Introduction
Autophagy, programmed cell death type 2, serves as a
catabolic process involving the degradation of intracellular
proteins through autophagosomic-lysosmal mechanisms. Autophagy may
be triggered by different stimuli, such as growth factor and
nutrient deprivation, hypoxia, cancer and other pathological
conditions (1–4). Autophagy is usually known to be
important for cell survival and demonstrates pro-tumor properties
in carcinogenesis (5).
Apoptosis, programmed cell death type 1, differs
from autophagy in morphological characteristics and physiological
procedures. In general, apoptosis constitutes the primary type of
cell death, and it plays an anti-survival role in a variety of
intracellular and extracellular conditions. However, tanglesome
interrelationships still exist between autophagy and apoptosis.
Both synergetic and antergic effects are exerted between autophagy
and apoptosis, depending on the cellular environment, oncogenic
stage and stimuli (6,7). Multiple regulatory signaling
pathways are involved in autophagy, such as PI3K, mTOR and p53
(8,9).
Caveolin-1, a 22–24-kDa scaffolding protein, acts as
a critical structural component of caveolae, which constitute the
main invaginations of the plasma membrane (10). Accumulating evidence has indicated
that caveolin-1 functions as a critical regulatory factor in a
diversity of cellular and extracellular processes, including
endocytois, cholesterol homeostasis and signal transduction
(11–13). Furthermore, caveolin-1 plays
important roles in tumorigenesis and demonstrates tumor-promoting
and suppressing effects depending on the tumor cell types and
subtypes (14,15). Studies have demonstrated that
caveolin-1 plays key roles in the retulation of autophagy and
apoptosis (16,17).
17β-estradiol (E2) constitutes one of the 3 major
estrogens (17β-estradiol, estrone and estriol) in women and exerts
genomic and non-genomic effects in a variety of cell types, such as
breast cancer cells, ostebolasts, neurons and pancreatic cells
(18–20). Several studies have suggested that
E2 exerts protective effects by promoting autophagy and inhibiting
the apoptosis of cells (21).
However, E2 displays a protective role by suppressing ferrous
citrate-induced autophagy in neurons (22). Therefore, the role and molecular
mechanisms of action of E2 in autophagy and apoptosis remain
elusive and controversial.
In this study, we demonstrate that in BT474 human
breast cancer cells (ER+), caveolin-1 is highly
expressed and serves as a crucial regulator of E2-mediated
autophagy and apoptosis.
Materials and methods
Cell culture and treatment with E2
The BT474 human breast cancer cell line was
purchased from the American Type Cell Culture (ATCC; Manassas, VA,
USA) and the cells were cultured in RPMI-1640 medium (Gibco-Life
Technologies, Grand Island, NY, USA) supplemented with 10% fetal
bovine serum (FBS; Gibco-Life Technologies) and 1%
penicillin/streptomycin (Beyotime Institute of Biotechnology,
Nanjing, Jiangsu, China) in the presence of 5% CO2 at
37°C.
E2 was purchased from Sigma-Aldrich Corp. (St.
Louis, MO, USA). The cells were plaed in 6-well plates at a density
of 5,000 cells/well. The cells were washed and starved for 24 h in
serum-free RPMI-1640 medium (Gibco-Life Technologies) after up to
60–80% confluence to eliminate the potential influence of estrogen
in fetal bovine serum, and to trigger starvation-induced autophagy.
The cells were then cultured in serum-free RPMI-1640 medium in the
presence of 200 ng/ml E2 or 0.02% ethanol as the vehicle. The cells
were exposed to E2 or the vehicle for 24, 48 and 72 h before they
were harvested.
Antibodies
The antibodies were used were as follows: rabbit
anti-caveolin 1, rabbit anti-high mobility group box 1 protein
(HMGB1), rabbit anti-light chain (LC)3, rabbit anti-Beclin-1,
rabbit anti-Atg12 and rabbit anti-cleaved-caspase-3 were purchased
from Cell Signaling Technology (Beverly, MA, USA). Rabbit
anti-GAPDH was purchased from Santa Cruz Biotechnology (Santa Cruz,
CA, USA).
Small interfering RNA (siRNA) and
transfection
The human caveolin-1, human HMGB1 and negative
control siRNA were designed and constructed by Guangzhou Ribio
Biotech Co., Ltd. (Guangzhou, China). The sequences of caveolin-1
and HMGB1 siRNA were as follows: caveolin-1,
5′-GCAUCAACUUGCAGAAAGAdTdT-3′ and 3′-dTdTCGUAGUUGAACGUCUUUCU-5′;
HMGB1, 5′-GGAGGAAGAUGAAGAAGAUdTdT-3′ and
3′-dTdTCCUCCUUCUACUUCUUCUA-5′. Twenty-four hours prior to
transfection with siRNA, the medium was replaced with
penicillin/streptomycin-free RPMI-1640 complete medium. The BT474
cells were then transfected with siRNA using Lipofectamine™ 2000
(Invitrogen-Life Technologies, Carlsbad, CA, USA) in OPTI-MEM
(Gibco-Life Technologies) for 6 h, and then the medium was replaced
with antibiotics-free RPMI-1640 complete medium. After being
transfected for 48 h, the cells were then exposed to E2 for a
further 24 h.
Western blot analysis
The BT474 cells were planted in 6-well plates until
they were grown to 60–80% confluence and they were treated with
different stimuli for the indicated periods of time. The extraction
of protein was performed using an assay kit (Cell Signaling
Technology), and the concentration of total protein was examined
using the BCA Protein Assay kit (Beyotime Institute of
Biotechnology). For western blot analysis, SDS-PAGE (Beyotime
Institute of Biotechnology) was used to separated equal amounts (30
μg) of total protein and then the protein was transferred onto PVDF
membranes (Millipore, Billerica, MA, USA) using transfer buffer
(200 mM glycin, 40 mM Tris and 20% methanol) at 240 mA for 30–90
min, depending on the molecular weight of the detected proteins.
The membranes were blocked with 5% non-fat milk (Cell Signaling
Technology) in 0.1% TBST (Cell Signaling Technology) for 1 h, at
37°C and then incubated with the primary antibody overnight at 4°C.
After washing in 0.1% TBST 3 times, 5 min each, the membranes were
incubated in horseradish peroxidase (HRP)-conjugated anti-rabbit
secondary antibody (Cell Signaling Technology) at a dilution
1:3,000 for 1 h, 37°C. The membranes were then washed in 0.1% TBST
3 times, 5 min each. The ECL Chemiluminescent Substrate Reagent kit
(Cell Signaling Technology) was added to the membranes and the
quantification of the band intensity of the blots was measured
using Image Lab software (Bio-Rad Laboratories, Hercules, CA, USA).
The internal standard was GAPDH.
Immunofluorescence
The BT474 cells were seeded at sterile glass slides
in 6-well plates. After they reached 50–60% confluence, the cells
were treated with different stimuli as indicated. The medium was
then discarded, and the cells were washed with cold PBS (Cell
Signaling Technology) 3 times. The cells were fixed with 100% cold
methanol at −20°C for 15 min. Following washing 3 times again in
cold PBS, the slides were blocked with 1% bovine serum albumin in
0.1% PBST (Cell Signaling Technology) for 1 h at room temperature.
The slides were incubated with primary antibody overnight at 4°C.
FITC-conjugated goat-anti-rabbit IgG (Cell Signaling Technology)
was used to detect the primary antibody for 1 h at room temperature
in the dark. Subsequently, DAPI (0.3 μmol/l) was used to label the
nuclei for 2 min at room temperature in the dark. The slides were
then washed and images were acquired using an Olympus IX71
epifluorescence microscope (Olympus, Tokyo, Japan).
Monodansycadaverine (MDC) staining
MDC (Sigma-Aldrich), an autofluorescent dye, is used
to detect autophagy. Briefly, the BT474 cells were incubated with
50 μmol/l MDC for 15 min at 37°C to monitor autophagosomes. The
cells were then washed with PBS 3 times immediately followed by
image acquisition. The MDC-positive cells were observed under a
fluorescence microscope (Olympus).
Flow cytometric analysis for
apoptosis
For the apoptosis assay, an Annexin V-FITV/PI
apoptosis kit (Nanjing KeyGen Biotech, Co., Ltd., Nanjing, China)
was used according to the manufacturer’s instructions. Briefly, the
cells (1–5×105) were isolated using EDTA-free Trypsin
(Beyotime Institute of Biotechnology) and collected. The cells were
then centrifuged at 5,000 × g for 5 min, and they were washed 3
times with cold PBS. The cells were resuspended in 500 μl binding
buffer containing 5 μl Annexin V-FITC and 5 μl propidium iodide
(PI). The apoptotic cells were detected by flow cytometry using a
flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA).
CCK-8 cell viability assay
The BT474 cells (5×103 cells/well) were
plated in 96-well plates and cultured for the indicated periods of
time. Cell viability was measured using the CCK-8 kit (Dōjindo
Laboratories, Kumamoto, Japan). Briefly, the medium was removed
followed by the addition of 100 μl CCK-8 in a dilution of 1:9 to
each well. The cells were then incubated at 37°C for 0.5–2 h,
avoiding light. The absorbance of the optical densities was
measured using a microplate reader (Thermo Fisher Scientific,
Waltham, MA, USA) at 450 nm.
Statistical analysis
Results obtained from 3 repeat experiments are
expressed as the means ± standard deviation (SD), and analyzed
using SPSS 11.0 software (SPSS, Inc., Chicago, IL, USA). The
Student’s t-test was used to determine the differences between the
groups. A value of P<0.05 was considered to indicate a
statistically significant difference.
Results
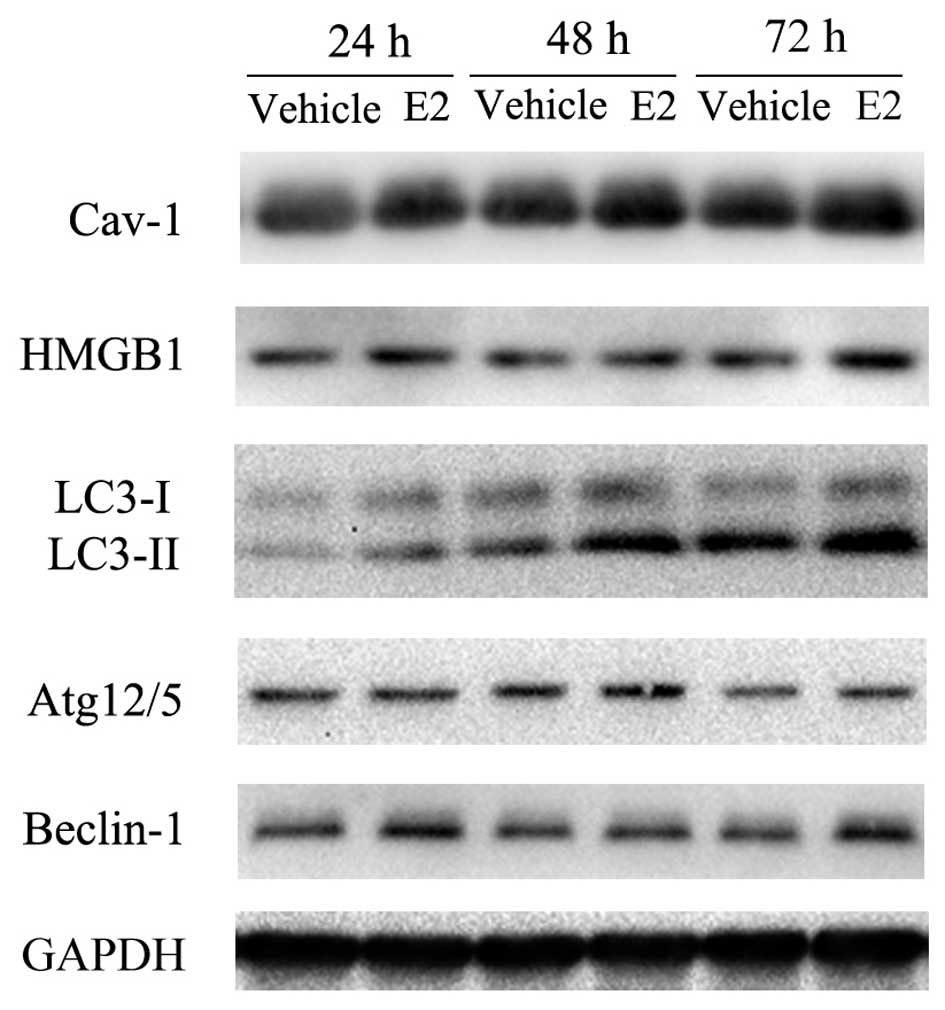
E2 increases the expression of
caveolin-1, HMGB1 and induces autophagy in BT474 cells
To investigate the involvement of E2 in the
autophagy of breast cancer cells, the BT474 (ER+) cells
were pre-incubated with serum-free RPMI-1640 medium for 24 h to
induce autophagy. The cells were then treated with 200 ng/ml E2 or
0.02% ethanol as the vehicle, for various periods of time (24, 48
and 72 h). The expression of caveolin-1, HMGB1 and
autophagy-related proteins (Beclin-1, LC3-II and Atg12) was
detected by western blot analysis. Our results revealed that
compared with the vehicle-treated group, E2 induced a
time-dependent increase in the overall expression of caveolin-1,
HMGB1 and autophagy-related proteins (Beclin-1, LC3-II and Atg12/5)
in the BT474 cells (Fig. 1).
Knockdown of caveolin-1 inhibits
E2-induced autophagy in BT474 cells
To further assess the association between caveolin-1
and HMGB1 and autophagy-related proteins, caveolin-1 siRNA was used
to knock down caveolin-1 expression. At approximately 48 h
following transfection, 200 ng/ml E2 or 0.02% ethanol were added to
the medium for a further 24 h. The results revealed that caveolin-1
knockdown inhibited the E2-induced upregulation of HMGB1, LC3-II
and Atg12/5, but did not affect the E2-induced upregulation of
Beclin-1 (Fig. 2). These results
indicate that E2-induced autophagy is regulated by cavelin-1 by
affecting the expression of LC3-II and Atg12, but not Beclin-1.
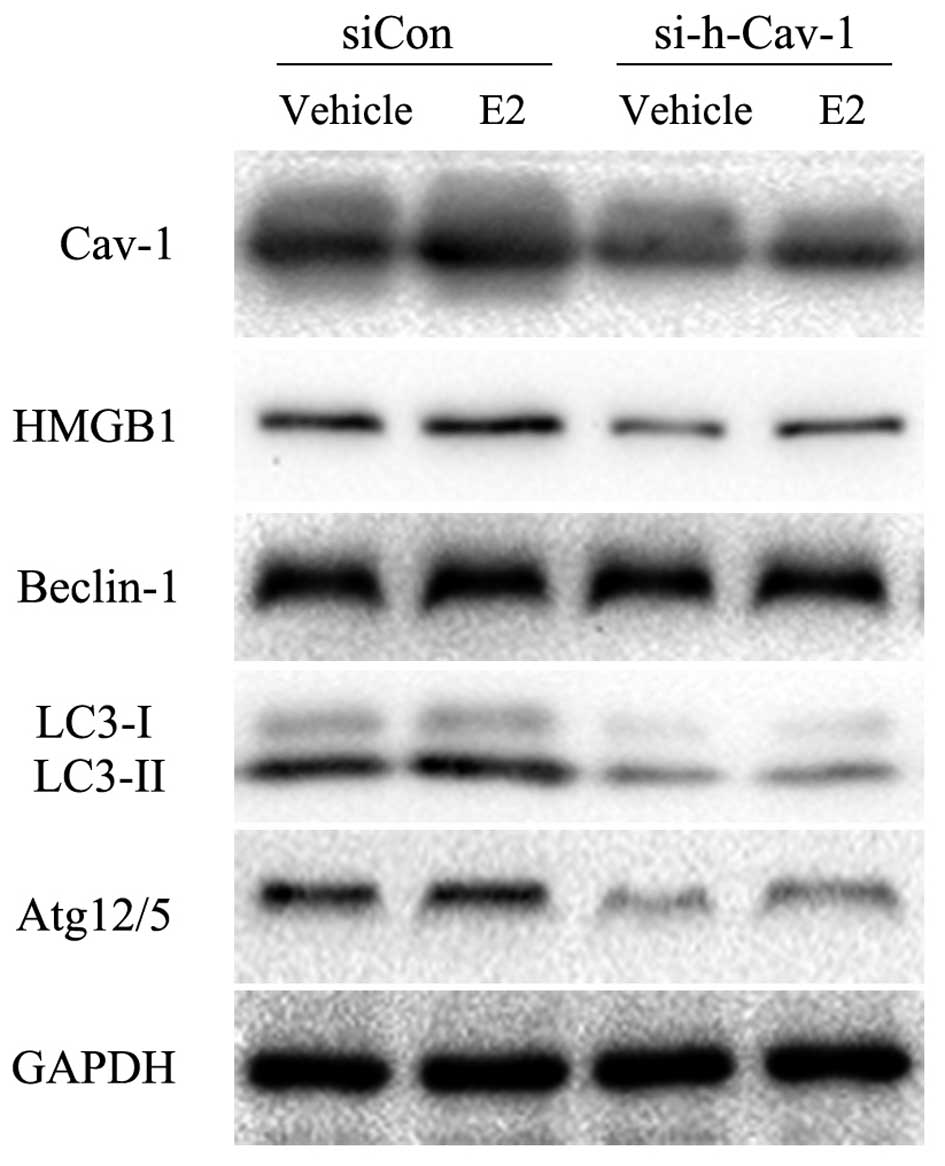 | Figure 2Caveolin-1 knockdown inhibits
E2-induced autophagy in BT474 cells. Western blot analysis was
performed to determine the effects of caveolin-1 knockdown on the
protein expression of HMGB1 and autophagy-related proteins induced
by E2. The results revealed that caveolin-1 knockdown markedly
attenuated E2-induced LC3-II and Atg12/5 protein expression, and
also suppressed HMGB1 protein expression. However, the protein
expression of Beclin-1 was not affected. Experiments were performed
in triplicate, and results were compared with those of the group
transfected with siCon. Cav-1, caveolin-1; si-h-Cav-1, small
interfering RNA against human caveolin-1; siCon, control siRNA; E2,
17β-estradiol; HMGB1, high mobility group box 1; LC3, light chain
3; Atg, autophagy-related gene. |
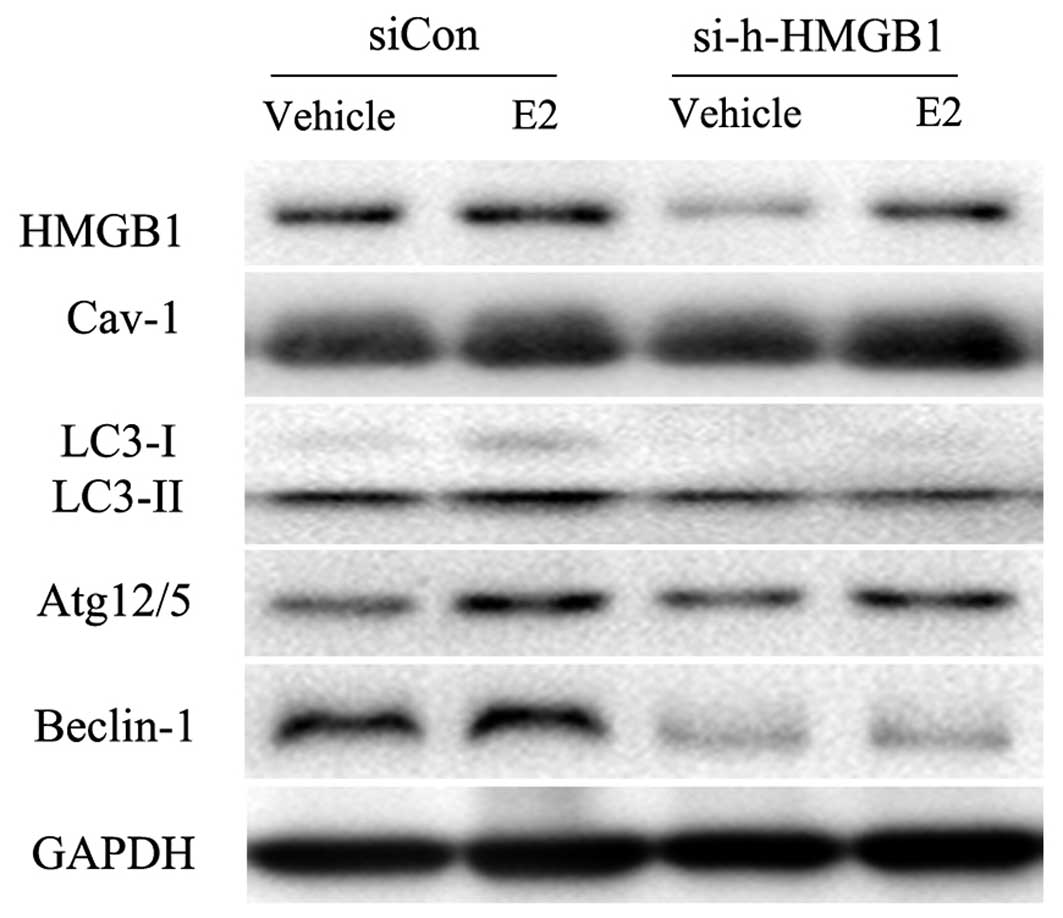
HMGB1 is the link between caveolin-1 and
LC3-II expression in BT474 cells
As caveolin-1 siRNA also decreased HMGB1 expression,
which is deeply related to autophagy, we then determined whether
caveolin-1 regulates autophagy by mediating HMGB1 expression. HMGB1
siRNA was constructed and used to knock down the expression of
HMGB1. We found that the knockdown of HMGB1 resulted in the
inhibition of LC3-II and Beclin-1 expression (Fig. 3). However, caveolin-1 expression
was not impaired by HMGB1 knockdown (Fig. 3), suggesting that HMGB1 may be
involved in the E2/caveolin-1-induced LC3-related autophagy.
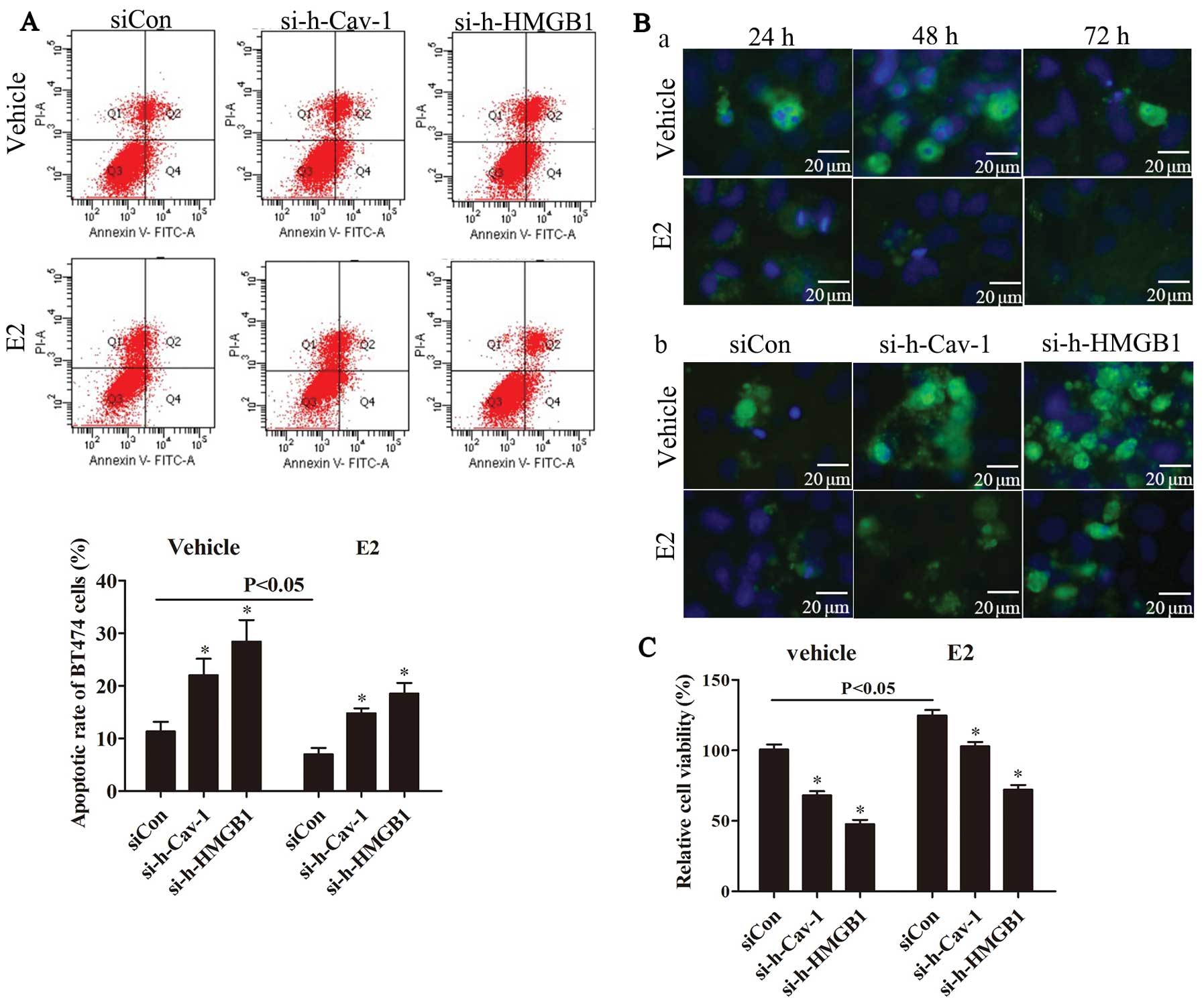
The blockade of caveolin-1 or HMGB1
attenuates E2-induced autophagosome formation
To evaluate autophagy, the LC3 punctate was directly
detected by immunofluoresence staining for LC3, and MDC staining
was further performed to detect auophagosome formation. Our results
revealed that compared with the vehicle-treated group, E2 promoted
the formation of both LC3 punctates (Fig. 4A-a) and autophagosomes (Fig. 4A-b). Compared with the group
transfected with negative control siRNA (siCon), caveolin-1 or
HMGB1 knockdown markedly decreased the E2-induced formation of LC3
punctates (Fig. 4B-a and C-a) and
autophagosomes (Fig. 4B-b and
C-b).
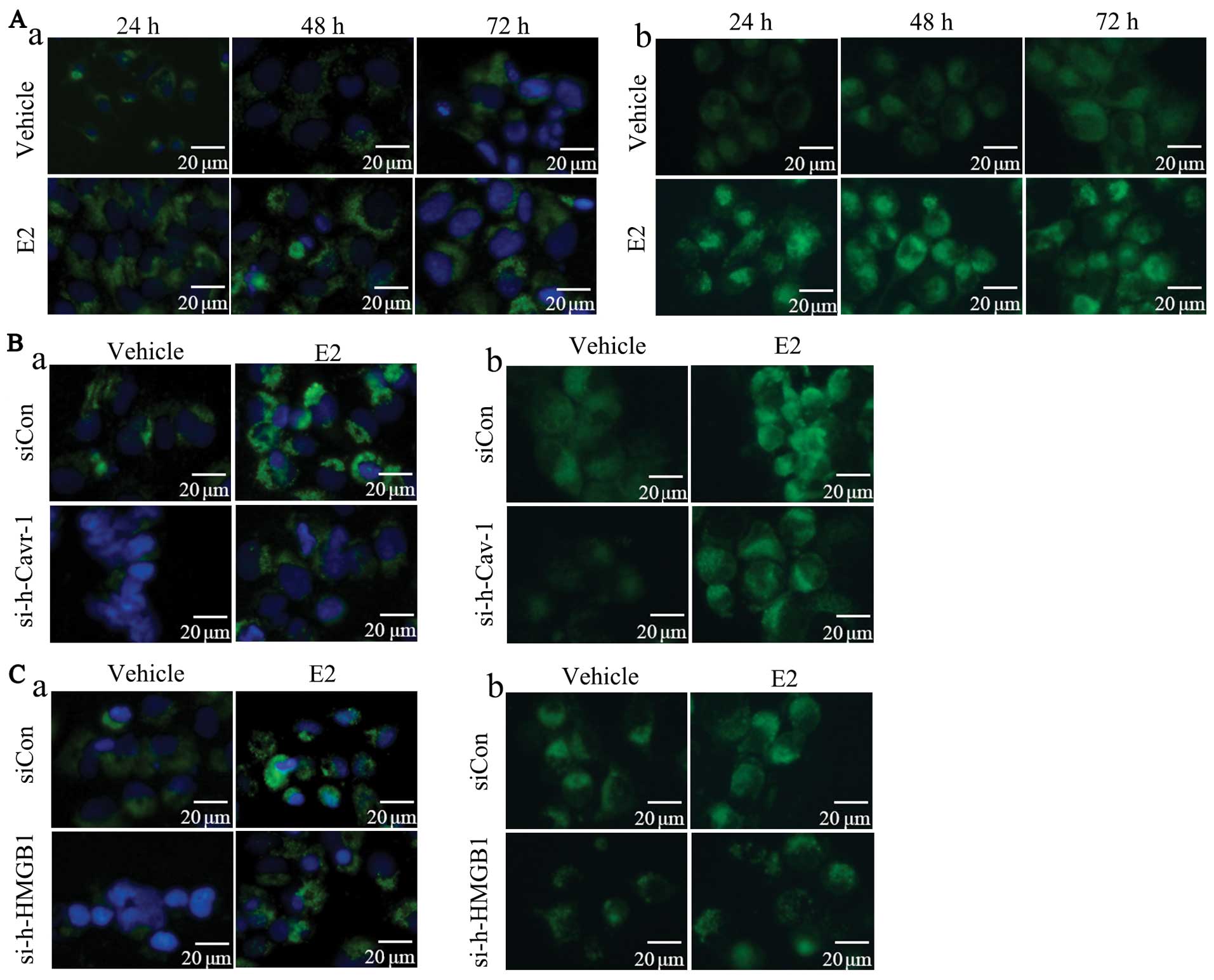 | Figure 4Caveolin-1 or HMGB1 blockade
attenuates the E2-induced formation of LC3 punctates and
autophagesomes. Immnuofluorescence staining for LC3 was used to
detect the LC3 punctates and MDC staining was performed to evaluate
the autophagosome formation in BT474 cells. (A-a) Compared with the
vehicle-treated group, LC3 punctates were induced by E2 in a
time-dependent manner. (A-b) Compare with the vehicle-treated
group, E2 promoted autophagosome formation. (B-a) The results
revealed that caveolin-1 knockdown inhibited E2-induced LC3
punctate formation compared with the siCon group. (B-b) Caveolin-1
knockdown suppressed autophagosome formation compared with siCon
group. (C-a) Compare with the siCon group, HMGB1 knockdwon
decreased E2-induced LC3 punctate formation. (C-b) Compared with
the siCon group, HMGB1 knockdown attenuated E2-induced
autophagosome formation. Experiments were performed in triplicate.
E2, 17β-estradiol; Cav-1, caveolin-1; si-h-Cav-1, small interfering
RNA against human caveolin-1; siCon, control siRNA; HMGB1, high
mobility group box 1; si-h-HMGB1, small interfering RNA against
human HMGB1; LC3, light chain 3; MDC, monodansycadaverine. |
Caveolin-1 or HMGB1 knockdown promotes
apoptosis in BT474 cells
We then explored the regulation of caveolin-1 and
HMGB1 knockdown in the apoptosis and E2-induced cell growth of
BT474 cells. Flow cytometric analysis and immnuofluorescence
staining for cleaved caspase-3 were used to assess the apoptosis of
BT474 cells, and CCK-8 assay was performed in order to evaluate the
viability of the BT474 cells. The results demonstrated that
compared with the vehicle-treted group, E2 inhibited apoptosis and
the formation of cleaved caspase-3 punctates (Fig. 5A and B-a). In addition, compared
with the siCon group, caveolin-1 or HMGB1 knockdown markedly
promoted apoptosis which had been inhibited by E2 (Fig. 5A and B-b). The results from the
evaluation of cell viability by CCK-8 assay revealed that
caveolin-1 or HMGB1 knockdown markedly suppressed the E2-induced
cell growth (Fig. 5C).
Discussion
Autophagy and apoptosis are two key regulatory
mechanisms for cell survival or death. The balance between
autophagy and apoptosis determines the cell status. Previous
studies have revealed that E2 and estradiol analogues affect cell
death and metastasis through the regulation of autophagic and
apoptotic pathways under various conditions, such as starvation,
ischemia and hypoxia (21–23).
In those studies, it was demonstrated that E2 promoted cell
survival by increasing autophagy and inhibiting apoptosis. However,
the exact molecular and functional mechanisms of E2-mediated
autophagy and apoptosis have not yet been elucidated.
LC3, Atg12 and Beclin-1 have been indentified as
autophagy markers. LC3 is an autophagy-related protein and LC3-I is
changed to LC3-II in the process of autophagy. LC3-II aggregates on
the autophagosome memebranes and the ratio of LC3-II/LC3-I
represents a maker for autophagy (24). Autophagosome formation demands an
ubiquitin-like conjugation system. In this system, Atg12 is bound
to Atg5 to target the autophagosome vesicles (25). Beclin-1 is the mammalian ortholog
of yeast Apg6/Vps30 and plays a crucial role in the formation of
autophagosomes (26).
The role for caveolin-1 in autophagy and apoptosis
has been supported by several studies. Recent research has
demonstrated that caveolin-1 binds LC3B through its scaffolding
domain (27). It has been
reported that caveolin-1 depletion increases the expression of
autophagy markers (LC3-II, Atg5/12 and p-ULK) (28). In addition, the expression of
caveolin-1 abrogates apoptosis in hepatocytes, small cell lung
cancer cells and thymocytes (29–31). However, certain studies have
reported that caveolin-1 sensitizes cells to drug-induced apoptosis
(32,33). These data suggest that the effects
of caveolin-1 on autophagy and apoptosis are dependent on specific
cell types and stimuli.
HMGB1 protein, a chromatin-binding factor, presents
as a highly conserved nuclear protein and bends DNA. HMGB1 is also
involved in various cellular processes, such as cell
differentiation, wound healing, cell survival and death, as well as
cell apoptosis and autphagy (34–36). Although a number of studies have
revealed that HMGB1 is closely related to autophagy and apoptosis,
the role of HMGB1 in E2-mediated autophagy remains unclear.
In the present study, we provide a possible
explanation for E2-mediated autophagy and apoptotic processes in
breast cancer cells. We demonstrate that E2 promotes cell growth by
mediating autophagy and apoptosis in BT474 breast cancer cells,
which are regulated by the caveolin-1/HMGB1 pathway. We found that
E2 promoted the expression of both caveolin-1 and HMGB1, as well as
that of autophagy-related proteins (LC3-II, Beclin-1 and Atg12/5).
E2 also inhibited apoptosis and the formation of cleaved-caspase-3
punctates. Caveolin-1 knockdown downregulated HMGB1 and
autophagy-related proteins (LC3-II and Atg12/5) and suppressed
autophagosome formation, and promoted apoptosis. Furthermore, the
blockade of HMGB1 inhibited the expression of autophagy-related
proteins (LC3-II and Beclin-1) and significantly promoted apoptosis
in the BT474 cells. Based on this evidence, it can be concluded
that the caveolin-1/HMGB1 pathway contributes to the E2-mediated
pro-autophagy and anti-apoptotic processes.
Taken together, the data from the present study
provide further insight into the relationship and interaction
between E2 and autophagy and apoptosis. However, the significance
of E2 and caveolin-1 in mediating autophagy and the apoptosis of
breast cancer cells requires further investigation.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81001171) and the
Key Technologies R&D Program of Hubei Province (grant no.
2007AA302B07).
References
|
1
|
Stanton MJ, Dutta S, Polavaram NS, Roy S,
Muders MH and Datta K: Angiogenic growth factor axis in autophagy
regulation. Autophagy. 9:789–790. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kuma A, Hatano M, Matsui M, et al: The
role of autophagy during the early neonatal starvation period.
Nature. 432:1032–1036. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pouyssegur J, Dayan F and Mazure NM:
Hypoxia signalling in cancer and approaches to enforce tumour
regression. Nature. 441:437–443. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Guo JY, Xia B and White E:
Autophagy-mediated tumor promotion. Cell. 155:1216–1219. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wei H and Guan JL: Pro-tumorigenic
function of autophagy in mammary oncogenesis. Autophagy. 8:129–131.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ye L, Zhao X, Lu J, Qian G, Zheng JC and
Ge S: Knockdown of TIGAR by RNA interference induces apoptosis and
autophagy in HepG2 hepatocellular carcinoma cells. Biochem Biophys
Res Commun. 437:300–306. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nepal S and Park PH: Activation of
autophagy by globular adiponectin attenuates ethanol-induced
apoptosis in HepG2 cells: involvement of AMPK/FoxO3A axis. Biochim
Biophys Acta. 1833:2111–2125. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang DM, Liu JS, Deng LJ, et al: a
natural bufadienolide from toad venom, induces apoptosis and
autophagy in human hepatocellular carcinoma cells through
inhibition of PI3K/Akt/mTOR pathway. Carcinogenesis. 34:1331–1342.
2013. View Article : Google Scholar
|
|
9
|
Wang EY, Gang H, Aviv Y, Dhingra R,
Margulets V and Kirshenbaum LA: p53 mediates autophagy and cell
death by a mechanism contingent on Bnip3. Hypertension. 62:70–77.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Okamoto T, Schlegel A, Scherer PE and
Lisanti MP: Caveolins, a family of scaffolding proteins for
organizing ‘preassembled signaling complexes’ at the plasma
membrane. J Biol Chem. 273:5419–5422. 1998.
|
|
11
|
Gonzalez MI, Krizman-Genda E and Robinson
MB: Caveolin-1 regulates the delivery and endocytosis of the
glutamate transporter, excitatory amino acid carrier 1. J Biol
Chem. 282:29855–29865. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bosch M, Mari M, Herms A, et al:
Caveolin-1 deficiency causes cholesterol-dependent mitochondrial
dysfunction and apoptotic susceptibility. Curr Biol. 21:681–686.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Svensson KJ, Christianson HC, Wittrup A,
et al: Exosome uptake depends on ERK1/2-heat shock protein 27
signaling and lipid Raft-mediated endocytosis negatively regulated
by caveolin-1. J Biol Chem. 288:17713–17724. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kannan A, Krishnan A, Ali M, Subramaniam
S, Halagowder D and Sivasithamparam ND: Caveolin-1 promotes gastric
cancer progression by up-regulating epithelial to mesenchymal
transition by crosstalk of signalling mechanisms under hypoxic
condition. Eur J Cancer. 50:204–215. 2014. View Article : Google Scholar
|
|
15
|
Ayala G, Morello M, Frolov A, et al: Loss
of caveolin-1 in prostate cancer stroma correlates with reduced
relapse-free survival and is functionally relevant to tumour
progression. J Pathol. 231:77–87. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ha TK and Ch SG: CAV1/caveolin 1 enhances
aerobic glycolysis in colon cancer cells via activation of
SLC2A3/GLUT3 transcription. Autophagy. 8:1684–1685. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guo YH, Hernandez I, Isermann B, et al:
Caveolin-1-dependent apoptosis induced by fibrin degradation
products. Blood. 113:4431–4439. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jiang QF, Wu TT, Yang JY, et al:
17β-estradiol promotes the invasion and migration of nuclear
estrogen receptor-negative breast cancer cells through cross-talk
between GPER1 and CXCR1. J Steroid Biochem Mol Biol. 138:314–324.
2013.
|
|
19
|
Fester L, Labitzke J, Hinz R, et al:
Estradiol responsiveness of synaptopodin in hippocampal neurons is
mediated by estrogen receptor β. J Steroid Biochem Mol Biol.
138:455–461. 2013.PubMed/NCBI
|
|
20
|
Rapid regulation of K(ATP) channel
activity by 17{beta}-estradiol in pancreatic {beta}-cells involves
the estrogen receptor {beta} and the atrial natriuretic peptide
receptor. Mol Endocrinol. 23:1973–1982. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang YH, Chen K, Li B, et al: Estradiol
inhibits osteoblast apoptosis via promotion of autophagy through
the ER-ERK-mTOR pathway. Apoptosis. 18:1363–1375. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen CW, Chen TY, Tsai KL, et al:
Inhibition of autophagy as a therapeutic strategy of iron-induced
brain injury after hemorrhage. Autophagy. 8:1510–1520. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Visagie MH and Joubert AM: The in vitro
effects of 2-methoxyestradiol-bis-sulphamate on cell numbers,
membrane integrity and cell morphology, and the possible induction
of apoptosis and autophagy in a non-tumorigenic breast epithelial
cell line. Cell Mol Biol Lett. 15:564–581. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kabeya Y, Mizushima N, Ueno T, et al: LC3,
a mammalian homologue of yeast Apg8p, is localized in autophagosome
membranes after processing. EMBO J. 19:5720–5728. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mizushima N, Sugita H, Yoshimori T and
Ohsumi Y: A new protein conjugation system in human. The
counterpart of the yeast Apg12p conjugation system essential for
autophagy. J Biol Chem. 273:33889–33892. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kametaka S, Okano T, Ohsumi M and Ohsumi
Y: Apg14p and Apg6/Vps30p form a protein complex essential for
autophagy in the yeast, Saccharomyces cerevisiae. J Biol
Chem. 273:22284–22291. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ryter SW, Lam HC, Chen ZH and Choi AM:
Deadly triplex: smoke, autophagy and apoptosis. Autophagy.
7:436–437. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ha TK, Her NG, Lee MG, et al: Caveolin-1
increases aerobic glycolysis in colorectal cancers by stimulating
HMGA1-mediated GLUT3 transcription. Cancer Res. 72:4097–4109. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Meyer C, Liu Y, Kaul A, Peipe I and Dooley
S: Caveolin-1 abrogates TGF-β mediated hepatocyte apoptosis. Cell
Death Dis. 4:e4662013.PubMed/NCBI
|
|
30
|
Yang X, Xiong H, Guan ZZ, et al: Higher
expression of Caveolin-1 inhibits human small cell lung cancer
(SCLC) apoptosis in vitro. Cancer Invest. 30:453–462. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Feng H, Guo L, Song Z, Gao H, et al:
Caveolin-1 protects against sepsis by modulating inflammatory
response, alleviating bacterial burden, and suppressing thymocyte
apoptosis. J Biol Chem. 285:25154–25160. 2010. View Article : Google Scholar
|
|
32
|
Ingueneau C, Huynh UD, Marcheix B, et al:
TRPC1 is regulated by caveolin-1 and is involved in oxidized
LDL-induced apoptosis of vascular smooth muscle cells. J Cell Mol
Med. 13:1620–1631. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pongjit K and Chanvorachote P: Caveolin-1
sensitizes cisplatin-induced lung cancer cell apoptosis via
superoxide anion-dependent mechanism. Mol Cell Biochem.
358:365–373. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huang J, Liu K, Yu Y, et al: Targeting
HMGB1-mediated autophagy as a novel therapeutic strategy for
osteosarcoma. Autophagy. 8:275–277. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kang R, Livesey KM, Zeh HJ III, Lotze MT
and Tang D: HMGB1 as an autophagy sensor in oxidative stress.
Autophagy. 7:904–906. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tang D, Kang R, Livesey KM, et al:
Endogenous HMGB1 regulates autophagy. J Cell Biol. 190:881–892.
2010. View Article : Google Scholar
|