Introduction
Osteoclasts have unique bone resorption activity.
The dynamic equilibrium between osteoclast-mediated bone resorption
and osteogenesis plays an important role in the bone remodeling
process. Excessive bone resorption can cause a variety of diseases,
such as osteoporosis, rheumatoid arthritis, multiple myeloma and
periodontitis (1,2). Therefore, investigation of bone
absorption by osteoclasts is crucial for the development of
effective therapeutic strategies.
The sealing zone is a cytoskeletal structure unique
to osteoclasts that consists of a tight arrangement of the actin
belt formed by podosomes (3,4).
The sealing zone provides a closed microenvironment for
osteoclast-mediated bone resorption, thus, defining the area of
bone resorption. Osteoclasts secrete protons and proteinases, which
enter the microenvironment formed by the sealing zone to
participate in the dissolution and absorption of the bone matrix
and minerals (5–7). Furthermore, the sealing zone allows
the tight adherence of osteoclasts to the surface of bone (8,9).
The podosome, which is the structural unit of the
sealing zone (6), is formed by
the F-actin-binding core and the radiating actin fibers. During the
osteoclast maturation process, the arrangement of podosomes changes
from a spot-like to a cluster-like pattern that eventually
transforms into a podosome belt or the sealing zone (10,11). There is no essential difference
between the podosome belt and the sealing zone, apart from the
difference in density (4,6). Myeloid cells, such as mononuclear
macrophages and osteoclasts produce podosomes instead of the stress
fibers and focal adhesions that are produced in other cells.
Podosomes, focal adhesions and stress fibers share many common
components (5,12). The podosome has actin at its
center, surrounded by scaffold proteins, kinases and integrins
(6). Previous studies have
indicated that the integrity of the sealing zone plays a decisive
role in osteoclast-mediated bone resorption (4,13).
RhoGTPases, which include at least 22 family
members, affect the sealing zone by regulating the generation and
arrangement of podosomes (4,5,14–16). These enzymes are inactivated in
the GDP-bound form and are activated in the GTP-bound form, playing
an important role in cell signal transduction and the regulation of
F-actin in the cytoskeleton (15,16). Rho guanine nucleotide exchange
factors (RhoGEFs) induced the upstream activation of RhoGTPases
(17). Studies have shown that
the expression of only Arhgef8/Net1 and DOCK5 among the RhoGEFs is
altered during the formation and maturation of osteoclasts
(18,19); thus, it can be speculated that
they play an important role in the regulation of the sealing zone.
ROCK is the downstream effector of the Rho subfamily, the most
extensively investigated member of this family, which includes
ROCK1 and ROCK2 (20,21).
Osteoprotegerin (OPG) markedly inhibits the
transformation of osteoclast precursor cells into osteoclasts.
Studies on the effects of OPG on bone resorption by osteoclasts
have focused mainly on the OPG-induced inhibition of osteoclast
differentiation through the OPG/receptor activator of the nuclear
factor kappa-B ligand (RANKL)/RANK system (22–24). It has been reported that OPG
promotes the separation of osteoclasts adhering to the bone matrix
from the bone surface (25).
However, the mechanisms involved have not yet been elucidated. In
addition, to the best of our knowledge, the effects of OPG on the
sealing zone and the underlying molecular mechanisms have not been
reported to date.
In this study, the effects of OPG on the sealing
zone of purified osteoclasts were observed. Furthermore, the
expression of the RhoGTPase family members, Net1, DOCK5, ROCK
(ROCK1 and ROCK2) was analyzed, in order to investigate the
mechanisms responsible for OPG action on the sealing zone.
Materials and methods
Induction and purification of
osteoclasts
Murine macrophage-like RAW264.7 cells were purchased
from the Type Culture Collection of the Chinese Academy of
Sciences, Shanghai, China. Studies have indicated that massive
apoptosis of monocytes occurs during the generation of osteoclasts
(26). According to this rule,
the yield of osteoclasts can be increased by reducing the density
of Raw264.7 cells (a mouse leukemic monocyte/macrophage cell line)
at the early stage of inoculation and increasing the duration of
induction. This will cause the greatest degree of apoptosis of
undifferentiated monocytes. After the formation of a large number
of osteoclasts, the monocytes can be removed by aspiration. This
method was used for the generation of all osteoclasts used in this
study.
For investigations of the formation of the sealing
zone, RAW264.7 cells (1562.5 cells/cm2) were incubated
in α-MEM culture medium containing 50 ng/ml M-CSF and 60 ng/ml
RANKL (PeproTech, Rocky Hill, NJ, USA), 10% fetal bovine serum
(FBS) (HyClone, Logan, UT, USA) and 100 U/ml
penicillin/streptomycin (Sigma-Aldrich, St. Louis, MO, USA) for 1–5
days. Immunofluorescence (IF) staining (rhodamine phalloidin) were
performed daily. IF staining was conducted to monitor the formation
of the sealing zone. The time required for the formation of the
sealing zone and the characteristics of the cells during the
osteoclast differentiation process were observed using a Hoffman
microscope (Leica, Wetzlar, Germany) on days 1–5.
For investigations of the effects of OPG (PeproTech)
on the sealing zone, RAW264.7 cells (1,562.5 cells/cm2)
were incubated in α-MEM culture medium containing 50 ng/ml M-CSF
and 60 ng/ml RANKL (PeproTech), 10% FBS (HyClone) and 100 U/ml
penicillin/streptomycin (Sigma-Aldrich). Based on the results of
this staining protocol, osteoclasts with a visible sealing zone
were incubated for 12 h with OPG (0, 20, 40 and 80 ng/ml). The
changes of living osteoclasts in the sealing zone in these
experiments were captured by Hoffman modulation contrast (HMC)
microscopy. Moreover, IF staining (rhodamine phalloidin) was
performed to monitor the effects of OPG (PeproTech) on the sealing
zone.
IF staining
The cells obtained as described above were fixed for
15 min with 4% paraformaldehyde (Sigma-Aldrich), and washed 3 times
with phosphate-buffered saline (PBS). Subsequently, the cells were
washed with 0.2% Triton X-100 (Amresco, Solon, OH, USA) for 10 min,
then washed 3 times with PBS. The cells were sealed for 1 h with
500 μl 2% bovine serum albumin (BSA; Sigma-Aldrich). Subsequently,
the cells were stained for 1 h with rhodamine phalloidin
(Invitrogen, Carlsbad, CA, USA) diluted in 2% BSA and washed 3
times with PBS. During the analysis of the sealing zone formation,
the cells were photographed immediately using an inverted
fluorescence microscope (LSM510; Carl Zeiss, Jena, Germany).
However, during the analysis of the effects of OPG on the sealing
zone, the cells were stained for 15 min with
4′,6-diamidino-2-phenylindole (DAPI), and washed 3 times with PBS.
Finally, images were acquired using an inverted fluorescence
microscope (Leica).
Quantitative reverse transcription
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the cells treated with
OPG using TRIzol reagent (Invitrogen) according to manufacturer’s
instructions. Reverse transcription was performed using the
PrimeScript RT reagent kit with gDNA eraser (Takara Bio, Inc.,
Shiga, Japan). Subsequently, the mRNA expression levels of
Arhgef8/Net1, DOCK5, the RhoGTPase family members, DOCK2 and other
genes (MMP-9 and CAII) associated with bone resorption were
detected with the 7500 Real-Time PCR System using SYBR-Green
(Applied Biosystems, Carlsbad, CA, USA) following the
manufacturer’s instructions. The primers used in this study are
presented in Table I.
 | Table IPrimers used in real-time PCR. |
Table I
Primers used in real-time PCR.
| Gene name | Accession no. | Upstream | Downstream |
|---|
| RhoA | NM_016802 |
CAAGGACCAGTTCCCAGAGG |
CGCAGGCGGTCATAATCTTC |
| RhoB | NM_007483 |
AAGTGGGTGCCCGAGGTA |
CGAGGTAGTCATAGGCTTGGAT |
| RhoC | NM_007484 |
GGAAGACCTGCCTCCTCATT |
TCCACCTGCTTGCCATCCAC |
| Rac1 | NM_009007 |
GCCTGCTCATCAGTTACACG |
GACGCAATCTGTCATAATCTTC |
| Rac2 | NM_009008 |
TGTGATGGTGGACAGTAAGCC |
GAACCACTTGGCACGGACAT |
| Rac3 | NM_133223 |
TGGTGAGCCCAGCCTCCTTT |
CCGCAGCCGTTCAATCGTAT |
| RhoG | NM_019566 |
CTGCTATACAACTAACGCCTTC |
GTCCCACAGGTTTAGGTTCACG |
| Cdc42v1 | NM_009861 |
CCTTTCTTGCTTGTTGGGAC |
TGCGGCTCTTCTTCGGTT |
| Cdc42v2 | NM_001243769.1 |
GTGGTCTCTCCATCCTCATT |
GCCTCATCAAACACATTCTTC |
| RhoU (Wrch1) | NM_133955 |
GAGAAGCCGGTGCCTGAAGA |
GCTGGGAGTCTGAGTGCTGGAT |
| Rnd1 | NM_172612 |
AGTGGTGGTCAGGTGTAAGC |
GCACATAGGTCTCGGGATAG |
| Rnd2 | NM_009708 |
GGTGCTGGTTGGCTGTAAG |
CAGAAGATCGGGAGGAACA |
| RhoF (Rif) | NM_175092 |
CCCGTCGGTGTTTGAGAAGTA |
GAGGACGTTGTCGTAACTGGTG |
| RhoBTB1 | NM_001252638 |
AAGGAGGAAGACCACTACCAG |
GGACGAATGCCAGAAACAC |
| ROCK2 | NM_009072.2 |
TTCACGTCCGACCTGTTAC |
GGCACCTACGGCACTCTA |
| DOCK5 | XM_619261 |
TGGACCTGCTAGGCTTACT |
GATGTCTCCTATCAGCGAAA |
| Arhgef8/Net1 | NM_019671 |
TGCCAGGCTTAAACGCTTGC |
TGAATGCAGAAGGCGAACCG |
| CAII | K00811.1 |
CATTACTGTCAGCAGCGAGCA |
GACGCCAGTTGTCCACCATC |
| MMP-9 | NM_013599.3 |
GCCCTGGAACTCACACGACA |
TTGGAAACTCACACGCCAGAAG |
| GAPDH | GU214026 |
ATGGTGAAGGTCGGTGTG |
TGAAGGGGTCGTTGATGG |
Western blot analysis
Total proteins were extracted from the OPG-treated
cells. The concentration of protein was determined using the BCA
protein assay kit (Beyotime, Shanghai, China). Equal amounts of
proteins were separated by sodium dodecyl sulfate-polyacrylamide
gel electrophoresis [SDS-PAGE; 12% gel for glyceraldehyde
3-phosphate dehydrogenase (GAPDH) and 6% gel for ROCK1]. The
proteins were then transferred onto nitrocellulose membranes and
non-specific binding was blocked by incubation with 5% non-fat milk
for 2 h at room temperature. Subsequently, the cells were incubated
overnight at 4°C with primary anti-rabbit monoclonal antibodies
(GAPDH and ROCK1) (Cell Signaling Technology, Beverly, MA, USA)
diluted with 5% BSA in TBST. The membranes were then washed 6 times
in TBST piror to incubation with HRP-coupled goat anti-rabbit
monoclonal antibody (Cell Signaling Technology) diluted with 5% BSA
in TBST. The membranes were then washed 6 times with TBST. The
immune complexes were detected using the ECL system by standard
scanning densitometry with the normalization of desitometry
measures to GAPDH. The gray values of the blots were detected using
Image Lab software.
Observation of bone resorption lacunae by
scanning electron microscopy
Osteoclast purification and induction from the
RAW264.7 cells was performed as described above, with the
modification of cells being plated in 24-well plates containing
bovine cortical bone slices. Following the formation of the sealing
zone in osteoclasts, the cells were treated with 0 ng/ml and 20
ng/ml OPG for 12 h. Finally, the bone resorption lacunae of
osteoclasts were observed by scanning electron microscopy.
Statistical analysis
In this study, each reaction was carried out in
triplicate. The data of mRNA expression were analyzed by comparing
their 2−ΔΔCt values. The results are presented
statistically as the means ± standard deviation (SD). Significance
was assessed by one-way ANOVA. The results were compared between
groups using ANOVA and the least significant difference (LSD)
post-hoc test following the appropriate transformation to
normalized data and equalized variance where necessary. Statistical
analysis was performed using SAS 9.1.3 software (SAS Institute
Inc., Cary, NC, USA); Values of P<0.05 and P<0.01 were
considered to indicate statistically significant and highly
significant differences, respectively.
Results
Differentiation and purification of
osteoclasts
In order to reduce the effects of monocytes on the
experimental results, the osteoclasts were purified at the stage
where large quantities of osteoclasts were produced. When the
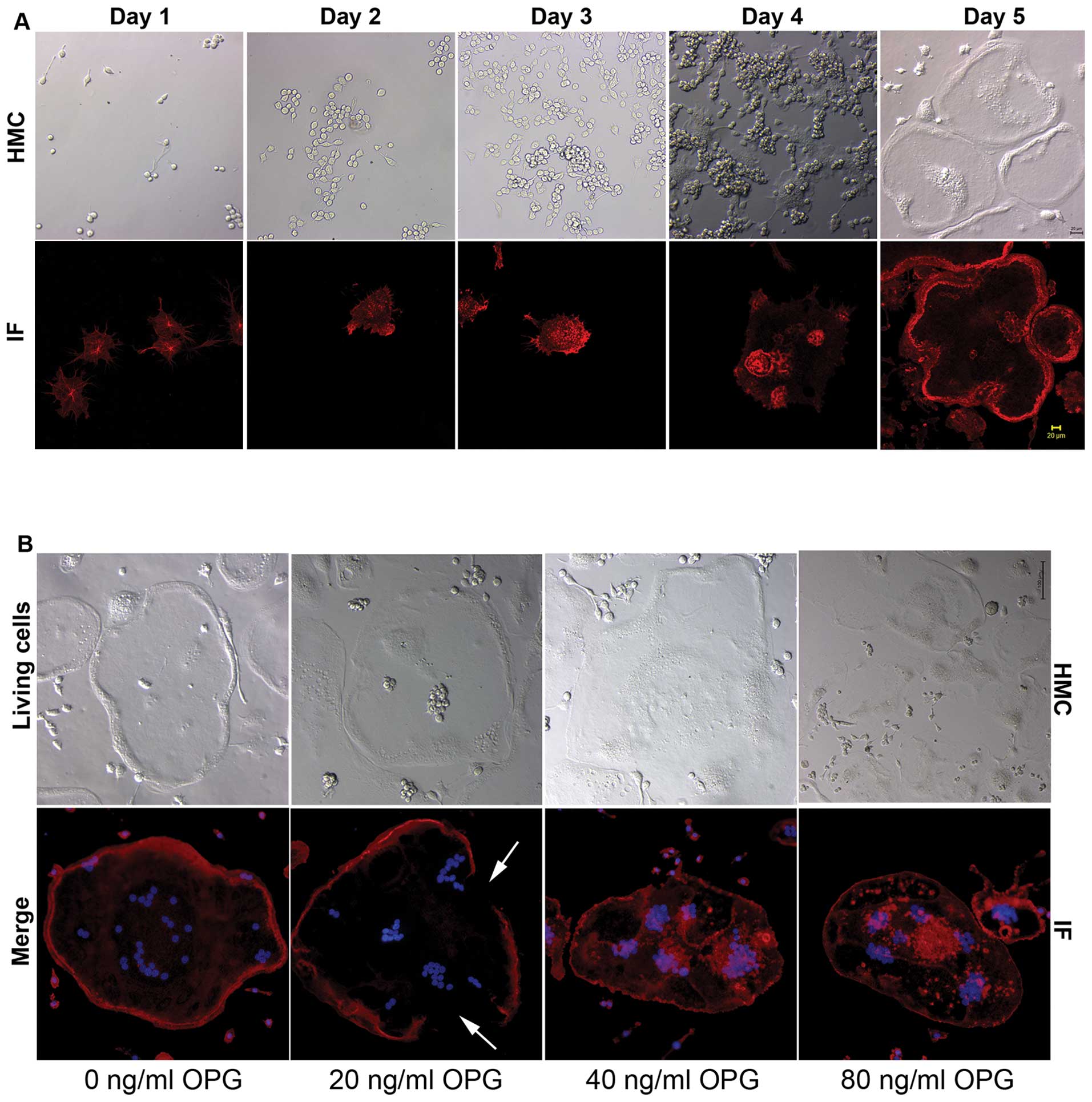
sealing zone was formed, very few monocytes were found in the
culture plate (Fig. 1A; HMC, day
5). Therefore, the presence of monocytes had little impact on the
research results.
The characteristics of the osteoclasts were observed
at the stage where the sealing zone was formed, which provided a
reference for the addition of OPG. During the process of osteoclast
differentiation, phalloidin-based IF staining was performed
together with photographing live osteoclasts using an HMC
microscope.
The sealing zone is composed of F-actin, to which
phalloidin specifically binds. IF staining revealed that the
formation of the sealing zone was mostly complete on day 5. HMC
microscopy revealed that the cell morphology at this stage was
characterized by a smooth cell edge, belt-like protuberances, no
filopodia and a scattered arrangement of nuclei (Fig. 1A; HMC, day 5). In the subsequent
experiments, OPG was added at the time at which the sealing zone
had just appeared.
Effects of OPG on the sealing zone of
osteoclasts
The evaluation of IF staining using an inverted
fluorescence microscope revealed that, in the control group without
the addition of OPG, the podosomes of the osteoclasts showed a
strip-like, compact arrangement. On the inner side of the edge of
cell, there were no podosomes; the sealing zone was intact
(Fig. 1B, IF), and the strip-like
protuberances on the edge of the living cell were intact (Fig. 1B, HMC). By contrast, following
treatment with 20 ng/ml OPG, the sealing zone of the osteoclasts
showed defects (Fig. 1B, IF).
There were defects in the strip-like protuberances on the edge of
the cell (Fig. 1B, HMC). It was
surprising to note that the sealing zone of the osteoclasts in the
40 ng/ml and 80 ng/ml OPG-treated groups had completely
disappeared. Between the basal surface of the osteoclasts and the
well surface, the podosomes showed a scattered or clustered
distribution (Fig. 1B).
Expression of Arhgef8/Net1, DOCK5,
RhoGTPase family members, DOCK and other genes associated with bone
resorption
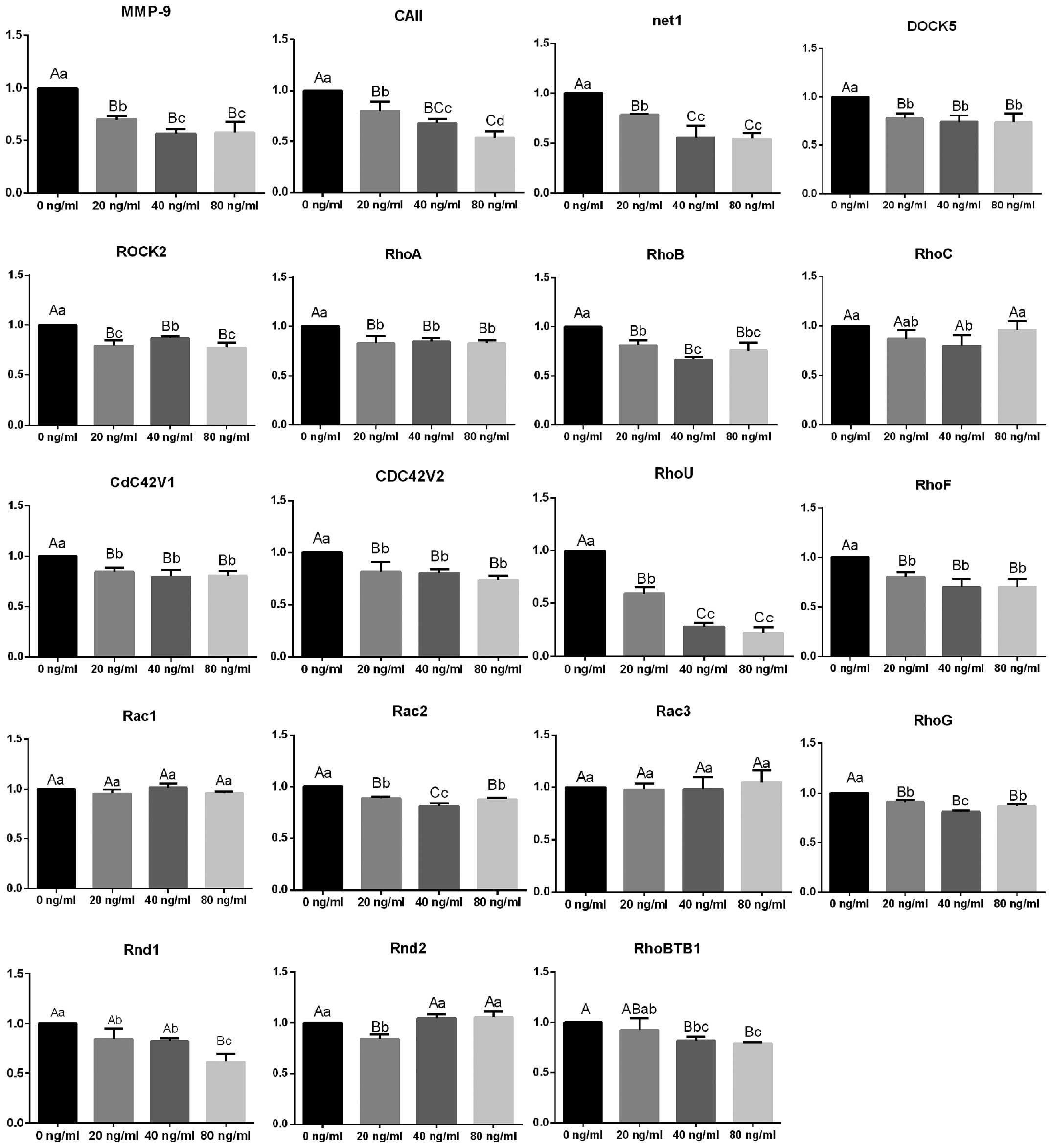
Although no significant differences in the
expression of RhoC, Rac1, Rac3 and Rnd2 were observed following
treatment with OPG at different concentrations, the mRNA expression
levels of Arhgef8/Net1, DOCK5, other RhoGTPases and ROCK2 were all
significantly downregulated (Fig.
2). Following treatment with OPG at 40 and 80 ng/ml, the mRNA
expression levels of MMP-9 and CAII were reduced to approximately
50% of the levels detected in the control group, while RhoU mRNA
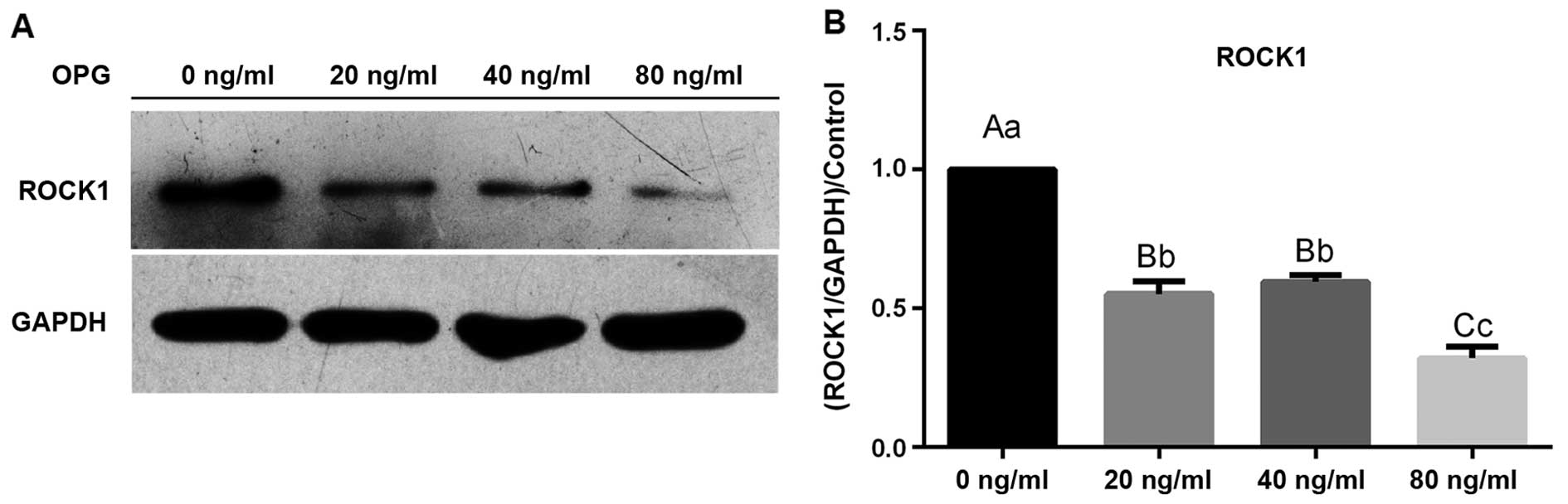
expression was reduced by approximately 80%. Western blot analysis
revealed that ROCK1 protein expression was sharply reduced
following treatment with OPG in a concentration-dependent manner
(Fig. 3).
Effects of OPG on resorption lacunae in
osteoclasts
Following treatment with OPG, no visible resorption
lacunae were observed in the bovine cortical bone slices (Fig. 4). However, in the control group
not treated with OPG, many bead-like resorption lacunae were
observed in the osteoclasts. These observations indicated the loss
of osteoclast bone resorption function following treatment with
OPG.
Discussion
In this study, the effects of OPG on the existing
sealing zones of osteoclasts were investigated. Variations in the
expression of genes (Arhgef8/Net1, DOCK5, RhoGTPase family members
and ROCK) that play key regulatory roles in the formation of the
sealing zone were determined before and after treatment with OPG.
It was found that OPG treatment caused damage or complete
destruction of the existing sealing zones of the osteoclasts. The
osteoclasts lost the bone resorption function during this time. The
expression of Arhgef8/Net1 and DOCK5 belonging to the RhoGEF
family, 10 of 18 RhoGTPase family members, as well as 2 members of
the ROCK family (ROCK1 and ROCK2) was downregulated significantly
following treatment with OPG.
During the osteoclast differentiation process, IF
staining revealed that the formation of the sealing zone was
complete on day 5 (approximately). At this stage, HMC microscopy
revealed that the cell morphology was characterized by a smooth
cell edge, no filopodia, a scattered distribution of nuclei and
strip-like protuberances at the edge of cell. These characteristics
were used as indicators for the addition of OPG in subsequent
experiments to examine the effects of OPG on the existing sealing
zone.
The sealing zone is a cytoskeletal structure unique
to mature osteoclasts. It not only provides a favorable closed
microenvironment for bone resorption, but also allows the firm
adhesion of osteoclasts onto the bone surface. The integrity of the
sealing zone is the prerequisite for the normal bone resorption
function of osteoclasts (4,13).
In our study, in the control group not treated with OPG, the
sealing zone remained intact. Following treatment with OPG at
different concentrations, the sealing zones showed defects or had
completely disappeared, and there was a scattered or clustered
arrangement of podosomes. In addition, investigations of the
resorption lacunae showed that the osteoclasts had lost their bone
resorption function in response to treatment with 20 ng/ml OPG.
MMP-9 and CAII are genes involved in the process of
osteoclast-mediated bone resorption. The proteins synthesized by
the 2 genes are secreted into the sealing zone and participate in
the decomposition of ossein and minerals (27,28). The downregulation of MMP-9 and
CAII mRNA expression corresponded to the effects of OPG on the bone
resorption activity of osteoclasts. All these findings suggested
that OPG caused the fracture or disappearance of the sealing zone
by altering the arrangement of podosomes. Hence, the bone
resorption activity of the osteoclasts was affected.
RhoGTPases are the molecular switches of signal
transduction pathways, allowing the regulation of a variety of
functions of the cell (14,16). A number of studies have also
confirmed that RhoGTPases control F-actin polymerization and
adhesion, and play an important role in the formation of the
sealing zone in osteoclasts (29–33). As shown by our previous study
(34) and a study by other
researchers (18), RhoH, RhoJ/TCL
and RhoE/Rnd3 are not expressed during osteoclast differentiation
and maturation. In addition, the expression of RhoQ/TC10, RhoV/Chp1
and RhoBTB2 is markedly inhibited during the formation of
osteoclasts. It is thus indicated that RhoH, RhoJ/TCL and Rnd3, as
well as RhoQ/TC10, RhoV/Chp1 and RhoBTB2 play no role in the
sealing zone formation process. Moreover, the effect of the Miro
family on actin is not known (15). The mRNA expression of the
remaining 14 members of the RhoGTPase family was analyzed in
osteoclasts following the disruption or disappearance of the
sealing zone as a result of OPG treatment. No significant changes
were detected in the mRNA expression of RhoC, Rac1, RhoE/Rnd3 and
Rnd2 following treatment with OPG; however, a significant
downregulation was observed in the mRNA expression of all the
remaining 10 members (RhoA, RhoB, cdc42v1, cdc42v2, RhoU/Wrch1,
RhoF/Rif, Rac2, RhoG, Rnd1 and RhoBTB1) of the RhoGTPase family
(P<0.01). In fact, RhoU expression was only approximately 20% of
that observed in the control group following treatment with OPG.
These results indicate that these 10 RhoGTPases play key a
regulatory role in the OPG-induced defects or disappearance of the
sealing zone in osteoclasts, with RhoU plays a more dominant
role.
The upstream kinase, RhoGEF, activates RhoGTPases
and catalyzes the conversion of GDP to GTP (17,35). In our study, Arhgef8/Net1 and
DOCK5 were significantly downregulated following treatment with
OPG. In light of the interaction between RhoGEF and RhoGTPases and
their roles in the formation of the sealing zone, we hypothesized
that Arhgef8/Net1 and DOCK5 are involved in the regulation of
RhoGTPases and are responsible for the damaging effects of OPG on
the sealing zone.
As the major effector of the Rho GTPase family, ROCK
receives the activating signal transmitted by RhoGTPases, which
provide conditions that are critical for dynamic changes in the
cytoskeleton, in particular, the nucleation and polymerization of
actin (36,37). The podosome contains an F-actin
core, surrounded by scaffold proteins, kinases and integrins. The
sealing zone is a large, compactly arranged F-actin ring composed
of podosomes. Currently, there are no antibodies available for the
detection of ROCK2; therefore, in our investigations, we detected
the mRNA expression levels of ROCK1 protein and ROCK2, both of
which were significantly downregulated following treatment with
OPG. These results indicate that ROCK1 and ROCK2 are involved in
the OPG-induced arrangement of podosomes, which result in defects
or destruction of the sealing zone. However, further studies are
required to investigate the complex mechanism of OPG action on the
sealing zone through the RhoGTPase signaling pathway.
In conclusion, OPG inhibited the expression of
Arhgef8/Net1 and DOCK5 (RhoGEF) and 10 of the 18 members of the
RhoGTPase family (RhoA, RhoB, cdc42v1, cdc42v2, RhoU/Wrch1,
RhoF/Rif, Rac2, RhoG, Rnd1 and RhoBTB1), all of which are involved
in the RhoGTPase signaling pathway. OPG also inhibited the 2
members of the ROCK family, ROCK1 and ROCK2. The OPG-mediated
effects on the arrangement and formation of podosomes, resulted in
defects or the destruction of the sealing zone and concomitant
reduction in the bone resorption activity of the osteoclasts.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (nos. 31172373, 31302154 and
31372495), the Specialized Research Fund for the Doctoral Program
of Higher Education (no. 20113250110003), the Priority Academic
Program Development of Jiangsu Higher Education Institutions and
the Graduate Innovation Project of Jiangsu Province
(CXZZ12_0917).
References
|
1
|
Boyle WJ, Simonet WS and Lacey DL:
Osteoclast differentiation and activation. Nature. 423:337–342.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lane NE and Rehman Q: Osteoporosis in the
rheumatic disease patient. Lupus. 11:675–679. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zambonin-Zallone A, Teti A, Carano A and
Marchisio PC: The distribution of podosomes in osteoclasts cultured
on bone laminae: effect of retinol. J Bone Miner Res. 3:517–523.
1988. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jurdic P, Saltel F, Chabadel A and
Destaing O: Podosome and sealing zone: specificity of the
osteoclast model. Eur J Cell Biol. 85:195–202. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ory S, Brazier H, Pawlak G and Blangy A:
Rho GTPases in osteoclasts: orchestrators of podosome arrangement.
Eur J Cell Biol. 87:469–477. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Luxenburg C, Geblinger D, Klein E, et al:
The architecture of the adhesive apparatus of cultured osteoclasts:
from podosome formation to sealing zone assembly. PLoS One.
2:e1792007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nesbitt SA and Horton MA: Trafficking of
matrix collagens through bone-resorbing osteoclasts. Science.
276:266–269. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Anderegg F, Geblinger D, Horvath P, et al:
Substrate adhesion regulates sealing zone architecture and dynamics
in cultured osteoclasts. PLoS One. 6:e285832011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Saltel F, Destaing O, Bard F, Eichert D
and Jurdic P: Apatite-mediated actin dynamics in resorbing
osteoclasts. Mol Biol Cell. 15:5231–5241. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Luxenburg C, Parsons JT, Addadi L and
Geiger B: Involvement of the Src-cortactin pathway in podosome
formation and turnover during polarization of cultured osteoclasts.
J Cell Sci. 119:4878–4888. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Akisaka T, Yoshida H and Suzuki R: The
ruffled border and attachment regions of the apposing membrane of
resorbing osteoclasts as visualized from the cytoplasmic face of
the membrane. J Electron Microsc (Tokyo). 55:53–61. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schachtner H, Calaminus SD, Thomas SG and
Machesky LM: Podosomes in adhesion, migration, mechanosensing and
matrix remodeling. Cytoskeleton (Hoboken). 70:572–589. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chiusaroli R, Knobler H, Luxenburg C, et
al: Tyrosine phosphatase epsilon is a positive regulator of
osteoclast function in vitro and in vivo. Mol Biol Cell.
15:234–244. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Itzstein C, Coxon FP and Rogers MJ: The
regulation of osteoclast function and bone resorption by small
GTPases. Small GTPases. 2:117–130. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Vega FM and Ridley AJ: SnapShot: Rho
family GTPases. Cell. 129:14302007.PubMed/NCBI
|
|
16
|
Coxon FP and Rogers MJ: The role of
prenylated small GTP-binding proteins in the regulation of
osteoclast function. Calcif Tissue Int. 72:80–84. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rossman KL, Der CJ and Sondek J: GEF means
go: turning on RHO GTPases with guanine nucleotide-exchange
factors. Nat Rev Mol Cell Biol. 6:167–180. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Brazier H, Stephens S, Ory S, Fort P,
Morrison N and Blangy A: Expression profile of RhoGTPases and
RhoGEFs during RANKL-stimulated osteoclastogenesis: identification
of essential genes in osteoclasts. J Bone Miner Res. 21:1387–1398.
2006. View Article : Google Scholar
|
|
19
|
Vives V, Laurin M, Cres G, et al: The Rac1
exchange factor Dock5 is essential for bone resorption by
osteoclasts. J Bone Miner Res. 26:1099–1110. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee JH, Katakai T, Hara T, Gonda H, Sugai
M and Shimizu A: Roles of p-ERM and Rho-ROCK signaling in
lymphocyte polarity and uropod formation. J Cell Biol. 167:327–337.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nakagawa O, Fujisawa K, Ishizaki T, Saito
Y, Nakao K and Narumiya S: ROCK-I and ROCK-II, two isoforms of
Rho-associated coiled-coil forming protein serine/threonine kinase
in mice. FEBS Lett. 392:189–193. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang J, Chen TY, Qin S, Duan Y and Wang G:
Inhibitory effect of metformin on bone metastasis of cancer via
OPG/RANKL/RANK system. Med Hypotheses. 81:805–806. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hofbauer LC, Kühne CA and Viereck V: The
OPG/RANKL/RANK system in metabolic bone diseases. J Musculoskelet
Neuronal Interact. 4:268–275. 2004.PubMed/NCBI
|
|
24
|
Shiotani A, Takami M, Itoh K, Shibasaki Y
and Sasaki T: Regulation of osteoclast differentiation and function
by receptor activator of NFkB ligand and osteoprotegerin. Anat Rec.
268:137–146. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
O’Brien EA, Williams JH and Marshall MJ:
Osteoprotegerin is produced when prostaglandin synthesis is
inhibited causing osteoclasts to detach from the surface of mouse
parietal bone and attach to the endocranial membrane. Bone.
28:208–214. 2001.
|
|
26
|
Song RL, Liu XZ, Zhu JQ, et al: RhoV
mediated the apoptosis of RAW2647 macrophages caused by osteoclast
differentiation. Mol Med Rep. (In press).
|
|
27
|
Borthwick KJ, Kandemir N, Topaloglu R, et
al: A phenocopy of CAII deficiency: a novel genetic explanation for
inherited infantile osteopetrosis with distal renal tubular
acidosis. J Med Genet. 40:115–121. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Everts V, Delaissé JM, Korper W, Niehof A,
Vaes G and Beertsen W: Degradation of collagen in the
bone-resorbing compartment underlying the osteoclast involves both
cysteine-proteinases and matrix metalloproteinases. J Cell Physiol.
150:221–231. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dovas A, Gevrey JC, Grossi A, Park H,
Abou-Kheir W and Cox D: Regulation of podosome dynamics by WASp
phosphorylation: implication in matrix degradation and chemotaxis
in macrophages. J Cell Sci. 122:3873–3882. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gringel A, Walz D, Rosenberger G, et al:
PAK4 and alphaPIX determine podosome size and number in macrophages
through localized actin regulation. J Cell Physiol. 209:568–579.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Guegan F, Tatin F, Leste-Lasserre T,
Drutel G, Genot E and Moreau V: p190B RhoGAP regulates
endothelial-cell-associated proteolysis through MT1-MMP and MMP2. J
Cell Sci. 121:2054–2061. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Osiak AE, Zenner G and Linder S:
Subconfluent endothelial cells form podosomes downstream of
cytokine and RhoGTPase signaling. Exp Cell Res. 307:342–353. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tatin F, Grise F, Reuzeau E, Genot E and
Moreau V: Sodium fluoride induces podosome formation in endothelial
cells. Biol Cell. 102:489–498. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Song RL, Liu XZ, Zhu JQ, et al: New roles
of filopodia and podosomes in the differentiation and fusion
process of osteoclasts. Genet Mol Res. 13:4776–4787. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Schmidt A and Hall A: Guanine nucleotide
exchange factors for Rho GTPases: turning on the switch. Genes Dev.
16:1587–1609. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Schofield AV and Bernard O: Rho-associated
coiled-coil kinase (ROCK) signaling and disease. Crit Rev Biochem
Mol Biol. 48:301–316. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zeidan A, Paylor B, Steinhoff KJ, et al:
Actin cytoskeleton dynamics promotes leptin-induced vascular smooth
muscle hypertrophy via RhoA/ROCK- and phosphatidylinositol
3-kinase/protein kinase B-dependent pathways. J Pharmacol Exp Ther.
322:1110–1116. 2007. View Article : Google Scholar
|