Introduction
Hepatitis B, which is caused by hepatitis B virus
(HBV), is a key infectious disease and a major health concern
worldwide. In China alone, there are 130 million HBV carriers and
30 million liver disease patients, of whom 300 thousand succumb to
the disease annually (1).
Vaccines and appropriate immunization strategies have decreased the
infectious rate of HBV, especially in young people. However, with
regard to the infected and chronic hepatitis B patients, because of
drug resistance and side-effects, the curative effects of current
anti-HBV therapies (nucleoside analogues and interferons) are still
not ideal (2).
The HBV genome contains the coding regions P, S, C
and X. The proteins (antigens) encoded by these regions are key
markers for the detection of HBV infection. It is believed that
targeting the proteins involved in the life cycle of HBV is an
ideal strategy or orientation for the treatment of HBV and related
liver diseases (3). The core
protein (HBc) encoded by the C region is crucial in the assembly of
the nucleocapsid of HBV, which is composed of 180 or 240 copies of
core proteins, and affects its envelopment (4). Thus, drugs targeting the core
protein may effectively inhibit the maturation of HBV, which is the
end-point of its complete life cycle (5). Therefore, focus on the intervention
of HBc is essential in the identification of anti-HBV therapy.
As a new class of adaptive molecules, aptamer is a
single-strand DNA or RNA screened through the SELEX (systematic
evolution of ligands by exponential enrichment) technique that can
target a wide range of metallic ions, polypeptides, proteins or
even cells (6,7). Considered as a nucleic acid
antibody, the sensitivity and specificity of aptamer is comparable
or even superior to conventional antibodies. The classical example
of the application of aptamer is pegaptanib (8,9),
which is the aptamer targeting vascular endothelial growth factor
(VEGF) and used to treat age-related macular degeneration. As the
first approved aptamer drug, it has become a milestone in studies
on aptamers.
HIV (10) and HCV
(11–13) have been the focus of many
antiviral aptamer studies. The most significant came from an in
vivo therapeutic attempt of an anti-HIV gp120 aptamer
conjugated with siRNA (14,15). Concerning HBV, aptamers targeting
S protein (16) and the
polymerase (17) were also
reported. These findings may definitely improve the use of drugs in
clinic as well as drug development.
In the present study, we successfully screened a DNA
aptamer targeting HBc. To the best of our knowledge, the findings
of this study, reported for the first time, hold great prospect in
inhibiting the maturation and replication of HBV. Therefore, the
aptamer is a potentially useful tool that can be used in the
diagnosis of HBV-related diseases and the development of a
drug-targeted delivery system (18).
Materials and methods
Expression and purification of core
protein dimers
The core protein sequence was amplified from the
plasmid pBS_HBV3.6II, which was obtained from the Shanghai Medical
School, Fudan University and contained complete sequences of HBV
genome using the primers: 5′-GCCCATATGGACA TTGACCCGTA-3′ and
5′-GCCCTCGAGTCAAACAACAG TAGTTT-3′. The sequence was then cloned
into the NdeI/XhoI site of pET28a. The recombinant
plasmid pET28a-HBc was transformed into Escherichia coli
BL21(DE3) to express the core protein at 32°C. The plain pET28a was
also transformed as a negative control.
Following induction with 1.0 mM IPTG for 4 h, the
transformed BL21(DE3) cells were washed and re-suspended in the
binding buffer (50 mM NaH2PO4, 300 mM NaCl,
10 mM imidazole, pH 8.0). Ultrasonication was carried out in an ice
bath with 2–5 sec of 100 W ultrasound 60–80 times. The supernatant
was mixed with half the volume of Ni-NTA Agarose beads (Qiagen) and
agitated for 1 h. After being washed by the wash buffer (binding
buffer supplemented with 20 mM imidazole) and eluted by the elution
buffer (binding buffer supplemented with 250 mM imidazole), the
purified proteins were ultrafiltrated to change the solvent from
the elution buffer to the binding buffer containing 3 M urea, pH
9.5 (to prevent core protein dimers from auto-assembling into
nucleocapsids). The proteins were again mixed with half the volume
of Ni-NTA Agarose beads and agitated for 30 min at 4°C. The
conjugates (Ni-NTA agarose beads conjugated with core protein
dimers or the negative control) were washed and suspended in the
binding buffer with the addition of Complete Protease Inhibitor
(Roche Diagnostics, Mannheim, Germany) and stored at 4°C.
SELEX
An ssDNA library (5′-ACGCTCGGATGCCACTA
CAGN40CTCATGGACGTGCTGGTGAC-3′) was synthesized by Sangon
Biotech (Shanghai, China). The ssDNA pool (200 pmol) (2.5 nmol
ssDNA library was used for the initial round) was dissolved in
PBS+Mg (PBS supplemented with 1 mM MgCl2) and was
denatured at 95°C for 8 min and quickly cooled on ice. Negative
control conjugates (40 pmol) were washed twice by PBS+Mg and
incubated with the ssDNA pool in a total volume of 1,000 μl of
PBS+Mg+tRNA (PBS+Mg supplemented with 0.1 mg/ml yeast tRNA) for 30
min at 37°C, in an orbital shaker. The percolate containing unbound
ssDNA sequences was then obtained through ultrafiltration, and was
incubated with positive conjugates (consisting of core proteins and
also washed with PBS+Mg twice), in an orbital shaker for 30 min at
37°C. The mixture was ultrafiltrated and the remaining conjugates
were washed three times by PBS+Mg to remove unbound ssDNA
sequences. The bound sequences were eluted by 10% trypsin (10 min
at 37°C) followed by heating at 95°C for 10 min and centrifugation
at 5,000 rpm for 5 min. The sequences were amplified by PCR using
the forward primer (5′-ACGCTCGGATGCCACTA CAG-3′) and biotin-labeled
reverse primer (5′-GTCACCAG CACGTCCATGAG-3′). The selected sense
DNA strands were separated from the biotinylated antisense strands
by alkaline denaturation (0.2 M NaOH) and affinity purification
with streptavidin-coated Sepharose beads (GE Healthcare, Uppsala,
Sweden). The entire selection procedure was repeated for several
rounds according to the extent of enrichment. The enriched pool was
amplified by PCR using unlabeled primers and the PCR products were
cloned into Escherichia coli DH5a using pMD19 T-vector
(Takara Bio Inc., Shiga, Japan) and the positive clones were
sequenced and aligned by MACAW software (NIH, Bethesda, MD,
USA).
Monitoring the enrichment of aptamer
pools
The aptamer pool of each SELEX round was incubated
with positive conjugates, ultrafiltrated, washed and eluted as
described above. Quantitative PCR (qPCR) reactions (10 μl reaction
volume) were carried out using SYBR Premix Ex Taq™ (Takara Bio
Inc.) according to the manufacturer’s instructions in an Applied
Biosystems 7500 sequence detection system (Applied Biosystems,
Foster City, CA, USA). The proportion of bound sequences, which
indicated the enrichment of the aptamer pool, was calculated as: 2
(Ct of input pool − Ct of elusion) × 100%.
Dot blot
Purified HBc (2 μl; 10 μM) was spotted onto a
polyvinylidene difluoride (PVDF) membrane (Bio-Rad Laboratories,
Hercules, CA, USA) pretreated with methyl alcohol. After drying,
the membrane was incubated with 5% BSA in PBS+Mg+tRNA (1 h, room
temperature) to block non-specific sites. Subsequently, it was
incubated with 200 μl candidate aptamer (0.1 μM) in PBS+Mg+tRNA (1
h, 37°C). After three washes with TBST, it was incubated with 200
μl streptavidin-horseradish peroxidase conjugate (Rpn1231; Amersham
Biosciences, Piscataway, NJ, USA) at a dilution of 1:1,000 in TBST
(45 min, 37°C). After another three washes, it was detected by
SuperSignal West Pico Chemiluminescent Substrate (Thermo Fisher
Scientific, Rockford, IL, USA) according to the manufacturer’s
instructions.
Cell culture and transfection
HBV DNA integrated cell line HepG2.2.15, which is
derived from the HepG2 human hepatoma cell line and is
characterized by stable HBV expression and replication, was
maintained in Dulbecco’s modified Eagle’s medium (DMEM) containing
10% fetal bovine serum (FBS) at 37°C in a humidified atmosphere
with 5% CO2.
For the transfection experiments, HepG2.2.15 cells
were seeded in 6-well plates at a density of 4×105 in 2
ml medium/well. After 24 h of incubation, transfection was
accomplished with Hifectin III transfection reagent (Applygen
Technologies, Beijing, China) according to the manufacturer’s
instructions. In brief, 8 μg aptamer mixed with 8 μl transfection
reagent was applied to each well. Transfection reagent alone was
applied as an experimental control. The cells were grown for a
further 24, 48 and 72 h prior to harvest.
Isolation and quantification of HBV DNA,
HBsAg and HBeAg
The intracellular HBV DNA was isolated by TRI
reagent (Sigma-Aldrich Inc., St. Louis, MO, USA) according to the
manufacturer’s instructions. The extracellular HBV DNA in culture
medium was extracted by the AxyPrep Blood Genomic DNA Miniprep kit
(Axygen Biosciences, Union City, CA, USA). Following the
extraction, the absorbance at 260/280 nm was measured using a
NanoDrop ND-1000 spectrophotometer (Bio-Lab Australia Pty Ltd.,
Mulgrave, VIC, Australia) and the concentrations were
calculated.
HBV DNA was quantified by qPCR as described above,
with the primers described by Jun-Bin et al (19). The amplification program was:
pre-denaturation at 95°C for 30 sec, 40 cycles of 95°C for 5 sec
and 64°C for 34 sec.
HBsAg and HBeAg in culture medium were determined by
using the relative quantitative ELISA kits (Shanghai Kehua
Bio-engineering Co., Ltd., Shanghai, China) (20). In brief, the culture media were
diluted to a ratio of 1:6 with PBS and added to plates coated with
antibody against HBsAg or HBeAg, respectively. After 30 min of
incubation at 37°C, the plates were washed and incubated with
substrate solution A or B, respectively (15 min at 37°C, protected
from light). The reaction was stopped by the termination solution
and the optical density (OD) value at 450 nm was measured.
Assay for HBV nucleocapsids
To separate the nucleocapsids based on Calvert and
Summers’ protocol (21), cell
lysates, which were achieved by cell lysis buffer for western blot
analysis and IP (Beyotime Institute of Biotechnology, Haimen,
China), were loaded onto native 0.8% agarose-Tris-acetate-EDTA
(TAE) gel electrophoresis. The bands were blotted onto PVDF
membrane overnight at 4°C by capillary transfer in 10X SSC buffer
(1.5M NaCl, 150 mM sodium citrate, pH 7.0). After drying, similar
to the general western blot analysis, the membrane was rinsed in
TBST containing 5% skim milk to block non-specific sites, followed
by three washes in TBST, with agitation for 10 min each. The
membrane was then incubated with the antibody against HBc (Santa
Cruz Biotechnology, Santa Cruz, Inc., CA, USA) at a dilution of
1:500 in solution 1 of the SignalBoost Immunoreaction Enhancer kit
(Merck KGaA, Darmstadt, Germany) for 2 h. It was then washed as
described above and incubated with a horseradish peroxidase (HRP)
conjugated secondary antibody (Santa Cruz Biotechnology, Inc.) at a
dilution of 1:1,000 in solution 2 of the SignalBoost Immunoreaction
Enhancer kit for 2 h. After another three washes, it was detected
by Immobilon Western Chemiluminescent HRP Substrate (Millipore
Corp., Billerica, MA, USA) according to the manufacturer’s
instructions. The β-actin bands in cell lysates, obtained by
general western blot analysis, were used as an internal
reference.
Results
An aptamer pool binding to HBc was
screened from an ssDNA library
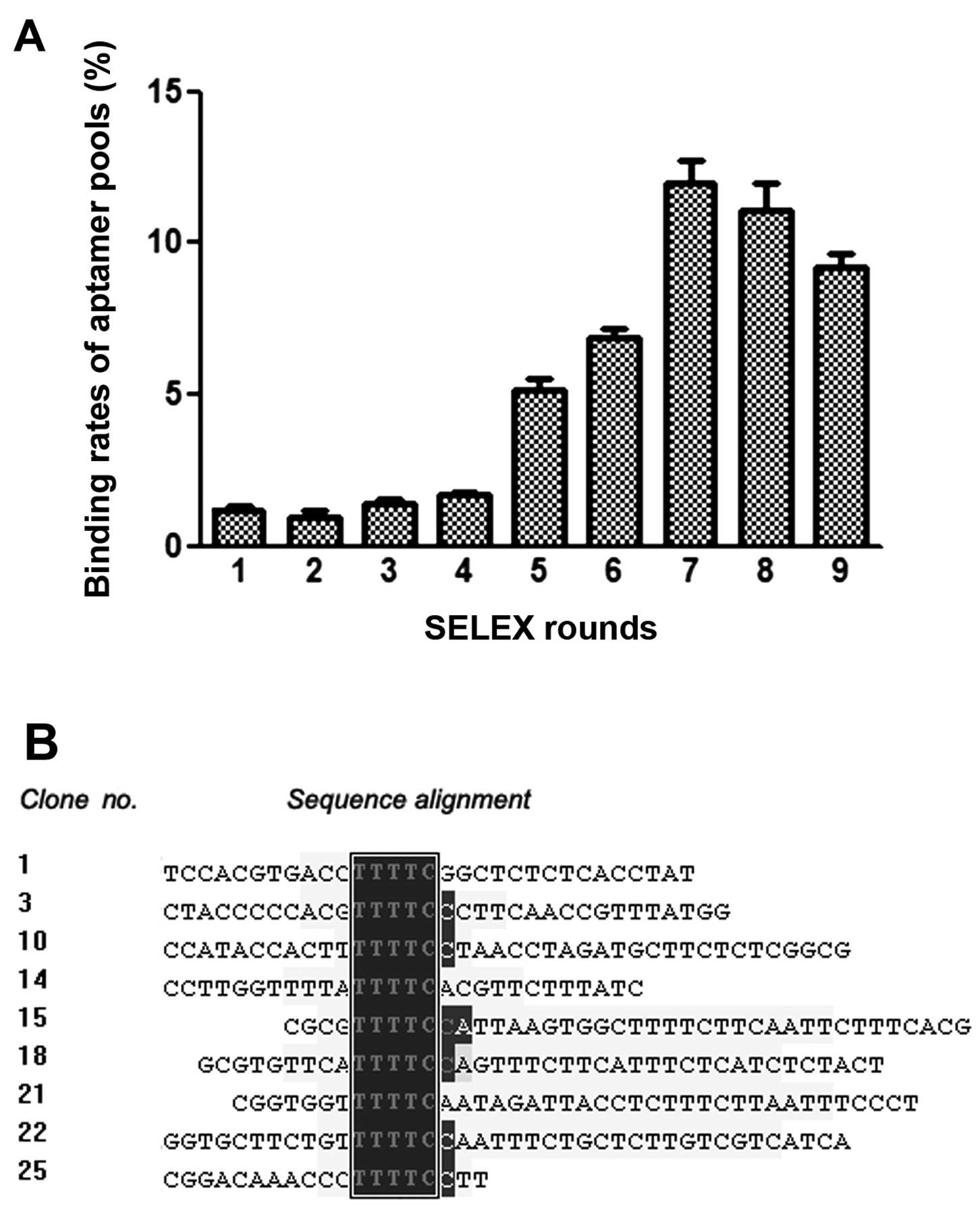
To screen for aptamers against HBc, we performed
nine rounds of SELEX with His-tagged HBc as target and His-tagged
plain vector as a counter-selector. Enrichment was observed
following qPCR of ssDNA pools of all rounds (Fig. 1A). It was shown that the
proportion of bound sequences increased with the proceeding of the
SELEX, until round seven. Subsequently, the pools ceased to enrich
and were even slightly attenuated, possibly due to amplification
bias (22). Thus, the pool of
round seven was expected to contain the ideal aptamer(s) and was
subsequently amplified and cloned.
A total of 90 positive clones were sequenced. By
analysis in MACAW, most of the clones were subdivided into seven
groups, each of which contained a consensus five-letter sequence
(Fig. 1B). In particular, clone
No.28 had a replicate in sequence (No.33), as was the case in clone
No.29 (No.66). Thus, one clone each from the five largest groups
was selected for subsequent studies, i.e., clone No.1, 12, 16, 28
and 29 from group 1, 5, 4, 2 and 3, respectively, among which the
replicated clones (No.28 and No.29) were specifically selected
(Table I).
 | Table IThe selected aptamer candidates. |
Table I
The selected aptamer candidates.
| Clone no. | Sequence |
|---|
| 1 |
5′-ACGCTCGGATGCCACTACAGCGCTCCTTATCCACGTGACCTTTTCGGCTCTCTCACCTATCTCATGGACGTGCTGGTGAC-3′ |
| 12 |
5′-ACGCTCGGATGCCACTACAGCAGCGAGCTGGGCGCATTTTTGTTATCCCTAGTGCTCAAGCTCATGGACGTGCTGGTGAC-3′ |
| 16 |
5′-ACGCTCGGATGCCACTACAGGTACTCCGGTTAATCGCAATATTGCTACGAGTAGCTTCTCCTCATGGACGTGCTGGTGAC-3′ |
| 28 |
5′-ACGCTCGGATGCCACTACAGCTTCCCCTAATCTGGCGCTCTCATCTAATTTCCCTTCCTGCTCATGGACGTGCTGGTGAC-3′ |
| 29 |
5′-ACGCTCGGATGCCACTACAGGGTGGGATTCTATTTTATTCTGATTCTCCGTTCACTTTGCCTCATGGACGTGCTGGTGAC-3′ |
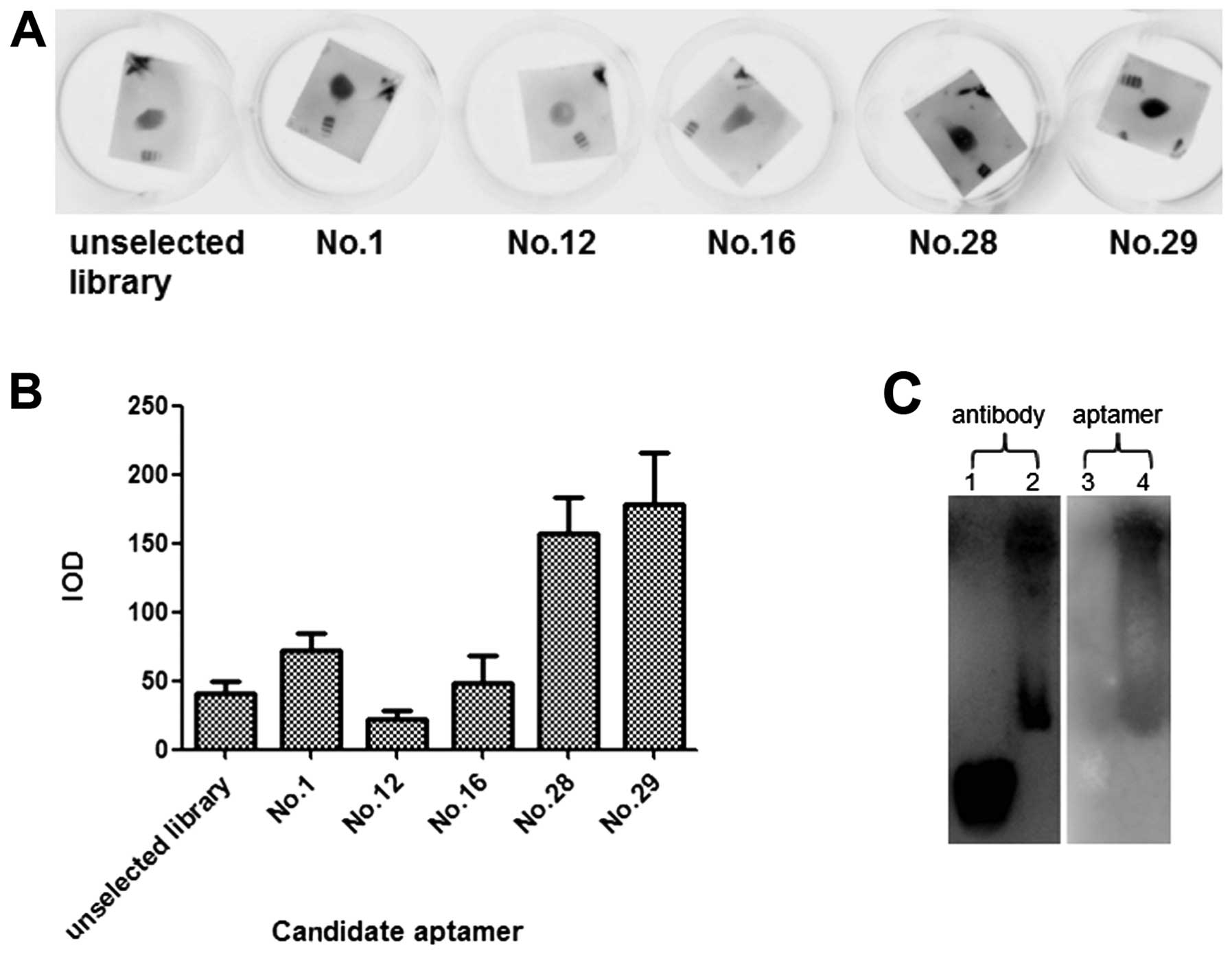
Candidate aptamer Apt.No.28 has high
affinity and specificity to HBc
To verify the binding affinity of the candidate
aptamers, a dot blot was carried out with purified HBc dotted on
PVDF membrane and candidate aptamers in the liquid phase. It was
shown that as compared to the unselected library, candidate
aptamers had divergent affinity to the target, among which
Apt.No.28 and No.29 had the highest (Fig. 2A and B). Thus, these two
candidates were subjected to the subsequent studies.
We also tested Apt.No.28 in the detection of HBV
nucleocapsids from cell lysates of HepG2.2.15 and compared it to
the HBc antibody. Both Apt.No.28 and the antibody were feasible in
the detection of the nucleocapsids (Fig. 2C). However, the antibody also
strongly stained the protein markers while the aptamer did not
stain these markers, suggesting that the aptamer was more specific
to HBc and the nucleocapsids.
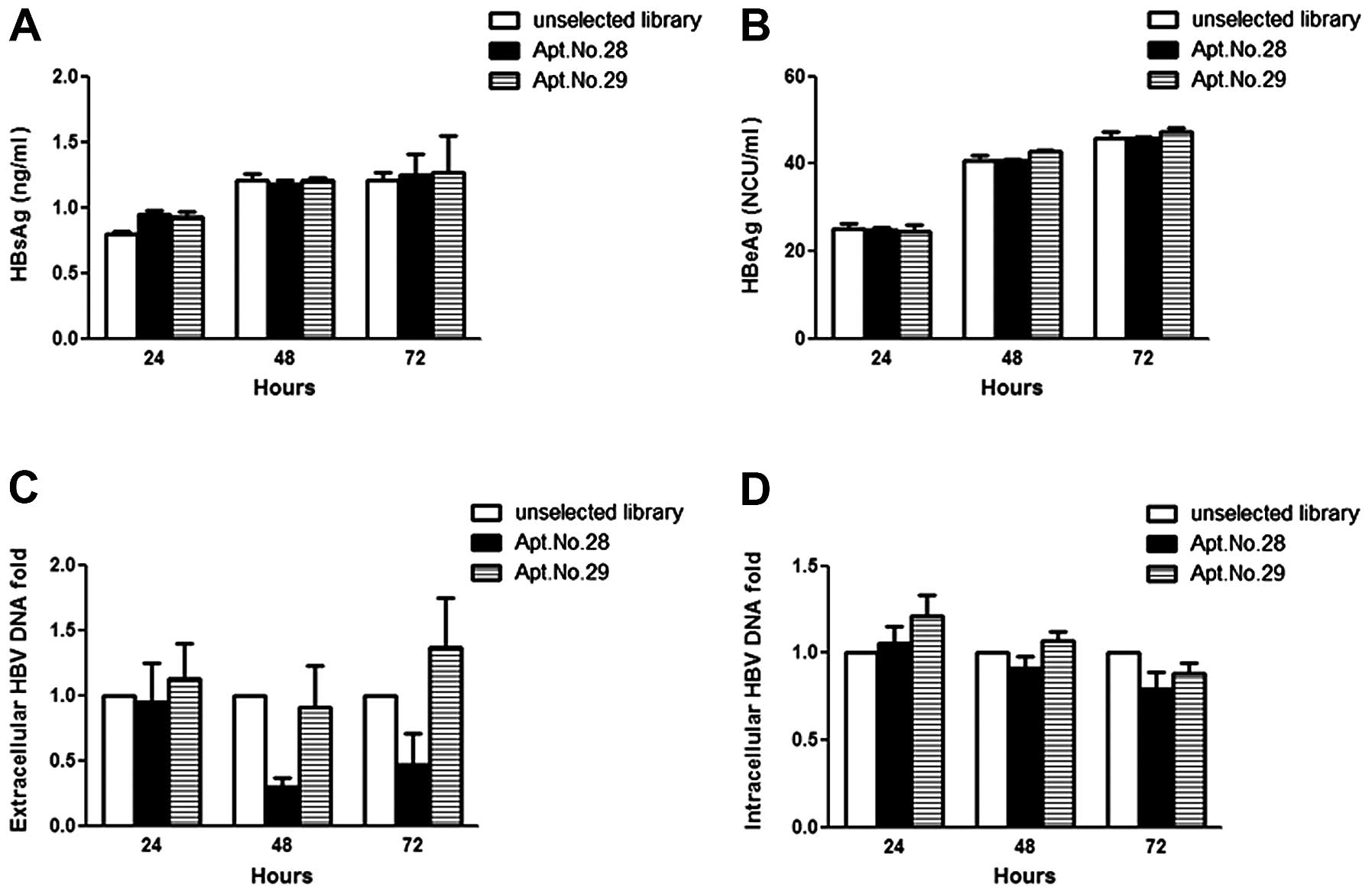
Apt.No.28 attenuates extracellular HBV
DNA but not the secretion of HBV capsids
In order to investigate whether the candidate
aptamers targeting HBc were able to interfere with the replication
of HBV, ELISA assay and qPCR were used to quantify the HBV of
aptamer-transfected HepG2.2.15. In the ELISA assay, HepG2.215 did
not lose the ability of secreting HBV capsids (no decrease in HBsAg
or HBeAg) following transfection with Apt.No.28 or No.29 (Fig. 3A and B). However, the alteration
was observed on extracellular HBV DNA by qPCR analysis. As shown in
Fig. 3C, 24 h after the
transfection, the level of extracellular HBV DNA in the groups
transfected with Apt.No.28 and No.29 showed no significant
difference to that transfected with the unselected library. At 48
h, the amount of extracellular HBV DNA in the group of Apt.No.28
was significantly less, while that in the group of Apt.No.29 showed
little difference. The trend at 72 h was similar to that at 48 h.
Apt.No.28 and No.29 did not reduce the intracellular level of HBV
DNA (Fig. 3D). These findings
indicated that Apt.No.28 was able to inhibit the replication of HBV
by suppressing the synthesis of HBV DNA.
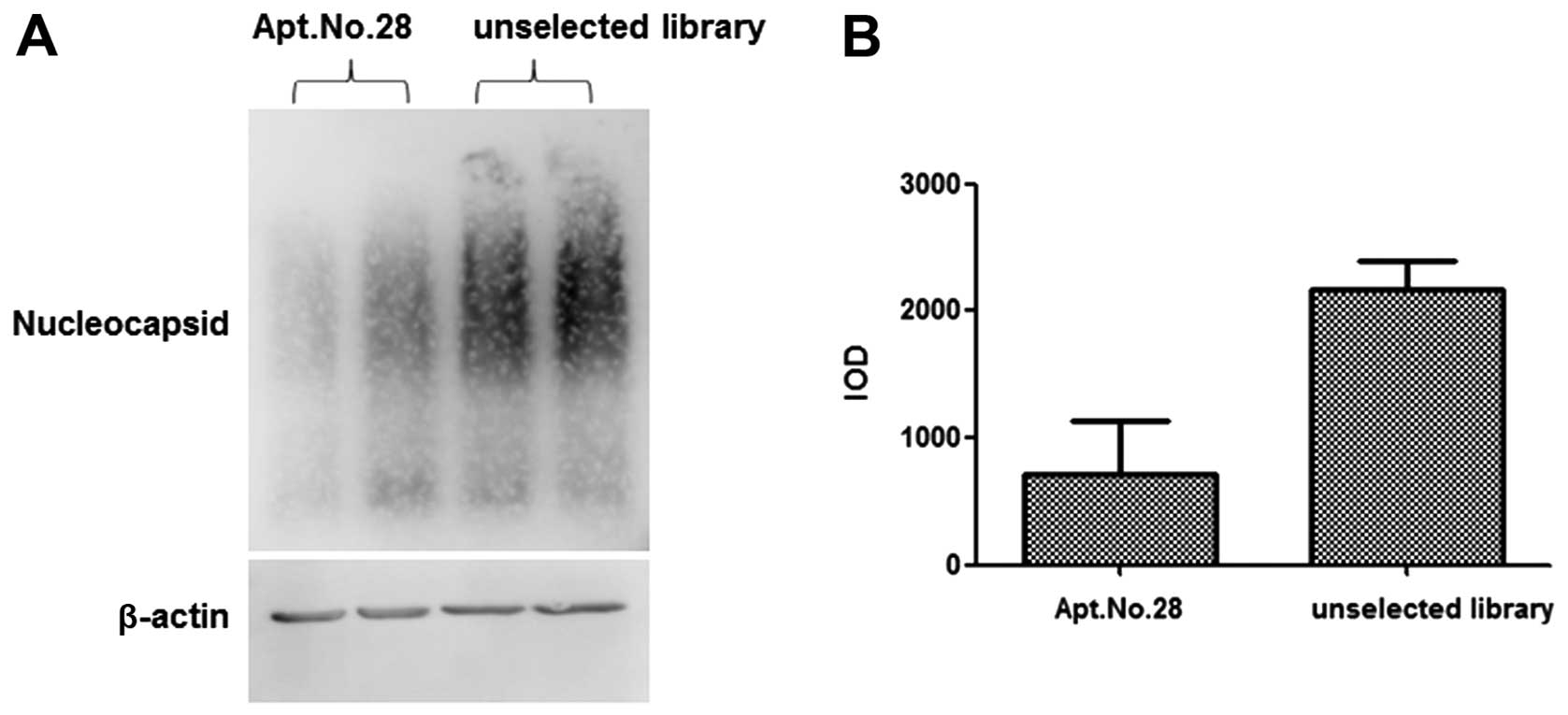
Apt.No.28 inhibits the assembly of
nucleocapsids
To study the mechanism involved in the inhibition of
HBV replication by Apt.No.28, the total intracellular protein was
extracted from HepG2.2.15 cells transfected with Apt.No.28 for 72
h. Using agarose gel electrophoresis, capillary transfer and
blotting, it was shown that the nucleocapsids formed by HBc were
significantly less in the group treated with Apt.No.28 than those
in the group treated with the library (Fig. 4A and B). These findings indicated
that Apt.No.28 inhibited HBV replication by interfering with the
assembly of nucleocapsid, which was essential to the synthesis of
HBV DNA.
Discussion
Although HBV was identified in 1885 and its basic
constructional elements have already been isolated and identified,
drugs with high specificity to HBV are under development, due to
the easy occurrence of side-effects and drug resistance (23). Thus, new technologies are required
to solve these problems and overcome limitations to HBV. Aptamer is
a newly developed adaptive molecule that shows high sensitivity and
specificity, while exhibiting no toxicity or immunogenicity
(8,24,25). It becomes more and more prevalent
in basic research as well as clinical practice. Thus, it is
believed that its use in antiviral research may reveal the
propagation, block the infection and inactivate the viability of
viruses. The aptamers for HCV and HIV have been developed thus far
and are altering antiviral investigations as well as clinical
diagnosis and treatments (10,13). In particular, the in vivo
therapeutic use of a HIV aptamer has been reported in Science
Translational Medicine (14).
However, the application of aptamers in anti-HBV research is in the
early stages, most probably due to the complexity of HBV.
HBc is the major component of the nucleocapsid
packaging HBV genome. It is involved in the processes of viral
replication and regulation, such as RNA packaging, DNA synthesis,
recognition of virus envelope, maturation and budding of virus. Of
note, HBc is capable of assembling the nucleocapsid in a native
shape (with 180 or 240 core protein units) automatically, in
circumstances lacking all other viral components (26,27). Therefore, the control of HBc and
the nucleocapsids may be essential to anti-HBV strategies. Thus, we
screened DNA aptamers targeting HBc in this study.
There are currently few reports regarding the
technology of the expression and purification of HBc. The most
common approach is to express HBc as inclusion bodies, which is
subsequently purified by a process of wash, denaturation and
renaturation (28–30). However, the manipulation of
inclusion bodies can easily cause the loss of proteins, while
renaturation remains a challenge (31). Thus, when the expression level is
relatively low, proteins dissolved in the bacterial cytoplasm are
easier to extract with large quantity and good solubility in the
supernatant following ultrasonication (32). Furthermore, since HBc was
automatically assembled into the nucleocapsid form while the dimer
form was required for aptamer selection, the extracted core
proteins were further depolymerized to dimers by the treatment of
urea and alkali and bound to the Ni-NTA agarose beads for
purification in the present study.
The fact that HBc is automatically assembled to
polymers in the neutral pH range makes the protein purification and
aptamer selection complicated and limits the methods available for
aptamer identification. In this study, dot blot was used to verify
the affinity of aptamers. In order to maintain its dimeric form,
HBc was also treated with urea and alkali, dotted onto a PVDF
membrane, dried and adjusted to a neutral pH range. In contrast to
the aptamer studies targeting S protein or the polymerase of HBV,
the automatic polymerization of HBc may account for the limitation
in the study of its aptamers. Findings of this study may facilitate
additional investigations of HBc.
The sequence replication of clones was a typical
characteristic of aptamer enrichment. Apt.No.28 and No.29, which
had sequence replicates, were shown to have high affinity to HBc,
although the possibility could not be excluded that clones with
higher affinity and even functional effects were not selected for
the present study.
To value the function of candidate aptamers in live
cells, we transferred them into HepG2.2.15 cells by transfection
reagent to test their inhibitory effect on HBV replication. As the
main indices of HBV level, it was identified that HBV capsid (shown
by HBsAg and HBeAg) was not inhibited by the aptamer Apt.No.28,
whereas extracellular HBV DNA was inhibited by the aptamer
Apt.No.28. This finding suggests that the inhibitory effect of
Apt.No.28 on HBV replication did not occur during the expression of
HBV proteins, but occurred prior to synthesis of HBV DNA reversely
transcribed by HBV RNA, which was after and depended on the
assembly of the nucleocapsid. By agarose gel electrophoresis,
capillary transfer and blotting, we found that the assembly of the
nucleocapsid was inhibited by Apt.No.28. Therefore, it was
confirmed that the aptamer inhibited the replication of HBV by
binding to core protein dimers and inhibiting the assembly of
nucleocapsid.
The inhibitory effect of Apt.No.28 was to some
extent limited by cell transfection. The transfection reagent used
in this study was Hifectin III, which was specific for the
transfection of ssDNA. We had also tried a general transfection
reagent Lipofectamine 2000 (Invitrogen), but yielded low
transfection efficiency. To strengthen the effect of Apt.No.28, a
targeted delivery system may be required. A conjugation with an
aptamer against hepatocytes is an ideal strategy (e.g., the aptamer
against hepatoma HA09) (22).
Previous studies have described peptide aptamers
that could specifically bind to HBc (20,33,34). However, considering certain
disadvantages of polypeptides and proteins such as instability and
immunogenicity, the DNA aptamer we identified, which interfered
with HBV replication at the stage of nucleocapsid assembly, was
thought to have greater potential in the detection of HBc, and in
the treatment of HBV-related diseases.
Nevertheless, additional studies are required to
identify the underlying structural basis and molecular mechanism of
the recognition and polymerization inhibition of HBc by the
aptamer. Studies should also be carried out to examine the
potential effects of the aptamer on HBV, such as the anti-HBV
effect of modified aptamers in vivo.
Acknowledgements
We would like to thank Dr Youhua Xie (from the
Shanghai Medical School, Fudan University) for kindly providing the
plasmid pBS_HBV3.6II. We also thank Drs Li Xu and Xiaohong Fang
(from the Institute of Chemistry, Academy of Sciences, China) for
their assistance in designing the SELEX protocol. The present study
was supported by grants from the National Natural Science
Foundation of China (nos. 81125001 and 91129702) and the Ministry
of Science and Technology of China (no. 2010CB732405).
References
|
1
|
Ma X, Lin C and Zhen W: Cancer care in
China: a general review. Biomed Imaging Interv J.
4:e392008.PubMed/NCBI
|
|
2
|
Vigano M and Lampertico P: Antiviral drugs
for HBV liver disease. Expert Opin Biol Ther. 11:285–300. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yan H, Zhong G, Xu G, et al: Sodium
taurocholate cotransporting polypeptide is a functional receptor
for human hepatitis B and D virus. Elife. 1:e000492012.PubMed/NCBI
|
|
4
|
Roseman AM, Berriman JA, Wynne SA, Butler
PJ and Crowther RA: A structural model for maturation of the
hepatitis B virus core. Proc Natl Acad Sci USA. 102:15821–15826.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Deres K, Schroder CH, Paessens A, et al:
Inhibition of hepatitis B virus replication by drug-induced
depletion of nucleocapsids. Science. 299:893–896. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kulbachinskiy AV: Methods for selection of
aptamers to protein targets. Biochemistry (Moscow). 72:1505–1518.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sefah K, Shangguan D, Xiong X, O’Donoghue
MB and Tan W: Development of DNA aptamers using Cell-SELEX. Nat
Protoc. 5:1169–1185. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chakravarthy U, Adamis AP, Cunningham ETJ,
et al: Year 2 efficacy results of 2 randomized controlled clinical
trials of pegaptanib for neovascular age-related macular
degeneration. Ophthalmology. 113:1508.e1501–1508.e1525.
2006.PubMed/NCBI
|
|
9
|
Ng EW, Shima DT, Calias P, Cunningham ET
Jr, Guyer DR and Adamis AP: Pegaptanib, a targeted anti-VEGF
aptamer for ocular vascular disease. Nat Rev Drug Discov.
5:123–132. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ramalingam D, Duclair S, Datta SA,
Ellington A, Rein A and Prasad VR: RNA aptamers directed to human
immunodeficiency virus type 1 Gag polyprotein bind to the matrix
and nucleocapsid domains and inhibit virus production. J Virol.
85:305–314. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kikuchi K, Umehara T, Fukuda K, Kuno A,
Hasegawa T and Nishikawa S: A hepatitis C virus (HCV) internal
ribosome entry site (IRES) domain III-IV-targeted aptamer inhibits
translation by binding to an apical loop of domain IIId. Nucleic
Acids Res. 33:683–692. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kikuchi K, Umehara T, Nishikawa F, Fukuda
K, Hasegawa T and Nishikawa S: Increased inhibitory ability of
conjugated RNA aptamers against the HCV IRES. Biochem Biophys Res
Commun. 386:118–123. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen F, Hu Y, Li D, Chen H and Zhang XL:
CS-SELEX generates high-affinity ssDNA aptamers as molecular probes
for hepatitis C virus envelope glycoprotein E2. PLoS One.
4:e81422009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Neff CP, Zhou J, Remling L, et al: An
aptamer-siRNA chimera suppresses HIV-1 viral loads and protects
from helper CD4+ T cell decline in humanized mice. Sci
Transl Med. 3:66ra662011.PubMed/NCBI
|
|
15
|
Zhou J, Li H, Li S, Zaia J and Rossi JJ:
Novel dual inhibitory function aptamer-siRNA delivery system for
HIV-1 therapy. Mol Ther. 16:1481–1489. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu J, Yang Y, Hu B, et al: Development of
HBsAg-binding aptamers that bind HepG2.2.15 cells via HBV surface
antigen. Virol Sin. 25:27–35. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Feng H, Beck J, Nassal M and Hu KH: A
SELEX-screened aptamer of human hepatitis B virus RNA encapsidation
signal suppresses viral replication. PLoS One. 6:e278622011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ray P and White RR: Aptamers for targeted
drug delivery. Pharmaceuticals. 3:1761–1778. 2010. View Article : Google Scholar
|
|
19
|
Jun-Bin S, Zhi C, Wei-Qin N and Jun F: A
quantitative method to detect HBV cccDNA by chimeric primer and
real-time polymerase chain reaction. J Virol Methods. 112:45–52.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang W, Ke W, Wu SS, et al: An
adenovirus-delivered peptide aptamer C1-1 targeting the core
protein of hepatitis B virus inhibits viral DNA replication and
production in vitro and in vivo. Peptides. 30:1816–1821. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Calvert J and Summers J: Two regions of an
avian hepadnavirus RNA pregenome are required in cis for
encapsidation. J Virol. 68:2084–2090. 1994.PubMed/NCBI
|
|
22
|
Lu B, Wang J, Zhang J, et al: Screening
and verification of ssDNA aptamers targeting human hepatocellular
carcinoma. Acta Biochim Biophys Sin (Shanghai). 46:128–135. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Devi U and Locarnini S: Hepatitis B
antivirals and resistance. Curr Opin Virol. 3:495–500. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ferapontova EE and Gothelf KV: Recent
advances in electrochemical aptamer-based sensors. Curr Org Chem.
15:498–505. 2011. View Article : Google Scholar
|
|
25
|
Chu TC, Marks JW III, Lavery LA, et al:
Aptamer: toxin conjugates that specifically target prostate tumor
cells. Cancer Res. 66:5989–5992. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang JC, Dhason MS and Zlotnick A:
Structural organization of pregenomic RNA and the carboxy-terminal
domain of the capsid protein of hepatitis B virus. PLoS Pathog.
8:e10029192012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Vanlandschoot P, Van Houtte F, Serruys B
and Leroux-Roels G: The arginine-rich carboxy-terminal domain of
the hepatitis B virus core protein mediates attachment of
nucleocapsids to cell-surface-expressed heparan sulfate. J Gen
Virol. 86:75–84. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jammeh S, Tavner F, Watson R, Thomas HC
and Karayiannis P: Effect of basal core promoter and pre-core
mutations on hepatitis B virus replication. J Gen Virol.
89:901–909. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Guo H, Jiang D, Zhou T, Cuconati A, Block
TM and Guo JT: Characterization of the intracellular deproteinized
relaxed circular DNA of hepatitis B virus: an intermediate of
covalently closed circular DNA formation. J Virol. 81:12472–12484.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schmitz A, Schwarz A, Foss M, et al:
Nucleoporin 153 arrests the nuclear import of hepatitis B virus
capsids in the nuclear basket. PLoS Pathog. 6:e10007412010.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Masri HP and Cornelissen CN: Specific
ligand binding attributable to individual epitopes of gonococcal
transferrin binding protein A. Infect Immun. 70:732–740. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yamada N and Tanoue E: Detection and
partial characterization of dissolved glycoproteins in oceanic
waters. Limnol Oceanogr. 48:1037–1048. 2003. View Article : Google Scholar
|
|
33
|
Tomai E, Butz K, Lohrey C, von Weizsacker
F, Zentgraf H and Hoppe-Seyler F: Peptide aptamer-mediated
inhibition of target proteins by sequestration into aggresomes. J
Biol Chem. 281:21345–21352. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Butz K, Denk C, Fitscher B, et al: Peptide
aptamers targeting the hepatitis B virus core protein: a new class
of molecules with antiviral activity. Oncogene. 20:6579–6586. 2001.
View Article : Google Scholar : PubMed/NCBI
|