Introduction
Retinitis pigmentosa (RP) is one of the leading
causes of vision impairment and blindness affecting millions of
people worldwide (1,2). The Royal College of Surgeons (RCS)
rat is a widely used animal model to study retinal degeneration
diseases and the process of retinal remodeling; it inherits a null
mutation in MER, a member of the Axl/Mer/Tyro3 receptor tyrosine
kinase family gene (MERTK) of the retinal pigment epithelium (RPE)
(3). This mutation results in RPE
dysfunction and photoreceptor degeneration (1). Congenic strains of RCS rats,
genetically similar to the inbred RCS strain, were produced in the
1970s and early 1980s (4,5). These strains comprise: i) pink-eyed
rats that are wild-type (+/+) at the retinal dystrophy locus (rdy
locus; the strain is designated as RCS-rdy+) and serve
as control animals for the pink-eyed dystrophic, inbred RCS strain;
ii) pigmented (black-hooded) dystrophic rats (the strain is
designated as RCS-p+); and iii) pigmented,
non-dystrophic rats for use as normal controls for the pigmented
dystrophic rat (the strain is designated as RCS-rdy+
p+).
Mammalian retinal degeneration is involved in
extensive structural changes in retinal neurons (6), loss of bipolar (7) and ganglion cells (8), heterotopic migration of neurons
(9), and the presence of ectopic
neurites and microneuromas (10).
The action potentials of retinal ganglion cells were reduced in
amplitude and frequency during retinal degeneration, although
morphological changes of retina ganglion cells are not apparent
(11). To date, the mechanism(s)
responsible for retinal neuronal dysfunction and loss of retinal
neurons at the terminal stage of degeneration remain unclear.
Glutamate is a major excitatory neurotransmitter in
the vertebrate retina, responsible for the input of visual signals
(12). In the normal state,
photoreceptors (rods and cones) and bipolar cells release
glutamate, which is toxic to retinal cells (13,14), glial L-glutamate/L-aspartate
transporter (GLAST), which is expressed in the membrane of Müller
cells, and glutamine synthetase (GS), which is expressed in the
cytoplasm of Müller cells to maintain the requisite glutamate
homeostasis (15,16). The glutamate in retinal neurons is
bound with vesicular glutamate transporter-1 (VGLUT-1) reserved in
the synaptic vesicle and is released during neuronal excitation.
After being released by neurons, free glutamate in the
extracellular space is bound with GLAST and taken up by Müller
cells. Glutamate is then transformed to glutamine by GS in Müller
cells. Subsequently, glutamine is released by Müller cells to the
extracellular space, and the glutamatergic neurons uptake glutamine
in order to synthesize glutamate. Finally, the glutamate reserves
in the synaptic vesicle bind with VGLUT-1. As a result, finely
tuned glutamate release, uptake, and degradation are essential for
avoiding neurotoxicity and transmitting normal signals from
photoreceptors to bipolar and ganglion cells.
In the present study, we investigated the
expressions of key factors involved in glutamate metabolism in the
degenerating RCS rat retina. Both mRNA and protein levels of
VGLUT-1, protein kinase Cα (PKCα), GS, and GLAST were evaluated by
quantitative real-time polymerase chain reaction (qPCR),
immunohistochemistry, and western blot analyses.
Materials and methods
Animals
All experiments were conducted with the approval of
the Animal Ethics Committee of the Southwest Hospital, Chongqing,
China, and the ethics guidelines set forth by the Laboratory Animal
Care and Use Committee of the Association for Research in Vision
and Ophthalmology were followed. All efforts were made to minimize
animal suffering and to use the fewest number of animals in the
study.
Black-eyed dystrophic strain (RCS-p+) and
black-eyed non-dystrophic control strain (RCS-rdy+
p+) rats were anesthetized by intraperitoneal injection
of 10% chloral hydrate, after which both eyes were enucleated.
Animals were then euthanized with an overdose of anesthetic.
Retinas from both groups of rats were analyzed at four scheduled
time-points: postnatal Days 15 (P15), P30, P60 and P90. At least
six rat retinas were used for each time-point.
Immunohistochemistry
Eyes for immunohistochemical analysis were fixed in
4% paraformaldehyde in 0.1 M phosphate-buffered saline (PBS) for 2
h at 4°C, and then cryoprotected in 30% sucrose in 0.1 M PBS for
1–2 days at 4°C. Eyecups were embedded with optimum cutting
temperature (OCT) compound prior to cryostat sectioning at 10 μM,
and they were stored at −80°C for subsequent use. Frozen tissue
sections were initially thawed and rehydrated in 0.01 M PBS (pH
7.2). Following incubation in 5% normal goat serum (Jackson
ImmunoResearch, USA) for 1 h at room temperature, sections were
incubated with mouse monoclonal PKCα antibody (1:500) (Abcam, UK)
overnight at 4°C. Following three rinses (5 min each) with PBS,
fluorescein isothiocyanate (FITC)-conjugated goat anti-mouse IgG
antibody (1:200) (Jackson ImmunoResearch) was added to the slides
and the slides were incubated for 1 h at room temperature. Finally,
4′,6-diamidino-2-phenylindole (DAPI) staining was performed for
labeling cell nuclei. After several rinses with PBS, the sections
were covered with coverslips.
Quantitative real-time polymerase chain
reaction
After enucleating, retinas were isolated and frozen
in liquid nitrogen until use. For qPCR analysis, total RNA was
isolated with TRIzol reagent, according to the manufacturer’s
instructions. After contaminated DNAs were removed using the TURBO
DNA-free™ kit (Applied Biosystems), cDNAs were synthesized from 2
μg of total RNA using the PrimeScript™ RT Reagent kit (Takara Bio,
Inc., Japan) in 20 μl of reaction mixture. qPCR was carried out
using the Bio-Rad 5-Color System (Bio-Rad, USA). All primers were
designed online using SciTools software, Integrated DNA
Technologies, Inc., (IDT). The sequences are listed in Table I. The expression change of a
target gene in RCS rats relative to control rats was calculated as
follows: fold change = 2−(ΔCT,Tg−ΔCT,control). The
following PCR scheme was used: 5 min at 94°C, (30 sec at 94°C, 30
sec at 63°C, 30 sec at 72°C) for 35 cycles, 10 min at 72°C, and
then 4°C thereafter.
 | Table IqPCR primers used in the present
study. |
Table I
qPCR primers used in the present
study.
| Gene (rat) | Forward primer | Reverse primer |
|---|
| PKCα (144
bp) |
GGGGGAAAGGGATGTCAGAG |
CTGCCCTCGTGTGAAGAACTT |
| VGLUT-1 (122
bp) |
TGCTGCTGGTGGTCGGATAC |
AGGGGCGATGTCCAAGTGGT |
| GAPDH (158
bp) |
GCCCATCACCATCTTCCAGGAG |
GAAGGGGCGGAGATGATGAC |
Western blot analysis
For detection of GLAST and GS protein expression
levels, RCS rat retinas from each group were homogenized in
ice-cold radio-immunoprecipitation assay (RIPA) lysis buffer [50 mM
of Tris-HCl buffer (pH 7.4), 150 mM NaCl, 1% Triton X-100, 1%
sodium deoxycholate, 0.1% SDS, 1 mM sodium orthovanadate, 25 mM
sodium fluoride, 1 mM EDTA, and 1 μg/ml leupeptin]. Homogenates
were then centrifuged at 12,000 × g for 5 min at 4°C, and the clear
supernatants were stored at − 80°C until use. Protein
concentrations were determined using the Bicinchoninic Acid kit
(Beyotime Institute of Biotechnology, China). Samples (50 μg of
protein/lane) were loaded and electrophoresed on 12%
SDS-polyacrylamide gels (SDS-PAGE) for 40 min at 120 V. Proteins
were then transferred to nitrocellulose (NC) membranes for 70 min
at 120 V. After transferring, the NC membranes were blocked with
blocking solution containing Tris-buffered saline, 0.1% Tween-20
(TBST), and 5% free-fat milk for 1 h at room temperature. Blots
were then washed three times (5 min each) with TBST and incubated
with rabbit polyclonal GLAST antibody or GS antibody (Abcam)
overnight at 4°C. After washing, all membranes were again incubated
with mouse monoclonal GAPDH antibody for 1 h at room temperature.
Subsequently, all membranes were incubated with the following
secondary antibodies: IRDye 680-labeled donkey anti-rabbit IgG and
IRDye 800-labeled donkey anti-mouse IgG (LI-COR Biosciences,
Lincoln, NE, USA) in turn for 1 h at room temperature with shaking.
Membranes were rinsed between incubations. Finally, NC membranes
were scanned for GLAST, GS and GAPDH bands using the Odyssey
infrared imaging system with Odyssey Application software V1.2.15.
Densitometry ratios between GLAST and GAPDH, and GS and GAPDH were
obtained to semi-quantify the relative levels of GLAST and GS.
Statistical analysis
All data are presented as the means ± standard error
(SE). Data were evaluated by one-way analysis of variance (ANOVA)
and the LSD-q test, with post hoc statistical analysis for the same
group, and t-tests for comparing means between experimental and
control groups at the same time-points. Values of p<0.05 were
considered to indicate statistically significant differences.
Results
Changes in outer nuclear layer thickness
during retinal degeneration
Immunofluorescence staining showed that the
thickness of the outer nuclear layer (ONL) in control group retinas
remained unchanged at all time-points (Fig. 1A–D). In the RCS rats, the ONL
thickness at P15 was similar to that of the control group, but a
thinner ONL was observed at P30. In addition, the number of
photoreceptor cells in RCS rats was decreased at P60, and
disappeared by P90 (Fig. 1E–H).
Meanwhile, in RCS rats, a significant fibrotic glial seal layer
(FGSL) between the ONL and outer plexiform layer (OPL) had
gradually formed. There were no significant differences in the mean
thickness of the inner nuclear layer (INL) between the two
groups.
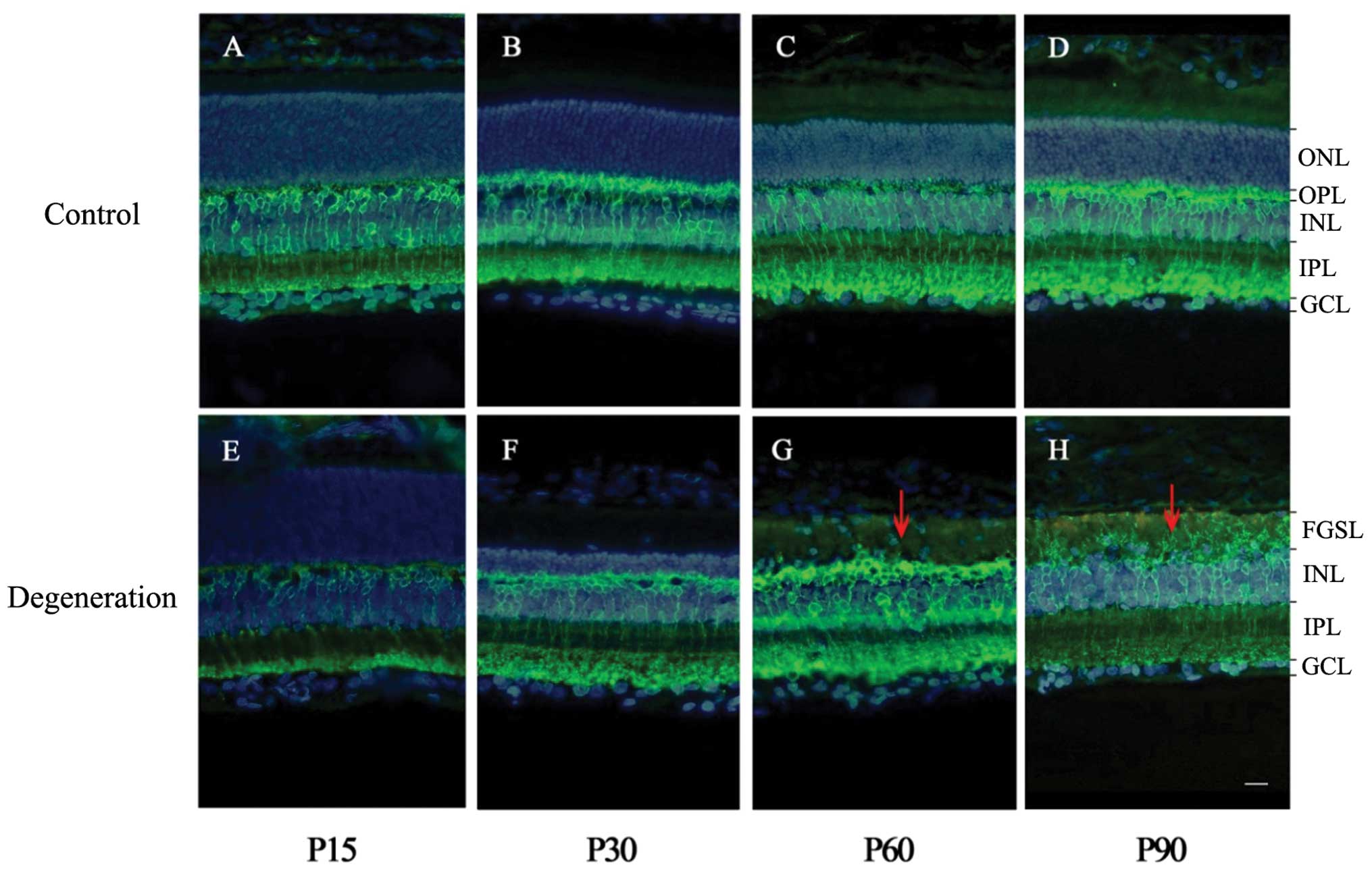 | Figure 1PKCα immunofluorescence staining of
rat retinas in RCS and control rats. PKCα (green) labeling shows
rod bipolar cells, including dendrites, soma, and axon terminals.
DAPI (blue) labeling shows whole cell nuclei in the retina. (A–D)
Digital images of vertical sections of rat retinas (control group).
(E–H) Digital images of vertical sections of RCS rat retinas
(degeneration group). The ONL thickness was the same between the
control and degeneration groups at P15, but was thinner at P30 in
the RCS rats, in which photoreceptor cells decreased in number at
P60 and were virtually nonexistent at P90. Meanwhile, the INL
remained almost unchanged. Red arrows indicate the fibrotic glial
seal of Müller cells. ONL, outer nuclear layer; OPL, outer
plexiform layer; INL, inner nuclear layer; IPL, inner plexiform
layer; GCL, ganglion cell layer; FGSL, fibrotic glial seal layer.
Scale: one bar, 10 μm. |
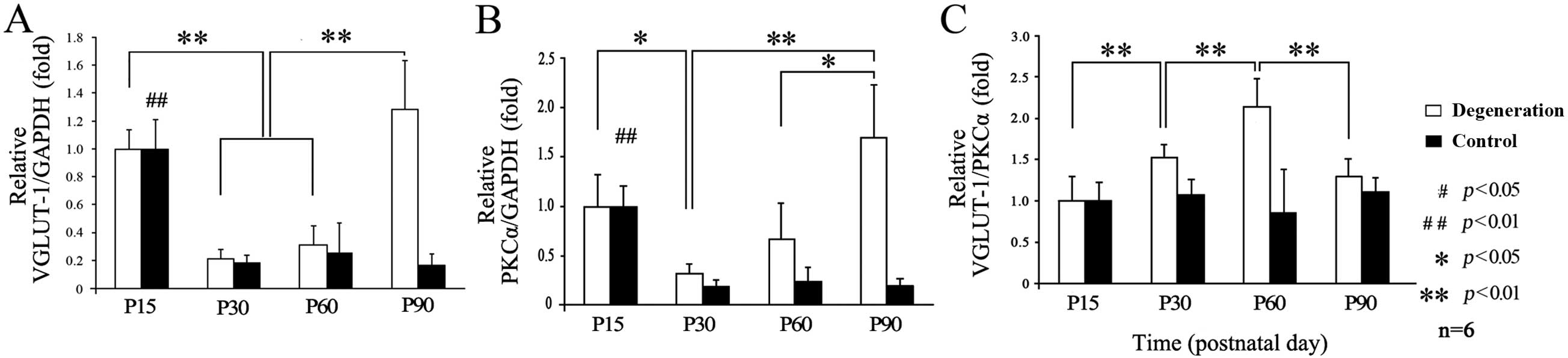
Changes in glutamate release during
retinal degeneration
qPCR results showed that PKCα and VGLUT-1 mRNA were
present in the retinas. No significant difference was found at P15
between RCS and control rats, although different tendencies existed
in both groups. In the control group, VGLUT-1 expression reached a
peak at P15 (Fig. 2A), decreased
rapidly at P30, and then stabilized to a lower level. By contrast,
VGLUT-1 expression in the RCS rats decreased slightly at P30, then
gradually increased at P60, and increased rapidly (as much as
3-fold) at P90. Differences were also seen in the PKCα expression
tendencies (Fig. 2B). In the
control group, the level of VGLUT-1 relative to PKCα remained
unchanged at all time-points (Fig.
2C), while in the RCS rats, it reached a peak at P60 and then
decreased at P90.
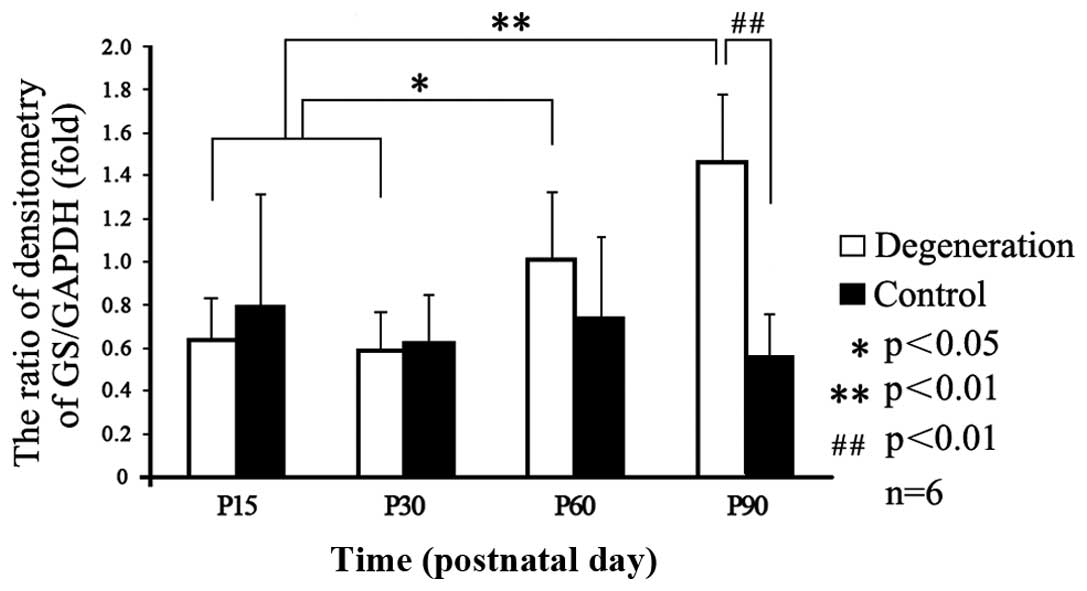
Transport and conversion capacity of
glutamate
Western blot analysis was used to analyze the
expression levels of GLAST and GS. Odyssey infrared imaging showed
that the GLAST levels did not change significantly in either group
at any time-point (Fig. 3).
However, analysis of GS expression relative to that of GAPDH showed
a time-dependent increase in the control group (Figs. 3 and 4), while it gradually increased from P15
to its peak value at P90 (p<0.05) in the degeneration group.
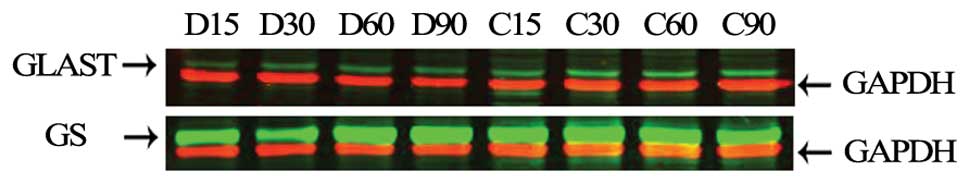 | Figure 3Western blot analysis of GLAST and GS
by Odyssey infrared imaging of the retinas of each group
(n=6/group). Representative western blot analyses for GLAST (60
kDa), GS (42 kDa), and GAPDH (36 kDa) are shown. Red blots are
GAPDH bands; green blots in each membrane represent GLAST and GS
bands. D15, D30, D60 and D90 represent the RCS rats at P15, P30,
P60 and P90, respectively. C15, C30, C60 and C90 represent the
control group at P15, P30, P60 and P90, respectively. |
Discussion
Glutamate is involved in neuronal survival during
postnatal development of the retina (17,18) and plays a key role in the
formation of retinal synaptic circuitry (18–21). In rats, the normal death periods
of bipolar cells and rods extend from birth to P4. Subsequently,
there is a gradual decrease in the incidence of cell death, and the
degeneration of bipolar cells continues until P48 (22). The apoptosis phase of
photoreceptor cells in the normal rat retina occurs between P12 and
P72, peaking at P23 (23). These
occurrences lead to decreasing VGLUT-1 and PKCα levels in normal
and degenerating rat retinas, consistent with our data showing that
the expression of VGLUT-1 and PKCα was significantly decreased at
P30 compared to P15.
It is known that retinal degeneration in RCS rats
involves initial rod photoreceptor loss (24,25). A very high percentage of inner
retinal neurons remained histologically intact, although loss of
cells in all retinal layers was found (8). The bipolar cells in the inner
plexiform layer (IPL) gradually became the predominant cells and
became the main source releasing glutamate following retinal
degeneration (25). The number of
rod bipolar cells in the IPL is relatively stable during retinal
degeneration, due to their characteristics of terminal
differentiation and incapability of proliferation. Since PKCα
exists mainly in bipolar cells in the retina, we indirectly
detected the number of bipolar cells in the INL by detecting the
levels of PKCα. VGLUT-1 was specifically localized to photoreceptor
and bipolar cell terminals in the retina (26) and accounts for the ability of
excitatory neurons to release glutamate by exocytosis (27). VGLUT-1 exhibits the glutamate
levels released and stored in the whole retina. Therefore, we
detected the ratio of VGLUT-1 to PKCα to indirectly show whether or
not the glutamate levels released and stored in the whole retina
had changed. Our data demonstrated that the ONL thickness
decreased, while the ratio of VGLUT-1 to PKCα increased during
retinal degeneration up to P60. These results indicate that the
percentage of living rod bipolar cells increased, while the bipolar
cells simultaneously became the main glutamate-releasing neurons in
the retina during retinal degeneration. Thus, the ability of
bipolar cells to release and store glutamate increased.
The relative level of VGLUT-1 to PKCα reached a peak
at P60, but subsequently decreased at P90, which indicated that
glutamate release increases gradually until the middle phase and
decreases during the latter phases of retinal degeneration. These
findings may be helpful in clarifying a contradiction regarding
glutamate changes during retinal degeneration. Certain studies have
reported that glutamate increases (28), whereas others have reported a
decreasing trend during later periods of degeneration (29). In this study, at the late stage of
retinal degeneration, the glutamate-releasing neurons in the retina
were mainly bipolar cells. Therefore, the decrease of this ratio
demonstrated that the release of glutamate from bipolar cells to
ganglion cells decreased, which suggests the deafferentation of
ganglion cells.
Glutamate toxicity has been demonstrated in both
inner retinal cells and photoreceptor terminals (15,16). Since there is no apparent
extracellular conversion of glutamate, retinal tissues require a
powerful uptake mechanism capable of quickly removing extracellular
glutamate to protect itself. Müller cells play a critical role in
the inactivation of glutamate released by glutamatergic neurons,
since they have a high affinity for Na+-linked
transporters that take up glutamate (30). In addition, unprotected neurons in
culture may be killed by even lower concentrations of glutamate in
the medium (31–33).
Glutamate uptake is accomplished by glutamate
transporter proteins located in the plasma membranes of retinal
Müller cells (34,35). GLAST has been described in retinal
Müller, astrocyte, and RPE cells in rats (36,37) and the consequences induced by its
absence from neuronal elements have also been reported (38). Our data show that GLAST expression
levels remain unchanged throughout retinal degeneration in RCS
rats, which is different from previous reports (28). We hypothesize that such a
discrepancy likely arises from the different animal models and
infrared imaging systems used. For instance, the probing of the
target and reference proteins simultaneously in a particular sample
indicates blots at ~60 kDa, without GLAST multimeric forms in our
results to abate any possible measurement errors. In addition, the
retinal degeneration mouse is a rapid retinal degeneration model,
unlike the RCS rat, which is a slow degeneration animal model.
Despite these differences, our results indicate that glutamate
uptake decreases (with GLAST levels remaining unchanged) lead to
inefficient extracellular glutamate transport into Müller cells
and, consequently, induce the accumulation of glutamate in the
extracellular space, resulting in toxicity to living neurons.
In our study, GS expression remained constant in the
control group at all time-points, while differences in the RCS rats
appeared from P30 onwards, with a significant increase at P90.
These findings are consistent with results of a previous study, in
which GS increased during the terminal stages of retinal
degeneration (28). In our study,
the GS expression pattern was similar to that of VGLUT-1 from P30
to P90. Increased GS expression does not result from Müller cell
proliferation or from any concentration change in the RCS retinas
(28). By P15 and P30, the
conversion of glutamate into glutamine via GS activity in Müller
cells was efficient; however, from P60 to P90, the level of GS in
Müller cells was gradually increased and the glutamate balance was
eventually disrupted, suggesting the existence of glutamate
conversion enhanced within Müller cells. In general, expression of
GS is regulated by glutamate (39). When the major glutamate-releasing
population of neurons and photoreceptors degenerate, GS expression
in Müller cells is reduced (40–42). However, in the present study, the
number of glutamatergic neurons reduced, while the level of GS
increased, indicating that intracellular glutamate likely increases
in Müller cells.
In conclusion, our results demonstrate that VGLUT-1
expression is high at P15 in both RCS and control rats. However, a
gradual increase in VGLUT-1 levels, accompanied by photoreceptor
degeneration, was observed after P30, while the relative expression
of VGLUT-1 to PKCα levels increased up to P60, and decreased in
P90, suggesting that glutamate release and storage gradually
increases until the middle stage of retinal degeneration, but it
decreases in the latter stage of retinal degeneration. Finally,
GLAST expression levels remained unchanged, while GS expression
increased gradually during the early stages and reached peak levels
during the latter stages of retinal degeneration.
Acknowledgements
This study was supported by the National Basic
Research Program of China (973 program) (Grant no. 2007CB512203).
The authors thank Jianrong He for technical assistance, and Lifeng
Chen and Yaochen Li for their suggestions and discussion on the
manuscript.
References
|
1
|
Olney JW: The toxic effects of glutamate
and related compounds in the retina and the brain. Retina.
2:341–359. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sullivan LS and Daiger SP: Inherited
retinal degeneration: exceptional genetic and clinical
heterogeneity. Mol Med Today. 2:380–386. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
D’Cruz PM, Yasumura D, Weir J, et al:
Mutation of the receptor tyrosine kinase gene Mertk in the retinal
dystrophic RCS rat. Hum Mol Genet. 9:645–651. 2000.PubMed/NCBI
|
|
4
|
LaVail MM: Photoreceptor characteristics
in congenic strains of RCS rats. Invest Ophthalmol Vis Sci.
20:671–675. 1981.PubMed/NCBI
|
|
5
|
LaVail MM, Sidman RL and Gerhardt CO:
Congenic strains of RCS rats with inherited retinal dystrophy. J
Hered. 66:242–244. 1975.PubMed/NCBI
|
|
6
|
Strettoi E, Porciatti V, Falsini B,
Pignatelli V and Rossi C: Morphological and functional
abnormalities in the inner retina of the rd/rd mouse. J Neurosci.
22:5492–5504. 2002.PubMed/NCBI
|
|
7
|
Strettoi E and Pignatelli V: Modifications
of retinal neurons in a mouse model of retinitis pigmentosa. Proc
Natl Acad Sci USA. 97:11020–11025. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Santos A, Humayun MS, Juan Ed, et al:
Preservation of the inner retina in retinitis pigmentosa. A
morphometric analysis. Arch Ophthalmol. 115:511–515. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jones BW and Marc RE: Retinal remodeling
during retinal degeneration. Exp Eye Res. 81:123–137. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Marc RE and Jones BW: Retinal remodeling
in inherited photoreceptor degenerations. Mol Neurobiol.
28:139–147. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen ZS, Yin ZQ, Chen S and Wang SJ:
Electrophysiological changes of retinal ganglion cells in Royal
College of Surgeons rats during retinal degeneration. Neuroreport.
16:971–975. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kalloniatis M and Tomisich G: Amino acid
neurochemistry of the vertebrate retina. Prog Retin Eye Res.
18:811–866. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Babai N, Morgans CW and Thoreson WB:
Calcium-induced calcium release contributes to synaptic release
from mouse rod photoreceptors. Neuroscience. 165:1447–1456. 2010.
View Article : Google Scholar
|
|
14
|
Yang XL: Characterization of receptors for
glutamate and GABA in retinal neurons. Prog Neurobiol. 73:127–150.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Karl MO, Hayes S, Nelson BR, Tan K,
Buckingham B and Reh TA: Stimulation of neural regeneration in the
mouse retina. Proc Natl Acad Sci USA. 105:19508–19513. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shen W, Li S, Chung SH and Gillies MC:
Retinal vascular changes after glial disruption in rats. J Neurosci
Res. 88:1485–1499. 2010.PubMed/NCBI
|
|
17
|
Nicoletti F, Bruno V, Copani A, Casabona G
and Knopfel T: Metabotropic glutamate receptors: a new target for
the therapy of neurodegenerative disorders? Trends Neurosci.
19:267–271. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Phillips MJ, Otteson DC and Sherry DM:
Progression of neuronal and synaptic remodeling in the rd10 mouse
model of retinitis pigmentosa. J Comp Neurol. 518:2071–2089. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cuenca N, Pinilla I, Sauve Y and Lund R:
Early changes in synaptic connectivity following progressive
photoreceptor degeneration in RCS rats. Eur J Neurosci.
22:1057–1072. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang J and Diamond JS: Subunit- and
pathway-specific localization of NMDA receptors and scaffolding
proteins at ganglion cell synapses in rat retina. J Neurosci.
29:4274–4286. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jarsky T, Tian M and Singer JH: Nanodomain
control of exocytosis is responsible for the signaling capability
of a retinal ribbon synapse. J Neurosci. 30:11885–11895. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vecino E, Hernandez M and Garcia M: Cell
death in the developing vertebrate retina. Int J Dev Biol.
48:965–974. 2004. View Article : Google Scholar
|
|
23
|
Vogel M and Moller K: Cellular decay in
the rat retina during normal post-natal development: a preliminary
quantitative analysis of the basic endogenous rhythm. Albrecht Von
Graefes Arch Klin Exp Ophthalmol. 212:243–260. 1980. View Article : Google Scholar
|
|
24
|
Jones BW, Watt CB and Marc RE: Retinal
remodelling. Clin Exp Optom. 88:282–291. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Marc RE, Jones BW, Anderson JR, et al:
Neural reprogramming in retinal degeneration. Invest Ophth Vis Sci.
48:3364–3371. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gong J, Jellali A, Mutterer J, Sahel JA,
Rendon A and Picaud S: Distribution of vesicular glutamate
transporters in rat and human retina. Brain Res. 1082:73–85. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fremeau RT, Kam K, Qureshi T, et al:
Vesicular glutamate transporters 1 and 2 target to functionally
distinct synaptic release sites. Science. 304:1815–1819. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Delyfer MN, Forster V, Neveux N, Picaud S,
Leveillard T and Sahel JA: Evidence for glutamate-mediated
excitotoxic mechanisms during photoreceptor degeneration in the rd1
mouse retina. Mol Vis. 11:688–696. 2005.PubMed/NCBI
|
|
29
|
Okada M, Okuma Y, Osumi Y, Nishihara M,
Yokotani K and Ueno H: Neurotransmitter contents in the retina of
RCS rat. Graefes Arch Clin Exp Ophthalmol. 238:998–1001. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Garlin AB, Sinor AD, Sinor JD, Jee SH,
Grinspan JB and Robinson MB: Pharmacology of sodium-dependent
high-affinity L-[3H]glutamate transport in glial cultures. J
Neurochem. 64:2572–2580. 1995.
|
|
31
|
Minami M, Oku H, Okuno T, Fukuhara M and
Ikeda T: High infusion pressure in conjunction with vitreous
surgery alters the morphology and function of the retina of
rabbits. Acta Ophthalmol Scan. 85:633–639. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sedaghat K, Finkelstein DI and Gundlach
AL: Effect of unilateral lesion of the nigrostriatal dopamine
pathway on survival and neurochemistry of parafascicular nucleus
neurons in the rat - evaluation of time-course and LGR8 expression.
Brain Res. 1271:83–94. 2009. View Article : Google Scholar
|
|
33
|
Friedman LK and Segal M: Early exposure of
cultured hippocampal neurons to excitatory amino acids protects
from later excitotoxicity. Int J Dev Neurosci. 28:195–205. 2010.
View Article : Google Scholar
|
|
34
|
Ward MM, Jobling AI, Kalloniatis M and
Fletcher EL: Glutamate uptake in retinal glial cells during
diabetes. Diabetologia. 48:351–360. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Namekata K, Harada C, Kohyama K, Matsumoto
Y and Harada T: Interleukin-1 stimulates glutamate uptake in glial
cells by accelerating membrane trafficking of
Na+/K+-ATPase via actin depolymerization. Mol
Cell Biol. 28:3273–3280. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Otori Y, Shimada S, Tanaka K, Ishimoto I,
Tano Y and Tohyama M: Marked increase in glutamate-aspartate
transporter (GLAST/GluT-1) mRNA following transient retinal
ischemia. Mol Brain Res. 27:310–314. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Derouiche A and Rauen T: Coincidence of
L-glutamate/L-aspartate transporter (GLAST) and glutamine
synthetase (GS) immunoreactions in retinal glia: evidence for
coupling of GLAST and GS in transmitter clearance. J Neurosci Res.
42:131–143. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lehre KP, Davanger S and Danbolt NC:
Localization of the glutamate transporter protein GLAST in rat
retina. Brain Res. 744:129–137. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shen F, Chen B, Danias J, et al:
Glutamate-induced glutamine synthetase expression in retinal Müller
cells after short-term ocular hypertension in the rat. Invest
Ophthalmol Vis Sci. 45:3107–3112. 2004.
|
|
40
|
Germer A, Jahnke C, Mack A, Enzmann V and
Reichenbach A: Modification of glutamine synthetase expression by
mammalian Müller (glial) cells in retinal organ cultures.
Neuroreport. 8:3067–3072. 1997.
|
|
41
|
Shen X and Xu G: Role of IL-1beta on the
glutamine synthetase in retinal Muller cells under high glucose
conditions. Curr Eye Res. 34:727–736. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Jablonski MM, Tombran-Tink J, Mrazek DA
and Iannaccone A: Pigment epithelium-derived factor supports normal
Müller cell development and glutamine synthetase expression after
removal of the retinal pigment epithelium. Glia. 35:14–25.
2001.
|