Introduction
Inflammatory bowel diseases (IBDs), including
ulcerative colitis (UC) and Crohn’s disease (CD), are chronic and
relapsing intestinal disorders of unknown etiology (1–5).
There is currently no pharmaceutical cure for IBD, and hence
treatment options aim for inducing and maintaining remission.
Steroids are used to induce remission but they cannot be used to
maintain remission (6).
Azathioprine, 6-mercaptopurine, methotrexate and biologics such as
infliximab or adalimumab are all options in treating IBD (7). Despite treatment with these agents,
a significant number of patients are resistant to conventional
therapies and require surgery during the course of the disease to
relieve their symptoms and to treat complications (8,9).
Regarding an alternative form of treatment, there is an increasing
number of reports demonstrating the clinical efficacy of tacrolimus
for steroid-dependent or steroid-refractory populations of IBD
patients (8–14).
Tacrolimus (FK506) is a macrolide immunosuppressive
agent isolated from Streptomyces tsukubaensis, and was first
used as an immunosuppressant in transplant patients (9,15–17). This immunomodulatory action of
tacrolimus is based on its ability to disrupt T-cell activation and
cytokine expression by binding to an intracellular protein called
FK binding protein (FKBP). The complex of tacrolimus plus FKBP
prevents the activation of calcineurin, resulting in an inhibition
of transcription factor NFAT, which plays an important role in the
transcriptional activation of genes for interleukin (IL)-2 and
interferon (IFN)-γ. IL-2 is a potent growth factor for T cells, and
IFN-γ is a strong inducer of Th1 response (18,19). Thus, the major immunosuppressive
effects of tacrolimus are considered to be mediated by a
suppression of T-cell responses via an inhibition of IL-2 and IFN-γ
secretion. However, whether tacrolimus has immunoregulatory effects
on other cell types, including mesenchymal cells, remains
unclear.
Chemokines and their receptors play a dominant role
in orchestrating the activity of monocytes/macrophages and T cells
in IBD (5), and, in particular,
there is evidence that CCL2 [monocyte chemoattractant protein-1
(MCP-1)] and CXCL10 [interferon-γ-inducible protein-10 (IP-10)] are
involved in human CD (20–23).
CCL2 signaling through CCR2 is a key factor in the recruitment and
activation of monocytes/macrophages, as well as in the recruitment
of T cells to mucosal lesions (24–26). CXCL10 is a ligand for the CXCR3
receptor, the activation of which results in the recruitment of T
cells and the perpetuation of mucosal inflammation (27).
In the present study, in order to investigate a
novel aspect of the molecular mechanisms underlying the
immunosuppressive effects of tacrolimus on intestinal inflammation,
we examined whether this agent had an impact on cytokine/chemokine
secretion in human colonic myofibroblasts. TNF-α plays a pivotal
role in the pathogenesis of IBD (5,28,29), and is widely-accepted as the major
target molecule of monoclonal antibody therapy for IBD (30). Therefore, we focused on the
effects of tacrolimus on TNF-α-stimulated cytokine/chemokine [IL-6,
CXCL8 (IL-8), CCL2 (MCP-1) and CXCL10 (IP-10)] expression in these
cells. To our knowledge, this is the first report describing the
immunosuppressive effects of tacrolimus on non-immune cells through
an inhibition of p38 MAP kinase activation.
Materials and methods
Reagents and antibodies
Recombinant human TNF-α and IFN-γ were purchased
from R&D Systems (Minneapolis, MN, USA). Inhibitors of p42/44
MAP kinases (PD98059), and an inhibitor of p38 MAP kinases
(SB203580) were purchased from Cell Signaling Technology, Inc.,
(Beverly, MA, USA). A PI3K inhibitor (LY294002) was purchased from
Calbiochem-Merck Co. (Darmstadt, Germany). All other reagents were
purchased from Sigma Chemical Co. (St. Louis, MO, USA). Goat
anti-p38 MAP kinase and anti-NF-κB (p65) antibodies were purchased
from Cell Signaling Technology, Inc. Tacrolimus was purchased from
Astellas Pharma, Inc. (Tokyo, Japan).
Culture of human colonic
myofibroblasts
Primary human colonic myofibroblasts were isolated
and cultured according to the method reported by Mahida et
al (31). The cellular
characteristics and culture conditions are also described in our
previous report (32).
Quantification of human CCL2 and
CXCL10
Antigenic CCL2 and CXCL10 in all samples were
quantified by sandwich enzyme-linked immunosorbent assay (ELISA)
kits purchased from R&D Systems.
Real-time PCR
The expression of mRNA in the samples was assessed
by real-time PCR analyses. Real-time PCR was performed using a
LightCycler 2.0 system (Roche Applied Science, Tokyo, Japan). The
PCR was performed using a SYBR-Green PCR Master Mix (Applied
Biosystems, Foster City, CA, USA). The oligonucleotide primers used
in this study are described in our previous reports (33–35). The data were normalized vs.
β-actin for human CCL2 and CXCL10.
Western blot analyses
The stimulated cells were lysed in an SDS sample
buffer containing orthovanadate. Western blot analyses were then
performed according to the method previously described (34). The detection was performed using
the enhanced chemiluminescence western blotting system.
Immunohistochemical analysis for p38 MAPK
activation
The cells were cultured in an 8-well chamber slide
system (Nalgene, Naperville, IL, USA) for predetermined times. The
cells were then fixed in cold methanol and acetone, and air dried.
The samples were incubated with an anti-p38 MAPK antibody and
subsequently incubated with a fluorescein isothiocyanate
(FITC)-labeled goat anti-rabbit IgG (Santa Cruz Biotechnology,
Inc., Santa Cruz, CA, USA).
Statistical analysis
The statistical significance of the differences was
determined by the Mann-Whitney U test (Statview version 4.5).
Differences resulting in P-values <0.05 were considered to be
statistically significant.
Results
Effects of tacrolimus on TNF-α-induced
cytokine/chemokine mRNA expression
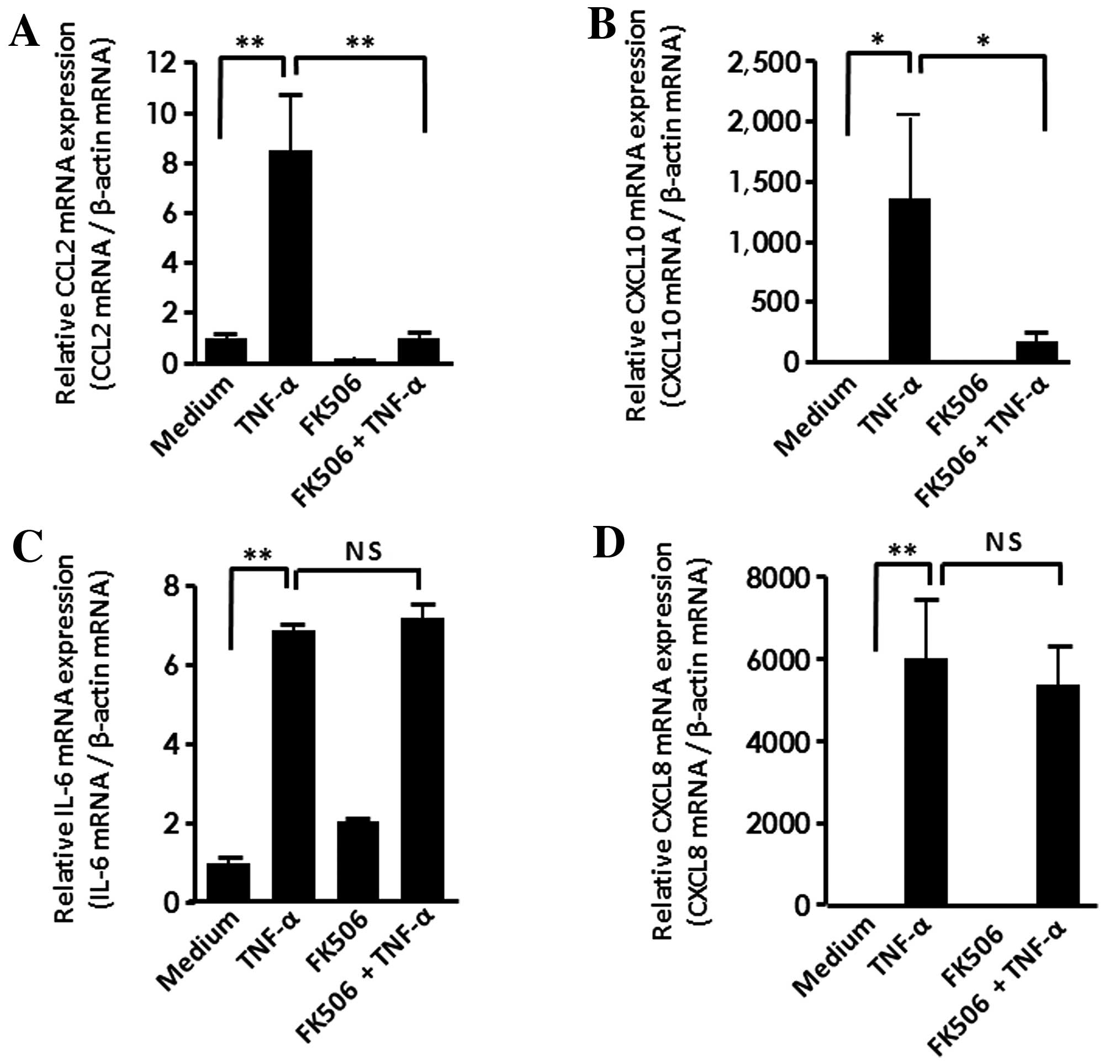
Human colonic myofibroblasts were stimulated for 24
h with TNF-α (30 ng/ml) in the presence or absence of tacrolimus (1
µM), and then the mRNA expression for CCL2, CXCL10, IL-6 and CXCL8
(IL-8) was analyzed by real-time PCR. TNF-α significantly
upregulated the mRNA expression for CCL2 and CXCL10, and the
presence of tacrolimus (1 µM) markedly suppressed the TNF-α-induced
CCL2 and CXCL10 mRNA expression (Fig.
1A and B). TNF-α also induced a significant increase in mRNA
expression for IL-6 and CXCL8, but tacrolimus failed to suppress
TNF-α-induced IL-6 and CXCL8 mRNA expression (Fig. 1C and D). These findings indicate
that tacrolimus selectively inhibits TNF-α-induced CCL2 and CXCL10
expression in human colonic myofibroblasts.
Dose-dependent inhibitory effects of
tacrolimus
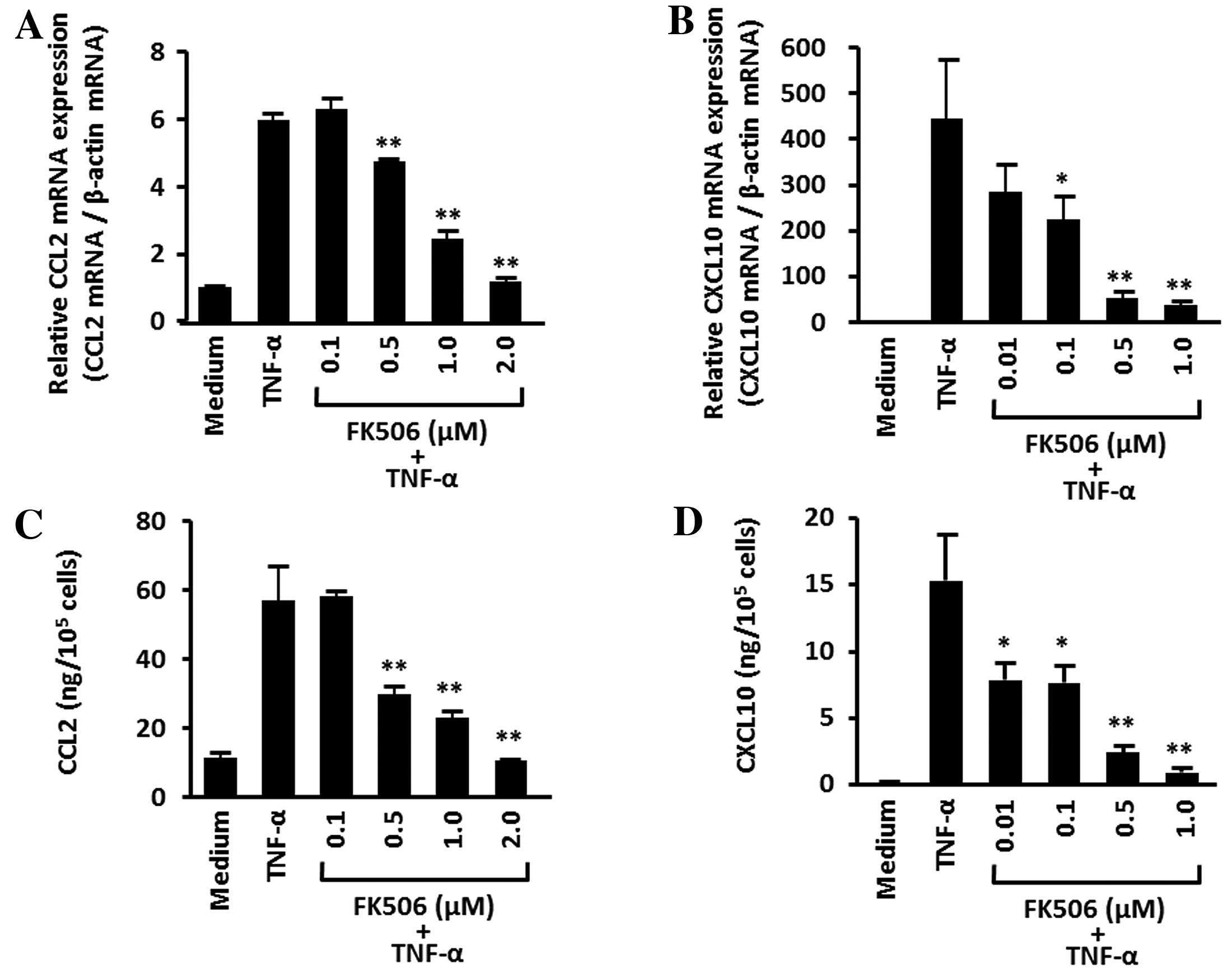
Human colonic myofibroblasts were stimulated for 24
h with TNF-α (30 ng/ml) in the presence of increasing
concentrations of tacrolimus, and the mRNA expression for CCL2 and
CXCL10 was analyzed by real-time PCR. Tacrolimus dose-dependently
suppressed TNF-α-induced CCL2 and CXCL10 mRNA expression (Fig. 2A and B). Significant inhibitory
effects for tacrolimus were observed at concentrations as low as
0.5 μM for CCL2 and 0.1 μM for CXCL10, respectively.
The inhibitory effects of tacrolimus were also
investigated at the protein level. The cells were stimulated for 24
h with TNF-α (30 ng/ml) in the presence of increasing
concentrations of tacrolimus, and the CCL2 and CXCL10 secretion
into the supernatants was analyzed by ELISA. Tacrolimus
dose-dependently suppressed TNF-α-induced CCL2 and CXCL10 protein
secretion (Fig. 2C and D).
Tacrolimus selectively inhibits p38 MAP
kinase activation
Previous studies have shown that the activation of
NF-κB and p38 MAP kinase plays an important role in TNF-α-induced
CXCL10 expression in various cell types (36–39). In addition, we have previously
demonstrated that the activation of NF-κB and p38 MAP kinase is
involved in TNF-α-induced IL-6, CXCL8 and CCL2 expression in human
colonic myofibroblasts (40).
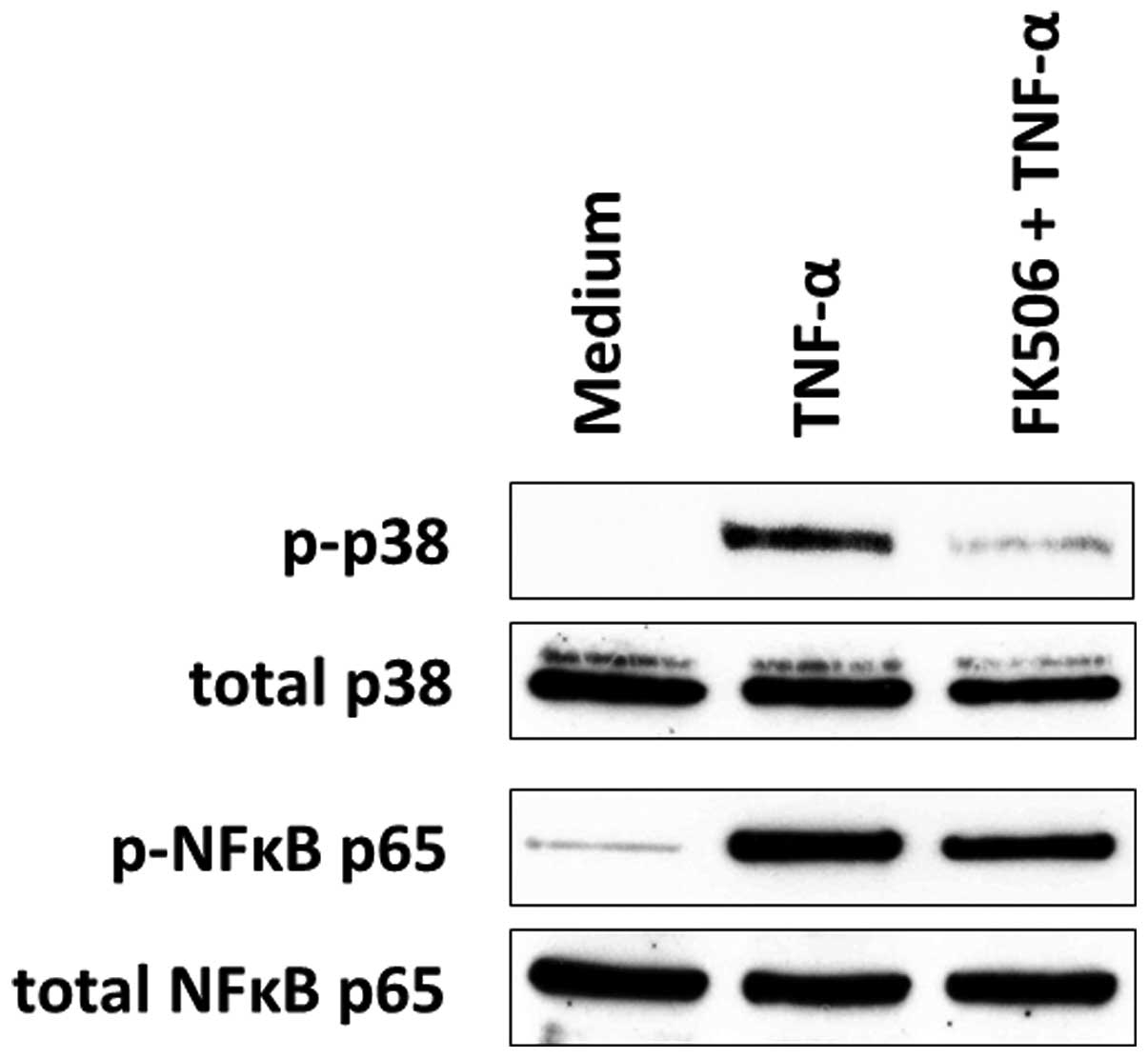
Based on this background information, we investigated how
tacrolimus affects the TNF-α-induced phosphorylation of NF-κB and
p38 MAP kinase in human colonic myofibroblasts. Tacrolimus had no
effect on NF-κB p65 phosphorylation, suggesting that the inhibition
of NF-κB activation is not involved in the inhibitory actions of
tacrolimus (Fig. 3). This is
supported by the finding that tacrolimus did not affect the
TNF-α-induced expression of IL-6 and CXCL8, which induction is
closely associated with NF-κB activation.
By contrast, tacrolimus strongly inhibited the
TNF-α-induced phosphorylation of p38 MAP kinase in these cells
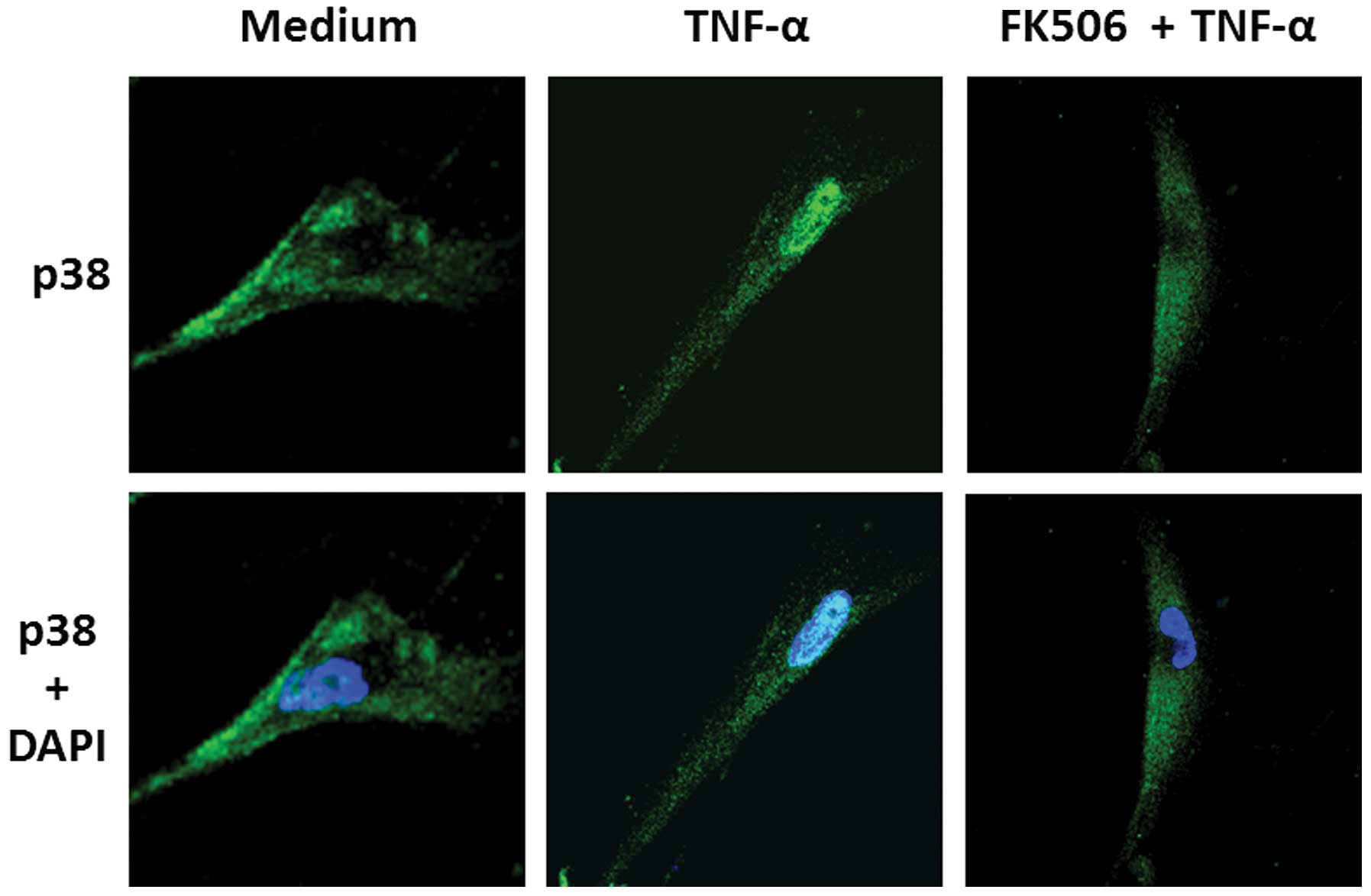
(Fig. 3). Furthermore,
immunohistochemical staining for p38 MAP kinase showed that TNF-α
rapidly induced the accumulation of p38 MAP kinase into the
nucleus, and that tacrolimus completely blocked this response
(Fig. 4). To confirm the role of
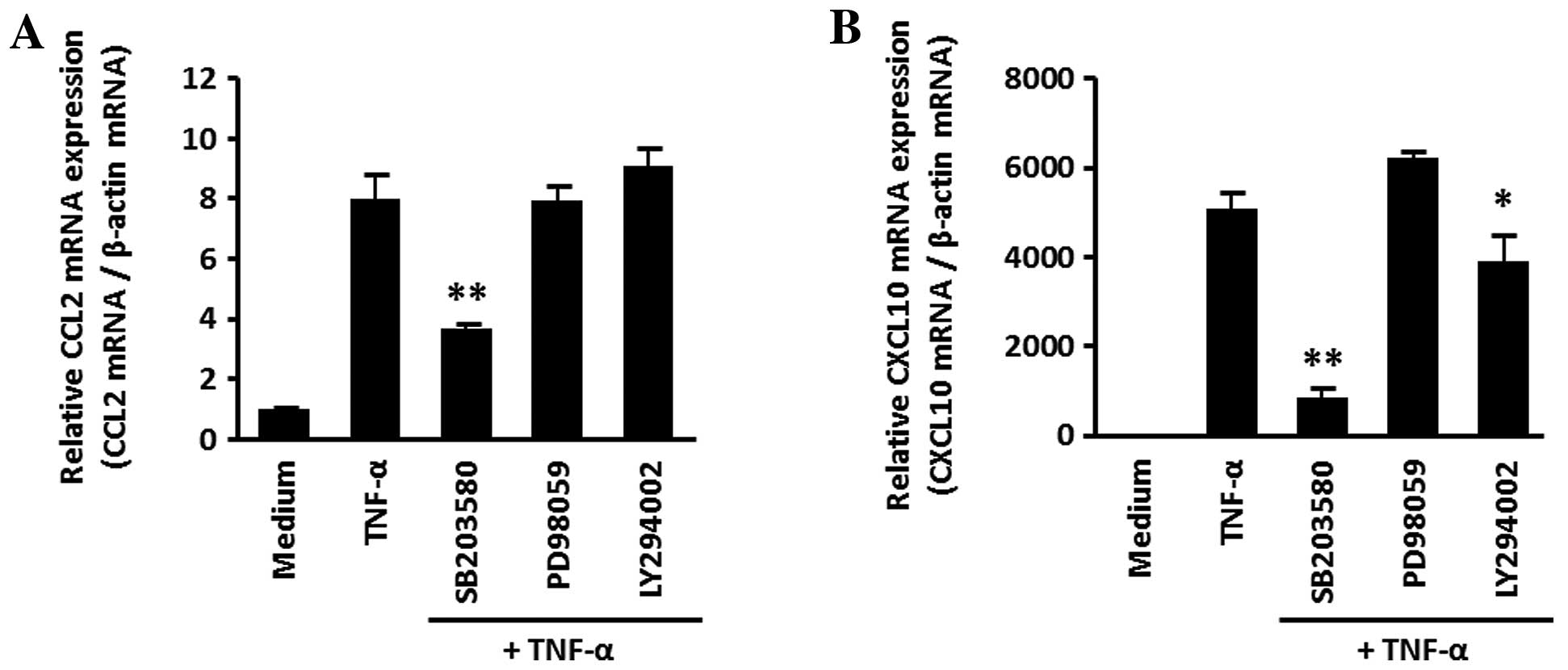
p38 MAP kinase in our system, the effects of various MAP kinase
inhibitors were studied. SB203580 (a specific inhibitor for p38 MAP
kinase) exhibited significant inhibition against TNF-α-induced CCL2
and CXCL10 expression, but p42/44 MAP kinase inhibitor (PD98059)
and a PI3 kinase inhibitor (LY294002) had no effect (Fig. 5).
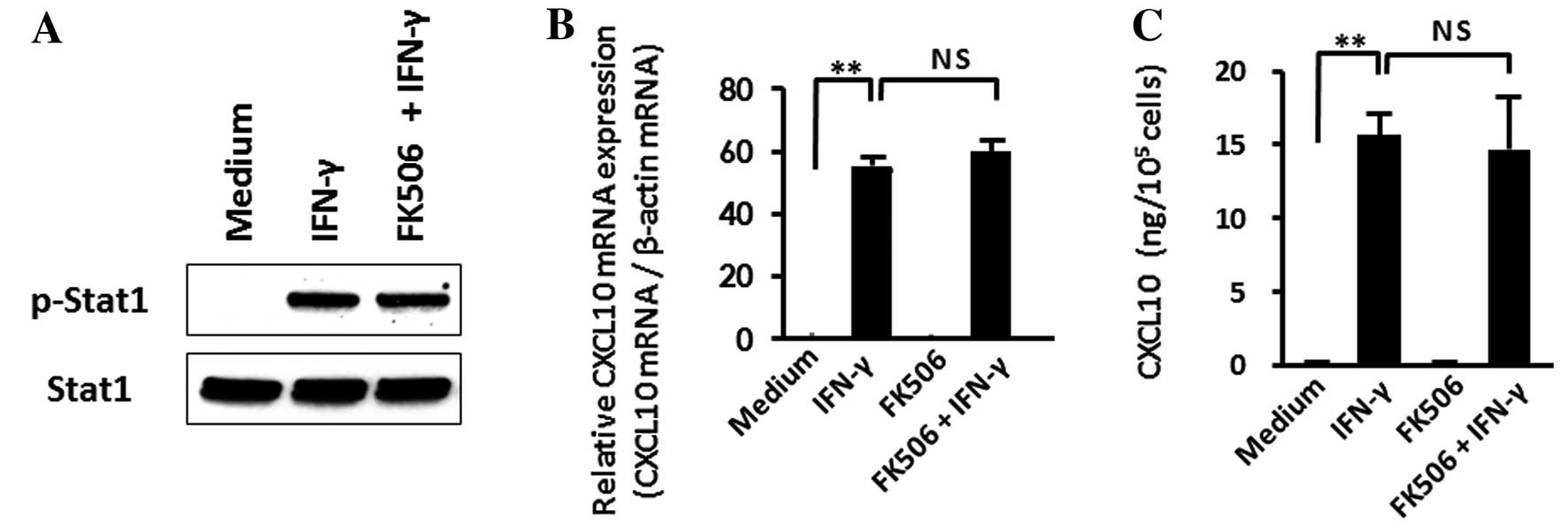
Tacrolimus does not affect IFN-γ-induced
CXCL10 expression
We previously demonstrated that IFN-γ induced CXCL10
expression via the activation of signaling transducer and activator
of transcription-1 (STAT1) in human colonic myofibroblasts
(35). Therefore, we tested the
effects of tacrolimus on this response. Tacrolimus did not affect
IFN-γ-induced STAT1 phosphorylation, nor did it modulate CXCL10
mRNA or protein expression (Fig.
6). These observations suggest that the inhibitory effects of
tacrolimus on CXCL10 expression are specific for TNF-α-induced
responses.
Discussion
In this study, we investigated whether tacrolimus
exerted inhibitory effects on human colonic myofibroblasts, and
found that tacrolimus directly inhibited TNF-α-stimulated chemokine
(CCL2 and CXCL10) production in these cells. Colonic myofibroblasts
are significantly involved in the regulation of a number of
epithelial cell functions, such as proliferation, differentiation,
and extracellular matrix (ECM) metabolism affecting the growth of
the basement membrane (40).
These findings indicate that tacrolimus modulates the immunological
functions of colonic myofibroblasts and contributes to the
therapeutic action of tacrolimus in IBD patients.
In human colonic myofibroblasts, tacrolimus did not
modulate TNF-α-induced IL-6 and CXCL8 expression, but selectively
inhibited TNF-α-induced CCL2 and CXCL10 production. Previous
studies have demonstrated that the activation of transcription
factor NF-κB is critical for the induction of all these
cytokine/chemokines (IL-6, CXCL8, CCL2 and CXCL10). However, the
selective inhibitory effects of tacrolimus on CCL2 and CXCL10
suggest that the modulation of NF-κB activation is not associated
with the inhibitory action of tacrolimus. This hypothesis is
supported by the finding that tacrolimus did not affect the
TNF-α-induced phosphorylation of the NF-κB p65 subunit. Thus, the
inhibitory actions of tacrolimus are possibly mediated by molecular
mechanisms independent of NF-κB activation.
In human colonic myofibroblasts, tacrolimus markedly
inhibited TNF-α-induced phosphorylation and translocation of p38
MAP kinase into the nucleus. The role of p38 MAP kinase in the
induction of CCL2 and CXCL10 has previously been reported (36–39), and the p38 MAP kinase inhibitor
SB203580 significantly blocked TNF-α-induced CCL2 and CXCL10
expression in human colonic myofibroblasts. These observations
suggest that the inhibitory actions of tacrolimus on CCL2 and
CXCL10 expression were mainly associated with an inhibition of p38
MAP kinase activation.
Several studies describe the role of CCL2/CCR2 and
CXCL10/CXCR3 in the pathophysiology of IBD (41–43). Immunohistochemical staining of gut
biopsy samples from CD patients revealed infiltrating
CD4+ T cells and monocytes/macrophages that are
uniformly positive for CCR2 (21). Furthermore, the disease phenotypes
of CD have been linked to polymorphisms in the CCL2 and CCR2 genes
(44,45). On the other hand, CXCL10
specifically activates the CXCR3 receptor, which is predominantly
expressed on activated T and B cells, natural killer cells,
dendritic and macrophage cells (46). Manousou et al (43) demonstrated an upregulation of CCR3
and its ligands in the inflamed mucosa of IBD patients.
CXCL10/CXCR3 is strongly expressed in IL-10 KO mice, a model of
human IBD, and the neutralization of IP-10 attenuated the colitis
in this model (47). Thus,
CCL2/CCR2- and CXCL10/CXCR3-mediated immune responses are regarded
as the key responses in the pathogenesis of IBD, and the inhibitory
actions of tacrolimus on CCL2 and CXCL10 production from colonic
myofibroblasts may be involved in the clinical immunosuppressive
effects of tacrolimus.
Tacrolimus is clinically used for prophylaxis
against organ rejection, autoimmune diseases and IBD. It is
generally accepted that the therapeutic effects of tacrolimus are a
result of its inhibitory actions on T-cell functions, but there is
little information concerning the inhibitory effects of tacrolimus
on non-immune cells such as colonic myofibroblasts. As an example
of the inhibitory actions of tacrolimus on non-T-cell populations,
Yoshino et al (48)
recently reported the immunosuppressive effects of tacrolimus on
macrophages. They demonstrated that the proinflammatory cytokine
production from tacrolimus-treated macrophages was significantly
lower than that from untreated cells. Tacrolimus suppressed the
LPS-induced activation of both NF-κB and MAPK in macrophages, and
induced the apoptosis of macrophages via the activation of
caspases-3 and −9. Combined with the observations of the present
study, tacrolimus may exert its immunosuppressive properties via
inhibitory actions on non-T cells, such as macrophages and colonic
myofibroblasts, as well as T cells.
In conclusion, our results confirm the potential
immunosuppressive effects of tacrolimus on human colonic
myofibroblasts. These inhibitory effects of tacrolimus on colonic
myofibroblasts may play a significant role in the treatment of
colonic inflammation in patients with IBD.
References
|
1.
|
DK PodolskyInflammatory bowel diseaseN
Engl J Med347417429200210.1056/NEJMra020831
|
|
2.
|
A MizoguchiE MizoguchiInflammatory bowel
disease, past, present and future: lessons from animal modelsJ
Gastroenterol43117200810.1007/s00535-007-2111-318297430
|
|
3.
|
RB SartorMechanisms of disease:
pathogenesis of Crohn’s disease and ulcerative colitisNat Clin
Pract Gastroenterol Hepatol33904072006
|
|
4.
|
A KoboriY YagiH ImaedaInterleukin-33
expression is specifically enhanced in inflamed mucosa of
ulcerative colitisJ
Gastroenterol459991007201010.1007/s00535-010-0245-120405148
|
|
5.
|
L MayerEvolving paradigms in the
pathogenesis of IBDJ
Gastroenterol45916201010.1007/s00535-009-0138-3
|
|
6.
|
WA Faubion JrEV Loftus JrWS HarmsenAR
ZinsmeisterWJ SandbornThe natural history of corticosteroid therapy
for inflammatory bowel disease: a population-based
studyGastroenterology121255260200110.1053/gast.2001.2627911487534
|
|
7.
|
KM TaylorPM IrvingOptimization of
conventional therapy in patients with IBDNat Rev Gastroenterol
Hepatol8646656201110.1038/nrgastro.2011.17221970871
|
|
8.
|
K McSharryAM DalzellK LeiperW
El-MatarySystematic review: the role of tacrolimus in the
management of Crohn’s diseaseAliment Pharmacol
Ther34128212942011
|
|
9.
|
M NaganumaT FujiiM WatanabeThe use of
traditional and newer calcineurin inhibitors in inflammatory bowel
diseaseJ
Gastroenterol46129137201110.1007/s00535-010-0352-z21132334
|
|
10.
|
A BensonT BarrettM SparbergAL
BuchmanEfficacy and safety of tacrolimus in refractory ulcerative
colitis and Crohn’s disease: a single-center experienceInflamm
Bowel Dis147122008
|
|
11.
|
H OgataT MatsuiM NakamuraA randomised dose
finding study of oral tacrolimus (FK506) therapy in refractory
ulcerative
colitisGut5512551262200610.1136/gut.2005.08179416484504
|
|
12.
|
H OgataJ KatoF HiraiDouble-blind,
placebo-controlled trial of oral tacrolimus (FK506) in the
management of hospitalized patients with steroid-refractory
ulcerative colitisInflamm Bowel
Dis18803808201210.1002/ibd.2185321887732
|
|
13.
|
S YamamotoH NakaseM MatsuuraS MasudaK
InuiT ChibaTacrolimus therapy as an alternative to thiopurines for
maintaining remission in patients with refractory ulcerative
colitisJ Clin
Gastroenterol45526530201110.1097/MCG.0b013e318209cdc421336140
|
|
14.
|
S YamamotoH NakaseS MikamiLong-term effect
of tacrolimus therapy in patients with refractory ulcerative
colitisAliment Pharmacol
Ther28589597200810.1111/j.1365-2036.2008.03764.x18549460
|
|
15.
|
RW BusuttilJR LakeRole of tacrolimus in
the evolution of liver
transplantationTransplantation77S44S51200410.1097/01.TP.0000126927.49589.3F15201686
|
|
16.
|
F VincentiA decade of progress in kidney
transplantationTransplantation77S52S61200410.1097/01.TP.0000126928.15055.DC15201687
|
|
17.
|
F RusnakP MertzCalcineurin: form and
functionPhysiological Rev8014831521200011015619
|
|
18.
|
W LiaoJX LinWJ LeonardIL-2 family
cytokines: new insights into the complex roles of IL-2 as a broad
regulator of T helper cell differentiationCurr Opin
Immunol23598604201110.1016/j.coi.2011.08.00321889323
|
|
19.
|
V LazarevicLH GlimcherT-bet in diseaseNat
Immunol12597606201110.1038/ni.205921685955
|
|
20.
|
UP SinghC VenkataramanR SinghJW Lillard
JrCXCR3 axis: role in inflammatory bowel disease and its
therapeutic implicationEndocr Metab Immune Disord Drug
Targets7111123200710.2174/18715300778083210917584151
|
|
21.
|
SJ ConnorN ParaskevopoulosR NewmanCCR2
expressing CD4+ T lymphocytes are preferentially
recruited to the ileum in Crohn’s
diseaseGut5312871294200415306587
|
|
22.
|
O GripS JanciauskieneAtorvastatin reduces
plasma levels of chemokine (CXCL10) in patients with Crohn’s
diseasePLoS One4e5263200919421322
|
|
23.
|
C BanksA BatemanR PayneP JohnsonN
SheronChemokine expression in IBD. Mucosal chemokine expression is
unselectively increased in both ulcerative colitis and Crohn’s
diseaseJ Pathol1992835200312474223
|
|
24.
|
HH ZhangK SongRL RabinCCR2 identifies a
stable population of human effector memory CD4+ T cells
equipped for rapid recall responseJ
Immunol18566466663201010.4049/jimmunol.090415620980630
|
|
25.
|
IF CharoW PetersChemokine receptor 2
(CCR2) in atherosclerosis, infectious diseases, and regulation of
T-cell
polarizationMicrocirculation10259264200310.1080/mic.10.3-4.259.26412851643
|
|
26.
|
YG KimN KamadaMH ShawThe Nod2 sensor
promotes intestinal pathogen eradication via the chemokine
CCL2-dependent recruitment of inflammatory
monocytesImmunity34769780201110.1016/j.immuni.2011.04.01321565531
|
|
27.
|
M LiuS GuoJM HibbertCXCL10/IP-10 in
infectious diseases pathogenesis and potential therapeutic
implicationsCytokine Growth Factor Rev22121130201121802343
|
|
28.
|
S DaneseJF ColombelW ReinischPJ
RutgeertsReview article: infliximab for Crohn’s disease treatment -
shifting therapeutic strategies after 10 years of clinical
experienceAliment Pharmacol Ther338578692011
|
|
29.
|
SB HanauerAA KornbluthJ MessickDT RubinWJ
SandbornBE SandsClinical scenarios in IBD: optimizing the use of
conventional and biologic agentsInflamm Bowel
Dis16S1S11201010.1002/ibd.2152921104734
|
|
30.
|
M AllezS VermeireN MozziconacciThe
efficacy and safety of a third anti-TNF monoclonal antibody in
Crohn’s disease after failure of two other anti-TNF
antibodiesAliment Pharmacol Ther31921012010
|
|
31.
|
YR MahidaJ BeltingerS MakhAdult human
colonic subepithelial myofibroblasts express extracellular matrix
proteins and cyclooxygenase-1 and −2Am J
Physiol273G1341G134819979435560
|
|
32.
|
T OkunoA AndohS BambaInterleukin-1beta and
tumor necrosis factor-alpha induce chemokine and matrix
metalloproteinase gene expression in human colonic subepithelial
myofibroblastsScand J
Gastroenterol37317324200210.1080/003655202317284228
|
|
33.
|
A AndohH TakayaT SaotomeCytokine
regulation of chemokine (IL-8, MCP-1, and RANTES) gene expression
in human pancreatic periacinar
myofibroblastsGastroenterology119211219200010.1053/gast.2000.853810889171
|
|
34.
|
M ShimadaA AndohK HataIL-6 secretion by
human pancreatic periacinar myofibroblasts in response to
inflammatory mediatorsJ
Immunol168861868200210.4049/jimmunol.168.2.86111777983
|
|
35.
|
O InatomiA AndohK KitamuraH YasuiZ ZhangY
FujiyamaButyrate blocks interferon-gamma-inducible protein-10
release in human intestinal subepithelial myofibroblastsJ
Gastroenterol40483489200510.1007/s00535-005-1573-4
|
|
36.
|
XF QiDH KimYS YoonEssential involvement of
cross-talk between IFN-gamma and TNF-alpha in CXCL10 production in
human THP-1 monocytesJ Cell
Physiol220690697200910.1002/jcp.2181519472212
|
|
37.
|
C WuY OhmoriS BandyopadhyayG SenT
HamiltonInterferon-stimulated response element and NF kappa B sites
cooperate to regulate double-stranded RNA-induced transcription of
the IP-10 geneJ Interferon
Res14357363199410.1089/jir.1994.14.3577897255
|
|
38.
|
DB ShultzMR RaniJD FullerRM RansohoffGR
StarkRoles of IKK-beta, IRF1, and p65 in the activation of
chemokine genes by interferon-gammaJ Interferon Cytokine
Res29817824200910.1089/jir.2009.003419929594
|
|
39.
|
CK WongCB WangWK IpYP TianCW LamRole of
p38 MAPK and NF-κB for chemokine release in coculture of human
eosinophils and bronchial epithelial cellsClin Exp
Immunol139901002005
|
|
40.
|
K HataA AndohM ShimadaIL-17 stimulates
inflammatory responses via NF-kappaB and MAP kinase pathways in
human colonic myofibroblastsAm J Physiol Gastrointest Liver
Physiol282G1035G1044200210.1152/ajpgi.00494.200112016129
|
|
41.
|
HC ReineckerEY LohDJ RinglerA MehtaJL
RombeauRP MacDermottMonocyte-chemoattractant protein 1 gene
expression in intestinal epithelial cells and inflammatory bowel
disease
mucosaGastroenterology1084050199510.1016/0016-5085(95)90006-3
|
|
42.
|
M UguccioniP GionchettiDF
RobbianiIncreased expression of IP-10, IL-8, MCP-1, and MCP-3 in
ulcerative colitisAm J
Pathol155331336199910.1016/S0002-9440(10)65128-010433925
|
|
43.
|
P ManousouG KoliosV ValatasIncreased
expression of chemokine receptor CCR3 and its ligands in ulcerative
colitis: the role of colonic epithelial cells in in vitro
studiesClin Exp
Immunol162337347201010.1111/j.1365-2249.2010.04248.x21077277
|
|
44.
|
H HerfarthB Pollok-KoppM GokeA PressM
OppermannPolymorphism of CC chemokine receptors CCR2 and CCR5 in
Crohn’s diseaseImmunol Lett77113117200111377705
|
|
45.
|
H HerfarthM GokeC HellerbrandPolymorphism
of monocyte chemoattractant protein 1 in Crohn’s diseaseInt J
Colorectal Dis184014052003
|
|
46.
|
CE EganMD CravenJ LengM MackKW SimpsonEY
DenkersCCR2-dependent intraepithelial lymphocytes mediate
inflammatory gut pathology during Toxoplasma gondii
infectionMucosal Immunol2527535200910.1038/mi.2009.105
|
|
47.
|
UP SinghS SinghDD TaubJW Lillard
JrInhibition of IFN-gamma-inducible protein-10 abrogates colitis in
IL-10-/-miceJ
Immunol17114011406200310.4049/jimmunol.171.3.140112874231
|
|
48.
|
T YoshinoH NakaseY
HonzawaImmunosuppressive effects of tacrolimus on macrophages
ameliorate experimental colitisInflamm Bowel
Dis1620222033201010.1002/ibd.2131820848491
|