Introduction
Sporothrix schenckii (S. schenckii),
the etiologic agent of sporotrichosis, is a dimorphic fungus that
produces lymphocutaneous lesions. The mechanisms involved in the
morphological transitions of S. schenckii remain unknown,
although knowledge on the experimental control of these processes
has increased in the past years (1,2).
Inhibition of the differentiation of S. schenckii mycelial
into the parasitic yeast forms may be a noteworthy alternative for
the design of antifungal drugs to be used in sporothrichosis.
Therefore, understanding the morphological transition process and,
consequently, how the morphological transition can be inhibited,
may aid in the development of novel strategies to effectively
manage this increasingly important clinical problem.
Dimorphic switching requires fungal cells to undergo
changes in polarized growth in response to environmental stimuli.
Polarity establishment allows the development of a wide variety of
cell morphologies and the differentiation of distinct cell types
(3). The family of Ste20-related
kinases is conserved from yeast to mammals and includes the p21
activated kinases (PAKs) and germinal centre kinases (GCKs). These
kinases have been shown to be involved in signaling through mitogen
activated protein kinase (MAPK) pathways and in morphogenesis
through the regulation of cytokinesis and actin-dependent polarized
growth (4). Saccharomyces
cerevisiae (S. cerevisiae) Ste20 is required for efficient
activity of the pheromone response pathway and an effective
Hog1-mediated hypertonic stress response. It appears to play a role
in the activation of myosin, adhesion of mating partners and
vegetative functions relating to budding (5). Homologues of S. cerevisiae
Ste20 have been described in a number of other fungal species.
These include Candida albicans (C. albicans) Ste20 (Q92212),
Cryptococcus neoformans (C. neoformans) Ste20 (AF542531.1),
Ustilago maydis Ste20 (AF299352.1) and Pneumocystis
carinii Ste20 (AF332388). These Ste20 proteins were proved to
play important roles in mediating the regulation of various aspects
of morphogenesis, conjugation, mating and pathogenicity (6). We previously reported differentially
expressed proteins between the mold and early yeast stage of S.
schenckii using 2DE (7). The
protein spot J is homologous to the Ste20-like protein kinase from
Paracoccidioides brasiliensis and increases in the early
yeast form of S. schenckii.
In this study, we described the molecular cloning of
the S. schenckii Ste20 homologue, designated SsSte20. We
performed necessary function analysis of the SsSte20 gene and
detected the differential gene expression in dimorphic switch of
S. schenckii. These will establish the primary foundation of
understanding the function of the SsSte20 gene from S.
schenckii. The cloning and characterization of the SsSte20 gene
from S. schenckii is reported for the first time.
Materials and methods
Fungal strain, media and growth
conditions
The strain of S. schenckii used, ATCC10268,
was maintained at the Research Center for Pathogenic Fungi, Dalian
Medical University, China. To obtain a mycelial culture, the
ATCC10268 isolate was inoculated on Sabouraud dextrose agar (SDA)
medium and incubated at 25°C. The mycelial colonies thus obtained
were inoculated in Sabouraud's fluid medium and cultured with
shaking at 100 rpm at 25°C for 96 h. To achieve the switch of S.
schenckii from the mycelial phase to the yeast phase, mycelial
colonies were transferred to brain heart infusion (BHI) liquid
medium at 37°C and shaken at 100 rpm for 96 h. Mycelial and yeast
pellets were collected by centrifugation and stored at −80°C.
Total RNA, genomic DNA isolation and gene
cloning
Approximately 100 mg samples of S. schenckii
mycelial and yeast were separately pulverized under liquid nitrogen
with a mortar and pestle. Total RNA isolation was carried out
according to the manufacturer's protocol using the TRIzol Reagent
Kit (Invitrogen, Carlsbad, CA, USA) and treated with the RNase-free
DNase I kit from Takara to eliminate DNA contamination. Genomic DNA
was isolated from yeast phase colonies following the manufacturer's
protocol using the InstaGene™ Matrix kit (Bio-Rad, Hercules, CA,
USA). cDNA was synthesized from 500 μg of total RNA of ATCC10268 by
murine leukemia virus reverse transcriptase (MLV-RT) (Takara,
Tokyo, Japan) primed with oligo(dT) following the manufacturer's
instructions, and used as template for PCR. Degenerate primers,
SsSte20-F1 and SsSte20-R1, were designed based on multiple
alignments of the high conserved Ste20 domains of C.
albicans Ste20 (Q92212), C. neoformans Ste20
(AF542531.1), Ustilago maydis Ste20 (AF299352.1), and
Pneumocystis carinii Ste20 (AF332388) amino acid sequences.
PCR product of expected size was cloned into pMD18 vector (Takara)
and sequenced. The degenerate primers yielded a 302 bp fragment
homologous to known Ste20. To obtain the full-length cDNA sequence
of the SsSte20 gene, 5′-RACE and 3′-RACE were performed with
5′-Full RACE Kit and 3′-Full RACE Core Set Ver.2.0 kit (Takara)
according to the manufacturer's instructions. Nest-PCR was
performed. Briefly, five specific primers CTE869 F and CTE869 R of
3′-RACE and R132-1, R132-2 and R132-3 of 5′-RACE were synthesized
based on the cDNA sequence obtained by the degenerate primers. PCR
products of 5′- and 3′-RACE were both cloned into pMD18 vector
(Takara) and sequenced.
To determine the nucleotide sequence of the genomic
DNA corresponding to the SsSte20, PCR was performed using the
primers SsSte20-P1 and SsSte20-B3 and genomic DNA as template. The
PCR products were then sequenced. The sequences of all the primers
used in this study are listed in Table I.
 | Table ISequence of primers in this study. |
Table I
Sequence of primers in this study.
| Primer | Sequence (5′-3′) |
|---|
| SsGCK-F1 |
CAYAARYTNTGGATHGTNATGGA |
| SsGCK-R1 |
TGYTGDATNACYTCNGGNGCCATCCA |
| CTE869F |
AGAAGCTCCACCGCGATATCAAGGCC |
| CTE869R |
CGCCACCATGACCAAGAAGAACACCT |
| R132-1(GSP1) |
TGGAGCCTAACAAGACA |
| R132-2(GSP2) |
TTCTTGTCGGCGTGGAGGTAG |
| R132-3(GSP3) |
CAATGATGGCAATGTAGTCTTCG |
| P1 |
ATGGCGAGCAGAGATGACG |
| B3 |
CACGGCCTCTTCGTCGAG |
| F |
GCAACAGCAACAATTAGC |
| R |
GACACGTTCTCTTCCATG |
| 24T | (FAM)
CCAACAGCAACTACAACAGCAACAG |
Bioinformatic and phylogenetic analysis
of SsSte20
Nucleotide sequences and deduced amino acid
sequences of the cloned SsSte20 gene were analyzed. The nucleotide
sequences were analyzed as previously described by us (8) using Sequencer software (Sequencer,
USA) and the BLAST network service of the National Center for
Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.gov/blast). The open reading
frame (ORF) was found by the ORF Finder (http://www.ncbi.nlm.nih.gov/gorf/gorf.html). For the
exact localization of the exon/intron boundaries, the
mRNA-to-genomic alignment program Spidey (http://www.ncbi.nlm.nih.gov/IEB/Research/Ostell/Spidey/index.html)
was used. The deduced amino acid sequence was analyzed with the
Expert Protein Analysis System (http://www.expasy.org/) and the protein domain
features of SsSte20 were determined by using Simple Modular
Architecture Research Tool (http://hits.isb-sib.ch/cgi-bin/PFSCAN). Isoelectric
point and molecular weight prediction were carried out at
(http://cn.expasy.org/tools/pi_tool.html). Multiple
alignments of SsSte20 were performed with the ClustalW Multiple
Alignment Program (http://www.ebi.ac.uk/clustalw/).
Differential expression of SsSte20 in two
stages during dimorphic switch
The expression of SsSte20 transcript in different
stages (mycelial, yeast) was measured by real-time RT-PCR. Primers
and a TaqMan probe for target genes were designed with Primer
Select in DNASTAR software (Lasergene) and are listed in Table I (24T, 8F and 58R). Fifty
nanograms of total RNA were assayed from two stages of S.
schenckii in triplicate using the PrimeScript RT-PCR Kit
(Takara). The minus-reverse transcriptase control was also
performed in triplicate. The amplification conditions were
optimized for the ABI-PRISM-7500 instrument (Applied Biosystems).
The cycling conditions using TaqMan probe detection were 95°C for 2
min followed by 40 cycles at 95°C for 10 sec, 61°C for 10 sec, 72°C
for 40 sec. 18srDNA was selected as the endogenous control.
Relative quantification of target gene expression was evaluated
using the comparative cycle threshold (CT) method as
previously described by Livak and Schmittgen (9). The ΔCT value was
determined by subtracting the target CT of each sample from its
respective 18srDNA CT value. Calculation of
ΔΔCT involved using the mycelial sample ΔCT
value as an arbitrary constant to subtract from yeast sample
ΔCT values. Differences in expression of target genes
were determined by 2−ΔΔCT. Data are expressed
as arithmetic means ± SD unless otherwise indicated. Comparison
between mycelial and yeast samples was performed using the
Student's t-test. Differences with a P-value of <0.05 were
considered statistically significant.
Results
Cloning and genomic structure of
SsSte20
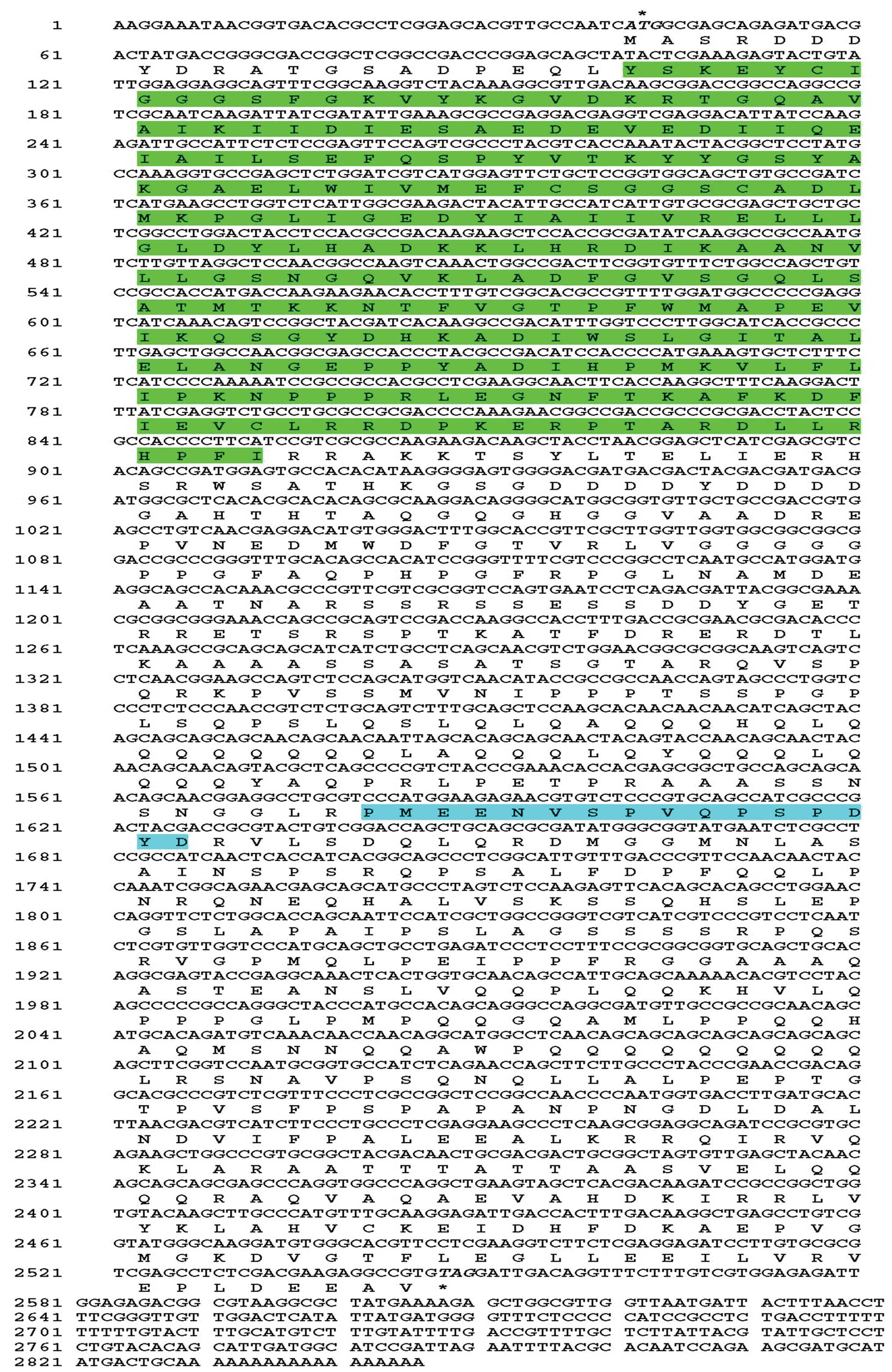
A full-length SsSte20 cDNA (2,846 bp) including an
ORF of 2505 bp, encoding 835 amino residues, was flanked by a 41 bp
5′-untranslated region (5′-UTR) and a 297 bp 3′-UTR. The SsSte20
genomic DNA is 2578 bp in length. The aligned results revealed that
there is one intron between the sequences of the genomic DNA and
the cDNA. Its 5′ and 3′ ends conformed to the basic consensus,
GT/AG, for the eukaryotic splice donor and acceptor site. Based on
the sequence of cDNA, the molecular weight of the predicted amino
acid is ∼91.31 kDa, the theoretical pI is 5.76.
Motif searches and sequence comparison showed that
SsSte20 consists of a 251-amino acid N-terminal kinase domain
(residues 21–271) (Fig. 1). In
the N-terminal extremity of the catalytic domain there is a
glycine-rich stretch of residues in the vicinity of a lysine
residue (K), which has been proved to be involved in ATP binding.
In the central part of the catalytic domain there is a conserved
aspartic acid residue (N) which is important for the catalytic
activity of the enzyme (Fig. 2).
One proline/glutamic acid/serine/threonine (PEST) motif was
searched using Epestfind in the C-terminal domain of SsSte20
(residues 514–529) (Fig. 1).
Database searches revealed that SsSte20 is a member of the Ste20
family of protein kinases. SsSte20 displays high homology
throughout the kinase domain and contains the amino acid sequence
GTPYFWMAPEV in the kinase domain (Fig. 2). This sequence motif, known as
the Ste20 signature, is characteristic of the Ste20 family of
protein kinases and is critical for the kinase activity of Ste20
(10).
Homology and phylogenetic analysis of
SsSte20
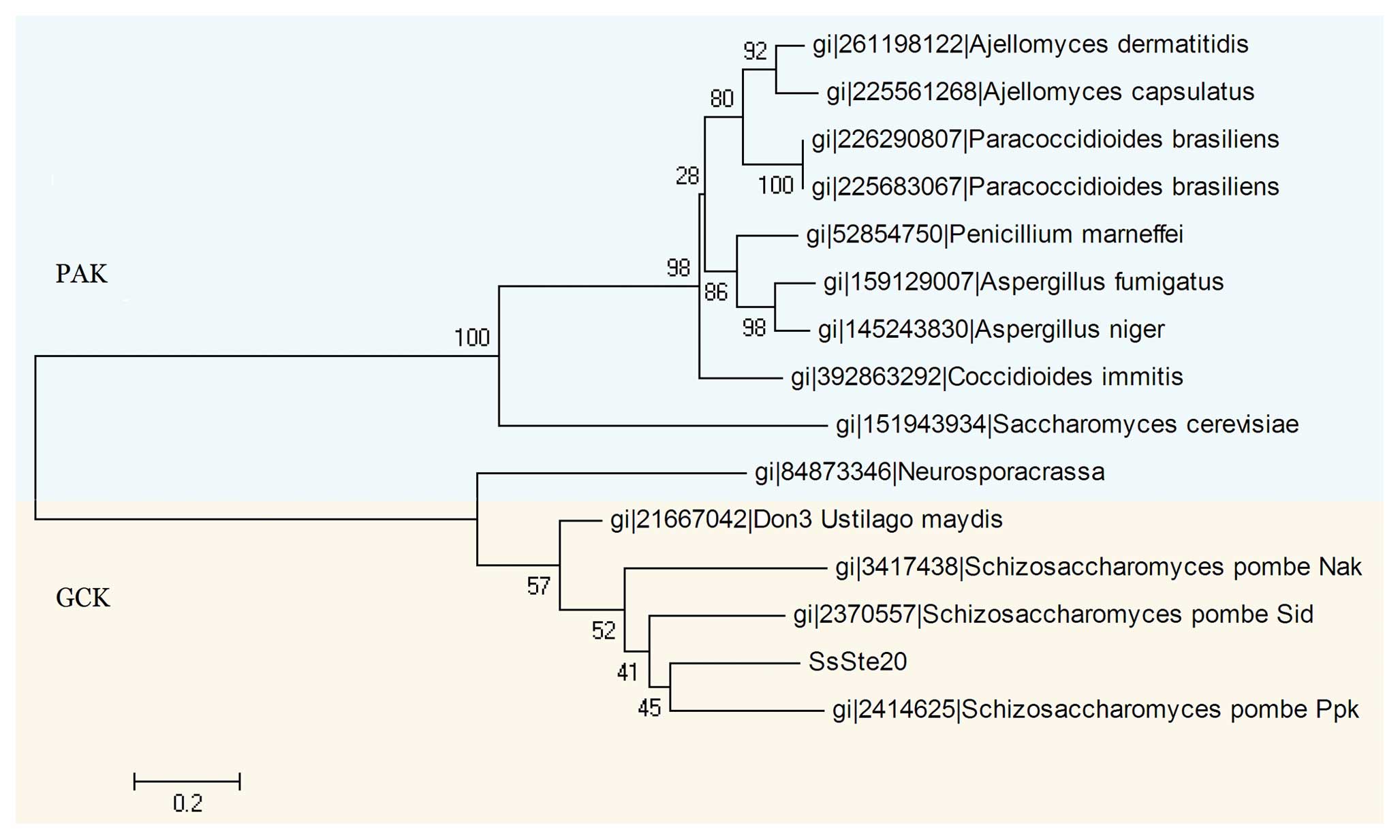
To clarify the relationship between SsSte20 and
other Ste20-related kinases, we calculated multiple sequence
alignments of these sequences. The derived evolutionary tree is
split into two main branches, one formed by some PAKs, the other
one by GCKs including SsSte20 sequence (Fig. 3). Further alignment clearly showed
that the SsSte20 kinase domain is highly homologous to that of
other GCKs, such as Don3 (Ustilago maydis, AF463451.1),
Ppk11 (Schizosaccharomyces pombe, NM_001019946.1), Sid1
(Schizosaccharomyces pombe, NM_001018997.1), Nak1
(Schizosaccharomyces pombe NM_001021931.1) and
polarity-defective 6 (Neurospora crassa, DQ336953.1). These
proteins share a similar structure and have 63, 56, 54, 47 and 38%
amino acid similarities in the catalytic domains, respectively.
These proteins have a highly conserved catalytic domain in common
but differ in their C-terminal regulatory domain, which does not
contain any identifiable sequence motifs.
Expression of SsSte20 in two stages of S.
schenckii
The mRNA expression of SsSte20 in different stages
was analyzed by real-time RT-PCR normalized against 18srDNA levels.
Expression was determined as fold increased
2−ΔΔCT levels relative to the stage with
lowest expression (mycelial) set to 1. The SsSte20 gene was
expressed in two stages of S. schenckii, with higher mRNA
levels observed in yeast (15.14-fold). There were significant
differences between the mycelial and the yeast form (Table II).
| Table IIRelative abundance of differential
expression gene as determined by real-time RT/PCR (mean ± SD)
(P<0.01). |
Table II
Relative abundance of differential
expression gene as determined by real-time RT/PCR (mean ± SD)
(P<0.01).
| cDNA name | Phase | Target
CT | 18srDNA
CT | ΔCT | ΔΔCT |
2−ΔΔCT |
|---|
| SsSte20 | Mycelial | 22.48±0.92 | 20.66±0.25 | 1.81±1.13 | 0±1.13 | 1 |
| Yeast | 19.99±0.23 | 22.09±0.60 | −2.11±0.79 | −3.92±0. 79 | 15.14 |
Accession number
The full length of cDNA sequence and the genomic DNA
sequence of the SsSte20 gene were submitted to the GenBank database
under the accession numbers JX312329 and JX857536,
respectively.
Discussion
The Ste20-like protein kinase family can be divided
into two subfamilies based on their domain structure and
regulation. The PAK/Ste20 subfamily contains a C-terminal catalytic
domain and an N-terminal binding site for the small G proteins
Cdc42 and Rac1, which are considered to be key regulatory molecules
linking surface receptors to the organization of the actin
cytoskeleton (11,12). By contrast, the kinases of the
second subfamily, the GCK subfamily, have an N-terminal kinase
domain and a C-terminal regulatory domain that does not have a
recognizable GTPase binding site (13). Furthermore, GCKs are subdivided
into two broad groups based on their structure. Group I GCKs have a
highly conserved C-terminal domain containing at least two PEST
motifs, two polyproline Src homology-3 (SH3) domains and an
additional highly conserved region of approximately 350 amino
acids. Group II GCKs share similarity to the catalytic domain of
group I; however, the C-terminal domain differs significantly. In
present study, SsSte20 was labelled as a GCK based on catalytic
domain located in the N-terminal site, which was also confirmed by
phylogenetic relationship analysis. We further identified SsSte20
as a group II GCK for its one PEST motif, no SH3 domain and no
conserved sequence in C-terminal domain.
Ste20 ortholog was proved to be involved in both the
morphogenesis and the pathogenesis of consequence of an inability
to grow in the pathogenic form. In C. albicans, filamentous
growth allows adherence and penetration of tissues during
infection. The cst20 (Ste20) mutants are unable to form hyphal
filaments in vitro and consequently show reduced kidney
colonization and virulence in a mouse model for systemic
candidiasis (14). While in
Penicillium marneffei (P. marneffei), infection occurs by
the inhalation of conidia that are phagocytosed by host pulmonary
alveolar macrophages where they germinate into yeast cells and
divide by fission. PakA (Ste20) plays an essential role in conidial
germination of P. marneffei at the host temperature of 37°C
such that ΔPakA mutant conidia fail to germinate during macrophage
infection (15). Although GCKs
currently have no described role in virulence, this may be due to a
lack of characterization rather than a lack of involvement. The
Kic1 (GCK) mutants in C. albicans and C. neoformans
probably have virulence defects as the C. albicans Kic1
mutant is unable to grow filamentously and the C. neoformans
mutant is unable to grow at 37°C, the temperature of the human
body, both of which are requirements for virulence in these
organisms (16,17). In S. schenckii, the
germination of conidia is a key pathogenicity determinant since
conidia are the infectious propagules. Does SsSte20, a GCK, have
the same function in the virulence of S. schenckii as Ste20
ortholog in other fungi? The mRNA expression of SsSte20 in yeast
cells of both ATCC10268 and a clinical S. schenckii isolated
from a patient with fixed sporotrichosis (data not shown) were
higher than the mycelial ones, which suggested that SsSte20 may be
involved in the morphogenesis or pathogenesis of S.
schenckii.
What is the environmental signal that SsSte20
responds to to participate in morphogenesis and pathogenesis? In
P. marneffei, PakB (Ste20) is essential for the formation of
yeast cells within macrophages and the ΔPakB mutant grows as
septate hyphae in vivo. PakB is not expressed at 37°C during
in vitro yeast growth but expression is induced during
macrophage infection (18). This
suggests that unlike PakA, which responds to temperature, PakB
responds to host cell inductive signals. In the present study, the
mycelial cells of S. schenckii switched to yeast cells with
obvious increased expression of SsSte20 when they were incubated in
BHI liquid medium at 37°C in vitro, which suggests SsSte20,
similar to PakA, can detect the change of temperature and nutrient
deprivation in the environment. Whether SsSte20 also responds to
other host signals remains to be elucidated. It is most likely that
there are other proteins that fungal cells come into contact with
in the early stages of infection. We are carrying out further
experiments on generating SsSte20-mutants to study its detailed
functions in this critical fungal pathogen.
Acknowledgements
This study was partly supported by a
grant from the National Natural Science Foundation of China (Grant
no. 81000069).
References
|
1
|
Valle-Aviles L, Valentin-Berrios S,
Gonzalez-Mendez RR and Rodriguez-Del Valle N: Functional, genetic
and bioinformatic characterization of a calcium/calmodulin kinase
gene in Sporothrix schenckii. BMC Microbiol. 7:1072007.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Valentín-Berríos S, González-Velázquez W,
Pérez-Sánchez L, et al: Cytosolic phospholipase A2: a
member of the signalling pathway of a new G protein α subunit in
Sporothrix schenckii. BMC Microbiol. 9:1002009.
|
|
3
|
Boyce KJ, Hynes MJ and Andrianopoulos A:
The Ras and Rho GTPases genetically interact to co-ordinately
regulate cell polarity during development in Penicillium
marneffei. Mol Microbiol. 55:1487–1501. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Boyce KJ and Andrianopoulos A:
Ste20-related kinases: effectors of signaling and morphogenesis in
fungi. Trends Microbiol. 19:400–410. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Calcagno AM, Bignell E, Rogers TR, Canedo
M, Muhlschlegel FA and Haynes K: Candida glabrata Ste20 is
involved in maintaining cell wall integrity and adaptation to
hypertonic stress, and is required for wild-type levels of
virulence. Yeast. 21:557–568. 2004. View
Article : Google Scholar
|
|
6
|
Hazan I and Liu H: Regulation of
morphogenesis by conserved developmental pathways in pathogenic
fungi. Pathogenic Fungi: Structural Biology and Taxonomy. San-Blas
G and Calderone RA: Caister Academic Press; Norfolk, UK: pp.
221–229. 2004
|
|
7
|
Zhang ZY, Hou BB, Xin Y and Liu XM:
Protein profiling of the dimorphic pathogenic fungus, Sporothrix
schenckii. Mycopathologia. 173:1–11. 2012. View Article : Google Scholar
|
|
8
|
Hou B, Zhang Z, Zheng F and Liu X:
Molecular cloning, characterization and differential expression of
DRK1 in Sporothrix schenckii. Int J Mol Med. 31:99–104.
2013.PubMed/NCBI
|
|
9
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
|
|
10
|
Knighton DR, Zheng JH, Ten Eyck LF, Xuong
NH, Taylor SS and Sowadski JM: Crystal structure of the catalytic
subunit of cyclic adenosine monophosphate-dependent protein kinase.
Science. 253:407–414. 1991. View Article : Google Scholar
|
|
11
|
Wu C, Whiteway M, Thomas DY and Leberer E:
Molecular characterization of Ste20p, a potential mitogen-activated
protein or extracellular signal-regulated kinase kinase (MEK)
kinase kinase from Saccharomyces cerevisiae. J Biol Chem.
270:15984–15992. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Herskowitz A, Choi S, Ansari AA and
Wesselingh S: Cytokine mRNA expression in postischemic/reperfused
myocardium. Am J Pathol. 146:419–428. 1995.PubMed/NCBI
|
|
13
|
Hall A: Rho GTPases and the actin
cytoskeleton. Science. 279:509–514. 1998. View Article : Google Scholar
|
|
14
|
Leberer E, Harcus D, Clark KL, Dignard D,
Ziegelbauer K, et al: Signal transduction through homologs of the
Ste20p and Ste7p protein kinases can trigger hyphal formation in
the pathogenic fungus Candida albicans. Proc Natl Acad Sci
USA. 93:13217–13222. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Boyce KJ and Andrianopoulos A: A
p21-activated kinase is required for conidial germination in
Penicillium marneffei. PLoS Pathog. 3:e1622007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Walton FG, Heitman J, Boone AC and Editor
M: Conserved elements of the RAM signaling pathway establish cell
polarity in the basidiomycete Cryptococcus neoformans in a
divergent fashion from other fungi. Mol Biol Cell. 17:3768–3780.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Song Y, Cheon SA, Lee KE, Lee S, Lee B, et
al: Role of the RAM network in cell polarity and hyphal
morphogenesis in Candida albicans. Mol Biol Cell.
19:5456–5477. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Boyce KJ, Schreider L and Andrianopoulos
A: In vivo yeast cell morphogenesis is regulated by a p21-activated
kinase in the human pathogen Penicillium marneffei. PLoS
Pathog. 5:e10006782009. View Article : Google Scholar : PubMed/NCBI
|