Introduction
Inflammatory bowel disease (IBD), which is a major
gastrointestinal disease consisting of Crohn’s disease (CD) and
ulcerative colitis (UC), is a recurring and remitting disorder
characterized by a range of symptoms, including abdominal pain,
severe diarrhoea, rectal bleeding and wasting. It has been
suggested that IBD results from environmental, genetic and
immunological factors. Moreover, compelling data have suggested
that the major contributing factor to IBD is an inappropriate
reaction of the intestinal immune system to normal luminal enteric
flora, according to individual genetic differences (1–3).
CD and UC are distinguished by the tissues that are affected: CD
can affect any region of the gastrointestinal tract in a
discontinuous and transmural manner, whereas UC is restricted to
the surface mucosa of the colon, in particular the rectum (4). Although the pathogenesis of these
diseases is not yet fully understood, transmural inflammation in
IBD, including lymphoid hyperplasia, submucosal edema, ulcerative
lesions, as well as fibrosis is typically associated with the
deficiency of T cell-mediated regulatory processes which would
normally prevent and/or terminate inflammation (5). However, anti-inflammatory and
immunosuppressive agents which we currently regard as important
treatment regimens are not curative and only reduce the degree of
intestinal inflammation associated with the disease (6). It is urgent to develop new
therapeutic approaches that are more effective in the treatment of
IBD.
Regulatory T cells (Tregs) are considered critical
for maintaining immune homeostasis by establishing tolerance to
foreign, non-pathogenic antigens that exist in commensal bacteria
and food (7,8). Patients who have non-functional or
absent Tregs associated with genetic mutations in Foxp3 always have
severe intestinal inflammation characterized by lymphocytic
infiltration (9,10). Similarly, mice lacking
Foxp3+ Tregs are more likely to develop severe colitis
(11). Previously, it has been
suggested that the restoration of Tregs can prevent and treat
colitis in mouse models of IBD (12).
Mesenchymal stem cells (MSCs) are multipotent cells
which can present in the bone marrow and may be isolated from a
variety of tissues, including adipose, liver, amniotic fluid, lung,
skeletal muscle, as well as kidney tissue (13). On the basis of their convenient
isolation, their low expression of human leukocyte antigen (HLA)
and co-stimulatory molecules, as well as the fact that their
isolation does not provoke ethical controversy, bone marrow-derived
MSCs (BM-MSCs) are a promising alternative for cellular
immunotherapy (14,15). MSCs can not only differentiate
into multiple cell lineages, including adipocytes, chondrocytes and
osteocytes, but they also exhibit immunomodulatory functions in
innate and adaptive immune systems. The immunosuppressive activity
of MSCs is associated with a range of immune cells, including T, B
and natural killer (NK) lymphocytes and dendritic cells (13). A number of in vitro and
in vivo studies have suggested that MSCs exert their
immunomodulatory effects through the expansion and/or induction of
Tregs (16).
In view of their ability to differentiate into
tissue-specific cell types, as well as their immunoregulatory
effects in a variety of immune cell types, MSCs have the potential
to prevent inflammatory disorders. A number of trials of MSC
infusion for the treatment of graft-versus-host disease and CD have
been published in recent years (17,18). However, the interactions between
MSCs and Tregs in IBD and whether MSCs can be used for the
treatment of IBD remain unclear. In this study, we infused BM-MSCs
into a 2,4,6-trinitrobenzene sulfonic acid (TNBS)-induced rat model
of colitis in the effort to confirm our hypothesis that MSCs can be
used for the treatment of IBD and to investigate the possible
mechanisms behind the immunosuppressive activity of MSCs and
Tregs.
Materials and methods
Animals
Female Sprague-Dawley (SD) rats of specific
pathogen-free grade, weighing approximately 200 to 250 g, were
purchased from the Hubei Provincial Center for Disease Control,
China. All rats were allowed free access to standard chow, as well
as water and were randomly assigned to 3 different groups [control
(untreated group), the TNBS + PBS group and the TNBS + MSC group].
The experimental protocol was approved by the Experimental Animal
Center of Tongji Medical College, Huazhong University of Science
and Technology, Wuhan, China.
Isolation and culture of BM-MSCs
The isolation and culture of the BM-MSCs was carried
out as previously described (19). Four-week-old male rats were
sacrificed by cervical dislocation. The rats were immersed in 75%
ethanol for 5 min, and the bone marrow was then isolated from the
bilateral femurs and tibias. The cell suspension was then loaded
onto a Percoll gradient (Sigma-Aldrich, St. Louis, MO, USA), and
the cells were centrifugated for 5 min at 352 × g. The top
supernatant was then transferred into a tube and washed with
phosphate-buffered saline (PBS). The cells were plated in a plastic
tissue culture flask (Corning, NY, USA) and cultured in low-glucose
complete cell culture medium consisting of α-minimum essential
medium (α-MEM; Gibco, Invitrogen Corp., Grand Island, NY, USA)
containing 10% fetal bovine serum (FBS; HyClone Laboratories, Inc.,
Logan, UT, USA). The non-adherent cells were removed by adding
fresh medium at 48 h and every 3–4 days thereafter. All the
cultures were maintained at 37°C in a 5% CO2 atmosphere.
For approximately 1–2 weeks, only adherent cells were collected
with 0.25% trypsin solution (Gibco, Invitrogen Corp.). The cells
were passaged at approximately 80% confluency. Third-passage cells
were used for all the experiments.
Cell-surface marker characterization of
BM-MSCs
For flow cytometric analysis, the BM-MSCs were
trypsinized and incubated for 30 min in the dark at 4°C with the
following fluorescent antibodies: anti-CD29-PE-Cy7,
anti-CD90-AlexaFluor®488, anti-CD45-PE and
anti-CD11b-AlexaFluor®647 (BioLegend, San Diego, CA,
USA). They were then washed twice with PBS, and the cells were
resuspended in 300 μl PBS. The detection of PE-Cy7/AlexaFluor
488/PE- and AlexaFluor 647-labeled cells was then carried out using
a flow cytometer (FACSCalibur flow cytometer; Becton-Dickinson,
Franklin Lakes, NJ, USA).
Transduction of BM-MSCs with GFP
Third-passage BM-MSCs at approximately 40%
confluence were seeded in fibronectin-coated 6-well plates
(Corning). The medium containing 10% FBS was removed, and the cells
were then added to the replication-defective recombinant lentiviral
vector carrying GFP (LV-GFP) supernatant containing 5 μg/ml
polybrene (Shanghai Genechem Co., Ltd., Shanghai, China), to a
final volume of 3 ml. The cells were transduced at a multiplicity
of infection (MOI) of 20 units according to the manufacturer’s
instructions. Following incubation with LV-GFP for 2 h, culture
medium containing 10% FBS was added to the plates. An additional
transduction was performed for 48 h. After the cells were washed
with PBS, the unabsorbed viral particles were removed. The BM-MSCs
were then trypsinized for 5 min and used in the following
experiments.
Induction of experimental colitis and
treatment
Morris et al developed a simple and
reproducible rat model of colonic inflammation in which they
administered the hapten, TNBS (5–30 mg), in 0.25 ml of 50% ethanol
by intraluminal instillation. They also suggested that
TNBS/ethanol-induced ulceration can persist for at least 8 weeks at
a dose of 30 mg (20). In our
study, we used TNBS (Sigma-Aldrich) to induce experimental colitis
according to the method mentioned above. Subsquently, on days 0, 3,
and 7, the GFP-transduced BM-MSCs were injected via the tail vein
at a dose of 1×106 cells in 0.3 ml PBS into the rats
with TNBS-induced colitis. In the control experiments, the animals
received 0.3 ml PBS without BM-MSCs according to the same protocol.
Stool condition and body weight, as well as activity were evaluated
daily to assess the disease activity index (DAI) as previously
described (21). On day 15, after
the mice were sacrificed, blood was collected by ventral aorta
puncture for the analysis of Tregs in peripheral blood and the
entire colon was excised. The colon was divided for histological
analysis and the evaluation of the expression of Foxp3 mRNA in
intestinal mucosa.
Histological analysis
After the rats were sacrificed, the colon samples
were fixed in 4% paraformaldehyde, embedded in paraffin and sliced
into sections (4-μm thick) before staining with hematoxylin and
eosin. Histological evaluation was completed by two expert
pathologists in a blinded manner according to a scoring scale
described previously (22). The
evaluated parameters were as follows: damage/necrosis, inflammatory
cell infiltration, submucosal edema, as well as hemorrhage of the
mucosa. Each of the parameters estimated was graded as follows: 0,
no change; 1, mild; 2, moderate; 3, severe changes. The
histological score was defined as the sum of these parameters.
Tracing of GFP-labeled BM-MSCs
To confirm the location of the infused BM-MSCs in
the inflamed colonic tissues, we transduced the BM-MSCs with
LV-GFP. Colon tissue samples excised from the colon of the inflamed
region and the non-inflamed region were embedded in optimum cutting
temperature (OCT) compound (Sakura Finetechnical Co., Ltd., Tokyo,
Japan), and then frozen on dry ice. One section was used to
detected GFP-positive cells by fluorescence confocal microscopy
(E600; Nikon, Tokyo, Japan) and the other was stained with an
antibody against GFP (Millipore, Billerica, MA, USA) and visualized
using FITC-conjugated secondary antibody (Santa Cruz Biotechnology,
Inc., Santa Cruz, CA, USA). Other sections were also prepared to
analyze GFP protein by western blot analysis. Total protein was
isolated and quantified using a BCA protein assay kit (Pierce,
Perbio Science, Tattenhall, UK). The primary antibody against GFP
was used at a dilution of 1:1,000. For the control protein loading,
anti-GAPDH was used at a dilution of 1:10,000. The protein extract
was resolved by SDS-PAGE electrophoresis and transferred onto
polyvinylidene difluoride (PVDF) membranes, which were blocked in
5% BSA and incubated with the relevant antibody.
Analysis of Tregs in peripheral blood by
flow cytometry
Peripheral blood mononuclear cells (2×106
cells/ml) were stained at 4°C for 30 min with anti-CD4-FITC and
anti-CD25-APC (all from Beckman Coulter, Brea, CA, USA). Following
incubation, the cells were fixed with Foxp3
fixation/permeabilization buffer 1 ml and permeabilized with Foxp3
permeabilization buffer 1 ml to determine intracellular Foxp3
levels. The cells were then incubated for 30 min at 4°C with
anti-Foxp3-PE antibody or isotype control (IgG2-FITC) (all from
Beckman Coulter). The stained cells were then analyzed by flow
cytometry.
mRNA expression of Foxp3 in the
colon
The colon segments were frozen in liquid nitrogen
until use at −80°C. We used TRIzol reagent (Invitrogen Corp.) to
isolate the total RNA from the individual colons according to the
manufacturer’s instructions. cDNA was synthesized from 0.5 μg of
total RNA using a reverse transcription kit (Toyobo Co., Osaka,
Japan) according to the manufacturer’s instructions. Subsequently,
a real-time PCR reaction with final volume of 20 μl using the
SYBR-Green Realtime PCR master mix (Takara Bio, Inc., Shiga, Japan)
was carried out using the ABI PRISM 7900HT sequence detector system
(Applied Biosystems, Life Technologies, Carlsbad, CA, USA). Primers
were designed according to the data from GenBank and evaluated by
nucleotide BLAST standard search to avoid cross-reactivity with
other known sequences. The designed primer sequences used for
real-time PCR were as follows: Foxp3 upstream,
5′-GGCAAACGGAGTCTGCAAG-3′ and downstream,
5′-TGCTCCAGAGACTGCACCAC-3′. The relative gene expression level,
including the levels of the target gene, normalized to the
endogenous and control gene was calculated using the comparative Ct
method 2−ΔΔCt.
Statistical analysis
Data are expressed as the means ± standard
deviation. Significant differences were assessed by the Statistical
Package for the Social Sciences (SPSS) version 18 and Microsoft
EXCEL version 2003. A p-value <0.05 was considered to indicate a
statistically significant difference. We use an unpaired t-test and
the Mann-Whitney U test for parametric and non-parametric analyses
between 2 groups.
Results
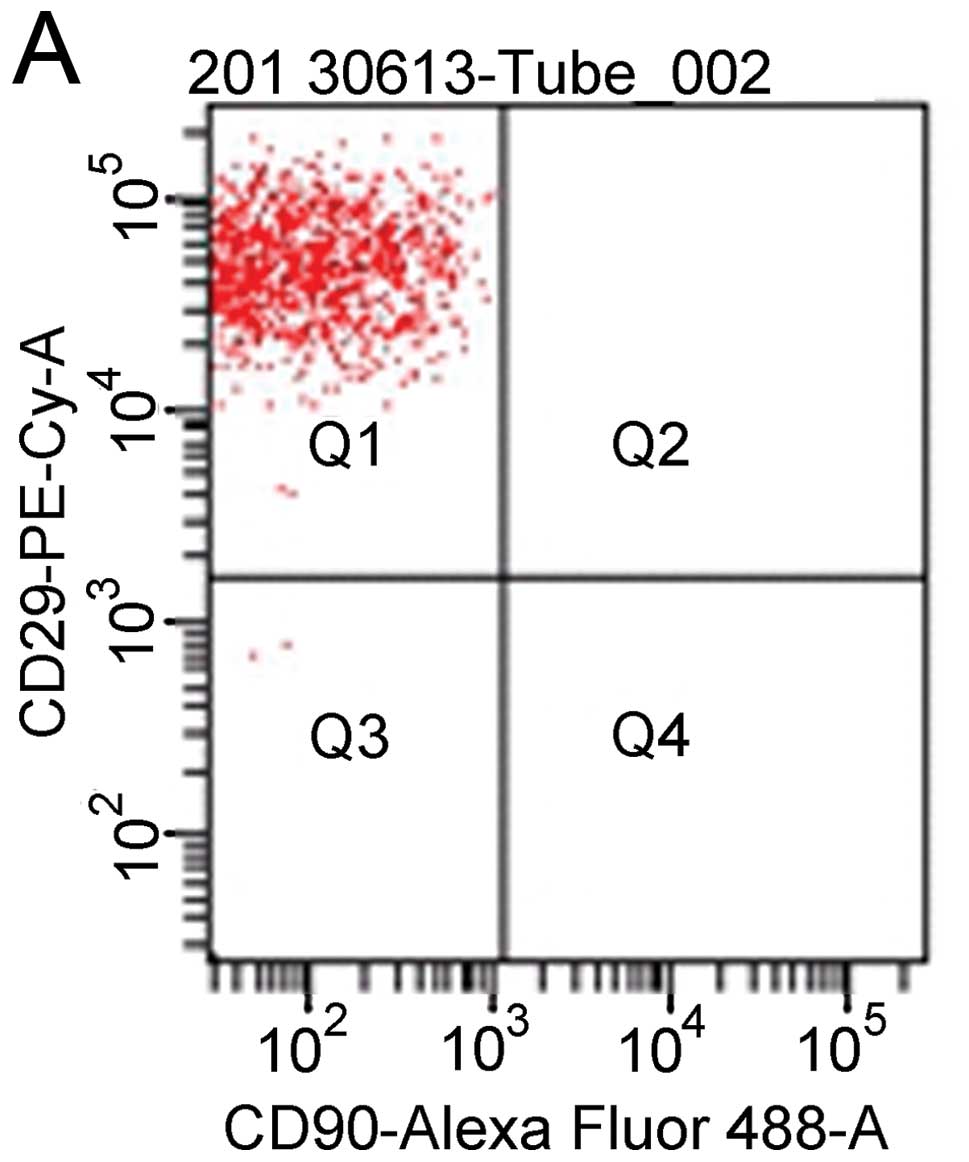
Characterization of BM-MSCs
At passage 3 or 4, the MSCs appeared to be a
homogenous population of spindle-shaped cells, such as fibroblasts
morphologically after the non-adherent cells were removed. Flow
cytometric analysis confirmed that the BM-MSCs were positive for
CD29 and CD90, but stained negative for the hematopoietic surface
markers, CD45 and CD11b (Fig.
1).
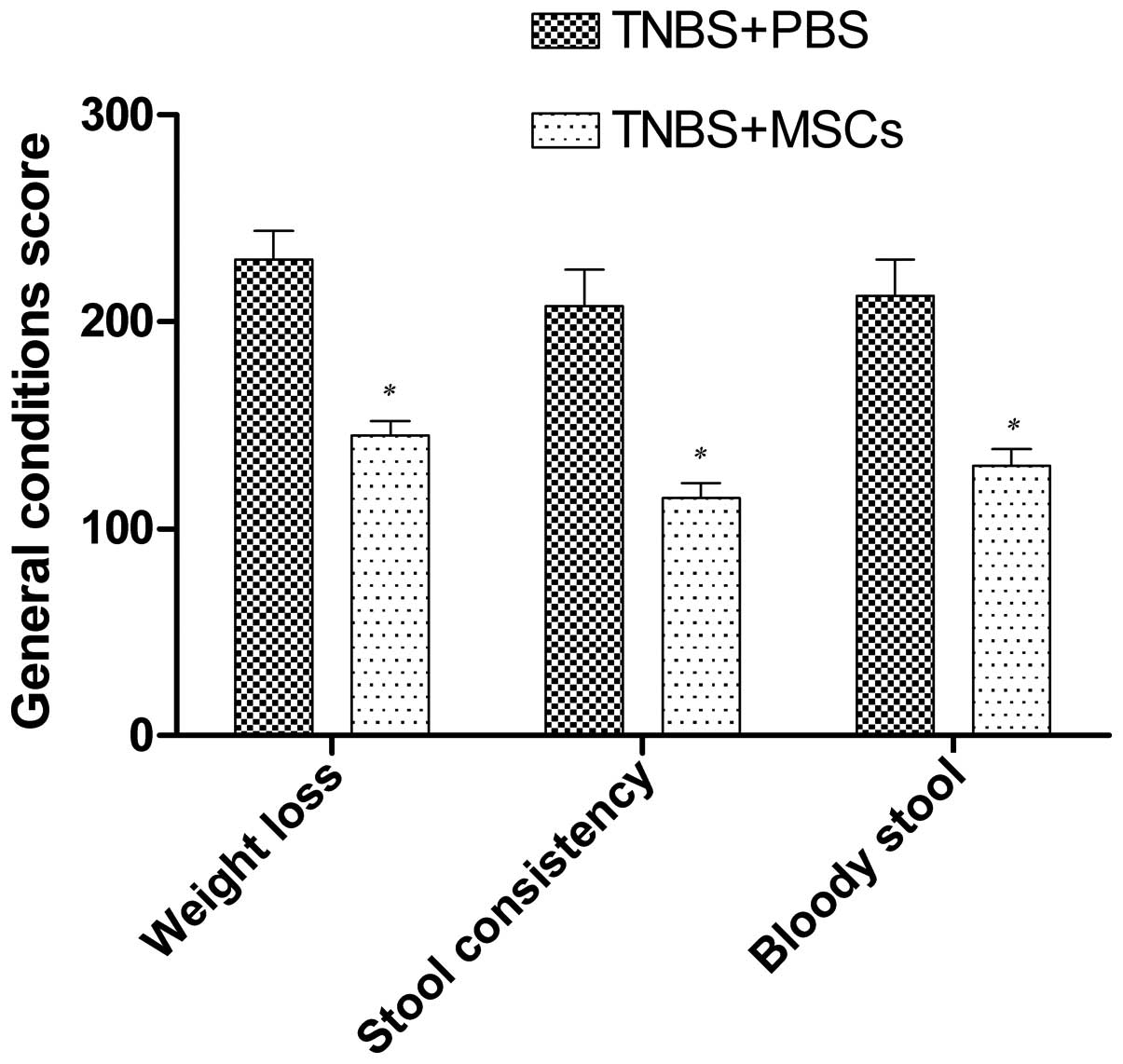
General conditions
All the TNBS-treated rats developed clinical
symptoms similar to IBD in humans 2–3 days after modeling,
including looser stool, bloody purulent stool, and loss of weight
and their activity significantly decreased. However, the rats that
were not administered TNBS did not show any of the abovementioned
signs and gained weight over time. Following the infusion of the
BM-MSCs, the symptoms of looser stool and bloody purulent stool
were significantly alleviated compared with the control group
(Fig. 2).
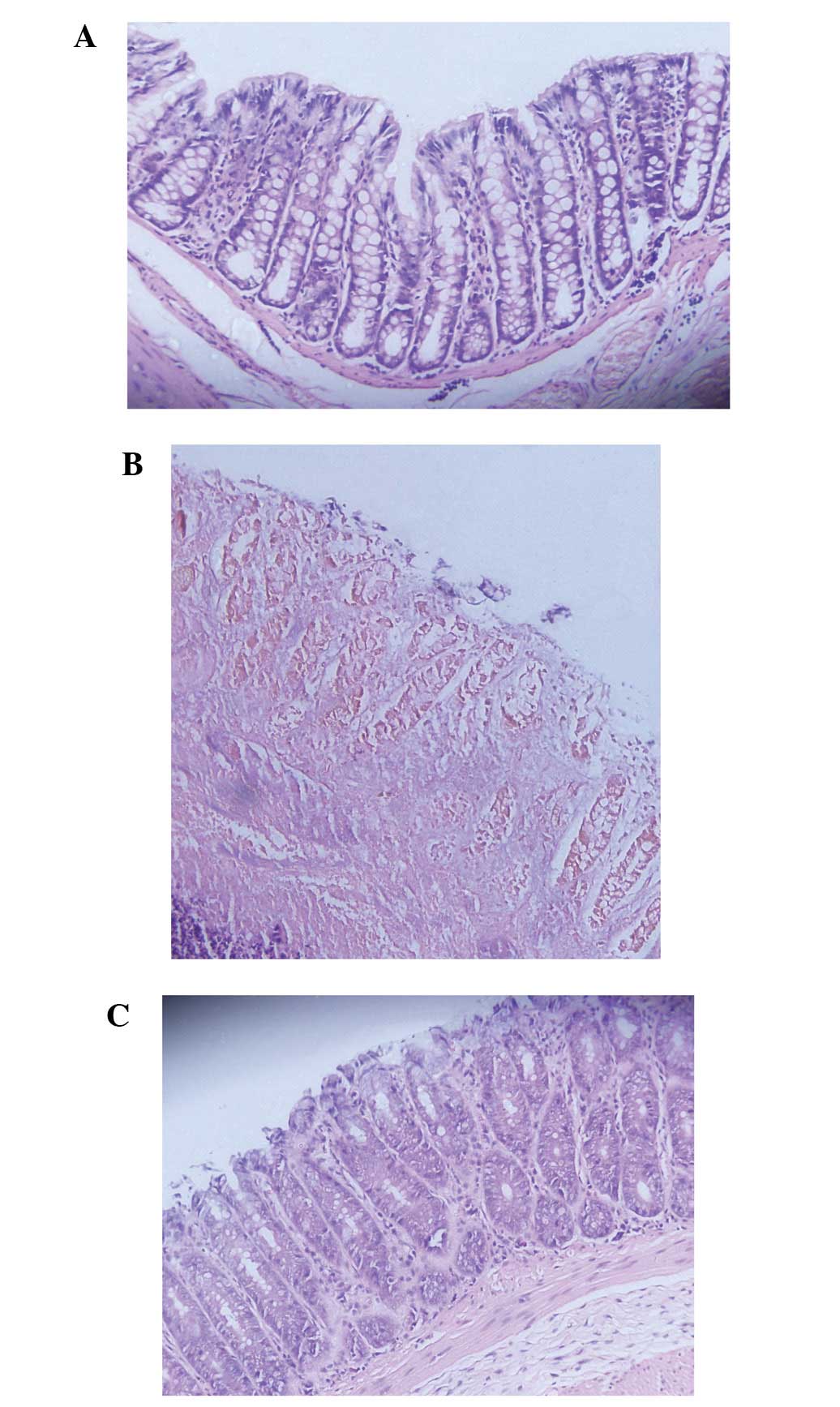
Histological improvement in the colon
following infusion of BM-MSCs
Microscopically, histological changes in the colon
samples from the rats with TNBS-induced colitis were observed. In
comparison to the control group, the rats in the MSC-treated group
showed a relatively intact structure of colonic mucosa which
consisted of more organized mucosal glands and more abundant goblet
cells, milder congestion and edema and less inflammatory cell
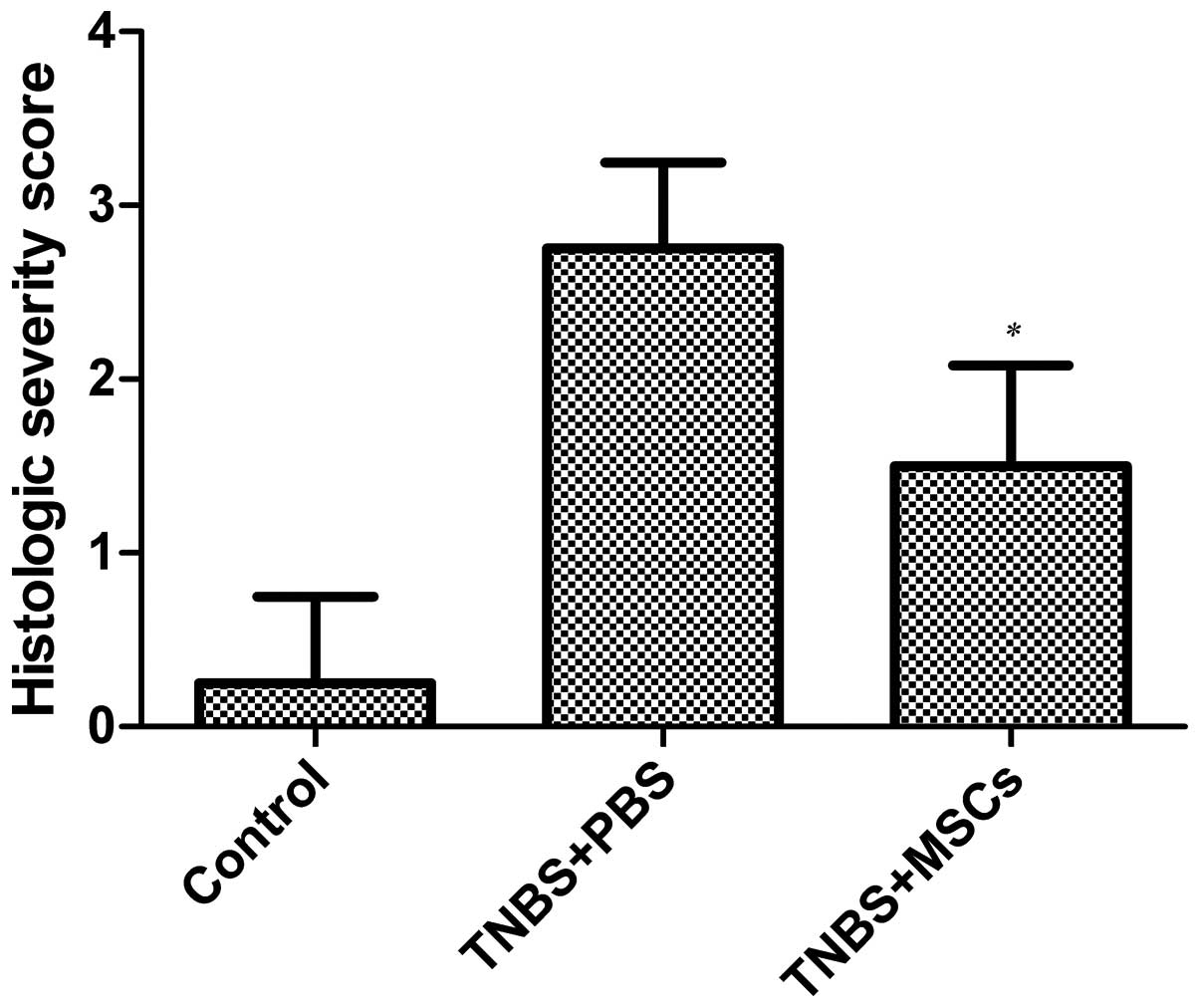
infiltration in the mucosa and submucosa (Fig. 3). The histological colitis score
defined as the sum of those parameters was significantly reduced in
the MSC-treated group (Fig.
4)
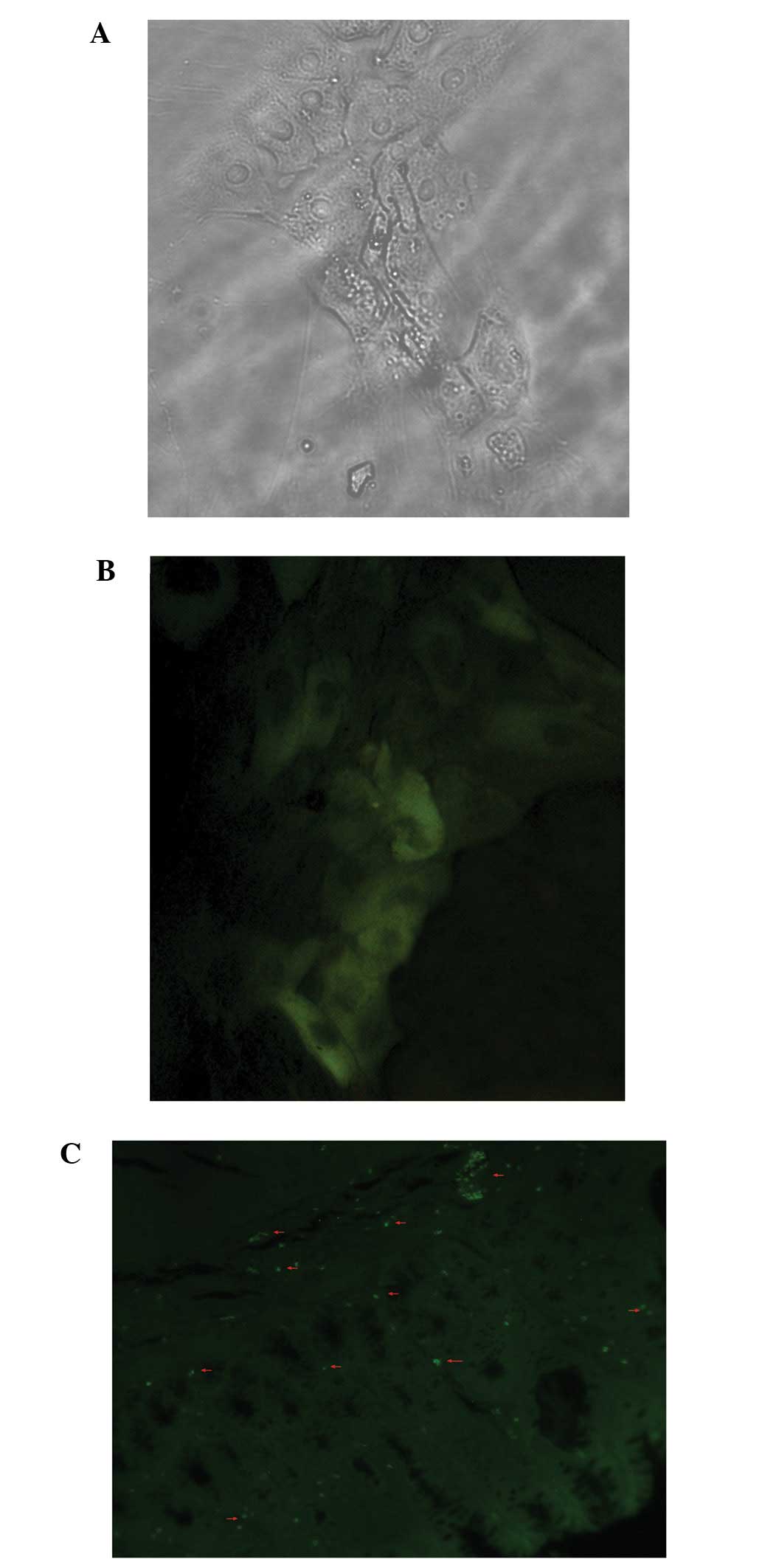
Localization of infused BM-MSCs in the
colon
In vivo, we transduced the BM-MSCs with GFP
to cofirm the localization of the infused BM-MSCs in the inflamed
colonic tissues. In vitro, a large proportion of GFP-labeled
BM-MSCs were observed (Fig. 5).
However, we did not detect GFP fluorescence after 14 days under a
confocal microscope in vivo. Distinct GFP-positive cells
were observed when we used a GFP antibody and FITC-conjugated
secondary antibody as described above, and the number of positive
cells was then observed (Fig. 5).
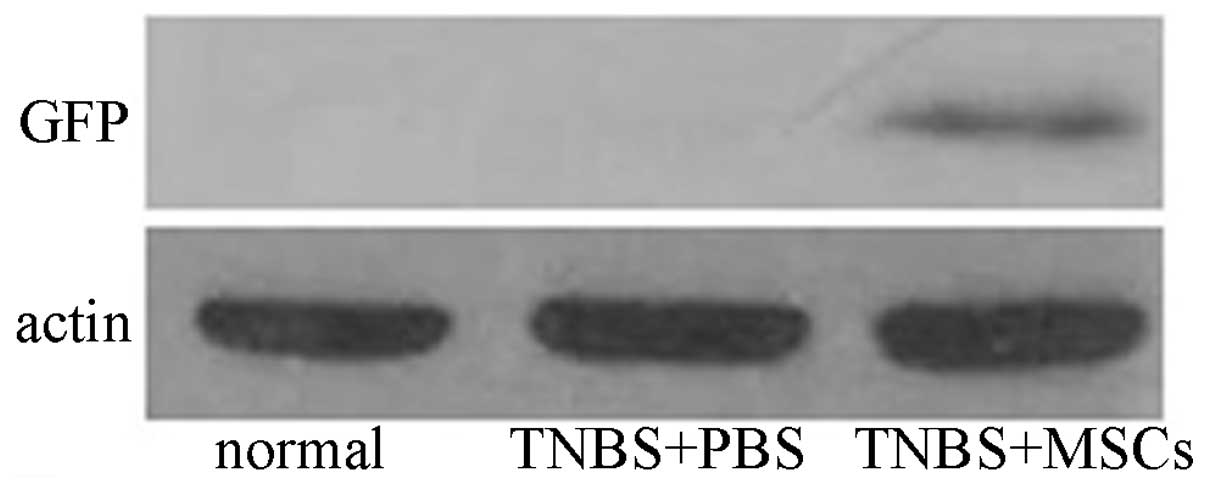
Moreover, high levels of expression of GFP protein were observed in
the GFP-labeled BM-MSC group (Fig.
6). By contrast, no immunoreactivity was detected in the
PBS-treated group and the normal control.
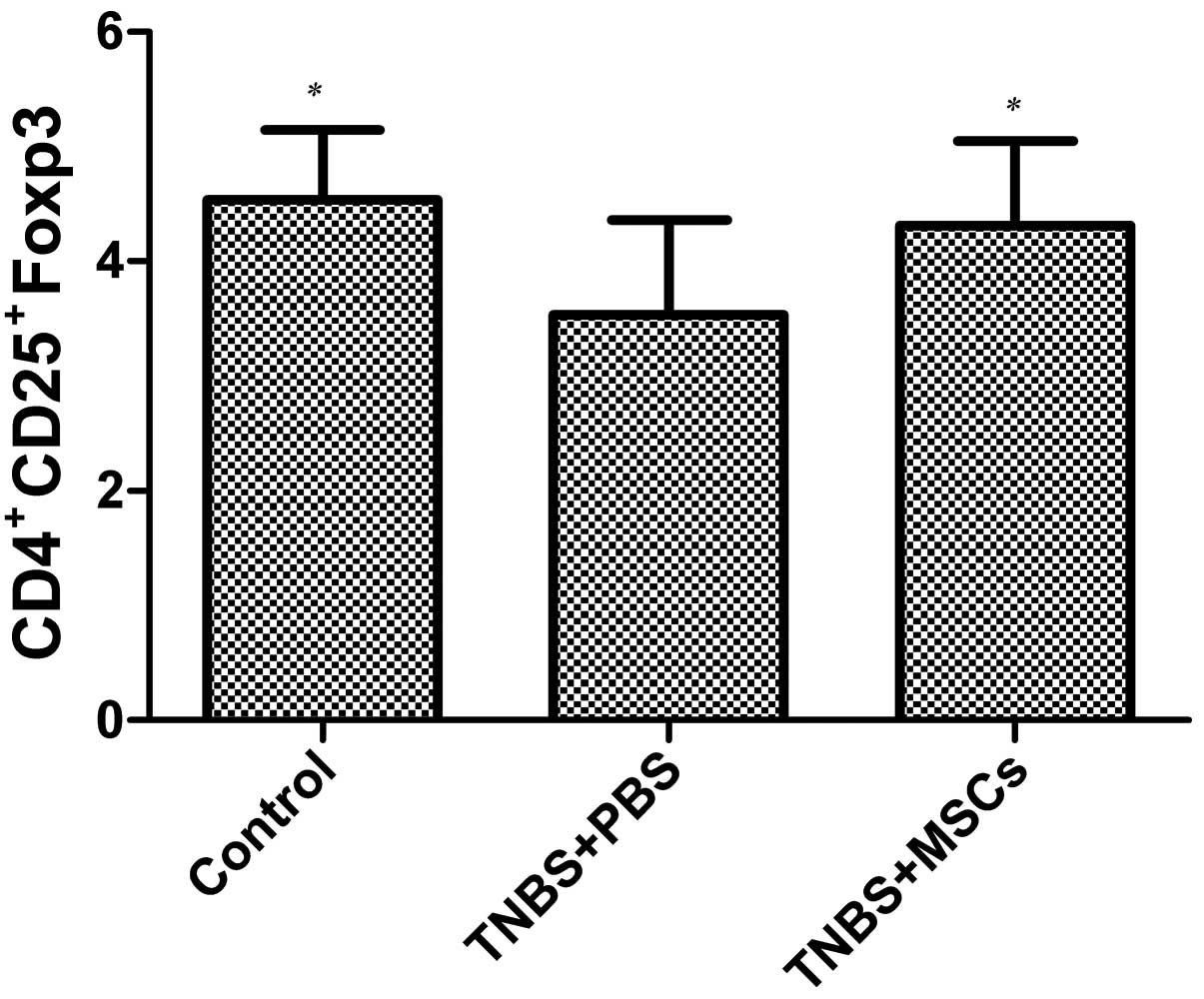
Effect of BM-MSC therapy on Tregs in
peripheral blood
A number of studies on animal models of IBD have
suggested that this disease is typically associated with a defect
in the number of Tregs and that the restoration of Tregs can
prevent and treat colitis (23,24). For this reason, in this study, we
evaluated the effects of infused BM-MSCs on Tregs in the peripheral
blood in the rats with TNBS-induced colitis. The proportion of
CD4+CD25+Foxp3 cells markedly decreased in
the PBS-treated rats with TNBS-induced colitis compared with the
control group (p<0.05), as shown by flow cytometry (Fig. 7). Following the infusion of
BM-MSCs, the proportion of CD4+CD25+Foxp3
cells incresed compared with the PBS-treated group (p<0.05)
(Fig. 7); however, no difference
was observed between the BM-MSC-treated group and the control
(untreated) group.
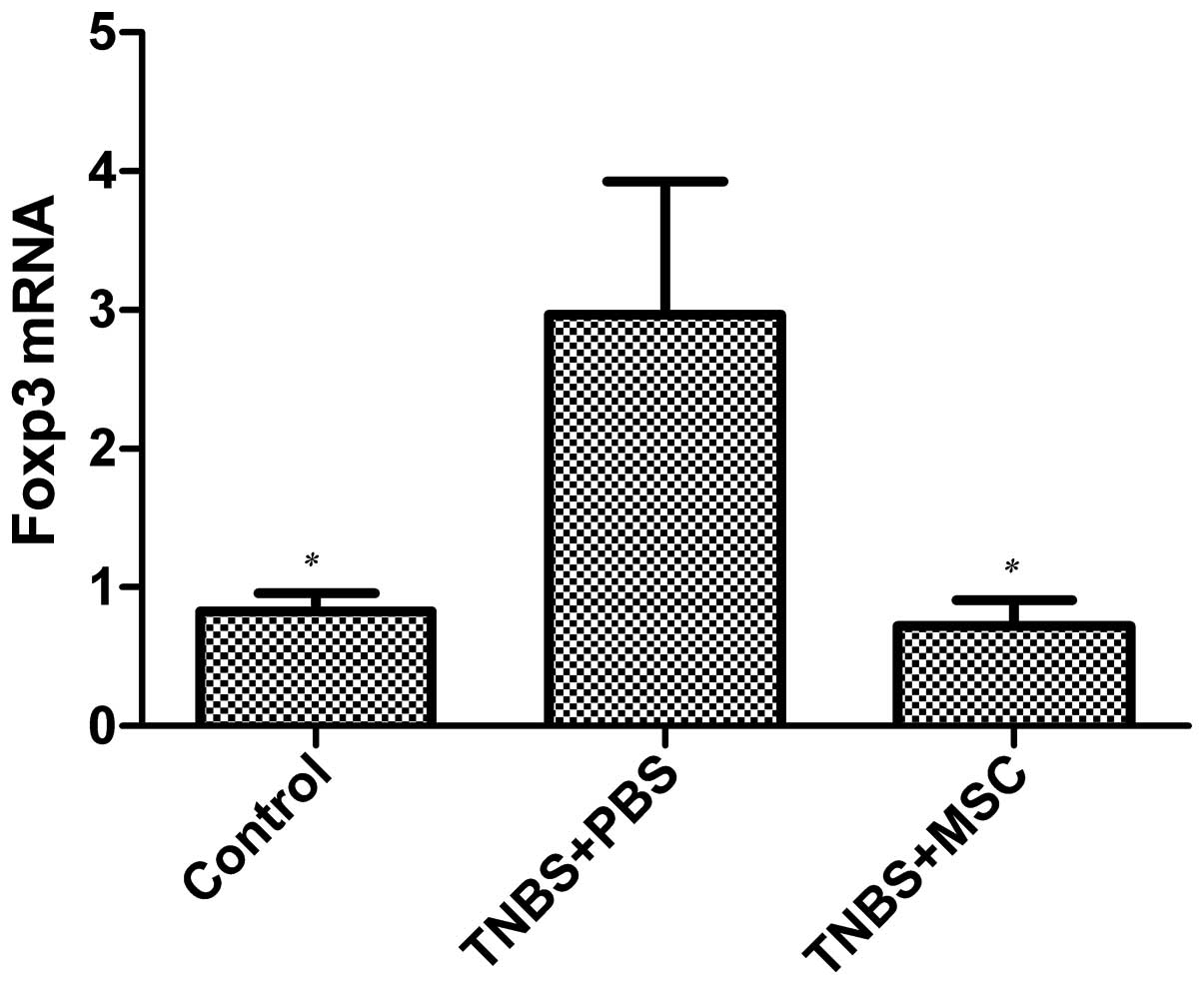
mRNA expression of Foxp3 in colonic
mucosa
The colonic mucosal mRNA expression of Foxp3 was
determined in the BM-MSC-treated rats with TNBS-induced colitis and
the PBS-treated group by real-time PCR. As shown in Fig. 8, an increase in the mRNA
expression of Foxp3 was observed in the PBS-treated group compared
with the BM-MSC-treated mice and the control group (p<0.05);
however, there was no difference in Foxp3 mRNA expression between
the BM-MSC-treated group and the control group.
Discussion
The use of animal models of mucosal inflammation in
studies on the pathogenesis of IBD was first implemented almost
half a century ago (25).
Although animal models of IBD cannot represent the human disease
completely, these types of studies are valuable and constitute
serious attempts at investigating the immunological basis of this
type of disease. In our study, we used TNBS to induce a rat model
of colitis.
The gut is an immunologically unique organ which is
in direct contact with the external environment and must retain the
ability to mount an adaptive response to commensal bacteria and
food simultaneously in order to maintain tolerance (26). The gut immune system protects the
host against pathological invasion and mediates the contact between
the host and microbes. Failure of these regulatory pathways and
imbalances of the microbiota can result in inflammatory processes,
such as IBD (27,28). A number of specialized immune
cells are involved in maintaining mucosal tolerance in the
intestine. Foxp3+ Tregs are the most significant cells
in modulating this interaction within the adaptive immune system.
One of the first studies to describe an existence of a peripheral
mechanism for the regulation of the immune system was published in
the 1960s (29). In the late
1990s, subsequent in vitro and in vivo studies
revealed the immunosuppressive properties of Tregs (30,31); however, the mechanisms involved
are complex and multifactorial. A few years later, Fontenot et
al (11) suggested that the
Foxp3 gene, which encodes the forkhead family transcription factor,
Foxp3, is the master regulatory gene for the development and
function of Tregs. In clinical practice, there is a severe
autoimmune syndrome termed immune dysregulation,
polvendocrinopathy, enteropathy, X-linked (IPEX), characterized by
a dysfunction in the development of Tregs and the consequent
activation of autoreactive T cells which is caused by mutations
within the Foxp3 gene in male infants (10,32). In addition to IPEX, the most
compelling evidence of the role of Tregs in IBD has come from the T
cell transfer model of colitis. In this model, naive
CD4+ T cells with depleted Tregs are adoptively
transferred into mice lacking B and T lymphocytes. In the
intestine, these T cells proliferate and become activated in
response to bacterial antigens, resulting in inflammation and
colitis. Thus, disease can be both induced and treated through the
elimination of the microbiota or the cotransfer of Tregs (33,34).
Interest in MSCs for their application in
transplantation was fostered not only due to their ability to
differentiate into different lineages, but also due to their
capacity to suppress the immune response (35). In recent years, the potential
application of both MSCs and Tregs in the treatment of a number of
chronic inflammatory and autoimmune diseases has gained significant
interest from immunologists worldwide, and the induction of Tregs
by activated MSCs is now a well-publicized phenomenon. In
transplantation, one of the first studies which demonstrated that
following the administration of MSCs there was an in vivo
induction of Tregs, was the study by Casiraghi et al
(36). In a subsequent study,
pre-transplant administration of donor-derived MSCs into the portal
vein in a semi-allogeneic heart transplant mouse model resulted in
T cell hyporesponsiveness, prolonged cardiac allograft survival and
expanded donor-specific Tregs expressing CD4, CD25 and Foxp3.
Further evidence that the production of Tregs by MSCs resulting
from graft survival was provided by a kidney allograft mouse model.
It was also demonstrated that there was a significant increase in
the number of intragraft Foxp3+ cells following
treatment with MSCs, through which it was suggested that Tregs
could recruit to the renal allograft (37).
In this study, we demonstrate that BM-MSCs can
ameliorate TNBS-induced colitis in rats. To the best of our
knowledge, this study is the first to demonstrate that BM-MSC
therapy affects Foxp3+ Treg cells in the blood and gut
mucosa, enhancing the treatment effects in IBD. After the rats with
colitis received exogenously infused BM-MSCs, we assessed stool,
body weight, as well as histological injury scores. Body weight and
a marked histological improvement was noted in the MSC-treated
group compared with the PBS-treated group. We hypothesize that the
beneficial effects of treatment with MSCs may be linked to the
induction of the activity of Tregs. Tregs in peripheral blood were
assayed by flow cytometry and the mRNA expression of Foxp3 in the
colon was determined by RT-PCR. Our data demonstrated that prior to
the administration of BM-MSCs, the rats with IBD had lower levels
of Foxp3+ cells in the peripheral blood compared with
the healthy controls (p<0.05) (Fig. 7), but had higher mRNA levels of
Foxp3 in the intestinal mucosa compared with the healthy controls
(p<0.05) (Fig. 8). The
decrease in the circulating population in the peripheral blood may
be due to an active pooling to the intestine and it is impossible
to repopulate the blood without ample numbers of new Tregs. Our
results on the expression of Foxp3 were similar to those from a
recent study (38). Following the
administration of BM-MSCs, the number of Foxp3+ cells in
the peripheral blood increased compared with the PBS-treated group
(p<0.05) (Fig. 7), but was
lower compared with the healthy controls. In the intestinal mucosa,
a decreased mucosal Foxp3 expression was observed in the
BM-MSC-treated group compared with the PBS-treated group
(p<0.05) (Fig. 8); there was
no significant difference between the control group and the
MSC-treated group. In conclusion, there was an inverse regulation
of mucosal and peripheral Foxp3 expression, which suggests that
MSCs redistributed the Tregs from the mucosa to the blood.
The more we understand the complex interactions
between MSCs and Tregs, the better we are likely to expore new
treatments for a number of immune diseases. However, a number of
studies have demonstrated that administered animal-derived MSCs can
enhance tumor growth in certain experimental models (39). Beyond that, there are factors,
such as the timing of delivery, the number of cells delivered, and
the site of MSC infusion, which may affect the engraftment
efficiency and the destination of exogenously delivered cells
(40). Therefore, the possible
long-term adverse effects and the details required to optimize the
protocol for MSC delivery require further and thorough
investigation. In conclusion, the interactions between
transplantated bone marrow MSCs and Tregs should be further
investigated. MSCs have tremendous potential for use in the
treatment of IBD.
Acknowledgements
Financial support for this study was provided by a
grant from the National Natural Science Foundation of China (no.
81102690)
References
|
1
|
Baumgart DC and Carding SR: Inflammatory
bowel disease: cause and immunobiology. Lancet. 369:1627–1640.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bouma G and Strober W: The immunological
and genetic basis of inflammatory bowel disease. Nat Rev Immunol.
3:521–533. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xavier RJ and Podolsky DK: Unravelling the
pathogenesis of inflammatory bowel disease. Nature. 448:427–434.
2007. View Article : Google Scholar
|
|
4
|
Baumgart DC and Sandborn WJ: Inflammatory
bowel disease: clinical aspects and established and evolving
therapies. Lancet. 369:1641–1657. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Himmel ME, Hardenberg G, Piccirillo CA, et
al: The role of T-regulatory cells and Toll-like receptors in the
pathogenesis of human inflammatory bowel disease. Immunology.
125:145–153. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fiocchi C: Towards a ‘cure’ for IBD. Dig
Dis. 30:428–433. 2012.
|
|
7
|
Shevach EM: Mechanisms of
foxp3+ T regulatory cell-mediated suppression. Immunity.
30:636–645. 2009.
|
|
8
|
Wing K and Sakaguchi S: Regulatory T cells
exert checks and balances on self tolerance and autoimmunity. Nat
Immunol. 11:7–13. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bacchetta R, Passerini L, Gambineri E, et
al: Defective regulatory and effector T cell functions in patients
with FOXP3 mutations. J Clin Invest. 116:1713–1722. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
McMurchy AN, Di Nunzio S, Roncarolo MG, et
al: Molecular regulation of cellular immunity by FOXP3. Adv Exp Med
Biol. 665:30–46. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fontenot JD, Gavin MA and Rudensky AY:
Foxp3 programs the development and function of
CD4+CD25+ regulatory T cells. Nat Immunol.
4:330–336. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mottet C, Uhlig HH and Powrie F: Cutting
edge: cure of colitis by CD4+CD25+ regulatory
T cells. J Immunol. 170:3939–3943. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yi T and Song SU: Immunomodulatory
properties of mesenchymal stem cells and their therapeutic
applications. Arch Pharm Res. 35:213–221. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Potian JA, Aviv H, Ponzio NM, et al:
Veto-like activity of mesenchymal stem cells: functional
discrimination between cellular responses to alloantigens and
recall antigens. J Immunol. 171:3426–3434. 2003. View Article : Google Scholar
|
|
15
|
Le Blanc K, Tammik C, Rosendahl K, et al:
HLA expression and immunologic properties of differentiated and
undifferentiated mesenchymal stem cells. Exp Hematol. 31:890–896.
2003.PubMed/NCBI
|
|
16
|
Engela AU, Baan CC, Dor FJ, et al: On the
interactions between mesenchymal stem cells and regulatory T cells
for immunomodulation in transplantation. Front Immunol. 3:1262012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Le Blanc K, Frassoni F, Ball L, et al:
Mesenchymal stem cells for treatment of steroid-resistant, severe,
acute graft-versus-host disease: a phase II study. Lancet.
371:1579–1586. 2008.
|
|
18
|
Dalal J, Gandy K and Domen J: Role of
mesenchymal stem cell therapy in Crohn’s disease. Pediatr Res.
71:445–451. 2012.
|
|
19
|
Sun S, Guo Z, Xiao X, et al: Isolation of
mouse marrow mesenchymal progenitors by a novel and reliable
method. Stem Cells. 21:527–535. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Morris GP, Beck PL, Herridge MS, et al:
Hapten-induced model of chronic inflammation and ulceration in the
rat colon. Gastroenterology. 96:795–803. 1989.PubMed/NCBI
|
|
21
|
Vowinkel T, Kalogeris TJ, Mori M, et al:
Impact of dextran sulfate sodium load on the severity of
inflammation in experimental colitis. Dig Dis Sci. 49:556–564.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
González R, Rodríguez S, Romay C, et al:
Anti-inflammatory activity of phycocyanin extract in acetic
acid-induced colitis in rats. Parmacol Res. 39:55–59. 1999.
|
|
23
|
Powrie F, Correaoliveira R, Mauze S, et
al: Regulatory interactions between Cd45Rb(high) and Cd45Rb(low)
Cd4(+) T-cells are important for the balance between protective and
pathogenic cell-mediated immunity. J Exp Med. 179:589–600.
1994.
|
|
24
|
Read S, Malmstrom V and Powrie F:
Cytotoxic T lymphocyte-associated antigen 4 plays an essential role
in the function of CD25(+)CD4(+) regulatory cells that control
intestinal inflammation. J Exp Med. 192:295–302. 2000.
|
|
25
|
Strober W: Animal models of inflammatory
bowel disease - an overview. Dig Dis Sci. 30:3S–10S. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Neuman MG: Signaling for inflammation and
repair in inflammatory bowel disease. Rom J Gastroenterol.
13:309–316. 2004.PubMed/NCBI
|
|
27
|
Reiff C and Kelly D: Inflammatory bowel
disease, gut bacteria and probiotic therapy. Int J Med Microbiol.
300:25–33. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cho JH: The genetics and
immunopathogenesis of inflammatory bowel disease. Nat Rev Immunol.
8:458–466. 2008. View
Article : Google Scholar
|
|
29
|
Chaperon EA, Selner JC and Claman HN:
Migration of antibody-forming cells and antigen-sensitive
precursors between spleen, thymus and bone marrow. Immunology.
14:553–561. 1968.PubMed/NCBI
|
|
30
|
Sakaguchi S, Sakaguchi N, Asano M, et al:
Immunologic self-tolerance maintained by activated T cells
expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single
mechanism of self-tolerance causes various autoimmune diseases. J
Immunol. 155:1151–1164. 1995.
|
|
31
|
Takahashi T, Kuniyasu Y, Toda M, Sakaguchi
N, Itoh M, Iwata M, et al: Immunologic self-tolerance maintained by
CD25+CD4+ naturally anergic and suppressive T
cells: induction of autoimmune disease by breaking their
anergic/suppressive state. Int Immunol. 10:1969–1980. 1998.
|
|
32
|
d’Hennezel E, Bin Dhuban K, Torgerson T,
et al: The immunogenetics of immune dysregulation,
polyendocrinopathy, enteropathy, X linked (IPEX) syndrome. J Med
Genet. 49:291–302. 2012.PubMed/NCBI
|
|
33
|
Maloy KJ: Induction and regulation of
inflammatory bowel disease in immunodeficient mice by distinct
CD4+ T-cell subsets. Methods Mol Biol. 380:327–335.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Eri R, McGuckin MA and Wadley R: T cell
transfer model of colitis: a great tool to assess the contribution
of T cells in chronic intestinal inflammation. Methods Mol Biol.
844:261–275. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tolar J, Le Blanc K, Keating A and Blazar
BR: Concise review: hitting the right spot with mesenchymal stromal
cells. Stem Cells. 28:1446–1455. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Casiraghi F, Azzollini N, Cassis P, et al:
Pretransplant infusion of mesenchymal stem cells prolongs the
survival of a semiallogeneic heart transplant through the
generation of regulatory T cells. J Immunol. 181:3933–3946. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ge W, Jiang J, Arp J, et al: Regulatory
T-cell generation and kidney allograft tolerance induced by
mesenchymal stem cells associated with indoleamine 2,3-dioxygenase
expression. Transplantation. 90:1312–1320. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li Z, Arijs I, De Hertogh G, et al:
Reciprocal changes of Foxp3 expression in blood and intestinal
mucosa in IBD patients responding to infliximab. Inflamm Bowel Dis.
16:1299–1310. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Djouad F, Plence P, Bony C, et al:
Immunosuppressive effect of mesenchymal stem cells favors tumor
growth in allogeneic animals. Blood. 102:3837–3844. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Karp JM and Leng Teo GS: Mesenchymal stem
cell homing: the devil is in the details. Cell Stem Cell.
4:206–216. 2009. View Article : Google Scholar : PubMed/NCBI
|