Introduction
Endothelial progenitor cells (EPCs) originate in the
bone marrow. They are recruited into peripheral circulation in
response to tissue ischemia or back to their site of origin to
areas of injured endothelium. In this manner, they participate in
the repair of damaged tissues, thereby improving blood flow and
attenuating the progression of atherosclerosis (1). Since both animal models and human
studies have shown that EPCs can contribute to neovascularization
and re-endothelialization, EPCs have been examined as potential
treatments for various types of ischemic disease, including stroke
(2), ischemic myocardium
(3,4), kidney injury (5), and ischemic vascular disease
(6). Moreover, it has been
reported that a low EPC level is an independent risk factor for
future cardiovascular events such as unstable angina and myocardial
infarction (MI) (7,8). MI (9) and limb ischemia (10,11) themselves have been associated with
an increase in circulating EPC numbers, and vascular traumas such
as acute MI with ST elevation (12) and percutaneous coronary
intervention (13) have been
suggested to induce the rapid mobilization of EPCs.
Evidence from in vitro and clinical studies
suggested that inflammatory and oxidative changes can influence EPC
apoptosis (14,15). Additionally, tumor necrosis
factor-α (TNF-α) is a contributing risk factor in atherosclerosis
and common metabolic disturbances including insulin resistance and
dyslipidemia (16). More
specifically, TNF-α plays a key role as both an inflammatory
mediator and inducer of apoptosis in endothelial cells (17,18). Evidence of these roles can be
found in the increased plasma levels of TNF-α in diabetic patients,
which have been shown to impair the function of endothelial cells,
as well as to enhance their aging and apoptosis (19–21). A previous in vitro study
has shown that incubation of EPCs with TNF-α increases
p38-phosphorylation, resulting in a reduction of total EPCs
(22). Clinically, statins are
widely used in the treatment of dyslipidemia and associated
vascular abnormalities. In fact, clinical trials with statins have
shown considerable benefits in patients with ischemic heart and
peripheral disease, irrespective of their cholesterol concentration
(23). Evidence from in
vitro and in vivo data have demonstrated that statins
exert pleiotropic actions beyond their lipid-lowering effects,
including immune regulation, anti-inflammatory effects, maintenance
of plaque stability, and anti-platelet effects, in addition to
anti-fibrotic and anti-oxidant properties (24–27).
Silent information regulator type-1 (SIRT1) is a
member of the sirtuin family of nicotinamide adenine dinucleotide
(NAD)-dependent class III histone deacetylases (HDACs). Previous
studies have shown that SIRT1 promotes cell survival by repressing
p53-dependent apoptosis in response to DNA damage and oxidative
stress. This repression occurs via physical interaction with both
p53 and the forkhead transcription factor (FOXO) family of proteins
(28–30). Additionally, evidence suggests
that SIRT1 may be involved in pathways of telomere maintenance,
giving them a putative role in aging and survival (31). It has been reported that statins
can activate SIRT1, which raises the possibility of their
epigenetic regulation of inflammatory responses. Moreover, SIRT1
plays a key role in the prevention of endothelial apoptosis and
senescence through its direct effect on endothelial nitric oxide
synthase (eNOS) activation (32,33). However, there is little evidence
that statins can attenuate TNF-α-induced apoptosis in EPCs.
Furthermore, the possible anti-apoptotic effects of SIRT1 are
currently unclear.
Thus, the present study aimed to clarify the roles
of statin application and activation of SIRT1 on EPC apoptosis. The
results showed that simvastatin, one of the most commonly used
statins, is capable of reducing TNF-α-induced apoptosis in EPCs,
and that SIRT1 may play a critical role in the prevention of EPC
apoptosis.
Materials and methods
Materials
TNF-α was obtained from PeproTech, Inc. (Rocky Hill,
NJ, USA). 3-(4,5-Dimethylthiazol-2-yl)-2,5- diphenyltetrazolium
bromide (MTT) was purchased from Sigma-Aldrich (St. Louis, MO,
USA). SIRT1 antibody was obtained from Cell Signaling Technology,
Inc. (Beverly, MA, USA). The FITC Annexin V Apoptosis Detection kit
was purchased from BD Pharmingen (San Diego, CA, USA). EGM-2MV was
obtained from Clonetics (San Diego, CA, USA), Lymphoprep™ was
purchased from Axis-shield (Oslo, Norway), and Matrigel from BD
Biosciences (San Diego, CA, USA).
Cell culture
EPCs derived from human umbilical cord blood were
obtained by Lymphoprep™ density gradient centrifugation at 800 × g,
for 20 min at a temperature of 4°C. Following centrifugation, the
mononuclear cells (MNCs) layer was harvested and washed twice in
0.9% saline. The MNCs were cultivated in endothelial
differentiation medium (EGM-2MV), which contained 5% fetal bovine
serum, VEGF, fibroblast growth factor-2, epidermal growth factor,
insulin-like growth factor-1 and ascorbic acid. Cell cultures were
maintained at 37°C with 5% CO2 and 21% O2 in
a humidified atmosphere. Three days after plating, the non-adherent
cells were discarded and fresh medium was applied. To maintain
optimal culture conditions, the medium was changed on alternate
days. The cells were selected for in vitro study after three
total passages.
Western blot analysis
EPCs were homogenized in phosphate-buffered saline
containing a protease inhibitor cocktail (50 mM Tris, pH 7.6, 150
mM NaCl, 1 mM EDTA, 1% Triton X-100, 0.5% sodium deoxycholate, 0.1%
SDS, and 10% glycerol). The samples were incubated overnight at 4°C
with rabbit anti-SIRT1 (1:500) (Cell Signaling Technology, Inc.).
Antibody signal was detected using a Chemiluminescent Detection kit
according to the manufacturer’s protocol (Beyotime, Haimen, China).
The results were normalized to GAPDH. The relative band intensities
of the blots were determined with Adobe Photoshop software.
Apoptosis analysis
After treatment for 48 h, EPCs were harvested,
washed in ice-cold PBS, resuspended in 500 μl of binding buffer,
and incubated in the dark with 5 μl of propidium iodide (PI) and 5
μl of Annexin V-fluorescein isothiocyanate for 15 min. The samples
were washed and resuspended in 500 μl PBS, before analyzing with
flow cytometry.
MTT growth curve
EPCs were detached and seeded into 96-well plates
(4,000 cells per well). At the indicated time-points (24, 48 and 72
h after culture), the cells were stained with MTT (5 mg/ml) in PBS
for 3 h, and then dissolved with 50% N,N-dimethylformamide and 10%
SDS for 3 h at 37°C. The optical density at 570 nm was then
determined. Each point was determined in triplicate and an average
was obtained for analysis.
Tube formation assay
To demonstrate capillary tube formation, 250 μl of
Matrigel was added to 24-well plates and solidified for 30 min at
37°C. EPCs were then seeded (5×104 cells/well) on the
solidified Matrigel and suspended in 300 μl of EGM2-MV medium.
After 12- and 24-h incubations, the cells were incubated at 37°C
with 5% CO2 for 5–24 h.
Statistical analysis
Data are expressed as mean ± SD. An unpaired
Student’s t-test was used to evaluate statistical differences
between the groups. Differences were considered significant at a
value of P<0.05.
Results
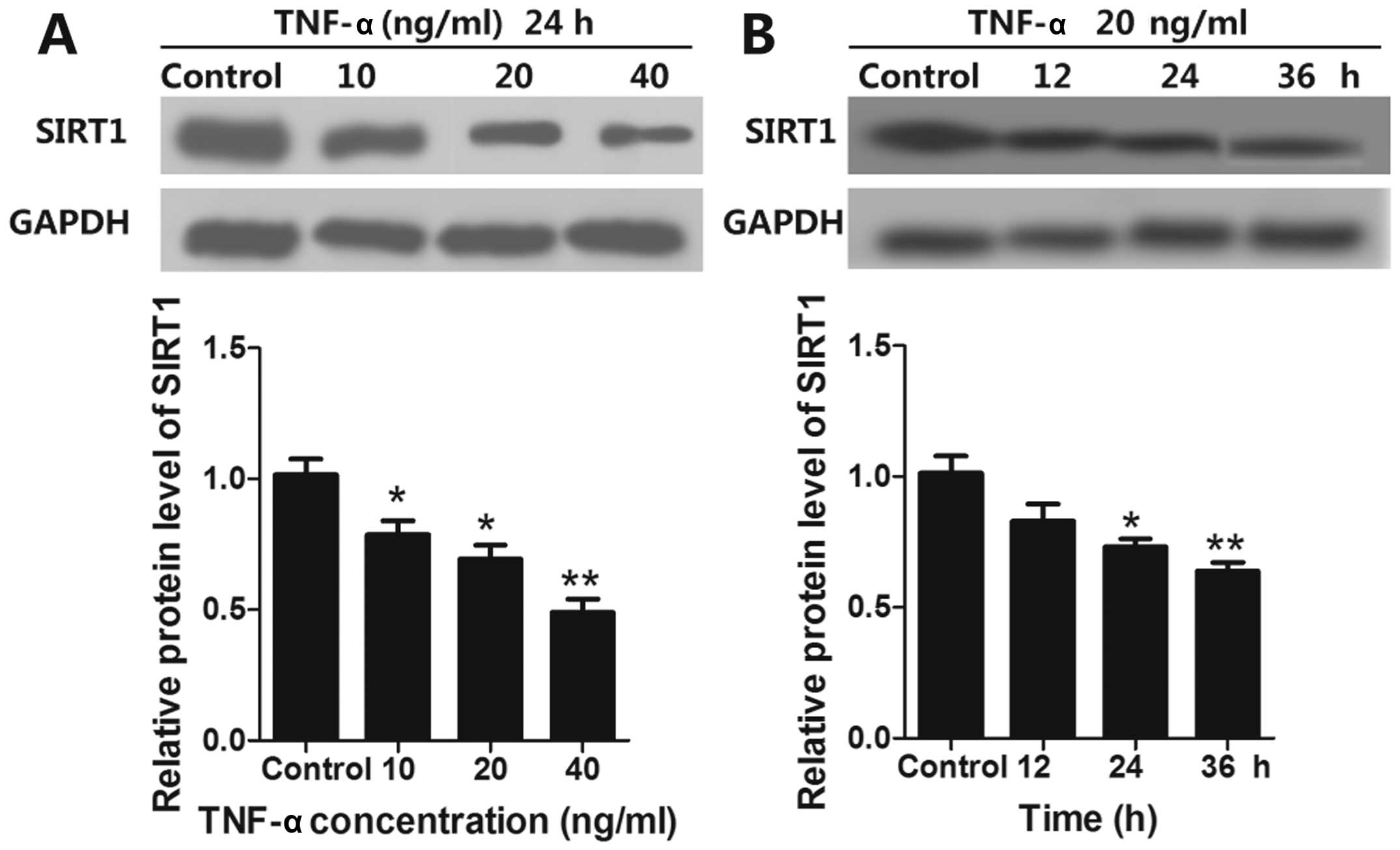
TNF-α induces apoptosis and decreases
SIRT1 expression of EPCs
To investigate whether SIRT1 expression could be
regulated by TNF-α in EPCs, EPCs were treated with TNF-α (10, 20
and 40 ng/ml) for 24 h and the expression of SIRT1 was determined
by western blot analysis. We found that TNF-α reduced SIRT1 protein
expression in EPCs in a concentration-dependent manner (Fig. 1A). We also studied the time course
of SIRT1 expression in EPCs by treating EPCs with TNF-α (20 ng/ml)
for 12, 24 and 36 h. Our results showed that SIRT1 protein
expression decreased in a time-dependent manner when compared with
the control (Fig. 1B).
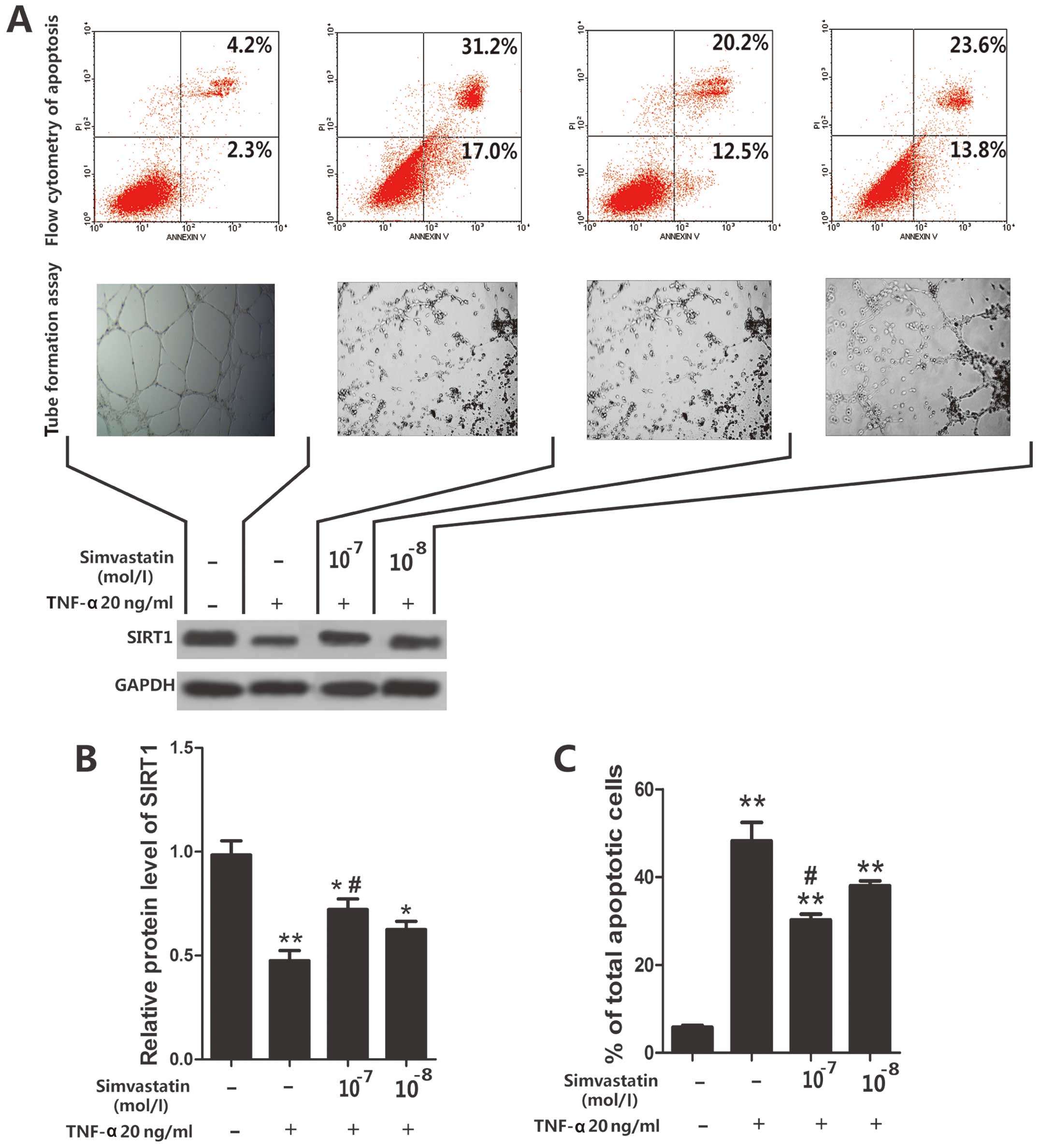
Simvastatin inhibits
TNF-α-induced-apoptosis of EPCs
EPCs were treated with TNF-α (20 ng/ml) for 24 h and
two groups of EPCs were then selected and treated with two
concentrations of simvastatin (10−7 or 10−8
mol/l). Cell apoptosis was then evaluated via flow cytometry. The
results showed that TNF-α caused a marked increase in the
percentage of apoptotic EPCs. However, application of simvastatin
decreased the number of apoptotic cells when compared with cells
treated solely with TNF-α. Moreover, we found that a higher
concentration of simvastatin had a greater reduction in apoptotic
cell number (P<0.05) (Fig. 2A and
C).
Simvastatin increases TNF-α-induced
decreases in SIRT1 levels
To investigate whether the TNF-α-induced decrease of
SIRT1 expression could be raised by simvastatin application, EPCs
were treated with TNF-α and a high or low concentration simvastatin
(10−7 or 10−8 mol/l). Expression of SIRT1 was
then determined by western blot analysis. We found that a high
concentration of simvastatin resulted in a marked increase in SIRT1
expression levels when compared to the TNF-α-only group (P<0.05)
(Fig. 2A and B). By contrast, we
found that a low concentration of simvastatin resulted in a less
obvious increase.
Simvastatin cannot restore the
angiopoietic ability of TNF-α-treated EPCs
The angiopoietic ability of simvastatin was assessed
using the tube formation assay. We found that the angiopoietic
ability of EPCs was significantly decreased by TNF-α (20 ng/ml, 24
h). We also found that two different concentrations of simvastatin
(10−7 or 10−8 mol/l) were not able to restore
the angiopoietic ability of the EPCs.
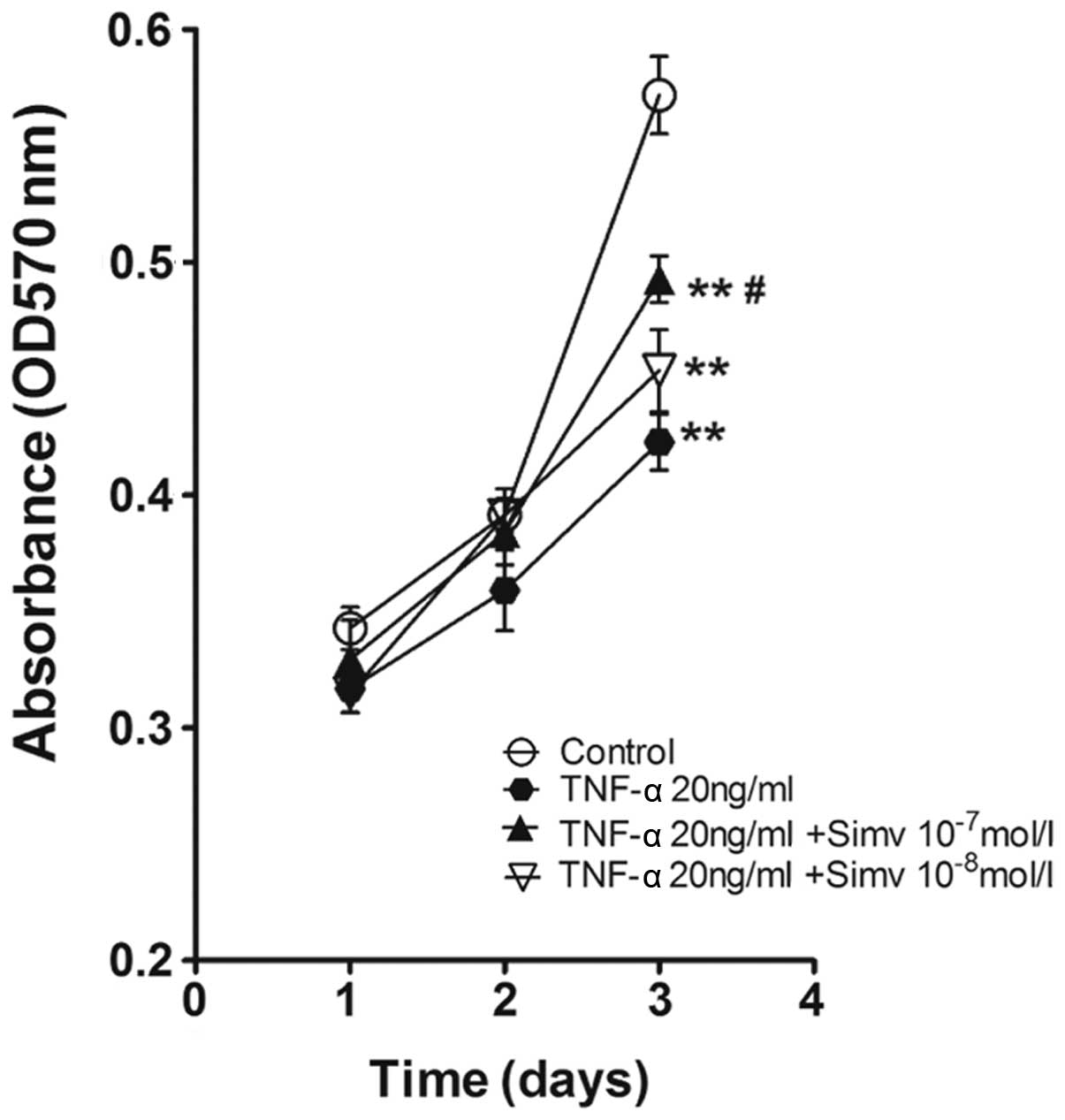
Simvastatin promotes cell proliferation
that was reduced by TNF-α application
The cell viability of EPCs was evaluated by an MTT
assay. As shown in Fig. 3, the
proliferation of EPCs treated with TNF-α (20 ng/ml) was markedly
inhibited when compared to the controls (P<0.01). High
concentrations of simvastatin were able to partially enhance cell
proliferation when compared to the TNF-α-only group (P<0.05). No
differences were observed in cell proliferation between the TNF-α
group and low concentrations of simvastatin.
Discussion
EPCs are critical for angiogenesis in ischemic
disease and inflammation. Numerous animal studies have suggested
that the vaso- and atheroprotective effects of EPCs are associated
with their cell replacement and non-cellular differentiation
effects. These include trophic support and enhancement of the
endogenous repair process (3,34,35). EPCs are heterogeneous and can be
classified into early or late EPCs. In ex vivo culture
systems, early EPCs appear within 4–7 days, while late EPCs develop
after 2–3 weeks. It has been hypothesized that early EPCs may
primarily provide trophic support, while late EPCs, which express a
variety of endothelial markers, differentiate into mature
endothelial cells and contribute to vascular repair (36). Considerable evidence indicates
that a reduction in the number of EPCs predicts future
cardiovascular events. Thus, enhancement of the number of EPCs is
of potential therapeutic benefit to individuals with cardiovascular
diseases. In this study, late EPCs were selected as our subject due
to their role in angiogenesis and proliferation (37).
Statins are beneficial for atherosclerosis via
mechanisms independent of their lipid-lowering ability. For
instance, increased EPC levels in the early post-infarction phase
by statin treatment were associated with improved cardiac function
and increased capillary density in the peri-infarct area after a MI
(38). A number of mechanisms
have been suggested by which statins may attenuate atherosclerosis
disease and cellular senescence in EPCs. Several of these
mechanisms involve the ability of statin therapy to enhance bone
marrow VEGF protein levels, their ability to phosphorylate Akt,
eNOS activity, and their ability to decrease oxidative DNA damage
and prevent telomere shortening due to oxidative stress in EPCs
(38–40). As a powerful factor in
inflammatory stimulation, TNF-α has been reported to induce
apoptosis and senescence of EPCs both in vitro and in
vivo. However, there are no previous studies focusing on the
ability of statins to attenuate TNF-α-induced apoptosis in EPCs
(41). In this study,
proliferation, apoptosis and angiogenesis of EPCs co-cultured with
TNF-α and simvastatin were examined. We found that simvastatin
exerts a mild anti-apoptotic effect on EPCs treated with TNF-α and
that it enhances proliferation of EPCs that were previously
inhibited by TNF-α incubation (Fig.
3). We found no effect of simvastatin treatment on the
restoration of angiopoiesis (Fig.
2).
SIRT1 was originally identified as a nuclear protein
that deacetylates proteins, contributing to cell regulation.
Increasing SIRT1 allows for greater cell survival, particularly
during periods of stress that usually trigger apoptosis. This
occurs through regulation of the activity of several proteins,
including FOXO, p53 and Ku70, all of which are involved in
apoptosis or the initiation of cell repair (42–44). SIRT1 is located in the nucleus and
translocated into the cytoplasm in response to hydrogen peroxide.
This translocation results in an increased sensitivity to apoptosis
(45). TNF-α is the prototypical
member of a family of cytokines that also includes FasL, CD40L, and
TRAIL. TNF-α is a potent inducer, which triggers inflammation,
apoptosis, differentiation, and cell activation. TNF-α is found in
the extracellular matrix, endothelium, and vessel walls of
fibro-vascular tissue of proliferative diabetic retinopathy
(46). Although SIRT1 levels can
alter as a result of TNF-α exposure, its effects may differ
depending on cell type, concentration, and time course. For
instance, Wang et al (47)
found that SIRT1 was highly expressed in the nucleus of vascular
adventitial fibroblasts (VAFs) and translocated into the cytoplasm
in response to TNF-α. Dvir-Ginzberg et al (48) also found that SIRT1 expression was
reduced in human chondrocytes that had been stimulated with TNF-α
(50 mg/ml) for 24 h. Conversely, results of a recent study showed
that the expression levels of SIRT1 mRNA and protein were markedly
increased in vascular smooth muscle cells (VSMCs) treated with
TNF-α at concentrations of 30 and 50 mg/ml for 8 h (49). Similarly, we found that TNF-α
reduced SIRT1 protein expression levels in EPCs in a concentration-
and time-dependent manner (Fig.
1). Our results have shown that the ability of angiopoiesis and
proliferation of EPCs was markedly inhibited by TNF-α. This
conclusion was based on our data, which showed that incubation of
EPCs with TNF-α significantly increased the percentage of apoptotic
EPCs and simultaneously decreased the expression of SIRT1 (Fig. 2). Thus, we conclude that SIRT1 is
involved in TNF-α-induced apoptosis in EPCs.
However, the molecular mechanism through which
statins exert their anti-apoptotic effect is unclear. Although we
found that the expression of SIRT1 can be decreased by TNF-α and
increased by simvastatin, additional experiments are required to
identify the molecular mechanism between statins, SIRT1, and TNF-α.
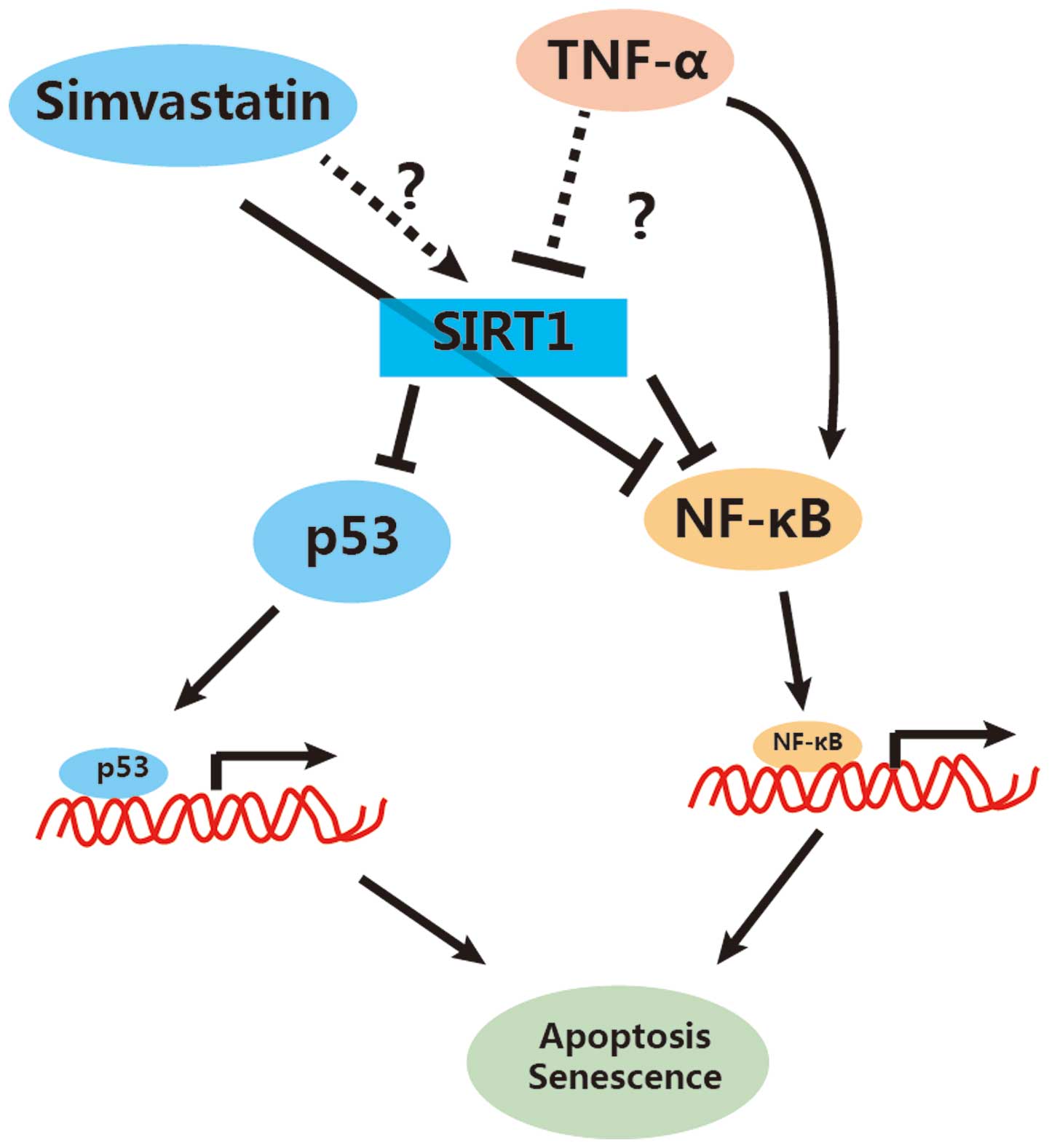
Based on our results and the available literature, we have proposed
a model demonstrating the role of simvastatin in apoptosis of EPCs
(Fig. 4). We suggest that
simvastatin inhibits cell apoptosis, in part, via the elevation of
SIRT1. This elevation consequently inactivates NF-κB activity by
decreasing its acetylation state (NF-κB-Ac). Additionally,
simvastatin may inhibit NF-κB activity directly, as has been
previously reported in the literature (50,51). Moreover, simvastatin-dependent
SIRT1 increases could also decrease the activity of p53 by
decreasing its acetylation, thus inhibiting the p53-dependent
apoptosis.
In summary, we have shown that simvastatin plays an
important role in the TNF-α-induced apoptosis of EPCs and that this
protection may involve SIRT1. These findings provide new evidence
for the anti-inflammatory role of statins, which surpass their
known effects on cholesterol metabolism.
Acknowledgements
This study has been supported by the Key Subject
Construction of The First Affiliated Hospital of Jinan University,
China.
References
|
1
|
Rouhl RP, van Oostenbrugge RJ, Damoiseaux
J, Tervaert JW and Lodder J: Endothelial progenitor cell research
in stroke: a potential shift in pathophysiological and
therapeutical concepts. Stroke. 39:2158–2165. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Decano JL, Moran AM, Giordano N,
Ruiz-Opazo N and Herrera VL: Analysis of CD45- [CD34+/KDR+]
endothelial progenitor cells as juvenile protective factors in a
rat model of ischemic-hemorrhagic stroke. PLoS One.
8:e552222013.
|
|
3
|
Huang H, Huang F and Huang JP:
Transplantation of bone marrow-derived endothelial progenitor cells
overexpressing Delta-like-4 enhances functional neovascularization
in ischemic myocardium. Mol Med Rep. 8:1556–1562. 2013.
|
|
4
|
Thal MA, Krishnamurthy P, Mackie AR, et
al: Enhanced angiogenic and cardiomyocyte differentiation capacity
of epigenetically reprogrammed mouse and human endothelial
progenitor cells augments their efficacy for ischemic myocardial
repair. Circ Res. 111:180–190. 2012. View Article : Google Scholar
|
|
5
|
Patschan D, Hildebrandt A, Rinneburger J,
et al: The hormone melatonin stimulates renoprotective effects of
‘early outgrowth’ endothelial progenitor cells in acute ischemic
kidney injury. Am J Physiol Renal Physiol. 302:F1305–F1312.
2012.PubMed/NCBI
|
|
6
|
Bouchentouf M, Forner K, Cuerquis J, et
al: A novel and simplified method of culture of human blood-derived
early endothelial progenitor cells for the treatment of ischemic
vascular disease. Cell Transplant. 20:1431–1443. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Głowińska-Olszewska B, Luczyński W and
Bossowski A: Endothelial progenitor cells as a new marker of
endothelial function with respect to risk of cardiovascular
disorders. Postepy Hig Med Dosw (Online). 65:8–15. 2011.(In
Polish).
|
|
8
|
Shantsila E, Watson T and Lip GY:
Endothelial progenitor cells in cardiovascular disorders. J Am Coll
Cardiol. 49:741–752. 2007. View Article : Google Scholar
|
|
9
|
Sun JY, Zhai L, Li QL, et al: Effects of
ACE inhibition on endothelial progenitor cell mobilization and
prognosis after acute myocardial infarction in type 2 diabetic
patients. Clinics (Sao Paulo). 68:665–673. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sandri M, Beck EB, Adams V, et al: Maximal
exercise, limb ischemia, and endothelial progenitor cells. Eur J
Cardiovasc Prev Rehabil. 18:55–64. 2011.PubMed/NCBI
|
|
11
|
Aicher A, Heeschen C, Sasaki K, Urbich C,
Zeiher AM and Dimmeler S: Low-energy shock wave for enhancing
recruitment of endothelial progenitor cells: a new modality to
increase efficacy of cell therapy in chronic hind limb ischemia.
Circulation. 114:2823–2830. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jiménez-Navarro MF, González FJ,
Caballero-Borrego J, et al: Coronary disease extension determines
mobilization of endothelial progenitor cells and cytokines after a
first myocardial infarction with ST elevation. Rev Esp Cardiol.
64:1123–1129. 2011.(In Spanish).
|
|
13
|
Bonello L, Harhouri K, Baumstarck K, et
al: Mobilization of CD34+ KDR+ endothelial progenitor cells
predicts target lesion revascularization. J Thromb Haemost.
10:1906–1913. 2012.
|
|
14
|
Kim JY, Park YJ, Kim KJ, Choi JJ, Kim WU
and Cho CS: Osteoprotegerin causes apoptosis of endothelial
progenitor cells by induction of oxidative stress. Arthritis Rheum.
65:2172–2182. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen J, Huang L, Song M, Yu S, Gao P and
Jing J: C-reactive protein upregulates receptor for advanced
glycation end products expression and alters antioxidant defenses
in rat endothelial progenitor cells. J Cardiovasc Pharmacol.
53:359–367. 2009. View Article : Google Scholar
|
|
16
|
Skoog T, Dichtl W, Boquist S, et al:
Plasma tumour necrosis factor-alpha and early carotid
atherosclerosis in healthy middle-aged men. Eur Heart J.
23:376–383. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ruan W, Xu JM, Li SB, Yuan LQ and Dai RP:
Effects of down-regulation of microRNA-23a on TNF-α-induced
endothelial cell apoptosis through caspase-dependent pathways.
Cardiovasc Res. 93:623–632. 2012.PubMed/NCBI
|
|
18
|
Markelic M, Velickovic K, Golic I, et al:
Endothelial cell apoptosis in brown adipose tissue of rats induced
by hyperinsulinaemia: the possible role of TNF-α. Eur J Histochem.
55:e342011.PubMed/NCBI
|
|
19
|
Nystrom T, Nygren A and Sjoholm A:
Increased levels of tumour necrosis factor-alpha (TNF-alpha) in
patients with Type II diabetes mellitus after myocardial infarction
are related to endothelial dysfunction. Clin Sci (Lond).
110:673–681. 2006. View Article : Google Scholar
|
|
20
|
Speciale A, Canali R, Chirafisi J, Saija
A, Virgili F and Cimino F: Cyanidin-3-O-glucoside protection
against TNF-α-induced endothelial dysfunction: involvement of
nuclear factor-κB signaling. J Agric Food Chem. 58:12048–12054.
2010.
|
|
21
|
Henrich D, Seebach C, Wilhelm K and Marzi
I: High dosage of simvastatin reduces TNF-alpha-induced apoptosis
of endothelial progenitor cells but fails to prevent apoptosis
induced by IL-1beta in vitro. J Surg Res. 142:13–19. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Seeger FH, Haendeler J, Walter DH, et al:
p38 mitogen-activated protein kinase downregulates endothelial
progenitor cells. Circulation. 111:1184–1191. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ziedén B and Olsson AG: The role of
statins in the prevention of ischemic stroke. Curr Atheroscler Rep.
7:364–368. 2005.
|
|
24
|
Margaritis M, Channon KM and Antoniades C:
Statins as regulators of redox state in the vascular endothelium:
beyond lipid lowering. Antioxid Redox Signal. 20:1198–1215. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhu Y, Casey PJ, Kumar AP and Pervaiz S:
Deciphering the signaling networks underlying simvastatin-induced
apoptosis in human cancer cells: evidence for non-canonical
activation of RhoA and Rac1 GTPases. Cell Death Dis. 4:e5682013.
View Article : Google Scholar
|
|
26
|
Piechota-Polanczyk A, Goraca A, Demyanets
S, et al: Simvastatin decreases free radicals formation in the
human abdominal aortic aneurysm wall via NF-κB. Eur J Vasc Endovasc
Surg. 44:133–137. 2012.
|
|
27
|
Fuhrmeister J, Tews M, Kromer A and
Moosmann B: Prooxidative toxicity and selenoprotein suppression by
cerivastatin in muscle cells. Toxicol Lett. 215:219–227. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hoffmann G, Breitenbücher F, Schuler M and
Ehrenhofer-Murray AE: A novel sirtuin 2 (SIRT2) inhibitor with
p53-dependent pro-apoptotic activity in non-small cell lung cancer.
J Biol Chem. 289:5208–5216. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nihal M, Ahmad N and Wood GS: SIRT1 is
upregulated in cutaneous T-cell lymphoma and its inhibition induces
growth arrest and apoptosis. Cell Cycle. 13:632–640. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hori YS, Kuno A, Hosoda R and Horio Y:
Regulation of FOXOs and p53 by SIRT1 modulators under oxidative
stress. PLoS One. 8:e738752013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kim S, Bi X, Czarny-Ratajczak M, et al:
Telomere maintenance genes SIRT1 and XRCC6 impact age-related
decline in telomere length but only SIRT1 is associated with human
longevity. Biogerontology. 13:119–131. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kok SH, Lin LD, Hou KL, et al: Simvastatin
inhibits cysteine-rich protein 61 expression in rheumatoid
arthritis synovial fibroblasts through the regulation of
sirtuin-1/FoxO3a signaling. Arthritis Rheum. 65:639–649. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ota H, Eto M, Kano MR, et al: Induction of
endothelial nitric oxide synthase, SIRT1, and catalase by statins
inhibits endothelial senescence through the Akt pathway.
Arterioscler Thromb Vasc Biol. 30:2205–2211. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dimitrova KR and Leitman IM:
Intramyocardial transplantation of endothelial progenitor cells and
erythropoietin: a new scope for the treatment of cardiovascular
disease. J Surg Res. 183:550–552. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hu CH, Ke X, Chen K, Yang DY, Du ZM and Wu
GF: Transplantation of human umbilical cord-derived endothelial
progenitor cells promotes re-endothelialization of the injured
carotid artery after balloon injury in New Zealand white rabbits.
Chin Med J (Engl). 126:1480–1485. 2013.
|
|
36
|
Liu P, Zhou B, Gu D, Zhang L and Han Z:
Endothelial progenitor cell therapy in atherosclerosis: a
double-edged sword? Ageing Res Rev. 8:83–93. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Du G, Zhou L, Ghang Q and Li Z: Inhibitory
effect of simvastatin on replicative senescence of endothelial
progenitor cells and its mechanism. J Jilin Univ (Medicine
Edition). 39:913–918. 2013.
|
|
38
|
Thum T, Fraccarollo D, Galuppo P, et al:
Bone marrow molecular alterations after myocardial infarction:
impact on endothelial progenitor cells. Cardiovasc Res. 70:50–60.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tousoulis D, Oikonomou E, Siasos G and
Stefanadis C: Statins in heart failure - with preserved and reduced
ejection fraction. An update Pharmacol Ther. 141:79–91. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Balakumar P, Kathuria S, Taneja G, Kalra S
and Mahadevan N: Is targeting eNOS a key mechanistic insight of
cardiovascular defensive potentials of statins? J Mol Cell Cardiol.
52:83–92. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xu S, Zhao Y, Yu L, Shen X, Ding F and Fu
G: Rosiglitazone attenuates endothelial progenitor cell apoptosis
induced by TNF-α via ERK/MAPK and NF-κB signal pathways. J
Pharmacol Sci. 117:265–274. 2011.PubMed/NCBI
|
|
42
|
Yuan H, Wang Z, Li L, et al: Activation of
stress response gene SIRT1 by BCR-ABL promotes leukemogenesis.
Blood. 119:1904–1914. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Huang PS, Son JH, Abbott LC and
Winzer-Serhan UH: Regulated expression of neuronal SIRT1 and
related genes by aging and neuronal β2-containing nicotinic
cholinergic receptors. Neuroscience. 196:189–202. 2011.PubMed/NCBI
|
|
44
|
Jeong J, Juhn K, Lee H, et al: SIRT1
promotes DNA repair activity and deacetylation of Ku70. Exp Mol
Med. 39:8–13. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Jin Q, Yan T, Ge X, Sun C, Shi X and Zhai
Q: Cytoplasm-localized SIRT1 enhances apoptosis. J Cell Physiol.
213:88–97. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kacimi R, Karliner JS, Koudssi F and Long
CS: Expression and regulation of adhesion molecules in cardiac
cells by cytokines: response to acute hypoxia. Circ Res.
82:576–586. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang W, Yan C, Zhang J, et al: SIRT1
inhibits TNF-α-induced apoptosis of vascular adventitial
fibroblasts partly through the deacetylation of FoxO1. Apoptosis.
18:689–701. 2013.
|
|
48
|
Dvir-Ginzberg M, Gagarina V, Lee EJ, Booth
R, Gabay O and Hall DJ: Tumor necrosis factor α-mediated cleavage
and inactivation of SirT1 in human osteoarthritic chondrocytes.
Arthritis Rheum. 63:2363–2373. 2011.
|
|
49
|
Zhang HN, Li L, Gao P, et al: Involvement
of the p65/RelA subunit of NF-kappaB in TNF-alpha-induced SIRT1
expression in vascular smooth muscle cells. Biochem Biophys Res
Commun. 397:569–575. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Yang J, Huang C, Yang J, Jiang H and Ding
J: Statins attenuate high mobility group box-1 protein induced
vascular endothelial activation: a key role for TLR4/NF-κB
signaling pathway. Mol Cell Biochem. 345:189–195. 2010.PubMed/NCBI
|
|
51
|
Ahn KS, Sethi G and Aggarwal BB: Reversal
of chemoresistance and enhancement of apoptosis by statins through
downregulation of the NF-κB pathway. Biochem Pharmacol. 75:907–913.
2008.PubMed/NCBI
|