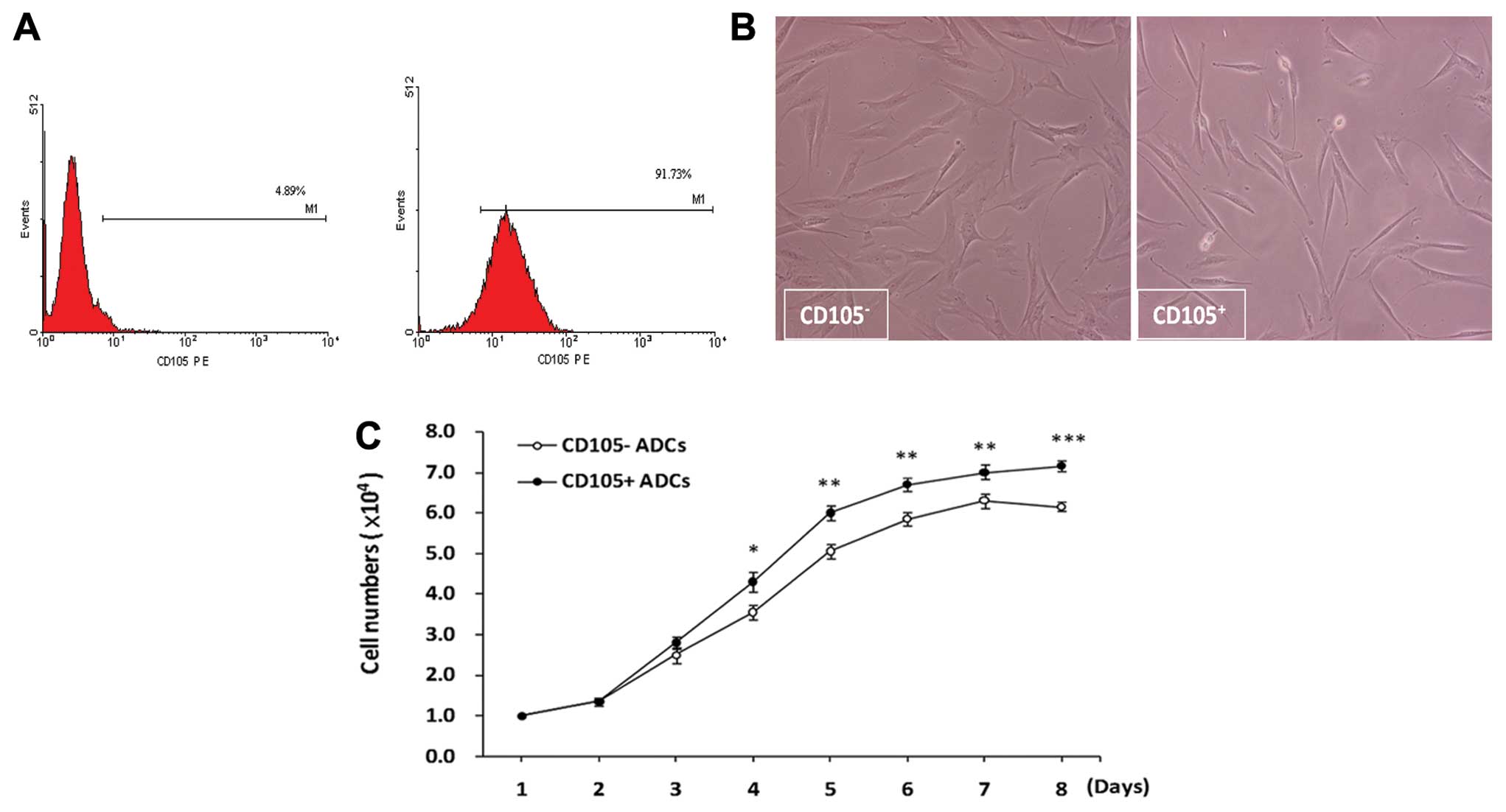
Prior to separation by CD105, 30% of ADCs were
positive for CD105. After enrichment for CD105 expression, the
proportion of CD105+ cells was increased to 90%,
suggesting that MACS successfully enriched CD105+
(Fig. 2A). After separation,
<5% of the CD105− population was positive for CD105
(Fig. 2A).
After separation, the morphology of
CD105− ADCs was fibroblast-like, and their growth was
directional, reaching near 100% confluence after five to 6 days of
culture (Fig. 2B, left panel).
The growth of CD105+ ADCs was relatively slow and was
characterized by clone-like growth; near 100% confluence was
reached after 6–7 days. CD105+ ADCs were largely
long-spindle shaped with a fraction of cells that were
short-spindle shaped, small-round or long-narrow-shaped (Fig. 2B, right panel).
CD105+ and CD105− ADC growth
was characterized by rapid growth after 3–4 days. CD105+
ADCs growth was continuous without a plateau. However,
CD105− ADC growth was suppressed after 6 days with a
doubling time of 72 h (Fig.
2C).
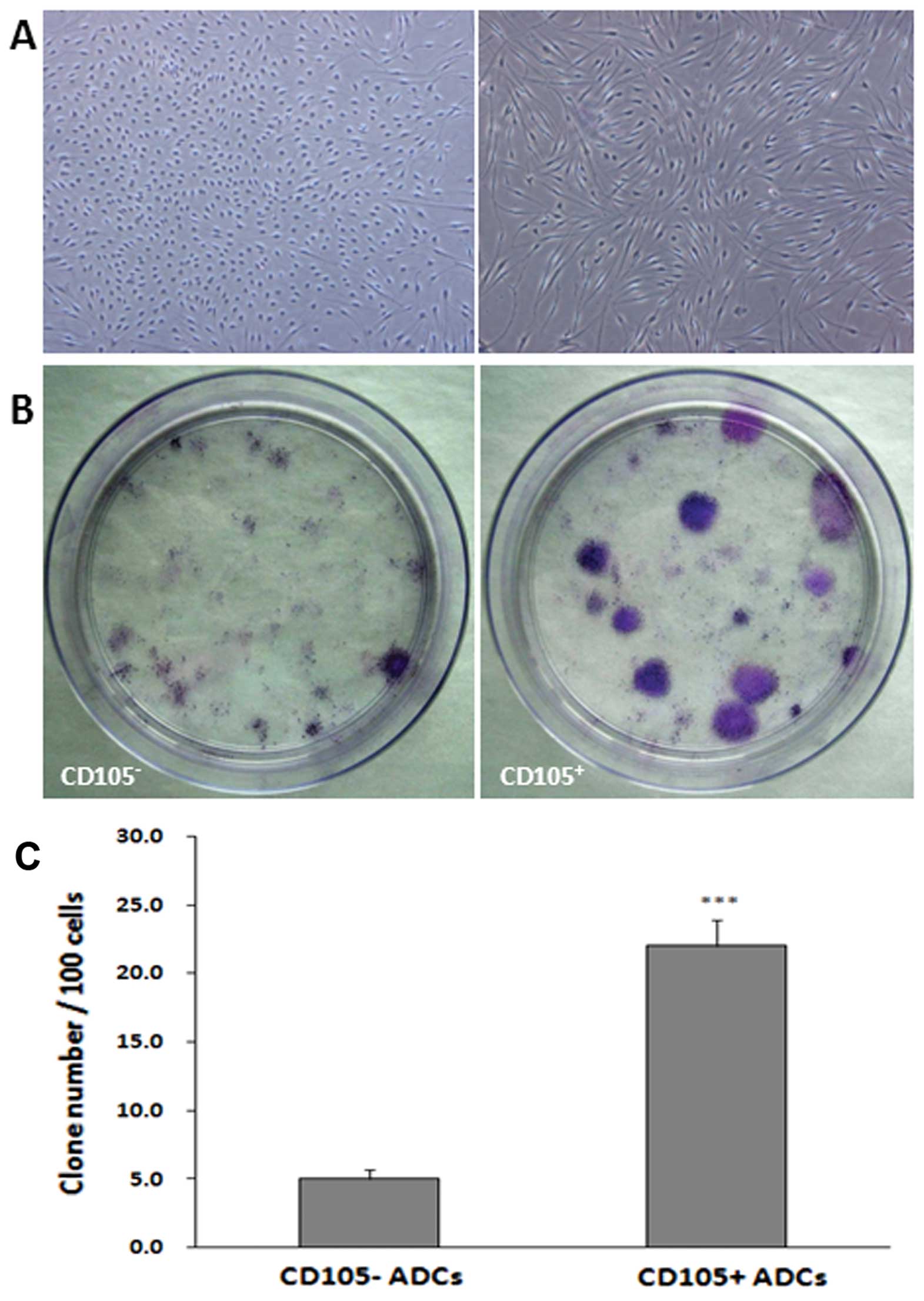
CD105+ cell colony
formation
Colony formation was assessed in both
CD105+ and CD105− cells (Fig. 3). After two weeks, single
cell-derived colonies were observed, consisting of 50–100 cells.
Cells within the colonies were long-spindle shaped, short-spindle
shaped, small-round or long-narrow shaped (Fig. 3A). CD105+ ADC colonies
enlarged gradually; the growth of CD105− ADCs was slow,
with some aging. After three weeks, Giemsa staining revealed
significantly larger CD105+ ADC colonies as compared to
CD105− ADCs (P<0.001) with diameters up to 1.5 cm
(Fig. 3B and C).
Differentiation potentials of
CD105+ and CD105 − ADCs
The potential for CD105+. ADCs to
differentiate into cartilage, bone and adipose tissue was assessed.
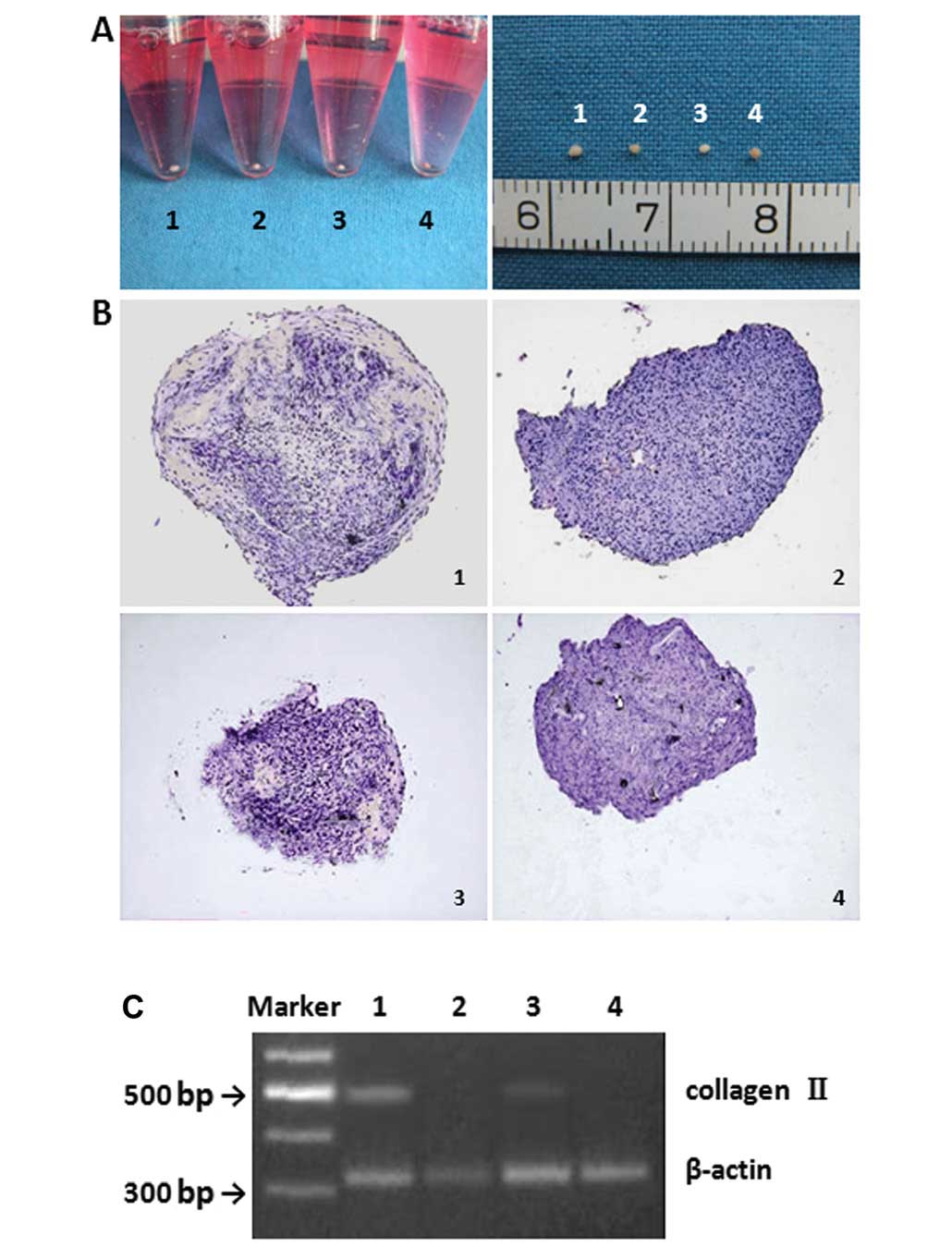
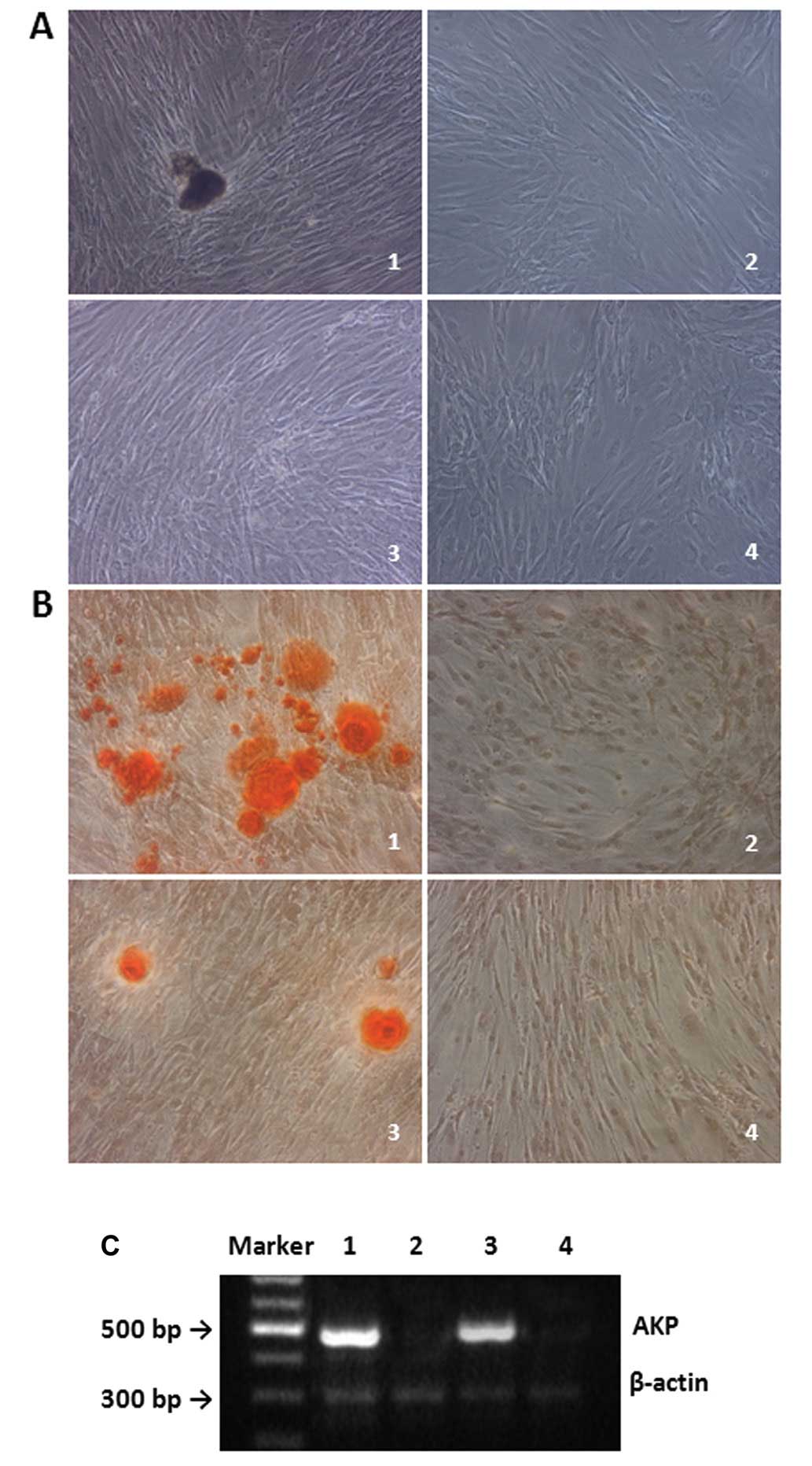
At 14 days after chondrogenetic induction, a round, pale, smooth,
elastic mass was observed (Fig.
4A). The mass observed from induced CD105+ ADC
(Fig. 4A, group 1) was larger and
more elastic than those derived from induced CD105− ADCs
(Fig. 4A, group 3). In the
non-induced groups (Fig. 4A,
groups 2 and 4), the cells were irregular, grey, soft and
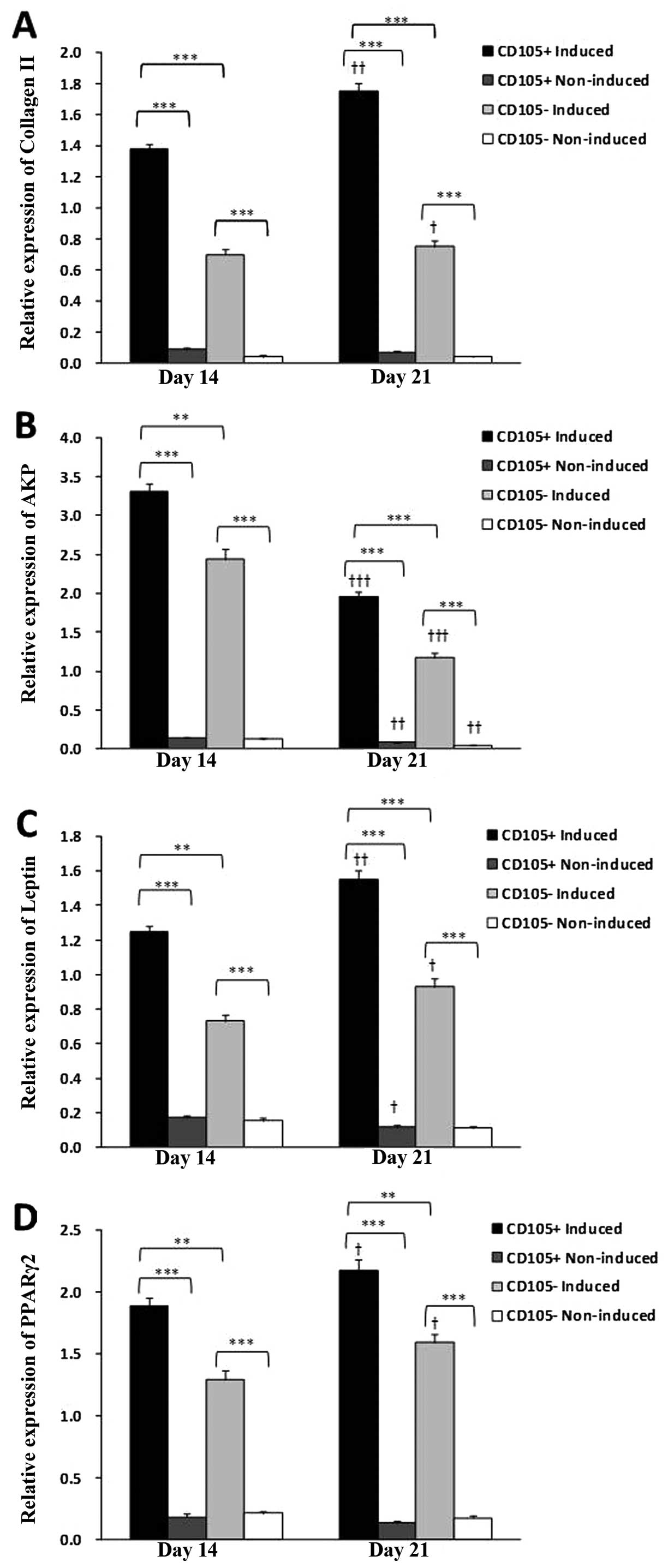
non-elastic. Collagen II expression was observed in both induced
groups; however, CD105+ ADCs had significantly greater
collagen II expression as compared to CD105− ADCs on
Days 14 and 21 (Figs. 4B and C
and 7A) (P<0.001). Collagen II
expression was significantly increased in both induced groups from
Days 14–21 (P<0.05). Both non-induced groups were negative for
type II collagen (Fig. 4B and
C).
Seven days after osteogenic induction, the long
spindle-shaped cells became flat, and had flake-like growth, which
was similar to osteocytes (Fig.
5A, groups 1 and 3). Fourteen days after osteogenic induction,
dark brown nodule-like deposits were found to be sporadic. The
number and volume of deposits were larger in CD105+
ADCs; no obvious changes in cell morphology were found in the
non-induced groups (Fig. 5A,
groups 2 and 4). Alizarin Red staining showed red, nodule-like
calcium deposits in the induced groups, which were not found in the
non-induced groups (Fig. 5B). AKP
expression was observed in both induced groups; however, it was
significantly higher in the induced CD105+ ADCs as
compared to induced CD105− ADCs (P<0.05) (Figs. 5C and 7B). AKP expression was significantly
decreased in both induced groups from Days 14–21 (P<0.001).
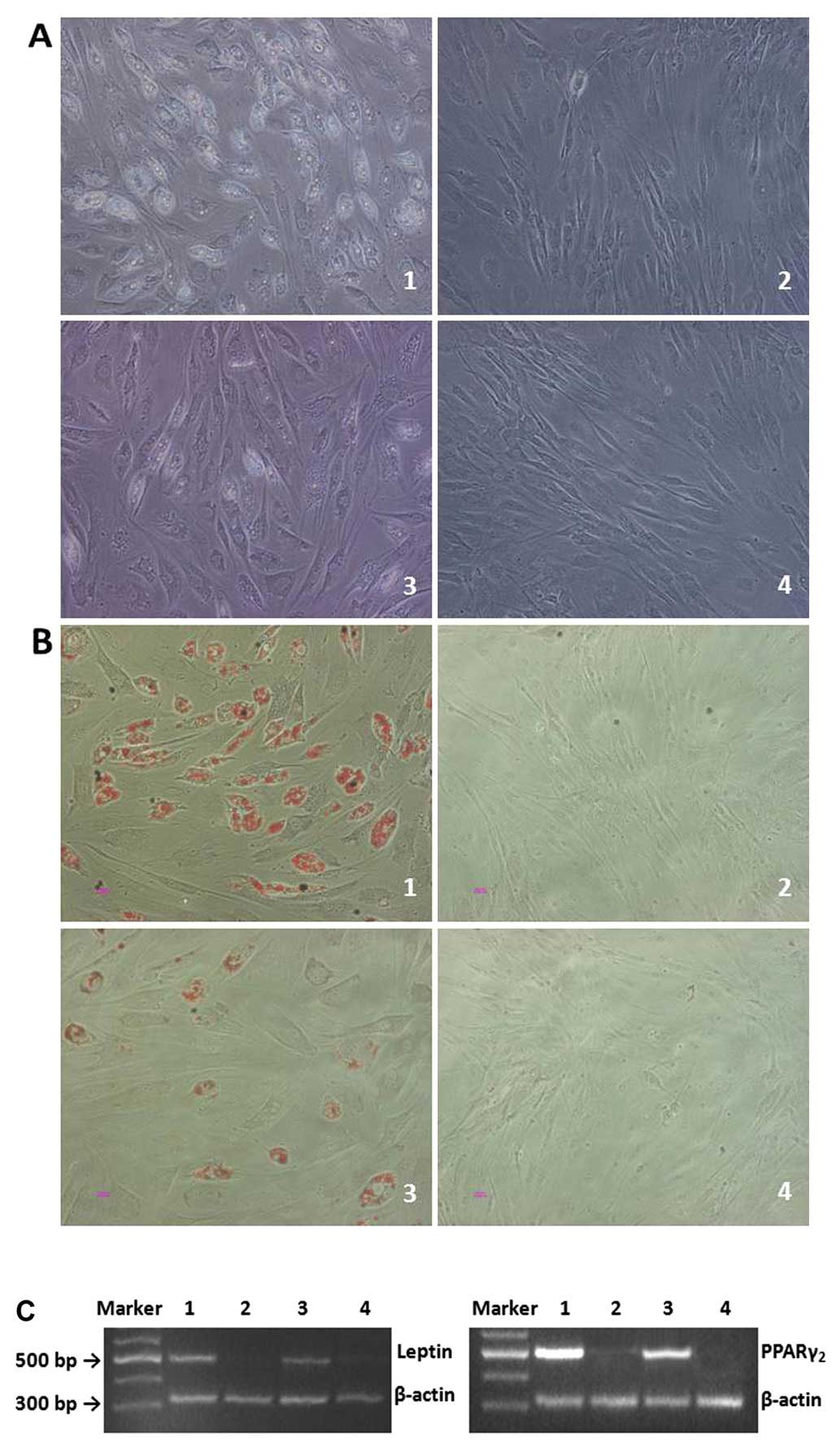
At 3 days after adipogenic induction, both
CD105+ and CD105− cells transformed from long
spindle-shaped to round-like. Seven days after induction, cell
growth became slow, and the morphology was also altered; small
lipid droplets were found in the cytoplasm; near mature adipocytes
were observed after 14 days (Fig.
6A, groups 1 and 3). The number of adipocytes and lipid
droplets was higher in the CD105+ ADC group than in the
CD105− ADC group. No obvious changes were observed in
the non-induced groups (Fig. 6A,
groups 2 and 4). Oil Red staining revealed lipid-containing
cytoplasms in induced cells, which were absent in the non-induced
groups (Fig. 6B). Increased
leptin and PPARγ2 expression was observed in both induced groups;
their expression was negative in both non-induced groups (Fig. 6C). Induced CD105+ ADCs
expressed significantly greater leptin and PPARγ2 expression as
compared to induced CD105-ADCs (P<0.01) (Fig. 7C and D). Leptin and PPARγ2
expression was significantly increased in both induced groups from
Days 14–21 (P<0.05).
Discussion
Although the advantages of ADSCs (i.e., simple
collection and high yield) are apparent, the maturity and
homogeneity of cartilages constructed using ADSCs were inferior to
those using BMSCs (8). This may
be attributed to the low proportion of MSCs in the isolated ADSCs.
In an effort to isolate ADSCs from a mixed-cell ADC population
isolated using suction-assisted lipectomy, ADCs were separated
based upon CD105 surface expression. CD105+ ADCs
displayed increased cell proliferation and colony formation as
compared to the CD105− cells. Upon induction,
CD105+ ADCs differentiated into chondrocytes,
osteocytes, and adipocytes to a greater extent than that observed
for CD105− ADCs, which is similar to other reports
demonstrating their ability to differentiate into chondrocytes
(8), osteoblasts (12), endothelial cells (13), muscle cells (14,15), and even hepatocytes (16).
Because culturing can change the expression of ADSCs
in mice (17), ADCs in the first
and second passage were analyzed for various stem cell-associated
surface antigens to determine which surface antigen may be useful
for isolation of ADSCs. Mitchell et al (18) similarly investigated the surface
markers on cells obtained from fat aspirates; the surface markers
on these cells included CD13, CD29, CD44, CD63, CD73, CD90, CD105
and CD166, which were similar to those observed on BMSCs. The
expression of these markers, including CD49D, CD44, CD90, CD105,
CD13 and CD71 were stable for up to 7 passages and 3 months in
culture (19). Izadpanah et
al (20) proposed that both
ADSCs and BMSCs were positive for the embryonic stem cell antigens,
Oct-4, Rex-1 and Sox-2. In the present study, ADCs were positive
for the MSC-related surface antigens, CD105, CD166 and STRO-1; the
percentage of cells expressing these markers increased from the
first to the second passage. In addition, ADCs expressed
hematopoietic and endothelial cell-related surface antigens,
including CD34 and CD31. These findings suggest that ADCs contain
MSCs as well as contaminant cells and possibly other non-MSCs.
To address the need for a simple method of obtaining
and purifying ADSCs from a mixed ADC population, we employed an
immunomagnetic bead assay with MACS similar as in Jiang et
al (21). Griesche et
al (22) compared the various
methods of obtaining homogeneous MSC populations from adipose
tissue, reporting a minor advantage of using immunomagnetic
isolation with the disadvantage of lower yeild. Because the
proportion of CD105+ cells was relatively stable between
the first and second passages, it was selected as the marker for
separation; the proportion of CD105+ cells was >90%
after separation, which is similar to that obtained using FACS
(21). Thus, separation of ADCs
using the immunomagnetic bead assay can achieve a target cell
population of high purity.
CD105 is a relatively specific marker for MSCs.
Within a population of BMSCs, CD105+ cells have been
shown to exhibit stronger stemness as compared to CD105−
cells (23). In addition,
CD105+ cells isolated from human (24) and rat (25) synovial fluid, human bone marrow
(26) as well as mouse
adipose-derived vascular fraction (27) have been reported to exhibit strong
chondrogenic potential.
To determine whether the separated cells had the
characteristics of stem cells, their morphology, self-renewal
capacity, and multipotentiality were assessed. CD105+
ADCs were largely long spindle-shaped with a small fraction of
cells being short spindle-shaped, small round, and long
narrow-shaped. The majority of CD105-ADCs were long spindle-shaped
with few processes; they were also larger. In addition, large
(>1 cm) colonies were observed from the CD105+ ADCs
after 3 weeks. Few single-cell colonies were found in
CD105− ADCs; the majority did not grow due to the
presence of aging cells, suggesting that the CD105− ADC
population contained differentiated cells. Furthermore,
CD105+ ADCs displayed significantly greater
proliferation as compared to CD105− ADCs. These results
differ from those published by Jiang et al (21). In their study ADCs were separated
based upon CD105 expression using FACS analysis; no differences in
proliferation or colony formation were noted. This may be caused by
differences in the tissue source, culture condition, and most
importantly the separation methods. Using magnetic beads might
improve the purity of the isolated cells, but parallel experiments
of these 2 methods should be performed to prove this
hypothesis.
The adipogenic, osteogenic, and chondrogenetic
potentials of the CD105+ cells were confirmed. Dramatic
differences in the chondrogenetic and osteogenic potentials were
observed between the CD105+ and CD105− ADCs,
which is similar to that observed by Jiang et al (21). Specifically, lacuna-like
structures were rarely found in cell micromass of CD105−
ADCs, and the amount of type II collagen was relatively small.
Differences in the adipogenic potential were relatively small
between the two cell populations, which may be attributed to the
presence of fat precursor cells in the CD105− ADCs.
Because the differentiation potential of MSCs is mainly reflected
in their chondrogenetic and osteogenic potentials, we speculate
that the proportion of MSCs in the CD105+ ADC population
is higher than that in CD105− ADCs. The limited
chondrogenetic and osteogenic potential observed for the
CD105− cells may be due to the following: i) a fraction
of CD105+ cells were present in the CD105−
cell population; ii) not all ADC-derived MSCs express CD105, and
ADSCs may consist of different cell types; and iii) the presence of
precursor cells, such as fat precursor cells, precursor
chondroblasts and precursor osteoblasts, in the CD105−
ADCs cannot be ruled out. However, further studies are required to
determine the therapeutic efficacy of using the CD105+
ADC population for tissue engineering.
The mechanism by which CD105 promotes MSC
chondrogenesis has yet to be determined. It is a member of the
TGF-β receptor superfamily, that binds to TGF-β1 and TGF-β3 by
associating with the TGF-β type II receptor and modulating cellular
responses to TGF-β (28).
Therefore, CD105 may mediate TGF-β-induced chondrogenesis (29,30). However, further studies are
necessary to determine the mechanism by which CD105 influences MSC
chondrogenesis.
The present study has its limitations. Firstly,
isolated ADSCs differentiated into chondrocytes, osteocytes, and
adipocytes all of which are mesodermal in origin. The capability
these isolated cells to differentiate to other germ layers needs
further analysis. Also, the differentiation potentials of these
cells was only assessed using in vitro studies; therefore,
further in vivo studies are necessary to confirm their
utility for tissue engineering purposes.
In conclusion, CD105+ ADCs were isolated
using MACS. The colony-formation ability and the chondrogenetic,
osteogenic, and adipogenic potentials of CD105+ ADCs
were superior as compared to CD105− ADCs. This study
provides theoretical and practical evidence for the identification
of new stem cell-specific markers and the use of MACS for the
separation and purification of ADSCs.
Abbreviations:
|
ADCs
|
adipose-derived cells;
|
|
ADSCs
|
adipose-derived stem cells;
|
|
AKP
|
alkaline phosphatase;
|
|
BMSCs
|
bone marrow stem cells;
|
|
CD105−
|
CD105-negative;
|
|
CD105+
|
CD105-positive;
|
|
FBS
|
fetal bovine serum;
|
|
H&E
|
hematoxylin and eosin;
|
|
IGF-1
|
insulin growth factor-1;
|
|
IBMX
|
3-isobutyl-1-methylxanthine;
|
|
PE
|
phycoerythrin;
|
|
SE
|
standard error;
|
|
TGF-β1
|
transforming growth factor-β1
|
Acknowledgements
We thank Dr Kai Liu and Dr Qun Zhang
of Shanghai Ninth People’s Hospital for the adipose tissue
collection; Mr. De-Min Yin and Ms. Juan-Juan Wu of Shanghai tissue
engineering center for the histological experiments; the
Superintendent, Dr Kai-ping Wang and the Director, Dr Xiao-chen
Tian of The Second Artillery General Hospital of the Chinese
People’s Liberation Army for suggestions with manuscript
preparation and financial support.
References
|
1.
|
S Heydarkhan-HagvallK Schenke-LaylandJQ
YangHuman adipose stem cells: a potential cell source for
cardiovascular tissue engineeringCells Tissues
Organs187263274200818196894
|
|
2.
|
DA RiderC DombrowskiAA SawyerAutocrine
fibroblast growth factor 2 increases the multipotentiality of human
adipose-derived mesenchymal stem cellsStem
Cells2615981608200810.1634/stemcells.2007-048018356575
|
|
3.
|
S BaglioniM FrancalanciR
SqueccoCharacterization of human adult stem-cell populations
isolated from visceral and subcutaneous adipose tissueFASEB
J2334943505200910.1096/fj.08-12694619584303
|
|
4.
|
L AustB DevlinSJ FosterYield of human
adipose-derived adult stem cells from liposuction
aspiratesCytotherapy6714200410.1080/1465324031000453914985162
|
|
5.
|
MJ VarmaRG BreulsTE SchoutenPhenotypical
and functional characterization of freshly isolated adipose
tissue-derived stem cellsStem Cells
Dev1691104200710.1089/scd.2006.002617348807
|
|
6.
|
JI HuangN KazmiMM DurbhakulaTM HeringJU
YooB JohnstoneChondrogenic potential of progenitor cells derived
fromhumanbonemarrowand adipose tissue: a patient-matched
comparisonJ Orthop
Res2313831389200510.1016/j.orthres.2005.03.008.110023062115936917
|
|
7.
|
GI ImYW ShinKB LeeDo adipose
tissue-derived mesenchymal stem cells have the same osteogenic and
chondrogenic potential as bone marrow-derived cells?Osteoarthritis
Cartilage13845853200510.1016/j.joca.2005.05.00516129630
|
|
8.
|
H AfizahZ YangJH HuiHW OuyangEH LeeA
comparison between the chondrogenic potential of human bone marrow
stem cells (BMSCs) and adipose-derived stem cells (ADSCs) taken
from the same donorsTissue
Eng13659666200710.1089/ten.2006.011817371203
|
|
9.
|
K YoshimuraT ShigeuraD
MatsumotoCharacterization of freshly isolated and cultured cells
derived from the fatty and fluid portions of liposuction aspiratesJ
Cell Physiol2086476200610.1002/jcp.2063616557516
|
|
10.
|
K LiuGD ZhouW LiuThe dependence of in vivo
stable ectopic chondrogenesis by human mesenchymal stem cells on
chondrogenic differentiation in
vitroBiomaterials2921832192200810.1016/j.biomaterials.2008.01.02118289667
|
|
11.
|
MP Hellio Le GraverandC RenoDA
HartInfluence of pregnancy on gene expression in rabbit articular
cartilageOsteoarthritis Cartilage6341350199810197169
|
|
12.
|
P NiemeyerM KornackerA MehlhornComparison
of immunological properties of bone marrow stromal cells and
adipose tissue-derived stem cells before and after osteogenic
differentiation in vitroTissue
Eng13111121200710.1089/ten.2006.0114
|
|
13.
|
V Planat-BenardJS SilvestreB
CousinPlasticity of human adipose lineage cells toward endothelial
cells: physiological and therapeutic
perspectivesCirculation109656663200410.1161/01.CIR.0000114522.38265.6114734516
|
|
14.
|
A Van DijkHW NiessenB Zandieh DoulabiFC
VisserFJ van MilligenDifferentiation of human adipose-derived stem
cells towards cardiomyocytes is facilitated by lamininCell Tissue
Res334457467200818989703
|
|
15.
|
Y ZhuT LiuK SongR NingX MaZ CuiADSCs
differentiated into cardiomyocytes in cardiac microenvironmentMol
Cell Biochem324117129200910.1007/s11010-008-9990-319107327
|
|
16.
|
A BanasT TerataniY YamamotoAdipose
tissue-derived mesenchymal stem cells as a source of human
hepatocytesHepatology46219228200717596885
|
|
17.
|
JR MaddoxX LiaoF LiC NiyibiziEffects of
culturing on the stability of the putative murine adipose derived
stem cells markersOpen Stem Cell
J15461200910.2174/187689380090101005419946473
|
|
18.
|
JB MitchellK McIntoshS
ZvonicImmunophenotype of human adipose-derived cells: temporal
changes in stromal-associated and stem cell-associated markersStem
Cells24376385200610.1634/stemcells.2005-023416322640
|
|
19.
|
V FolgieroE MiglianoM TedescoPurification
and characterization of adipose-derived stem cells from patients
with lipoaspirate transplantCell
Transplant1912251235201010.3727/09638910X51926521208530
|
|
20.
|
R IzadpanahC TryggB PatelC KriedtJ
DufourJM GimbleBA BunnellBiologic properties of mesenchymal stem
cells derived from bone marrow and adipose tissueJ Cell
Biochem9912851297200610.1002/jcb.2090416795045
|
|
21.
|
T JiangW LiuW LvPotent in vitro
chondrogenesis of CD105 enriched human adipose-derived stem
cellsBiomaterials3135643571201010.1016/j.biomaterials.2010.01.05020153525
|
|
22.
|
N GriescheW LuttmannA LuttmannT
StammermannH GeigerPC BaerA simple modification of the separation
method reduces heterogeneity of adipose-derived stem cellsCells
Tissues Organs192106115201010.1159/00028958620185896
|
|
23.
|
H AslanY ZilbermanL KandelOsteogenic
differentiation of noncultured immunoisolated bone marrow derived
CD105+ cellsStem
Cells2417281737200610.1634/stemcells.2005-054616601078
|
|
24.
|
MC ArufeA De la FuenteI Fuentes-BoqueteFJ
De ToroFJ BlancoDifferentiation of synovial CD-105(+) human
mesenchymal stem cells into chondrocyte-like cells through spheroid
formationJ Cell Biochem1081451552009
|
|
25.
|
J QiA ChenH YouK LiD ZhangF
GuoProliferation and chondrogenic differentiation of CD105-positive
enriched rat synovium-derived mesenchymal stem cells in
three-dimensional porous scaffoldsBiomed
Mater6015006201110.1088/1748-6041/6/1/01500621205995
|
|
26.
|
MK MajumdarV BanksDP PelusoEA
MorrisIsolation, characterization, and chondrogenic potential of
human bone marrow-derived multipotential stromal cellsJ Cell
Physiol18598106200010.1002/1097-4652(200010)185:1%3C98::AID-JCP9%3E3.0.CO;2-110942523
|
|
27.
|
D IshimuraN YamamotoK TajimaA OhnoY
YamamotoO WashimiH YamadaDifferentiation of adipose-derived stromal
vascular fraction culture cells into chondrocytes using the method
of cell sorting with a mesenchymal stem cell markerTohoku J Exp
Med216149156200810.1620/tjem.216.14918832797
|
|
28.
|
S CheifetzT BellónC CalésS VeraC BernabeuJ
MassaguéM LetarteEndoglin is a component of the transforming growth
factor-beta receptor system in human endothelial cellsJ Biol
Chem267190271903019921326540
|
|
29.
|
B JohnstoneTM HeringAI CaplanVM GoldbergJU
YooIn vitro chondrogenesis of bone marrow-derived mesenchymal
progenitor cellsExp Cell
Res238265272199810.1006/excr.1997.38589457080
|
|
30.
|
Y ItoP Bringas JrA MoghareiJ ZhaoC DengY
ChaiReceptor-regulated and inhibitory Smads are critical in
regulating transforming growth factor beta-mediated Meckel’s
cartilage developmentDev Dyn2246978200211984875
|