Introduction
Unilateral ureteral obstruction (UUO) in the rodent
is the most widely used model of chronic progressive renal disease,
quite similar to that developing in many clinical renal disorders
(1). Tubular atrophy and nephron
loss is the important pathogenic pathway that leads to progressive
reduction in the glomerular filtration rate in chronic renal
disease (2). The reduction in
proximal tubular mass appears to be the primary determinant of
renal parenchymal loss following UUO. In this process, proximal
renal tubules undergo oxidant injury, apoptosis and autophagy with
widespread mitochondrial injury, which results in tubular collapse
and decomposition (3).
Under a low level of injury, autophagy was found to
proceed apoptosis and to play a role in cell survival by clearing
the degradation of long-lived proteins and subcellular organelles
and to re-establish homeostasis (4). When excessive levels of autophagy
are activated due to severe injury, it may be detrimental and
responsible for non-apoptotic cell death (5). Autophagy induced by UUO has a
renoprotective role in the obstructed kidney at an early stage
(6). It has recently been
reported that under the stimulus of persistent UUO, cytokines and
growth factors and oxidative stress may induce proximal tubular
epithelial cell death by excessive autophagy and apoptosis
(7).
Experimental and clinical evidence indicates that
transforming growth factor (TGF)-β1 plays a central role in renal
fibrosis following obstructive nephropathy. Transgenic
overexpression of TGF-β1 localized to the proximal tubule induces
autophagy and then leads to tubular decomposition, peritubular
fibrosis and nephron loss. The process of tubular degeneration in
this model was similar in essential features to tubular
decomposition following UUO (8).
The persistent hypoxia, increasing levels of TGF-β1 and other
cytokines after UUO, stimulate the overproduction of reactive
oxygen species (ROS), which have been previously identified as a
key component to mediate autophagy and apoptosis (9). Inhibition of oxidative stress and
mitochondrial injury can ameliorate tubular injury and renal
interstitial fibrosis in obstructive nephropathy (10). Evidence has shown that selective
oxidant injury to the proximal tubules in response to UUO causes
widespread DNA damage and mitochondrial injury (11). In this condition, the induction of
autophagy may fail to prevent proximal tubule apoptosis, but may
contribute to progressive tubular atrophy.
In the present study, we aimed to ascertain whether
accumulation of reactive oxygen species induces autophagy and
promotes apoptosis, which contribute to renal proximal tubular cell
death, and represents an important mechanism of tubular
decomposition and nephron loss following UUO.
Materials and methods
Animals and experimental protocols
Sprague-Dawley (SD) male rats (weight, 200–230 g)
were subjected to UUO (30 animals) or sham operation (10 animals).
All animal experimental procedures were previously approved by the
Ethics Committee on Laboratory Animal Research at Fujian Medical
University, in accordance with the Guidelines of Experimental
Animals of China. The rats were anesthetized by intraperitoneal
injection of pentobarbital sodium at a dose of 50 mg/kg. In animals
undergoing UUO, the left ureter was ligated with 8-0 nylon, while
in the sham-operated mice the ureter was left undisturbed. The rats
were sacrificed 3, 7 and 14 days after UUO.
Cell culture
All experiments were performed using renal proximal
tubular epithelial cells (NRK-52E) (Chinese Academy of Sciences
Shanghai Institute). Cells were maintained in DMEM that contained
4.5 g/l glucose (Invitrogen, Carlsbad, CA) supplemented with 5%
fetal bovine serum (FBS), at 37°C in 5% CO2.
Materials and reagents
The antibodies used were as follows: anti-LC3-I and
anti-LC3-II (Cell Signaling Technology); anti-Beclin 1 (BD
Biosciences), β-actin (Sigma-Aldrich, St. Louis, MO), anti-NOX4
(Abcam), anti-Bax and anti-Bcl-2 (Santa Cruz Biotechnology, Inc.,
Santa Cruz, CA). The reagents used were as follows: a Trypan Blue
Staining Cell Viability Assay, Terminal deoxynucleotidyl
transferase-mediated dUTP nick translation end labeling (TUNEL)
cell apoptosis detection kit, Hoechst staining kit, and BCA protein
assay kit provided by Beyotime Institute of Biotechnology (Haimen,
China).
Pathological observation of the
kidneys
Kidneys were fixed using 10% formaldehyde overnight
and dehydrated using gradient ethanol. The transparent kidney that
was treated with xylene was embedded in paraffin. The pathological
changes in the tubulointerstitium were observed using Periodic
acid-Schiff (PAS) and Masson’s staining. The pathological changes
in the tubulointerstitium changes were semi-quantitatively observed
using a microscope and scored in a double-blind manner. Ten fields
of the PAS-stained slices that did not contain glomeruli were
randomly observed (×200). The pathological changes were scored
according to the expansion/atrophy of the renal tubules,
inflammatory infiltration and interstitial fibrosis. The scores
were as follows: normal, 0; injured area <25%, 1; injured area
between 25 and 50%, 2; and injured area >50%, 3. Collagen
deposition in the renal interstitium was semi-quantitatively
analyzed using Masson’s staining. Twenty renal section fields were
randomly selected and observed using a microscope (×200). The
semi-quantitative score was obtained by calculating the ratio of
the positive collagen-stained area to the area of the entire field.
The scores were as follows: positive area <2% (normal), 0;
positive area between 2 and 10% (light), 1; positive area between
11 and 20% (moderate), 2; positive area between 21 and 30%
(severe), 3; and positive area >30% (the most severe), 4.
Electron microscopic (EM) analysis
Autophagosomes were evaluated by electron
microscopy. Kidney block samples were fixed in glutaraldehyde and
osmium tetroxide, washed with PBS, dehydrated with a series of
graded ethyl alcohol solutions. After exchange through acetone, the
samples were later embedded in Epon 812. The tissue blocks were
made into ultra-thin sections (70–80 nm) after observation and
positioning, and were double stained by uranium and cobalt
following standard methods. All of the samples were examined using
transmission electron microscopy (TEM) (Philips EM 208).
Detection of SOD, MDA and GSH levels
The kidney was lysed and homogenized, and the
homogenate was centrifuged at 4°C. Levels of superoxide dismutase
(SOD), malonaldehyde (MDA) (Jianchen Bioengineering Institute,
Najing, China) and glutathione (GSH) (Beyotime Institute of
Biotechnology) in the supernatant were detected as per instructions
included in the kit.
Isolation of mitochondria from the
tissue
All steps were carried out at 4°C. Mitochondrial
isolation buffer (Sigma-Aldrich) was added to the tissue of the
renal cortex. The tissue was homogenized and lysated, then
subjected to centrifugation at 1,000 x g for 10 min. The
supernatants were then centrifuged at 3,000 x g for 10 min. The
pellets were resuspended and centrifuged at 1,500 x g for 10 min.
The resulting mitochondrial pellet was resuspended and centrifuged
at 3,000 and 15,000 x g and the final light mitochondrial pellet
was obtained.
TUNEL staining analysis
Detection of apoptosis in the obstructed kidney was
performed following the instructions of the TUNEL assay. Brown
granules in the nucleus or cytoplasm of renal tubular epithelial
cells represented positive apoptotic cells. Two sections were
observed and photographed for each rat. Ten consecutive fields
(×400) were randomly selected, and the nymber of positive cells was
assessed using an image analyzer.
Real-time reverse transcription-PCR
Total RNA from the mitochondrial fractions of the
obstructed renal cortex was extracted using TRIzol reagent
(Gibco-BRL). The sequences of the target gene were obtained from
nucleotides of the National Center for Biotechnology Information
(NCBI). Primers were synthesized by Sangon Co. (Shanghai, China).
The cDNA was obtained using AMV reverse transcriptase. The profile
of reverse transcription was as follows: 95°C for 15 sec, followed
by 60°C for 1 min. PCR was performed using the following primers:
NOX4, 5′-TCAACTGCAGCCTGATCCTTT-3′ (sense) and
5′-TCTGTGATCCGCGAAGGTAAG-3′ (antisense). The relative expression of
NOX4 was obtained as follows: Expression of NOX4/expression of
β-actin.
Immunoblot analyses
The kidney was homogenized in boiling lysis buffer
(50 mM Tris, pH 7.4, 1% sodium deoxycholate and 0.1% SDS). Cultured
cells were washed with ice-cold PBS and lysed in RIPA lysis buffer.
The concentration of proteins was detected by a BCA protein assay
kit. Immunoblot analysis was performed in a routine manner.
NADPH oxidase activity
NADPH oxidase activity of mitochondrial fractions in
the renal cortex was measured by the luminol
(5-amino-2,3-dihydro-1,4-phthalazinedione; Sigma-Aldrich)
technique. Samples were homogenized and centrifuged. The membrane
fraction was lysated and the concentration of proteins was
quantified. β-NADH and luminol in dimethyl sulfoxide were
incorporated, and NADPH oxidase activity was evaluated by
fluorescence emission. The values were expressed as relative
fluorescence units per microgram of protein.
Hoechst 33258 staining
Apoptosis in the cultured cells was evaluated by
staining the nuclei with Hoechst staining to determine those with
condensed chromatin. Human renal proximal tubular epithelial cells
(HRPTEpiCs) were cultured in 6-well flat-bottom plates. After
exposure to 0.1 mM H2O2 for 12 h, cells were
fixed, washed with PBS, and then stained with Hoechst 33258 (0.5
ml/well) for 5 min. Cells were observed under fluorescence
microscopy with a 350 nm excitation filter and 460 nm emission
filter.
Measurement of mitochondrial membrane
potential
To measure mitochondrial depolarization in NRK-52E
cells, a JC-1 probe was employed. JC-1 exhibits potential-dependent
accumulation in mitochondria, which is indicated by a fluorescence
emission shift from 529 to 590 nm. Mitochondrial depolarization is
indicated by an increase in the ratio of green:red fluorescence
intensity as described previously (12).
Statistical analysis
All experiments were performed in at least
triplicate. Data are presented as means ± the standard deviation
(SD) and analyzed using Statistical Product and Service Solutions
(SPSS) version 13.0. Between-group differences were analyzed using
one-way analysis of variance (ANOVA) and Bonferroni test. P<0.05
was considered as the level of significance.
Results
Tubular atrophy and interstitial fibrosis
following unilateral ureteral obstruction
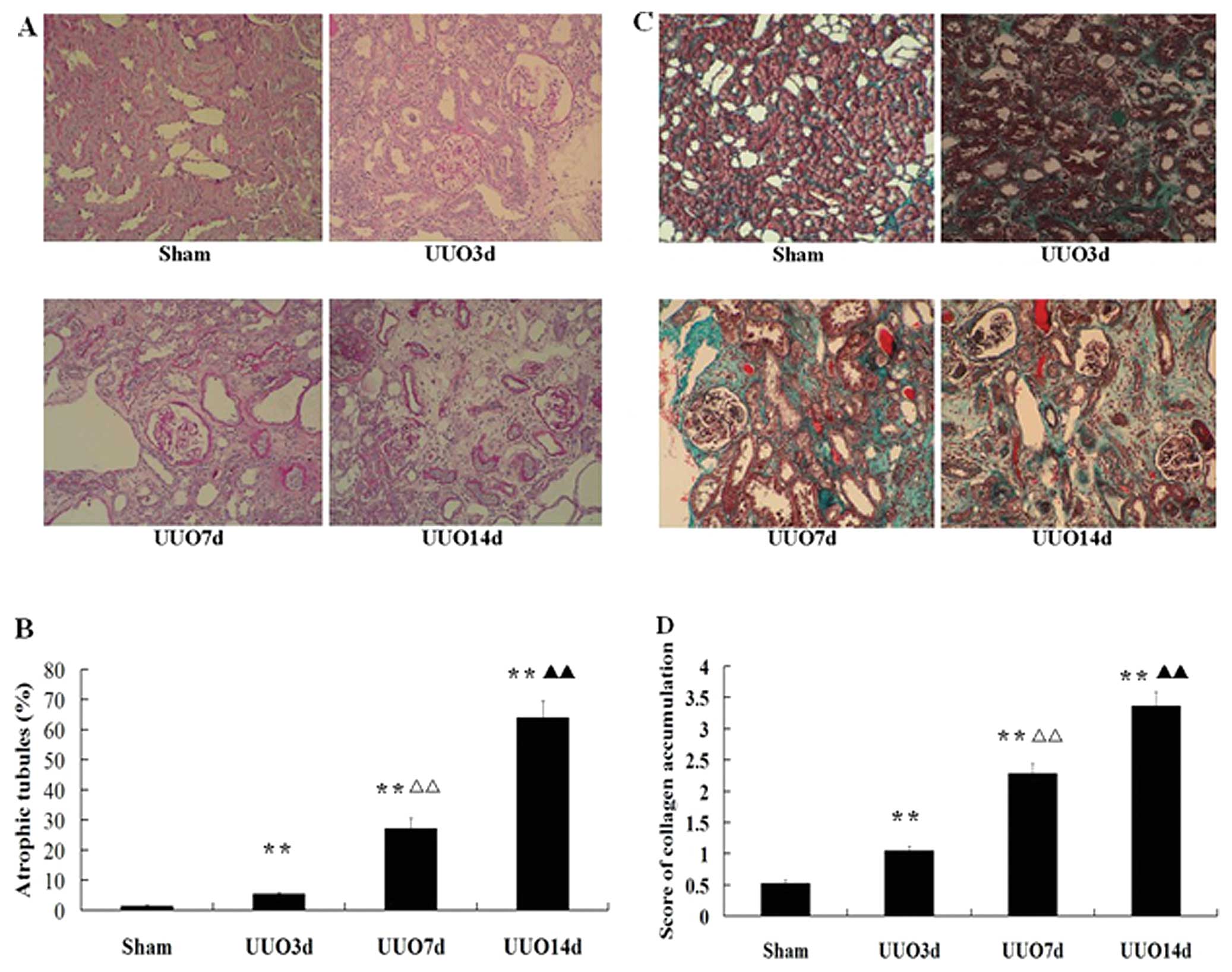
Periodic acid-Schiff staining was performed to
measure tubular atrophy. The proximal tubules were identified by
the presence of brush border. In sham-operated kidneys, rare
tubular atrophy was observed. Proximal tubular injury was evidenced
by the loss of brush border at 3 days, and tubule dilation was
commonly noted at 7 days with mild to moderate tubular atrophy
after UUO. A minimum of 1,000 tubules were examined for tubular
atrophy in each sample. At 14 days, tubular atrophy and wrinkling
of the TBM became more predominant (Fig. 1A and B).
Masson’s trichrome staining also showed matrix
accumulation and interstitial fibrosis in a time-dependent manner
in the obstructed kidney after UUO (Fig. 1C). In comparison to the
sham-operated groups, the renal cortex showed higher collagen
accumulation in the expanded interstitium after 14 days of UUO
(Fig. 1D).
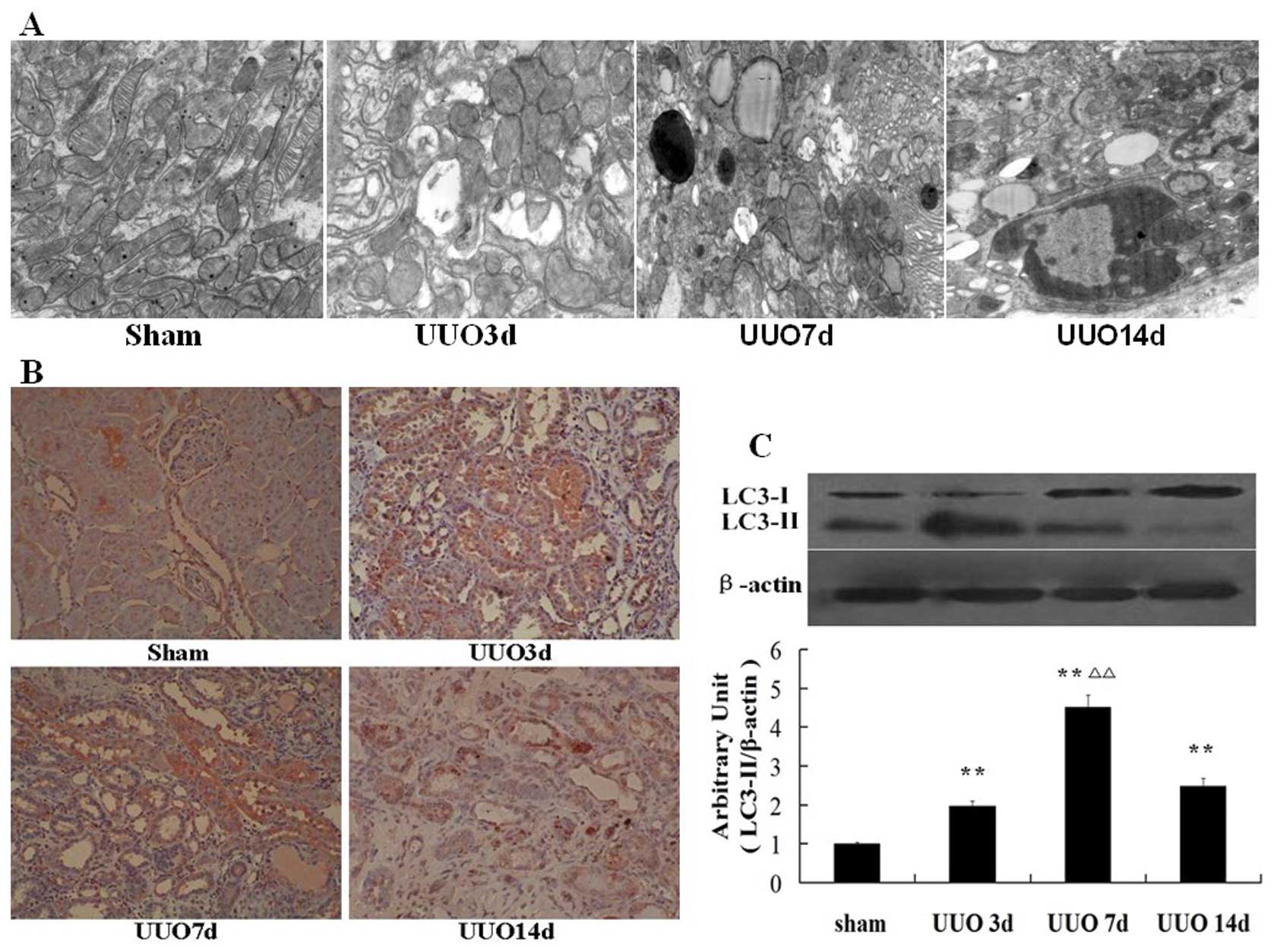
Autophagy was induced in the obstructed
renal tubules
Evidence suggests that tubular autophagy follows
UUO. To confirm the autophagy of proximal tubular cells, we first
examined autophagy by TEM. Rare autophagic vacuoles were detected
in the tubular cells of the sham operation rats. However, complete
urinary tract obstruction led to an increased level of autophagy
(Fig. 2A). Autophagic vacuoles
including autophagosomes and autophagolysosomes were markedly
increased in the proximal tubular cells at days 3 and 7 after UUO.
Double membrane vacuoles containing electron-dense material,
degenerating cytoplasmic organelles and cytosol, mitochondria with
loss of visible cristae were frequently observed in the tubules.
However, these autophagic vacuoles significantly decreased at day
14 after UUO.
To further confirm autophagy, we examined the
expression of microtubule-associated protein 1 light-chain 3
(LC3)-II. The conversion of LC3-I to LC3-II is considered a
standard marker of autophagy (13). Using immunohistochemical staining,
we also demonstrated that the expression of LC3-II to LC3-I
increased steadily up to day 7, then declined thereafter (Fig. 2B). In agreement with the
immunohistochemical staining, western blotting showed that the
contents of LC3-II to LC3-I markedly increased at days 3 and 7, and
then decreased towards basal levels at day 14 after UUO (Fig. 2C).
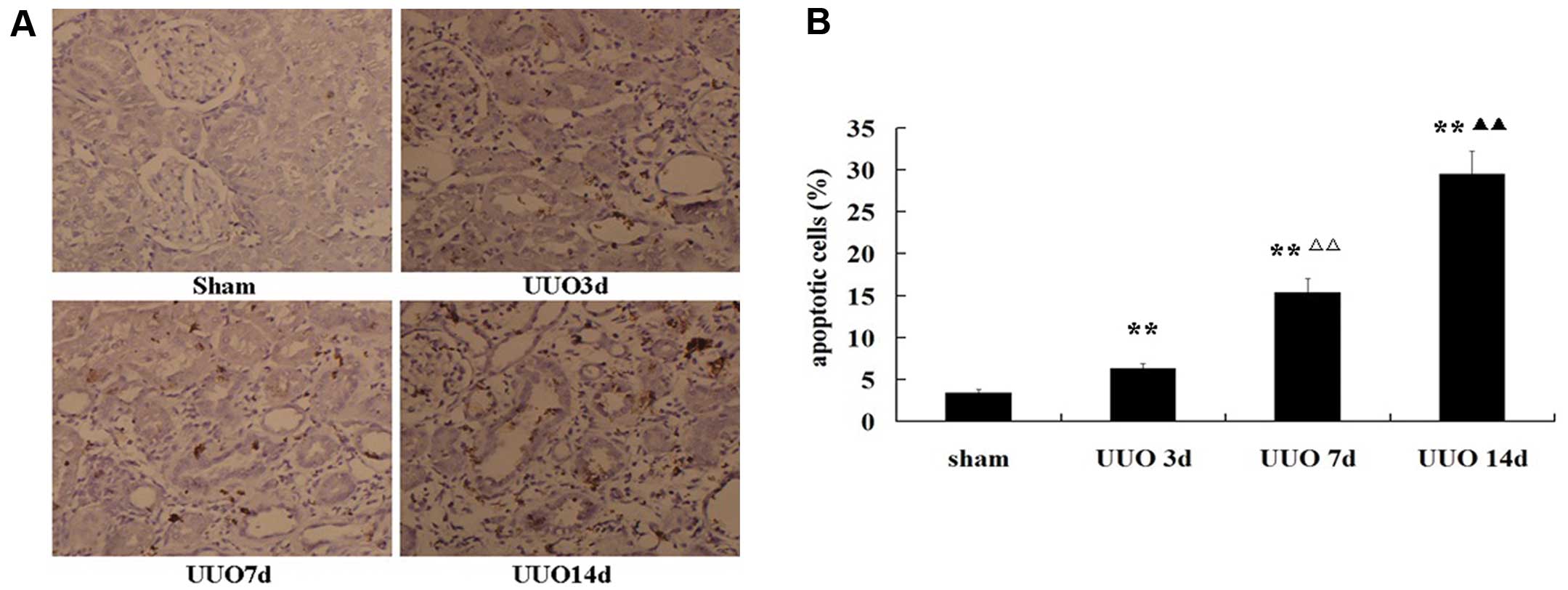
Increased apoptotic cell death following
UUO
In the sham-operated kidneys, rare apoptotic tubular
cells were observed. In the obstructed tubules, the number of
apoptotic cells steadily increased from day 3 to 14. The number of
TUNEL-positive cells increased in a time-dependent manner after UUO
(Fig. 3). Notably, in Fig. 2A, at 14 days after UUO, apoptotic
bodies were also noted. These results indicated the coexistence of
autophagy and apoptosis with tubular atrophy in the obstructed
proximal tubules. However, autophagy was induced earlier than
tubular apoptosis and decreased at day 17.
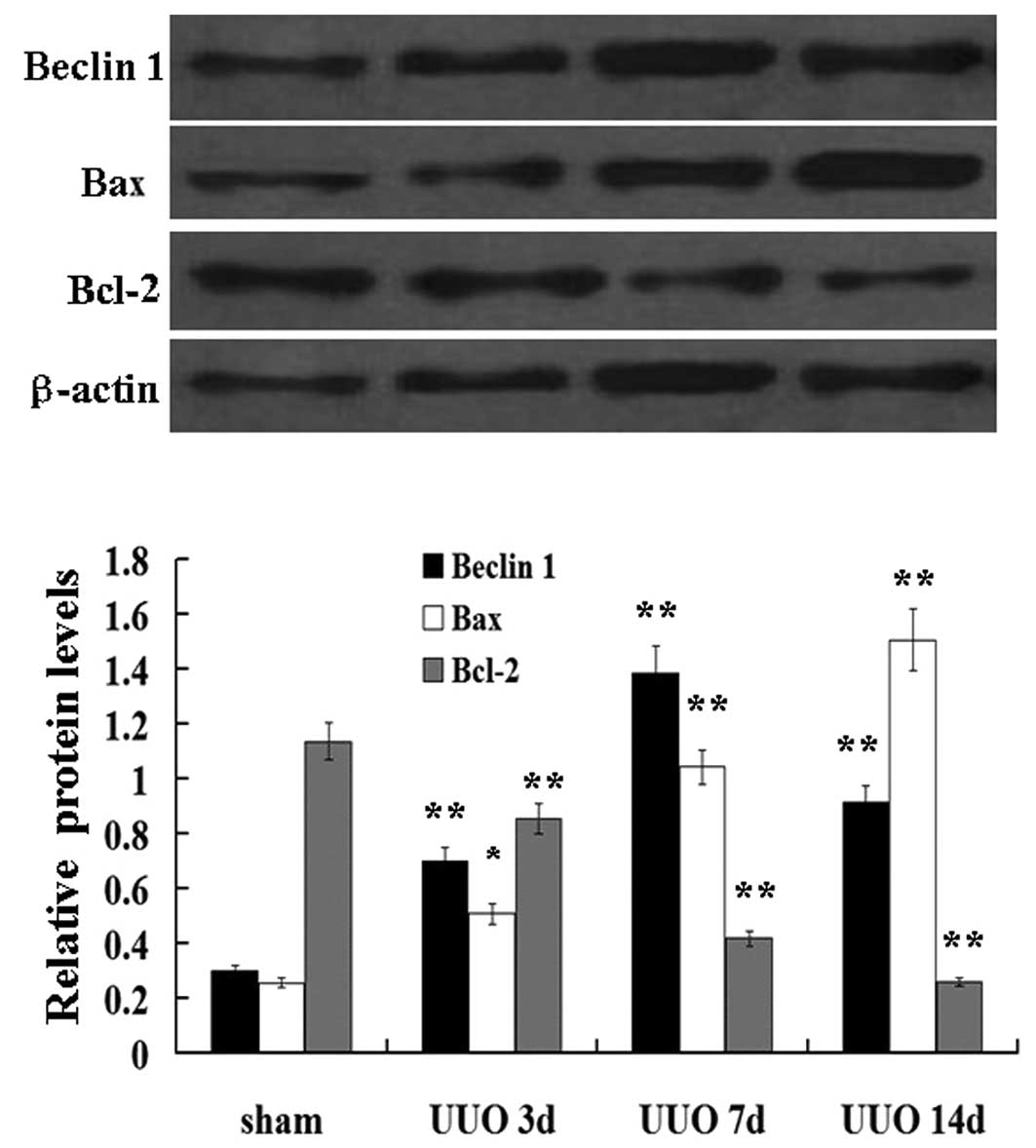
Expression of Bcl-2, Bax and Beclin 1
following UUO
Bcl-2 is known to play an anti-apoptotic role in
renal tubule damage (14). In
this UUO model, immunoblot analysis demonstrated that levels of
Bcl-2 protein were modestly increased at day 7, and markedly
decreased at day 14. However, the proapoptotic (Bax) protein was
not significantly upregulated in the obstructed kidney after 14
days. It is known that, Beclin 1 is involved in the initiation of
autophagosome formation by forming a multiprotein complex (4). We also confirmed the increased
expression of interacting protein Beclin 1 by immunoblot analysis.
Beclin 1 protein expression was also significantly increased 7 days
after UUO (Fig. 4).
Levels of MDA and antioxidant enzymes in
the obstructed kidney
Oxidative stress has been implicated as one of the
major underlying mechanisms behind the pathogenesis of obstructive
uropathy, which has been demonstrated to be related with activation
of tubular autophagy and apoptosis (15). MDA is the natural product of lipid
peroxidation. The concentration of MDA increases under conditions
of oxidative stress. Antioxidant supplementation protects against
the progression of renal fibrosis. SOD protects against
superoxide-mediated cytotoxicity by catalyzing
O2− to form H2O2. The
GSH antioxidant system is foremost among the cellular protective
mechanisms. Thus, we determined the level of MDA, SOD and GSH in
the obstructed kidney. The level of MDA increased while the levels
of SOD and GSH decreased in a time-dependent manner in the
obstructed kidney (Table I).
 | Table ILevels of MDA and antioxidant enzymes
in the obstructed kidney. |
Table I
Levels of MDA and antioxidant enzymes
in the obstructed kidney.
| Group | GSH (g/g) | SOD (U/mg) | MDA (nmol/mg) |
|---|
| Sham | 65.21±6.02 | 502.40±47.55 | 8.03±0.45 |
| UUO 3d | 51.51±4.78a |
411.77±38.13a | 16.19±1.48a |
| UUO 7d |
37.99±3.45a,b |
325.93±26.12a,b |
23.09±2.01a,b |
| UUO 14d |
31.88±2.71a,b |
256.32±22.79a,c |
28.59±2.64a,c |
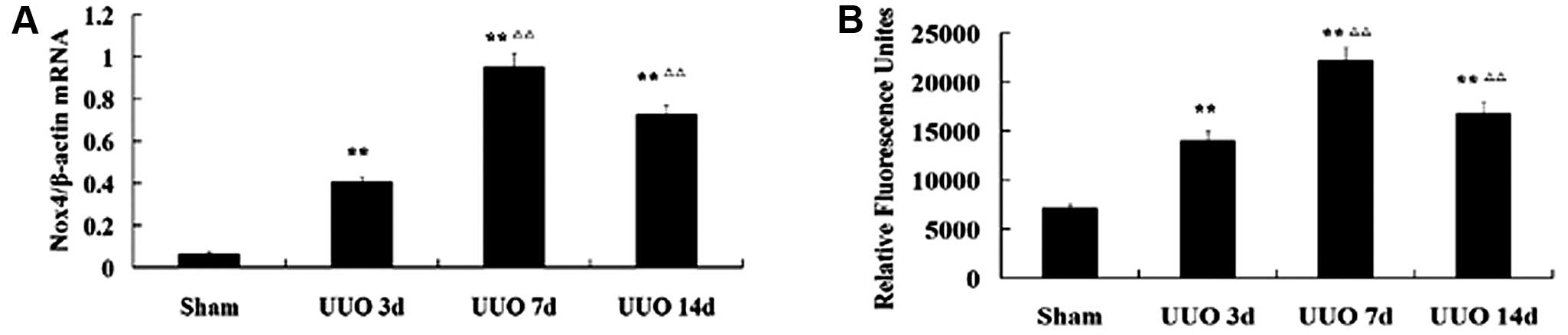
NOX4 expression and NADPH oxidase
activity in mitochondrial fractions of the renal cortex following
UUO
NOX4 is primarily localized in mitochondria by which
NOX4 stimulates cell dysfunction and increased cell death (16). The NOX4 mRNA expression was
significantly elevated in mitochondrial fractions beginning 3 days
after obstruction, and reached a peak on day 7. Parallel to this,
NADPH oxidase activity was notably enhanced in the renal cortex 7
days after UUO (Fig. 5).
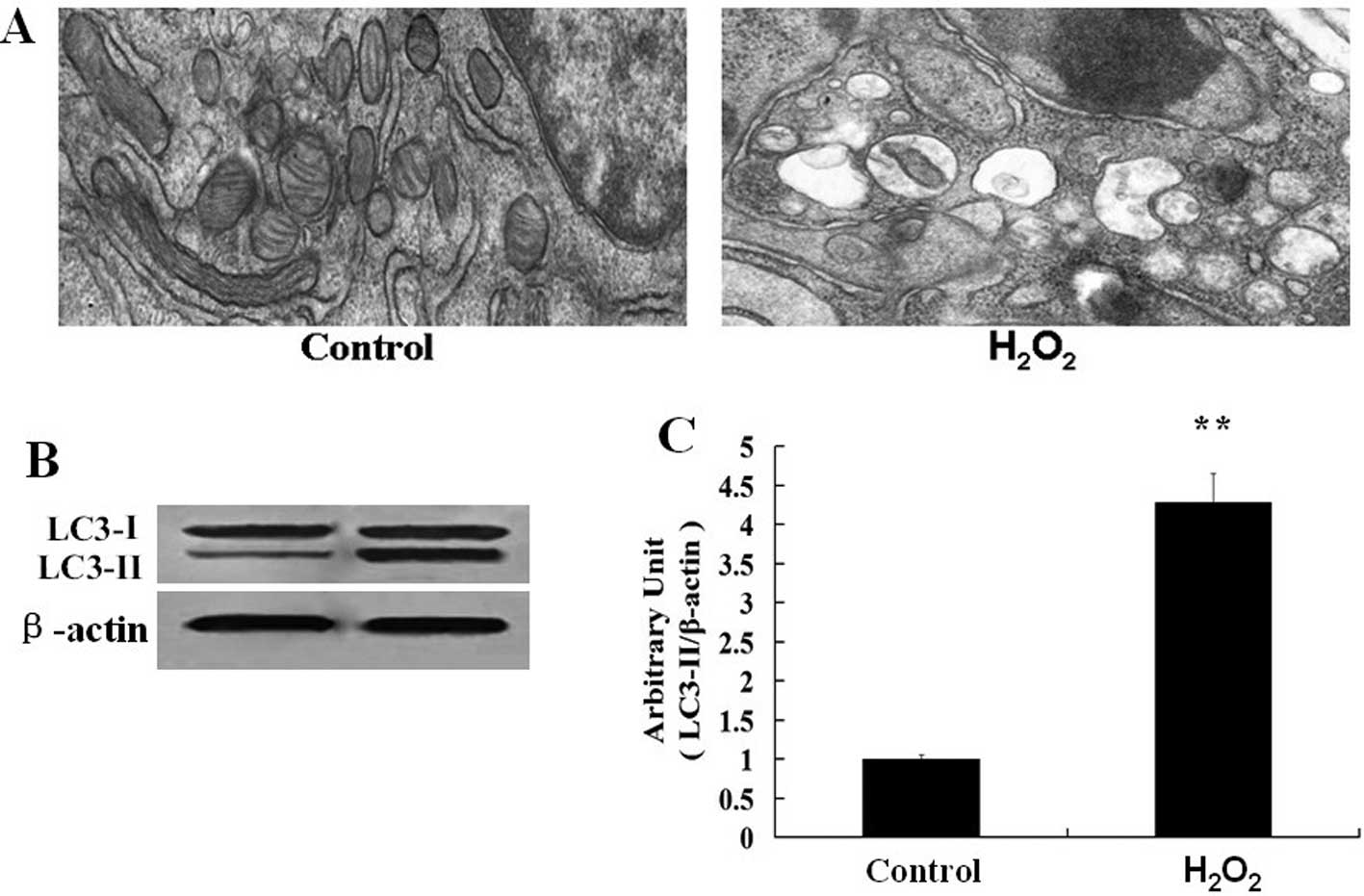
H2O2 induces
parallel apoptosis and autophagy in NRK-52E cells
NRK-52E cells were incubated in the control medium
or medium containing H2O2 for 12 h. We used
electron microscopy (EM) to image the autophagic vacuoles in
NRK-52E cells. As shown in Fig.
6A, in control cell nuclei and mitochondria were well preserved
structures with normal morphology. In contrast, abundant autophagic
vacuoles and abnormal mitochondria with dark cristae and condensed
matrix were noted in the cells cultured for 12 h with 0.1 mM
H2O2. Notably, some cells showed nuclei with
condensed chromatin or fragmented apoptotic bodies under TEM
(Fig. 6A). The conversion of
LC3-I to LC3-II by lipd phosphatidylethanolamine (PE) conjugation
in autophagy during autophagosome formation is one of the
estabished steps (4). Western
blot analysis indicated that the conversion of LC3-I to LC3-II
increased after exposure to H2O2 for 12 h
(Fig. 6B and C).
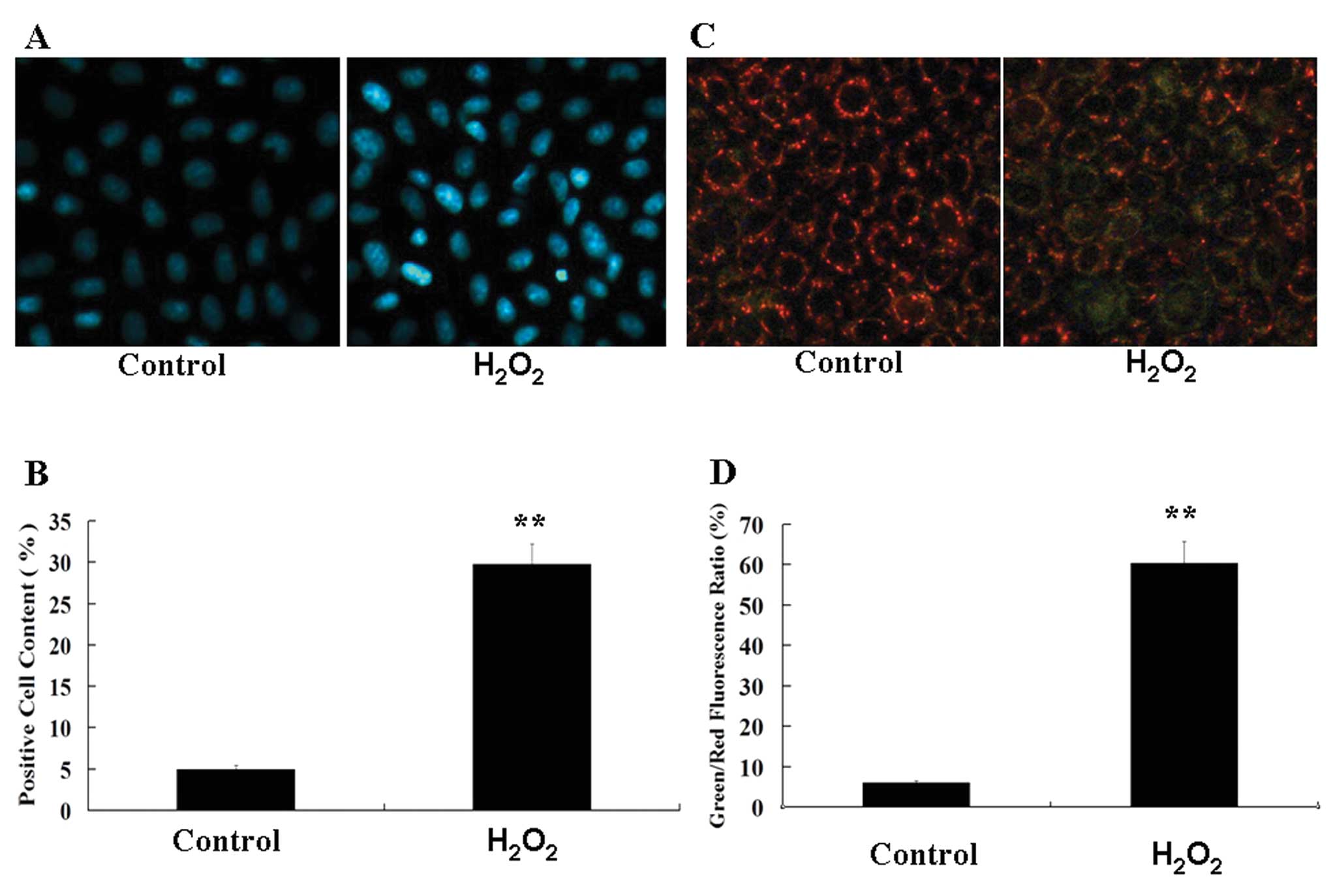
The percentage of apoptotic cells was visualized by
Hoechst staining. Compared with the control, the number of positive
cells markedly increased after exposed to
H2O2 (Fig. 7A
and B). Reduction in the mitochondrial membrane potential (ΔΨm)
and increase in mitochondrial membrane permeability correlated with
apoptosis. Red fluorescence indicates cells with a stable ΔΨm,
while green fluorescence indicates cells with a low ΔΨm. When
NRK-52E cells were exposed to H2O2 for 12 h,
the relative ratio of green fluorescence increased compared with
the untreated control cells, indicating increased number of cells
undergoing apoptosis (Fig. 7C and
D).
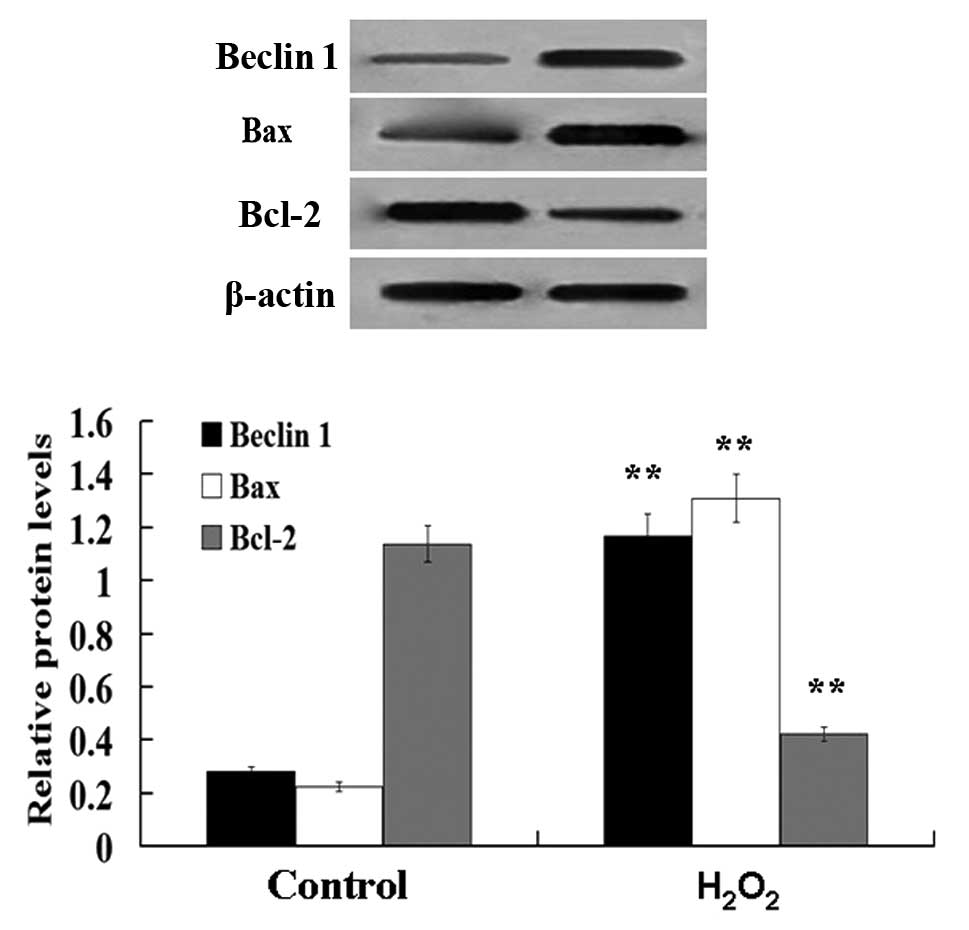
As a well-known anti-apoptotic gene, Bcl-2 is
involved in the regulation of autophagy and apoptosis. Evidence
shows that Bcl-2 inhibits Beclin 1-dependent autophagy by
interacting with Beclin 1. Overexpression of Bcl-2 significantly
inhibited H2O2-induced formation of
autophagic vacuoles and suppressed the mitochondrial fragmentation
(13). Immunoblot analysis
demonstrated that H2O2 downregulated
anti-apoptotic (Bcl-2) and stimulated pro-apoptotic (Bax) proteins,
and also increased Beclin 1 protein expression (Fig. 8).
Discussion
Surgical UUO in a variety of experimental animals
has been used as a model of progressive chronic kidney. Previous
studies have shown that epithelial-mesenchymal transition (EMT)
plays an important role in nephron loss and interstitial fibrosis
obstructive uropathy (1). Recent
evidence suggests another pathway for functional nephron loss in
UUO, which is proximal tubular injury and rapid formation of
atubular glomeruli (17). The
formation of atubular glomeruli proceeded to total decomposition of
tubular cells as a result of autophagy and apoptosis. It is well
known that apoptosis is increased in obstructed renal tubules
(18). However, others recently
reported that autophagy contributes to excessive cell death and
tubular atrophy is another potential mechanism of nephron loss in
obstructive uropathy (7).
Autophagy can also act as a mechanism of cell survival, but it can
lead to cell death in response to persistent stress (19). Under the stimulus of persistent
UUO, the induction of autophagy fails to prevent progressive
destruction of the proximal tubular mass (7). Transgenic overexpression of TGF-β1
localized to the proximal tubule also induces extensive autophagy,
and the autophagocytic process culminates in total disconnection of
cells from the TBM and tubular decomposition. In the present study,
we showed that autophagy and apoptosis proceeded by proximal tubule
injury contributed to tubular atrophy and renal fibrosis (8). However, autophagy increased within
the first week and then declined, while apoptosis continued to
increase through 2 weeks. Thus, the induction of autophagy in
proximal tubules failed to prevent apoptosis, but promoted
apoptosis in this UUO model.
Nephron loss in UUO has been associated with
increased free radical reactions and localized oxidant damage in
proximal tubules (10,18). Mitochondria are the major source
of ROS. NOX4 is the major NADPH oxidase isoform expressed in renal
cells and is primarily localized in mitochondria. It plays an
important role in mediating ROS generation (10,20). In this study, NOX4 expression
increased significantly in mitochondrial fractions of the
obstructed renal cortex during UUO. The mitochondria-rich proximal
tubules appeared to be the most susceptible to obstructive injury.
Electron microscopy showed that mitochondria were of an increased
size and contained dilated cristae and large spaces in their
interiors after UUO. The increased ROS may also be due to impaired
antioxidant capacity, such as an increased level of MDA and reduced
activity of SOD and GSH of the obstructed renal cortex. Thus,
oxidative stress in the renal cortex increased in response to
UUO.
Mitochondrial oxidative stress may contribute to the
decline in mitochondrial functions associated with the collapse of
mitochondrial membrane potential and further transient increase in
ROS generation. These events may result in autophagy, apoptosis or
necrosis (21). Autophagy is one
of the defense mechanisms protecting cells against oxidative
stress. However, persistent enhanced oxidative stress can induced
parallel apoptotic cell death and autophagic cell death (22). In this condition, autophagy may
cooperate with the apoptotic machinery by acting upstream of
apoptosis, converging with the apoptotic pathway. In this UUO
model, autophagy occurred earlier than apoptosis, but it did not
inhibit apoptosis. Thus, autophagy and apoptosis are coordinated
processes which contribute to tubular atrophy.
Oxidative stress plays an important role in various
renal pathological conditions. ROS are generated at high levels to
induce oxidation and damage DNA, mitochondria, endoplasmic
reticulum and other organelles or macromolecules (13,23). Reactive oxygen species (ROS) have
been shown to mediate cellular responses including EMT or
mitochondria-dependent apoptosis and autophagy in various types
cells (13,24). Previously, it was observed that
ROS mediated TGF-β1-induced autophagy, which promoted apoptosis in
renal tubular epithelial cells (19). Rasbach and Schnellmann (25) showed that exposure to oxidants
decreased mitochondrial function and biogenesis, which was
associated with autophagy in RPTCs (25). In the present study, we also
demonstrated that hydrogen peroxide (H2O2)
induced parallel autophagy and mitochondria dysfunction-mediated
apoptosis. However, it is still unclear whether
H2O2-induced autophagy contributes to
apoptosis in renal tubular epithelial cells.
It has recently been shown that Bcl-2 family
proteins, key modulators of apoptosis, represent a point of
convergence of the autophagic and apoptotic processes. The
anti-apoptotic proteins, such as Bcl-2, Bcl-xL and Bcl-B have been
shown to inhibit autophagy by binding to Beclin 1, a critical
protein involved in initiation of autophagosome formation (13,26,27). In this UUO model, we showed that
pro-apoptotic Bcl-2 family protein Bax increased while
anti-apoptotic protein Bcl-2 decreased in a time-dependent manner.
Notably, at day 7, Beclin 1 was significantly enhanced. These
results indicate that downregulation of Bcl-2 decreased its
capacity to interact with Beclin 1 and failed to suppress
autophagic cell death. Due to oxidative stress, the mitochondrial
dysfunction by accumulation of intracellular ROS has been
associated with autophagy and apoptosis (13). As a well-known anti-apoptotic
gene, overexpression of Bcl-2 stabilized mitochondria, blocked the
classic apoptotic pathways, affected binding with Beclin 1 and
inhibited autophagy (13,28). In vitro, our results showed
that H2O2 treatment increased the expression
of Beclin 1 protein and decreased the expression of Bcl-2, and also
induced mitochondria dysfunction and autophagy. However, under the
condition of oxidative stress, the insight into the interplay
between apoptosis and autophagy needs further investigation.
In summary, we demonstrated that both autophagy and
apoptosis occurred in the tubular epithelial cells following
obstructive uropathy, associated with oxidative stress. Under the
stimulus of persistent UUO, the induction of autophagy in the
proximal tubule did not play a protective mechanism, but
contributed to cell death. Further understanding of the mechanism
of the relationship between autophagy and apoptosis of tubular
epithelial cells in obstructive uropathy may facilitate the
development of therapeutic strategies to prevent end-stage renal
disease.
Acknowledgements
This study was supported in part by
the Nature Science Foundation from Fujian Provincial Department of
Science and Technology (no. 2011J01159), in part by an innovatory
research fund (no. 2011-CX-10) from the Department of Health of
Fujian Province, and partly by a Key Research Fund JA12136 from
Education Department of Fujian Province of China.
References
|
1
|
Yang JW and Liu YH: Blockage of tubular
epithelial to myofibroblast transition by hepatocyte growth factor
prevents renal interstitial fibrosis. J Am Soc Nephrol. 13:96–107.
2002.PubMed/NCBI
|
|
2
|
Zeisberg M and Neilson EG: Mechanisms of
tubulointerstitial fibrosis. J Am Soc Nephrol. 21:1819–1834. 2010.
View Article : Google Scholar
|
|
3
|
Forbes MS, Thornhill BA and Chevalier RL:
Proximal tubular injury and rapid formation of atubular glomeruli
in mice with unilateral ureteral obstruction: a new look at an old
model. Am J Physiol Renal Physiol. 301:F110–F117. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nishida K, Yamaguchi O and Otsu K:
Crosstalk between autophagy and apoptosis in heart disease. Circ
Res. 103:343–351. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Luo S and Rubinsztein DC: Apoptosis blocks
Beclin 1-dependent autophagosome synthesis: an effect rescued by
Bcl-xL. Cell Death Differ. 17:268–277. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kim WY, Nam SA, Song HC, et al: The role
of autophagy in unilateral ureteral obstruction rat model.
Nephrology. 17:148–159. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li L, Zepeda-Orozco D, Black R and Lin FM:
Autophagy is a component of epithelial cell fate in obstructive
uropathy. Am J Pathol. 176:1767–1778. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Koesters R, Kaissling B, Lehir M, et al:
Tubular overexpression of transforming growth factor-beta1 induces
autophagy and fibrosis but not mesenchymal transition of renal
epithelial cells. Am J Pathol. 177:632–643. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ferraro E and Cecconi F: Autophagic and
apoptotic response to stress signals in mammalian cells. Arch
Biochem Biophys. 462:210–219. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
García IM, Altamirano L, Mazzei L, et al:
Role of mitochondria in paricalcitol-mediated cytoprotection during
obstructive nephropathy. Am J Physiol Renal Physiol.
302:F1595–F1605. 2012.PubMed/NCBI
|
|
11
|
Forbes MS, Thornhill BA, Minor JJ, Gordon
KA, Galarreta CI and Chevalier RL: Fight-or-flight: murine
unilateral ureteral obstruction causes extensive proximal tubular
degeneration, collecting duct dilatation, and minimal fibrosis. Am
J Physiol Renal Physiol. 303:F120–F129. 2012. View Article : Google Scholar
|
|
12
|
Xu YF, Ruan SW, Xie H and Lin JM: Role of
LOX-1 in Ang II-induced oxidative functional damage in renal
tubular epithelial cells. Int J Mol Med. 26:679–690.
2010.PubMed/NCBI
|
|
13
|
Zhang H, Kong X, Kang J, Su J, Li Y, Zhong
JT and Sun LK: Oxidative stress induces parallel autophagy and
mitochondria dysfunction in human glioma U251 cells. Toxicol Sci.
110:376–388. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Miyajima A, Chen J, Lawrence C, et al:
Antibody to transforming growth factor-beta ameliorates tubular
apoptosis in unilateral ureteral obstruction. Kidney Int.
58:2301–2313. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Docherty NG, O’Sullivan OE, Healy DA,
Fitzpatrick JM and Watson RWG: Evidence that inhibition of tubular
cell apoptosis protects against renal damage and development of
fibrosis following ureteric obstruction. Am J Physiol Renal
Physiol. 290:F4–F13. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ago T, Kuroda J, Pain J, Fu C, Li H and
Sadoshima J: Upregulation of Nox4 by hypertrophic stimuli promotes
apoptosis and mitochondrial dysfunction in cardiac myocytes. Circ
Res. 106:1253–1264. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chevalier RL and Forbes MS: Generation and
evolution of atubular glomeruli in the progression of renal
disorders. J Am Soc Nephrol. 19:197–206. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chevalier RL: Pathogenesis of renal injury
in obstructive uropathy. Curr Opin Pediatr. 18:153–160. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xu YF, Yang SY, Huang JY, Ruan SW, Zheng Z
and Lin JM: TGF-β1 induced autophagy and promoted apoptosis in
renal tubular epithelial cells. Int J Mol Med. 29:781–790.
2012.
|
|
20
|
Kuroda J, Ago T, Matsushima SJ, Zhai PY,
Schneiderb MD and Sadoshim J: NADPH oxidase 4 (Nox4) is a major
source of oxidative stress in the failing heart. Proc Natl Acad Sci
USA. 107:15565–15570. 2010.PubMed/NCBI
|
|
21
|
Martinet W and De Meyer GR: Autophagy in
atherosclerosis: a cell survival and death phenomenon with
therapeutic potential. Circ Res. 104:304–317. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Addabbo F, Montagnani M and Goligorsky MS:
Mitochondria and reactive oxygen species. Hypertension. 53:885–892.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Periyasamy-Thandavan S, Jiang M, Wei Q,
Smith R, Yin XM and Dong Z: Autophagy is cytoprotective during
cisplatin injury of renal proximal tubular cells. Kidney Int.
74:631–640. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rhyu DY, Yang YQ, Ha H, Lee GT, Song JS,
Uh ST and Lee HB: Role of reactive oxygen species in
TGF-beta1-induced mitogen-activated protein kinase activation and
epithelial-mesenchymal transition in renal tubular epithelial
cells. J Am Soc Nephrol. 16:667–675. 2005. View Article : Google Scholar
|
|
25
|
Rasbach KA and Schnellmann RG: Signaling
of mitochondrial biogenesis following oxidant injury. J Biol Chem.
25:2355–2362. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Robert G, Gastaldi C, Puissant A, et al:
The anti-apoptotic Bcl-B protein inhibits BECN1-dependent
autophagic cell death. Autophagy. 8:1–13. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Oberstein A, Jeffrey PD and Shi Y: Crystal
structure of the Bcl-XL-Beclin 1 peptide complex: Beclin 1 is a
novel BH3-only protein. J Biol Chem. 282:13123–13132. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pattingre S, Tassa A, Qu X, Garuti R,
Liang XH, Mizushima N, et al: Bcl-2 antiapoptotic proteins inhibit
Beclin 1-dependent autophagy. Cell. 122:927–939. 2005. View Article : Google Scholar : PubMed/NCBI
|