Introduction
The neural precursor cell expressed, developmentally
downregulated 9 (NEDD9), also known as HEF1 and Cas-L (1,2),
acts as a scaffold protein and belongs to the family of
Crk-associated substrate (CAS) proteins that regulate protein
complexes controlling cell attachment, migration, invasion, cell
cycle, apoptosis, and oncogenic signal transduction (3). Overexpression of the NEDD9 protein
has now been strongly linked to poor prognosis in cancer, as well
as resistance to first-line chemotherapeutics in multiple tumor
types including breast cancer (4), glioblastoma (5) and melanoma (6). Lung cancer is still a serious health
problem and remains the first most common type of fatal cancer
worldwide (7). High levels of
NEDD9 mRNA and protein have been shown to be present in human lung
adenocarcinoma tissues (8). In
cell lines, NEDD9 has been identified as a metastasis-promoting
gene (9).
Small interfering RNA (siRNA) are short pieces of
double-stranded RNA (19–22 nt). The molecule works by cleaving and
destroying its cognate RNA. siRNA first assembles into RNA-induced
silencing complexes (RISCs), where it activates the complex by
unwinding its RNA strands. The unwound RNA strands subsequently
guide the complex to the complementary RNA molecules, where the
complex cleaves and destroys the cognate RNA, resulting in the RNA
interference (RNAi) phenomenon. RNAi has evolved into a powerful
tool to study gene functions and it also has potential applications
in gene therapy. RNAi has evolved into a powerful molecular
biological tool to study gene functions (10,11), with potential applications in gene
therapy.
The current study constructed and screened for short
hairpin RNA (shRNA) in response to the NEDD9 targeting gene. The
NEDD9 siRNA was transfected into the human lung adenocarcinoma cell
line A549 with Lipofectamine 2000. The cells were transfected with
siRNA and the untreated cells were used as controls. The inhibitory
effect of siRNA on the expression of NEDD9 mRNA and protein was
detected by fluorescence quantitative RT-PCR (FQ-PCR) and western
blotting.
Materials and methods
Reagents and instruments
The pRNAT-CMV3.2 plasmid was purchased from
GenScript Co. (USA). E. coli DH5α and restriction enzymes
were obtained from the Department of Microorganism and
Immunization, Preclinical Medicine, Zhengzhou University (China).
T4 DNA ligase and marker DL2000 were purchased from Takara Bio,
Inc. (Japan). Agarose was purchased from Oxoid (UK). The plasmid
DNA mini-preparation, the DNA gel extraction and PCR kits were
purchased from Axygen (USA). The inverted fluorescence microscope
was from Olympus Corporation (Japan).
Cell culture
The human lung adenocarcinoma cell line A549 was a
gift from the Sino-British Research Centre for Molecular Oncology
of Zhengzhou University. A549 cells were cultured in EMDM medium
(Hyclone, Logan, UT, USA) supplemented with 10% fetal bovine serum
(FBS) (Hyclone), 100 IU/ml penicillin and 100 μg/ml streptomycin
(Hyclone). Cells were maintained at 37°C in a humidified chamber
containing 5% CO2.
siRNA design
According to the principles of siRNA design and the
Homo sapien NEDD9 gene sequence (GenBank accession no.
NM_182966), three pairs of specific siRNA sequences
GCTGCCGAAATGAAGTATA (159–117), GGGCCTTATAT GACAATGT (193–211),
GTGTCCTATTTCTTAGTGA (648-666) and the negative control siRNA vector
sequence TTCTCCGAACGTCGCACGT (CO) were synthesized using software
provided at http://www.mpibpc.gwdg.de/abteilungen/100/105/sirna.html.
These siRNA sequences were composed of a sense strand, an antisense
strand, reverse complementary sequences and a loop. A TTTTT
termination signal was introduced to the 3′-end of the oligos.
BamHI and XhoI restriction enzyme digestion sites
were also designed in shRNA for further cloning. The eight
oligonucleotides were synthesized by Shanghai BioSune Biotechnology
Co. (Shanghai, China) (Table
I).
 | Table IDesigned oligonucleotide sequences
targeting the NEDD9 gene. |
Table I
Designed oligonucleotide sequences
targeting the NEDD9 gene.
| NEDD9 | Sequences
(5′→3′) |
|---|
| N159F |
5′-GATCCGCTGCCGAAATGAAGTATATTCAAGAGATATACTTCATTTCGGCAGCTTTTTTC-3′ |
| N159R |
5′-TCGAGAAAAAAGCTGCCGAAATGAAGTATATCTCTTGAATATACTTCATTTCGGCAGCG-3′ |
| N193F |
5′-GATCCGGGCCTTATATGACAATGTTTCAAGAGAACATTGTCATATAAGGCCCTTTTTTC-3′ |
| N193R |
5′-TCGAGAAAAAAGGGGCCTTATATGACAATGTTCTCTTGAAACATTGTCATATAAGGCCCG-3′ |
| N648F |
5′-GATCCGTGTCCTATTTCTTAGTGATTCAAGAGATCACTAAGAAATAGGACACTTTTTTC-3′ |
| N648R |
5′-TCGAGAAAAAAGTGTCCTATTTCTTAGTGATCTCTTGAATCACTAAGAAATAGGACACG-3′ |
| COF |
5′-GATCCTTCTCCGAACGTCGCACGTTTCAAGAGAACGTGACACGTTCGGAGAATTTTTTC-3′ |
| COR |
5′-TCGAGAAAAAATTCTCCGAACGTCGCACGTTCTCTTGAAACGTGACACGTTCGGAGAAG-3′ |
Annealing and purification of specific
target sequences
The four pairs of synthesized oligonucleotides
(N159F and N159R, N193F and N193R, N648F and N648R, COF and COR)
were dissolved in 50 μl ultrapure water. From these stocks, 13.5 μl
of each oligonucleotide pair was mixed with 3 μl of 10X buffer. The
annealing step was performed by a 5-min incubation at 95°C,
followed by a 10 min incubation at 70°C. The annealed oligos were
then slowly cooled to 37°C. The annealed NEDD9 and short hairpin
DNA control oligos were loaded onto 2.0% agarose gels for
separation and purification of the vector.
Annealed products ligated into the
pRNAT-CMV3.2 vector
DNA ligation was performed by incubating 10 μl of
the reaction solution containing 3 μl of the annealed
oligonucleotides, 1 μl linear pRNAT-CMV3.2 after BamHI and
XhoI enzyme double digestion, 1 μl of 10X T4 buffer and 1 μl
T4 DNA ligase and incubated at 16°C overnight. The recombinant
vector was transformed into DH5α competent cells. Positive clones
of vector pRNAT-CMV3.2-N159, pRNAT-CMV3.2-N193, pRNAT-CMV3.2-N648
and pRNAT-CMV3.2-C were randomly selected and inoculated into 30 ml
of LB medium supplemented with ampicillin sodium. After the cells
were cultured in a shaking incubator overnight (16–18 h) at 37°C,
the DNA plasmids were extracted using a plasmid DNA
mini-preparation kit according to the manufacturer’s instructions,
and identified by 1% agarose gel electrophoresis after restriction
enzyme digestion. DNA Marker 2000 was used to estimate the length
of the fragments. The confirmed positive clones producing the four
vectors were randomly selected and inoculated into 30 ml LB medium
supplemented with ampicillin sodium. After the cells were cultured
in a shaking incubator overnight (16–18 h) at 37°C, 1 ml of medium
was used for sequencing.
Plasmid transfection into the A549 cell
line
Liposomes were used to facilitate transfection.
Metafectene liposomes and the plasmid were mixed in a ratio of 3:1
(12). The culture medium
containing serum and antibiotic was removed prior to transfection
and washed three times with PBS, followed by the addition of
culture medium but without serum and antibiotics. After 1 h, the
A549 cells were transfected (when the cells reached 80–90%) by
placing the mixture in 6-well culture plates and adding 0.8 ml of
culture medium without serum; the plates were incubated at 37°C for
6–8 h. After incubation, 2 ml of the culture medium with 20% FBS
was added in 5% CO2 and cultured for a further 48 h at
37°C. Transfection efficiency was observed using an inverted
fluorescence microscope as the vector carries the coral green
fluorescent protein (cGFP) as a marker.
RNA isolation and establishment of a
method of FQ-PCR detection of NEDD9 mRNA
Cultured A549 cells were divided into six groups and
either transfected with the three vector plasmids
pRNAT-CMV3.2-N159, pRNAT-CMV3.2-N193 and pRNAT-CMV3.2-N648,
transfected with an empty vector plasmid, transfected with a
plasmid containing an unrelated pRNAT-CMV3.2-C sequence, or no
plasmid. The relative rate of NEDD9 mRNA expression in these six
groups was analyzed via FQ-PCR, and the rate of suppression of mRNA
expression was calculated.
Total RNA was purified from cells using a TRIzol
reagent (Invitrogen, Carlsbad, CA, USA). The first-strand of the
cDNA was synthesized using 2.5 μg RNA and AMV retroviridase
(Promega). Specific primers were designed in accordance with the
mRNA sequence of the NEDD9 gene and the NEDD9 and GAPDH segments
were amplified. The primers were provided by Shanghai
Bioengineering Co. Primers for the NEDD9 gene were upstream,
5′-CGTGGGTAAAAAGGTGTTCC-3′ and downstream,
5′-CAAGCCTCCAAACTCAGGAC-3′ (amplified segment 124 bp); primers for
GAPDH were upstream, 5′-TCGTGG AAGGACTCATGACC-3′ and downstream,
5′-AGGGATGA TGTTCTGGAGAG-3′ (amplified segment 97 bp).
NEDD9 mRNA levels were quantified by FQ-PCR based on
TaqMan™ technology, using the ABI PRISM 7500 Sequence Detection
System (Applied Biosystems, Foster City, CA, USA). The reaction
system consisted of 20 μl 2X real-time PCR buffer, 0.5 μl of the
upstream and downstream primers of NEDD9, 2 μl reverse
transcription product, 0.2 μl TaqDNA polymerase, and
ddH2O to increase the final volume to 40 μl. FQ-PCR
reaction conditions included pre-denaturation at 95°C for 3 min;
95°C for 15 sec, 65°C for 45 sec, 40 cycles; 72°C for 2 min. GAPDH
was simultaneously amplified as an internal reference. All
reactions were performed in triplicate. The results were analyzed
by calculating the Ct values for NEDD9 and GAPDH in the samples and
the relative expression of NEDD9 mRNA in each group [the relative
fold (RF)] using 2−ΔΔCt value was calculated.
Protein isolation and western blot
analysis
NEDD9 expression was examined in A549 cells
transfected with plasmid vectors using western blotting. Cells were
harvested in lysis buffer (2% SDS, 50 mM Tris, pH 7.4/1 mM
EDTA/protease inhibitor mixture) 48 h after transfection and
homogenized by sonication. Their protein concentrations were
determined using a DTX 880 Multimode Detector (Beckman Coulter,
Fullerton, CO, USA). Equal amounts of protein (40 μg) were
separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) on 8% gels and were transferred onto
nitrocellulose membranes (Hyclone). The membranes were blocked with
5% (v/v) skimmed milk and probed with phospho rabbit-anti-human
polyclonal primary antibodies for NEDD9 (93 kDa) (Abcam, San
Francisco, CA, USA), at 4°C overnight. The membranes were washed
and incubated with horseradish peroxidase (HRP)-conjugated
goat-anti-rabbit secondary antibody (Beijing Zhong Shan Golden
Bridge Biological Technology), at room temperature for 1 h.
Antibodies against p-actin (Santa Cruz Biotechnology, Inc., Santa
Cruz, CA, USA) were used to measure protein loading. The bound
antibodies were visualized using an electroche-miluminescence
system (Amersham Pharmacia Biotech, Buckinghamshire, UK).
Statistical analysis
The SPSS 17.0 (SPSS Inc., USA) for Windows
statistical software package was used for analysis. One-way
analysis of variance (ANOVA) was used to investigate the
differences in NEDD9 mRNA expression among the groups in
vitro. A P-value <0.05 indicated a significant difference,
and P-value <0.01 indicated a highly significant difference.
Results
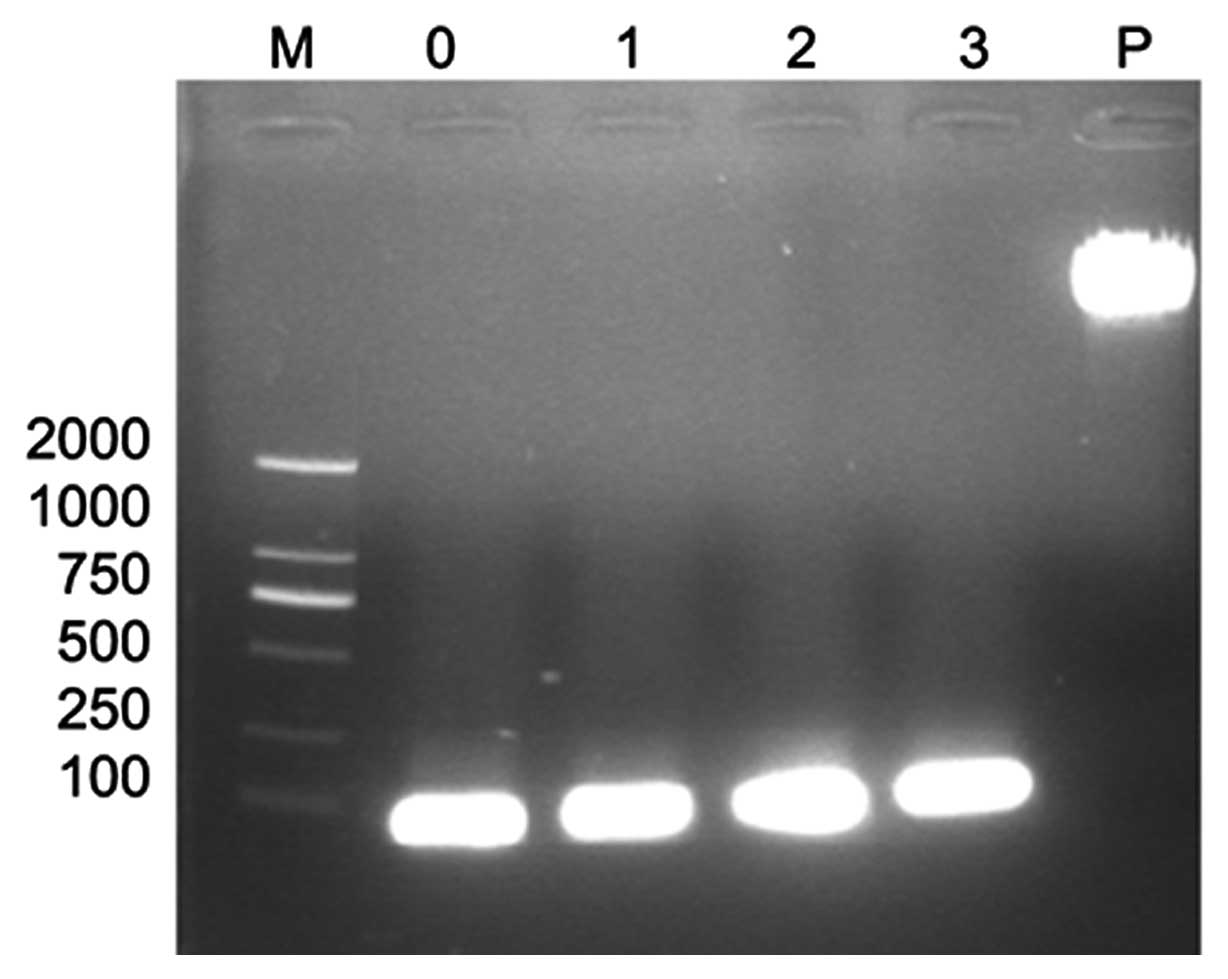
Annealed oligonucleotides
After annealing the single-strand oligonucleotides
(N159, N193, N648, CO) they were visualized by gel electrophoresis.
As expected, bright bands with a length of 59 bp were observed
(Fig. 1).
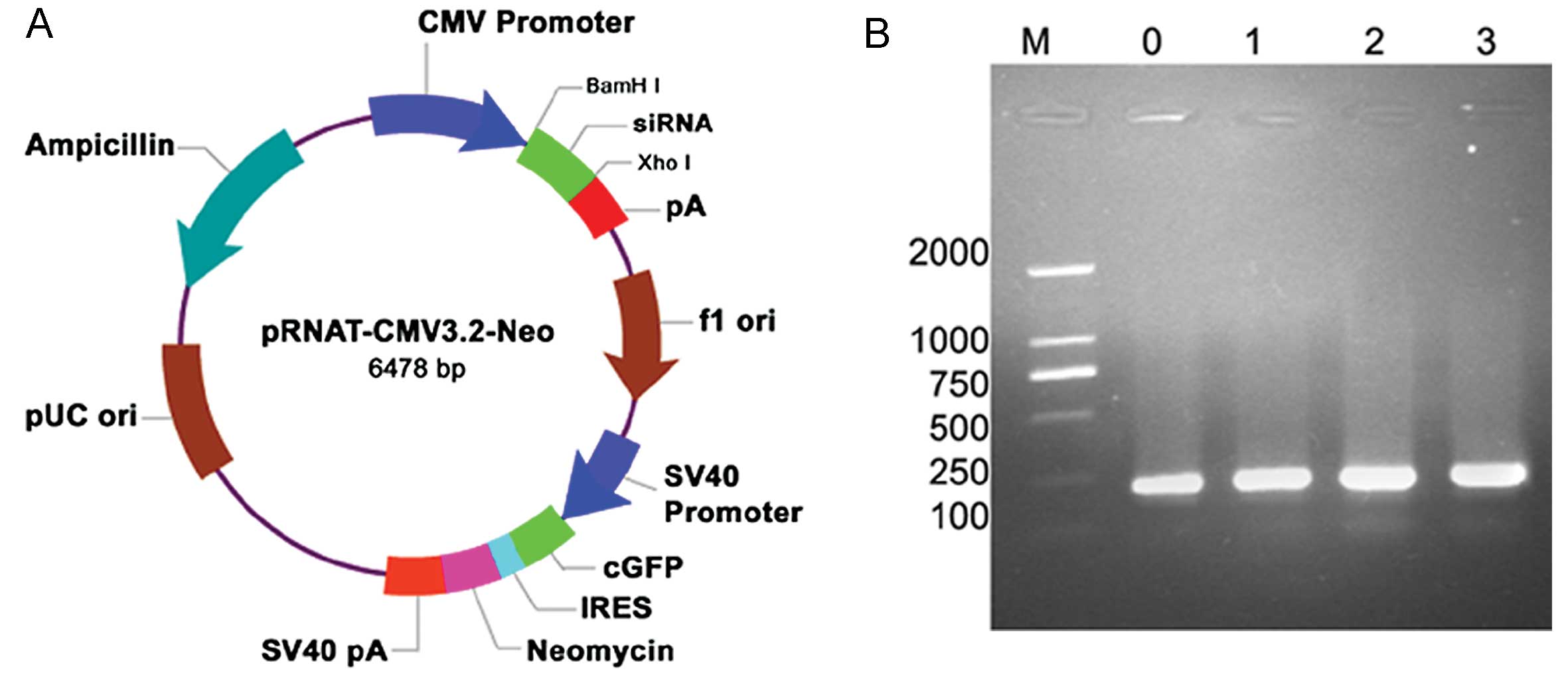
Identification of recombinant
pRNAT-CMV3.2-siRNA by electrophoresis
Positive clones were selected for plasmid
mini-preparation, and the products were digested using BamHI
and XhoI enzymes, forming linear DNA fragments (Fig. 2A). Fragments were inserted into
the pRNAT-CMV3.2 vector and 5 μl of the recombinant
pRNAT-CMV3.2-siRNA plasmid was visualized by 1% agar gel
electrophoresis. The PCR product of interest is 221 bp in length as
it contains the pRNAT-CMV3.2 insert, the forward and reverse
primers, and length of the hairpin DNA fragment. All three pairs of
specific NEDD9 and the unrelated control recombinant clones are
shown in Fig. 2B.
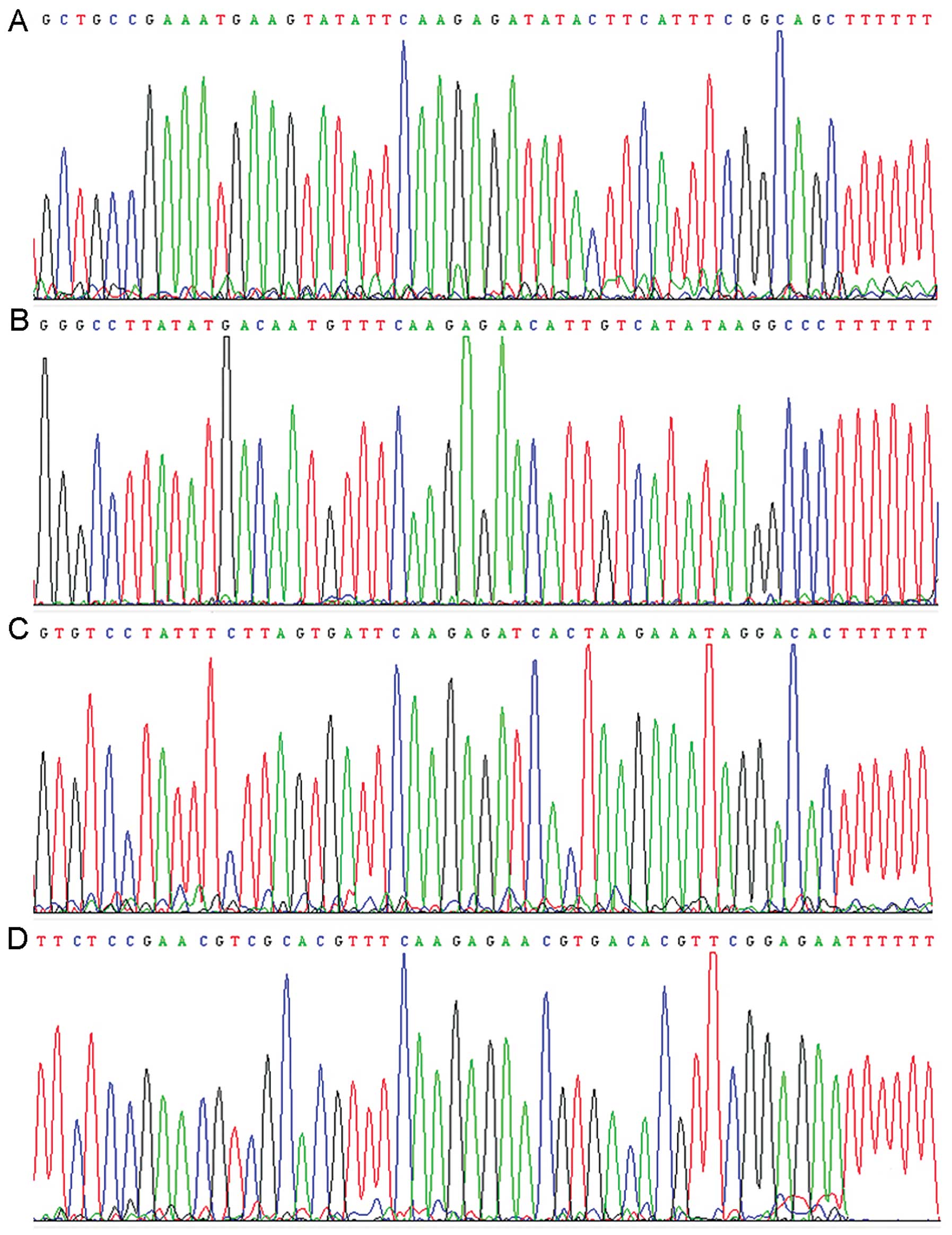
Identification of recombinant
pRNAT-CMV3.2-siRNA by DNA sequencing
The pRNAT-CMV3.2-siRNA cell culture was analyzed by
DNA sequencing. The sequence was in agreement with the GenBank
database (Fig. 3).
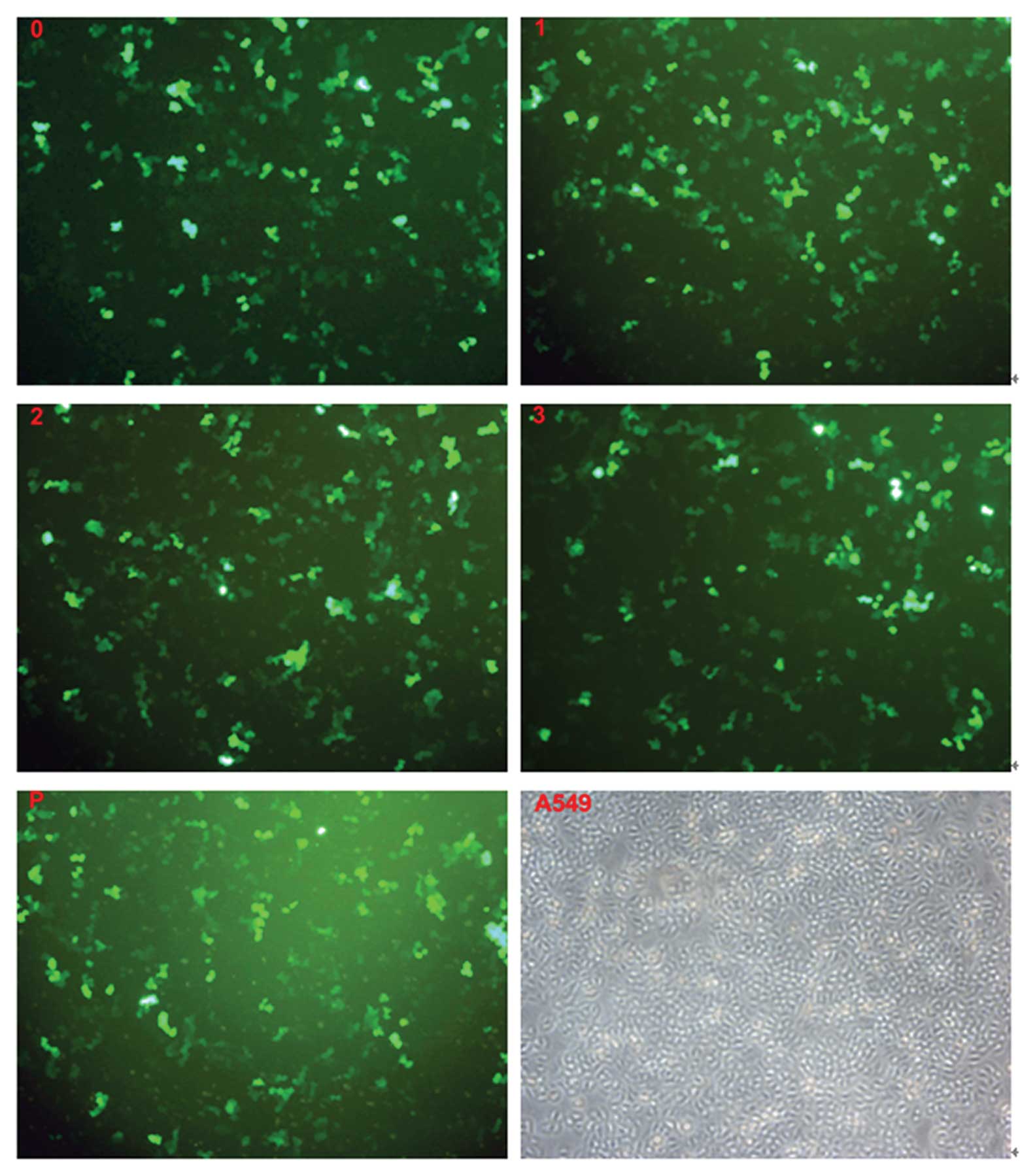
Results of the transfection of cells
The three recombinant pRNAT-CMV3.2-siRNA groups
(transfected with pRNAT-CMV3.2-NE159, pRNAT-CMV3.2-NE193,
pRNAT-CMV3.2-NE648), the unrelated siRNA group (transfected with
pRNAT-CMV3.2-C), and the empty vector plasmid group (transfected
with pRNAT-CMV3.2) all gave widespread expression of cGFP in A549
cells observed under an inverted fluorescence microscope (Fig. 4).
Inhibition of mRNA of the NEDD9 gene
after transfection
A logarithmic chart of the corresponding
concentrations according to the cycle threshold value (Ct) of GAPDH
and NEDD9 resulted in two straight lines. The relevant coefficients
of GAPDH and NEDD9, i.e., straight lines indicated by real-time
quantitative measurement, were −0.993 and −0.999, respectively, and
the gradients were both −3.24. According to the formula
E=10−1/S-1, where E is the amplification efficiency and
S is the rate of standard curve; the amplification efficiency of
the two genes was 100%. The Ct value for the plasmid
pRNAT-CMV3.2-NE648 was lower in the intervention group compared
with the blank group, and the rate of inhibition after correction
was 86%, which was significantly greater than other suppression
rates (P<0.05) (Table II).
| Table IIRelative expression of NEDD9. |
Table II
Relative expression of NEDD9.
| Group |
ΔCtmean | ΔΔCt | Fold (mean ±
SD) |
|---|
| Blank | 7.166 | 0 | 1 |
| pRNAT-CMV3.2 | 7.752 | 0.586 | 0.67±0.05a |
| pRNAT-CMV3.2-C | 7.763 | 0.597 | 0.68±0.17a |
|
pRNAT-CMV3.2-N159 | 9.505 | 2.339 |
0.19±0.02b,c |
|
pRNAT-CMV3.2-N193 | 8.915 | 1.749 |
0.30±0.03b,c |
|
pRNAT-CMV3.2-N648 | 10.185 | 3.019 |
0.11+0.01b,c |
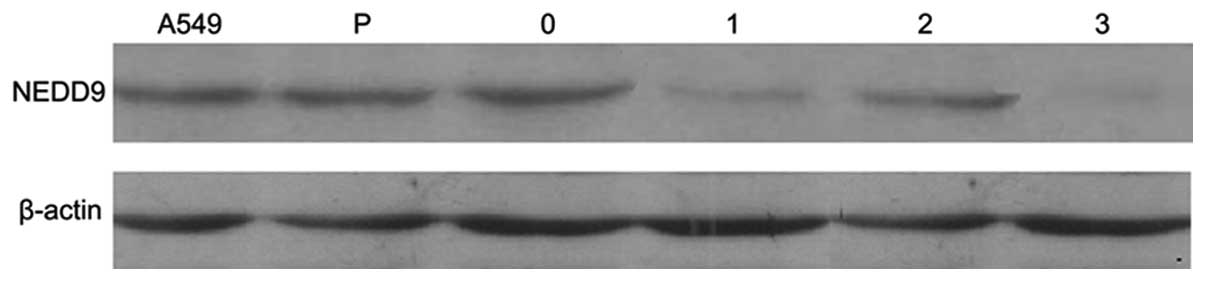
Inhibition of NEDD9 protein expression
after transfection
Western blot analysis indicated that the
experimental groups had low levels of NEDD9 protein expression
while the control group had high levels of expression. There were
no significant differences in p-actin protein expression between
the six groups (Fig. 5).
Discussion
NEDD9 is a member of the CAS (Crk-associated
substrate) protein family. Although it lacks any known enzymatic
function, it contains many functional modules for protein
interaction, leading to its classification as a scaffolding
protein. Because it appears to lack catalytic activity, NEDD9 is
not immediately promising as a target for directed drug
development, unless it is through agents intended to disrupt
protein-protein interactions, or through an siRNA-based approach to
deplete NEDD9 levels globally.
siRNA-mediated knockdown of NEDD9 reduces the number
of cells undergoing mitosis, and leads to cleavage furrow
regression and multinucleation (13,14). If NEDD9-directed drugs or siRNAs
should be developed, it is encouraging that genetic NEDD9-knockout
animals exhibit relatively limited defects, as this implies that
the loss of NEDD9 is well tolerated (15).
As a tool for gene therapy, RNAi technology has been
applied to a number of diseases in medical research. For example,
Lin et al (16)
downregulated cancer-related gene expression by RNAi in human
melanoma cells showing that the knockdown of gene expression
significantly suppressed the abnormal proliferation of melanoma
cells. Wang et al (17)
successfully downregulated the mRNA and protein expression of Bcl-2
in breast cancer MCF27 cells by RNAi, revealing the valuable
application of RNAi technology in basic research.
In this study, we designed and synthesized specific
siRNA sequences targeting NEDD9 and successfully constructed four
vectors, pRNAT-CMV3.2-N159, pRNAT-CMV3.2-N193, pRNAT-CMV3.2-N648
and pRNAT-CMV3.2-C. The human cytomegalovirus (CMV) promoter is one
of the strongest promoters described. Based on the RNA polymerase
II system, the CMV promoter drives higher-level constitutive
expression of genes in a greater variety of mammalian cell lines
compared with RNA polymerase III-based promoters such as U6 and H1.
In this vector, the CMV promoter drives the expression of siRNA and
a SV40 promoter drives the expression of the resistance gene. siRNA
cassettes can be easily inserted into the vectors between the
BamHI and XhoI sites. This vector also carries cGFP
(coral GFP) for convenient tracking of transfection efficiency.
FQ-PCR and western blotting showed substantially decreased mRNA and
protein expression of the NEDD9 gene in the transfected cells,
compared with the control group.
In conclusion, we designed a NEDD9 siRNA-expressing
plasmid and showed that the siRNA expression vector inhibited the
expression of NEDD9 in A549 cells. This study has laid the
foundation for further therapeutic study of NEDD9 inhibition in
lung adenocarcinoma.
Acknowledgements
The authors are grateful to all staff
at the study centre who contributed to this study.
References
|
1.
|
SF LawJ EstojakB WangHuman enhancer of
filamentation 1 (HEF1/NEDD9/CAS-L), a novel p130Cas-like docking
protein, associates with FAK, and induces pseudohyphal growth in
yeastMol Cell Biol163327333719968668148
|
|
2.
|
M MinegishiK TachibanaT SatoStructure and
function of Cas-L, a 105-kD Crk-associated substrate-related
protein that is involved in beta-1 integrin-mediated signaling in
lymphocytesJ Exp
Med18413651375199610.1084/jem.184.4.13658879209
|
|
3.
|
M SinghL CowellS SeoMolecular basis for
HEF1/NEDD9/Cas-L action as a multifunctional co-ordinator of
invasion, apoptosis and cell cycleCell Biochem
Biophys485472200710.1007/s12013-007-0036-317703068
|
|
4.
|
AJ MinnGP GuptaPM SiegelGenes that mediate
breast cancer metastasis to
lungNature436518524200510.1038/nature0379916049480
|
|
5.
|
M NatarajanJE StewartEA GolemisHEF1 is a
necessary and specific downstream effector of FAK that promotes the
migration of glioblastoma
cellsOncogene2517211732200610.1038/sj.onc.120919916288224
|
|
6.
|
M KimJD GansC NogueiraComparative
oncogenomics identifies NEDD9 as a melanoma metastasis
geneCell12512691281200610.1016/j.cell.2006.06.00816814714
|
|
7.
|
A JemalF BrayMM CenterGlobal cancer
statisticsCA Cancer J Clin616990201110.3322/caac.20107
|
|
8.
|
JX ChangF GaoGJ ZhangExpression and
clinical significance of NEDD9 in lung tissuesMed
OncolMarch242012(Epub ahead of
print)10.1007/s12032-012-0213-02012
|
|
9.
|
SF LawYZ ZhangAJ Klein-SzantoCell
cycle-regulated processing of HEF1 to multiple protein forms
differentially targeted to multiple subcellular compartmentsMol
Cell Biol18354035511998
|
|
10.
|
M RyuT KinoshitaM KonishiSegmental
resection of the duodenum including the papilla of Vater for focal
cancer in adenomaHepatogastroenterology4383583819968884299
|
|
11.
|
JA LowellRL RossiJL MunsonPrimary
adenocarcinoma of third and fourth portions of duodenumArch
Surg127557560199210.1001/archsurg.1992.014200500810101349472
|
|
12.
|
IG KaklamanosOF BatheD FranceschiExtent of
resection in the management of duodenal adenocarcinomaAm J
Surg1793741200010.1016/S0002-9610(99)00269-X10737576
|
|
13.
|
EN PugachevaEA GolemisThe focal adhesion
scaffolding protein HEF1 regulates activation of the Aurora-A and
Nek2 kinases at the centrosomeNat Cell
Biol7937946200510.1038/ncb130916184168
|
|
14.
|
D DadkeM JarnikEN PugachevaDeregulation of
HEF1 impairs M-phase progression by disrupting the RhoA activation
cycleMol Biol Cell1712041217200610.1091/mbc.E05-03-023716394104
|
|
15.
|
S SeoT AsaiT SaitoCrk-associated substrate
lymphocyte type is required for lymphocyte trafficking and marginal
zone B cell maintenanceJ
Immunol17534923501200510.4049/jimmunol.175.6.349216148091
|
|
16.
|
JQ YinY WanRNA-mediated gene regulation
system: Now and the futureInt J Mol Med10355365200212239579
|
|
17.
|
YH WangS LiuG ZhangKnockdown of c-Myc
expression by RNAi inhibits MCF-7 breast tumor cells growth in
vitro and in vivoBreast Cancer
Res7220228200510.1186/bcr97515743499
|