Introduction
Reactive oxygen species (ROS), including superoxide
anion radical (O2•−), hydroxyl radical (•OH)
and hydrogen peroxide (H2O2) have been
implicated in the development of a variety of diseases, such as
carcinogenesis, inflammation, aging and atherosclerosis. The
accumulation of ROS induces lipid peroxidation, the inactivation of
proteins and DNA damage in cells (1,2).
Protective enzymatic and non-enzymatic antioxidant defense
mechanisms reduce oxidative stress by scavenging ROS. To protect
themselves against toxic free radicals and ROS, cells have
developed a variety of antioxidant defenses (3). These include antioxidant enzymes,
such as superoxide dismutase (SOD), which catalyzes the dismutation
of superoxide anions to hydrogen peroxide; catalase (CAT), which
converts H2O2 into molecular oxygen and
water; glutathione peroxidase (GPx), which catalyzes the
degradation of H2O2 and hydroperoxides. On
the non-enzymatic level, certain vitamins and other antioxidant
compounds scavenge free radicals and delay the oxidation of
molecules (4,5). Phytochemicals provide further
protection against oxidative damage from free radicals. A large
number of studies have indicated that phytochemicals present in
fruits, vegetables and herbs, exert their antioxidant effects
against oxidative stress through the induction or activation of
these endogenous antioxidant enzymes (6–9).
In addition, increased ROS production not only directly damages
cells by oxidizing macromolecules, such as DNA and lipids, but also
indirectly by triggering mitogen-activated protein kinase (MAPK)
signaling pathways (10). MAPKs,
such as extracellular signal-regulated kinase (ERK), p38 and c-Jun
N-terminal kinase (JNK) are involved in crucial signaling pathways
in cell proliferation, differentiation and cell death in response
to various signals produced by growth factors, hormones and
cytokines, as well as genotoxic and oxidative stressors (11). Studies have demonstrated that MAPK
signaling pathways can also be modulated by the production of ROS
and antioxidant enzymatic activity and expression (12,13).
Ginseng (Panax ginseng C.A. Meyer) is
considered as one of the most popular medicinal herbs, and has well
known pharmacological activities which include anticancer (14), anti-aging (15), anti-diabetic (16), anti-stress (17) and neuroprotective effects
(18). Black ginseng is a
processed ginseng produced by a nine-time steaming at approximately
85°C and a nine-time drying process (repetitive steaming and
drying) using fresh ginseng, at which point the ginseng becomes
black in color (19). Fermented
black ginseng (FBG) is processed further by incubating black
ginseng with Saccharomyces cerevisiae for 24 h to produce
more active ginsenosides. Black ginseng has been shown to enhance
biological activities possibly due to the enrichment of the
bioactive chemical constituents during the heat and drying
processing stage (20). However,
to the best of our knowledge, studies on the antioxidant properties
and the underlying molecular mechanisms of FBG are limited. In the
present study, we investigated the antioxidant defense properties
of FBG at the enzymatic and cellular levels and its ability to
inhibit ROS production; we demonstrate that FBG induces both the
activity and the expression of antioxidant enzymes and modulates
upstream protein kinases, including MAPKs.
Materials and methods
Reagents
FBG was obtained from Ginseng-By-Pharm Co. (Wonju,
Korea). The main composition of FBG was determined using the liquid
chromatography-mass spectrometry (LC/MS) method (Joongbu
University, Geumsan, Korea) and is shown in Table I. KCN (potassium cyanide, 60178),
H2O2, sulfanilamide (S9251),
N-(1-naphthyl)ethylenediamine dihydrochloride (P7626), nitroblue
tetrazolium salt, xanthine, copper chloride, glutathione, xanthine
oxidase from bovine milk (0.1–0.4 U/mg of protein), glutathione
reductase from baker’s yeast (Saccharomyces cerevisiae;
100–300 U/mg of protein) and
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium, inner salt
(MTT) were purchased from Sigma-Aldrich (St. Louis, MO, USA).
2′,7′-Dichlorohydrofluorescein diacetate (DCF-DA) was purchased
from Molecular Probes, Inc. (Eugene, OR, USA). Antibodies against
CAT (14097), phosphorylated ERK (9101), phosphorylated JNK (9251),
phosphorylated p38 (9211) and horseradish peroxidase-conjugated
anti-rabbit IgG (7074) were purchased from Cell Signaling
Technology Inc. (Beverly, MA, USA). Antibodies against SOD
(sc-18504), GPx (sc-30147), β-actin (sc-1616) and horseradish
peroxidase-conjugated anti-goat IgG (sc-2350) were purchased from
Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). All other
reagents were of the highest quality generally available.
 | Table ISaponin and main ginsenosides found
in fermented black ginseng. |
Table I
Saponin and main ginsenosides found
in fermented black ginseng.
| Ginsenosides | Concentration
(μg/ml) |
|---|
| Crude saponin | 1,440 |
| Ginsenoside
Rg2 | 2.86 |
| Ginsenoside
Rg3 | 24.52 |
| Ginsenoside
Rh1 | 12.62 |
| Ginsenoside
Rh2 | 0.63 |
| Ginsenoside Rf | 1.32 |
Cell culture
HepG2 human hepatocellular carcinoma cells were
obtained from the American Type Culture Collection (ATCC; Manassas,
VA, USA). The cells were maintained in Dulbecco’s minimum essential
medium with 10% fetal bovine serum (FBS), 100 U/ml penicillin, 100
μg/ml streptomycin (all from HyClone, Logan, UT, USA), 1% essential
amino acids and 1% glutamax (both from Gibco, Grand Island, NY,
USA) at 37°C in a humidifying incubator containing 5%
CO2. For subculture, the HepG2 cells were harvested at
80–90% confluence.
Cell viability assay
MTT assay was used to measure cell viability. The
HepG2 cells were seeded in 24-well plates at a rate of
1×105 cells/well. After 24 h of incubation, the cells
were treated with distilled water (DW; control) or different doses
of FBG for 24 h. Subsequently, 50 μl of 1 mg/ml MTT were added and
the plates were incubated for 4 h. Following incubation at 37°C for
4 h, the MTT medium was removed by aspiration and 200 μl of
dimethyl sulfoxide (DMSO) were added to each well. After reacting
for 10 min at room temperature, formazan production was detected by
the measurement of the optical density (OD) at 570 nm using a
PowerWave XS microplate reader (BioTek Instruments, Winooski, VA,
USA). The data are expressed as the percentage cell viability
compared to the vehicle-treated control.
ROS formation assay
Intracellular ROS levels were determined by the
DCF-DA assay. Briefly, the cells were seeded in 96-well dark plate
at a rate of 1×104 cells/well and pre-incubated with 20
μM DCF-DA (dissolved in DMSO) for 1 h at 37°C in the dark. After
washing out the excess probe using 1× ice-cold phosphate-buffered
saline (PBS), the cells were treated with the vehicle (DW for
control), vitamin C (positive control), or FBG in the presence or
absence of 1 mM H2O2 for 12 h and then washed
twice with 1× ice-cold PBS. Fluorescence was detected by excitation
at 485 nm and emission at 535 nm using a fluorescence
multi-detection reader (BioTek Instruments). Vitamin C (100 μg/ml)
was used as positive control for current study. Vitamin C is well
known as a strong antioxidant and is widely used as a positive
control in antioxidant studies.
Assays for antioxidant enzymes
The cells were seeded in 6-well plates at a rate of
2×105 cells/well. After 24 h of incubation, the cells
were treated with DW (control), vitamin C (positive control), or
different doses of FBG for 24 h. For the CAT and GPx enzyme assays,
the cells were homogenized with 1 ml of 50 mM potassium phosphate
buffer (pH 7.0), and then centrifuged at 12,000 rpm for 20 min at
4°C. For the manganese-superoxide dismutase (Mn-SOD) assay, the
cells were homogenized with 1 ml of 65 mM phosphate buffer (pH
7.8), and then centrifuged at 12,000 rpm for 20 min at 4°C. The
cell lysis supernatant was analyzed to determine CAT and GPx
activity, while the cell pellet was used to detect Mn-SOD activity.
The protein concentration was determined using the BCA protein
assay (Pierce Biotechnology, Rockford, IL, USA) according to the
manufacturer’s instructions. The results are expressed as the
enzyme activity per milligram protein compared with the
corresponding control cultures.
Mn-SOD activity
The activity of Mn-SOD was measured according to the
method described in the study by Oyanagui (21). The remaining pellet (i.e., the
mitochondrial fraction) was dissolved in 0.1% Triton X-100 and used
for the determination of Mn-SOD activity. For the Mn-SOD activity
assay, 60 μl of 4 mM of KCN solution were added to the assay
mixture to inhibit copper- and zinc-containing superoxide dismutase
(Cu/Zn-SOD). The samples were pre-incubated with 15 μl of 75 mM
Na-xanthine and 15 μl of 10 mM hydroxylamine hydrochloride at 37°C
for 10 min. Subsequently, 0.1 units of xanthine oxidase were added
and the samples were incubated at 37°C for an additional 20 min.
The reaction was terminated by the addition of 1% sulphanilamide
and 0.02% ethylenediamine dihydrochloride. After standing at room
temperature for 20 min, the absorbance of the final mixture was
measured at 450 nm. Enzyme activity was expressed as units per
milligram protein.
CAT activity
The activity of CAT was measured as previously
described (22). The reaction
mixture contained 12 μl of 3% (vol/vol) H2O2
and 20 μg of cell lysates in 50 mM potassium phosphate buffer (pH
7.0) at a final volume of 1.0 ml. The samples were incubated for 5
min at 37°C and the absorbance of the samples was monitored for 5
min at 240 nm. The change in absorbance is proportional to the
breakdown of H2O2.
GPx activity
The activity of GPx was measured by as previously
described (23). A total of 20 μg
of supernatant, containing cytosolic fraction, was incubated with 1
mM EDTA, 1 mM sodium azide (NaN3), 5 mM GSH, 1 mM NADPH
and 1 unit glutathione reductase at room temperature for 5 min. The
reaction was initiated by the addition of 25 μl of 2.5 mM
H2O2. GPx activity was measured as the rate
of NADPH oxidation at 340 nm.
Western blot analysis
The cells were pre-treated with the vehicle (DW for
control), vitamin C (positive control), or FBG (10, 25 or 50 μg/ml)
for 1 h and then challenged with H2O2 for
another 1 h. Subsequently, the cells were washed 3 times with
ice-cold PBS (pH 7.4) and harvested with 200 μl of whole cell lysis
buffer (pH 7.4) containing 10 mM Tris-HCl, 50 mM sodium chloride,
30 mM sodium pyrophosphate, 50 mM sodium fluoride, 100 μM sodium
orthovanadate, 2 mM iodoacetic acid, 5 mM ZnCl2, 1 mM
phenylmethylsulfonyl fluoride and 0.5% Triton X-100. The cell
lysates were vigorously vortexed, homogenized in a sonicator for 10
sec and left on ice for 1 h. The homogenates were centrifuged at
13,000 × g for 10 min at 4°C. The supernatants were collected and
equal amounts of total protein, as determined by BCA protein assay
(Pierce Biotechnology), were mixed with 2× loading buffer and
heated at 95°C for 5 min. An equal amount (30 μg) of protein from
each cell lysate was separated by 12% SDS-polyacrylamide gel
electrophoresis and transferred onto polyvinylidene difluoride
(PVDF) membranes (Millipore, Billerica, MA, USA). The membranes
were blocked with 5% non-fat dry milk in 1× PBST buffer (0.1%
Tween-20 in PBS) for 1 h at room temperature and then incubated
overnight with the appropriate primary antibody. Following
hybridization with the primary antibody, the membranes were washed
3 times with PBST, and then incubated with anti-rabbit and
anti-goat antibodies with horseradish peroxidase for 1 h at room
temperature and washed with PBST 3 times. Final detection was
performed with enhanced chemiluminescence (ECL™) western blotting
reagents (Santa Cruz Biotechnology, Inc.).
Statistical analysis
The data are expressed as the means ± standard
deviation (SD) values using Microsoft Excel. The values were
compared with those of the control using analysis of variance,
followed by Bonferroni’s test (GraphPad Prism software version
5.01; GraphPad Software, San Diego, CA, USA). The significance
level was defined at P-values <0.05.
Results
Effects of FBG on cell viability
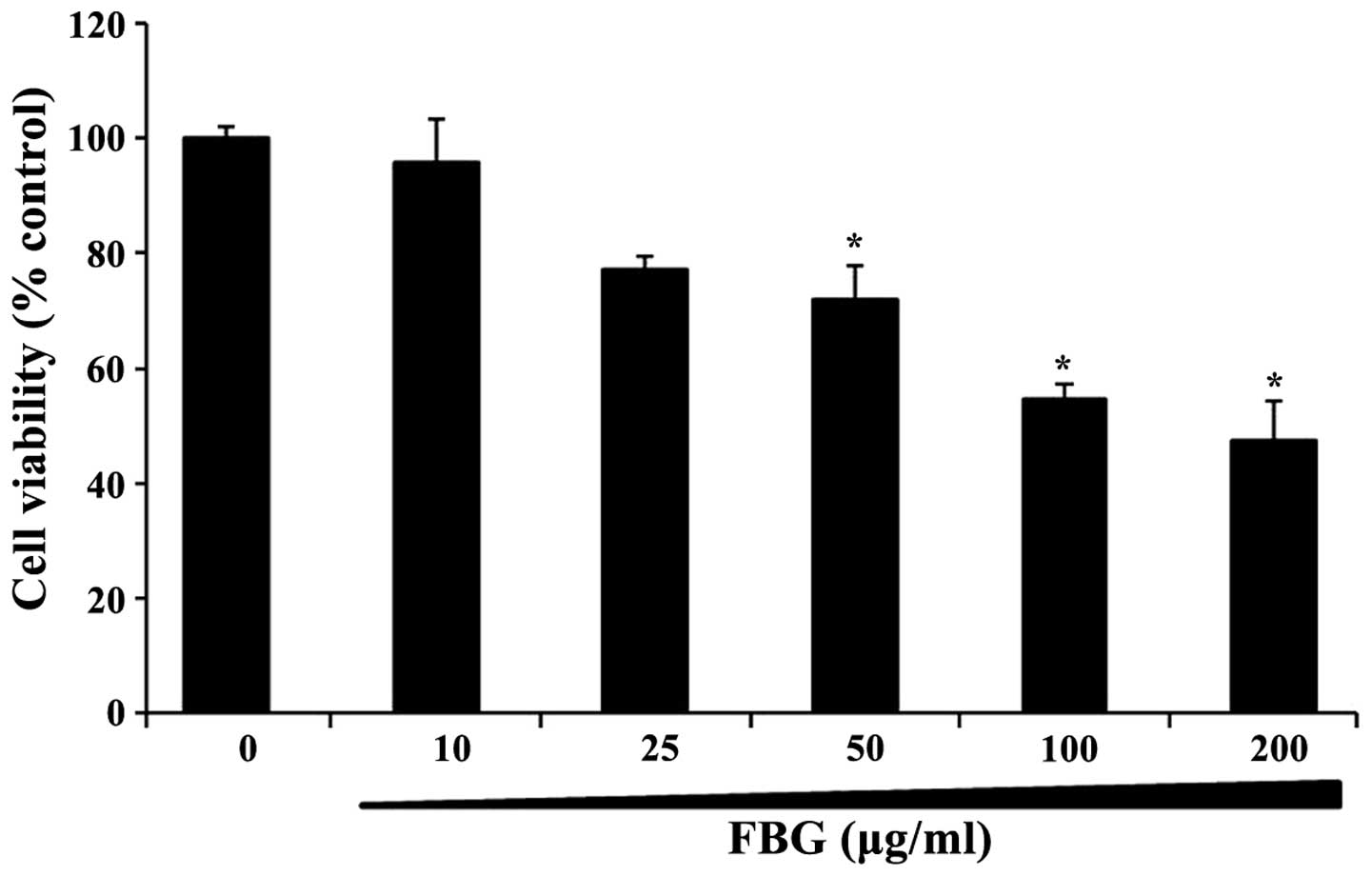
The cytotoxic effects of FBG on the HepG2 cells were
evaluated by MTT assay. The percentage of viable cells was
determined by MTT assay and compared to that of the control cells.
The cells were treated with FBG at the concentration range of 0–200
μg/ml for 24 h. Treatment with 10–200 μg/ml FBG inhibited cell
viability in a dose-dependent manner (Fig. 1). The survival rate of the cells
treated with 50 μg/ml of FBG was approximately 70% compared to that
of the control cells. Based on the cell viability data, subsequent
experiments were performed using concentrations of FBG below 50
μg/ml of FBG.
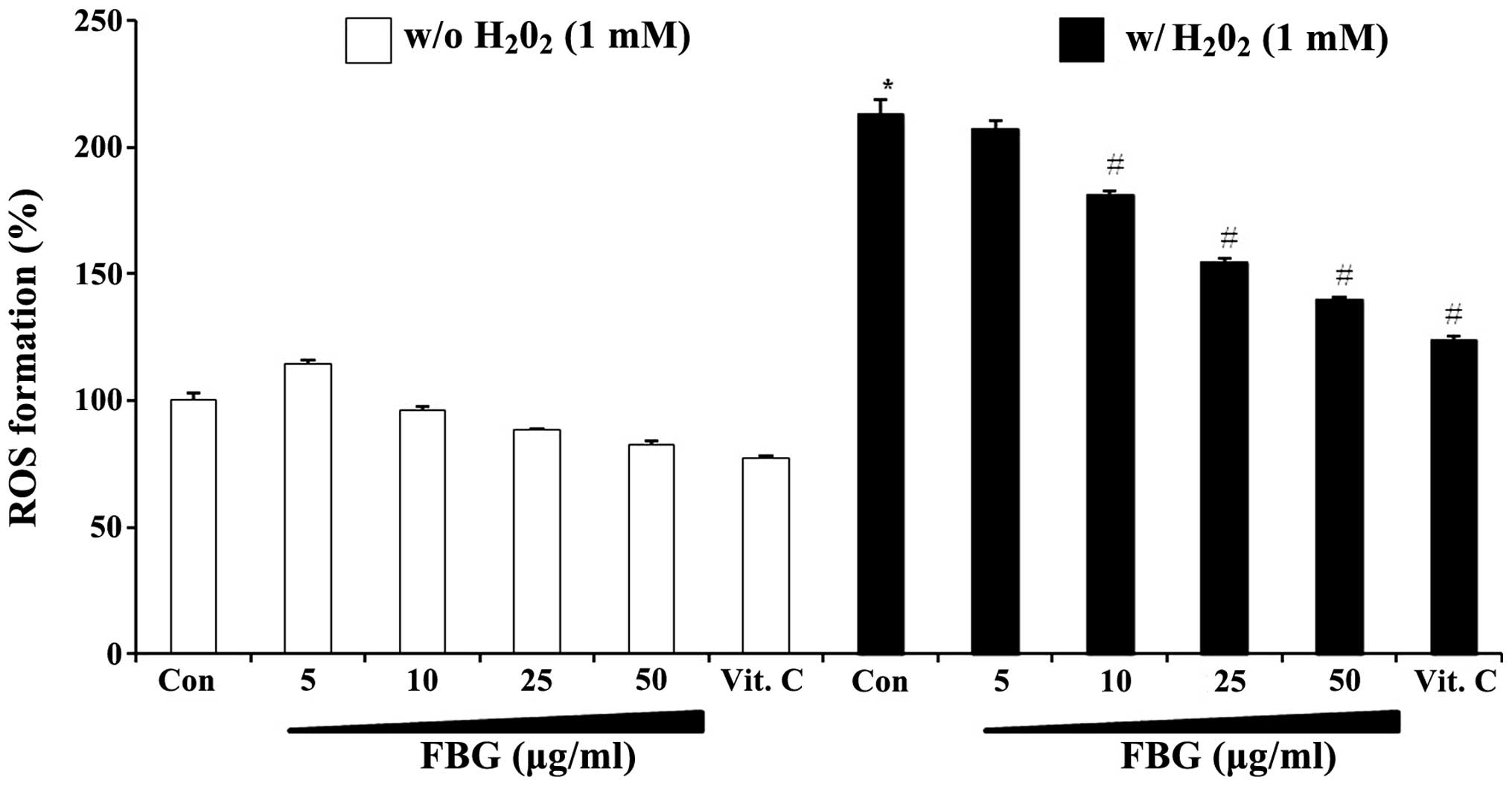
Effects of FBG on ROS production
We examined whether FBG exerts inhibitory effects on
the production of ROS using DCF-DA in
H2O2-treated HepG2 cells. When the cells were
treated with 1 mM H2O2, a >2.1-fold
increase in the generation of ROS compared to the vehicle-treated
controls was observed. Pre-treatment with FBG decreased the
H2O2-mediated production of ROS in a
dose-dependent manner and vitamin C (100 μg/ml) also markedly
decreased ROS formation in the presence and/or absence of
H2O2 in the HepG2 cells (Fig. 2). The dose-dependent effects of
FBG were observed even in the absence of
H2O2. These results suggest that FBG acts as
an antioxidant which can directly scavenge excessive ROS generation
in cells.
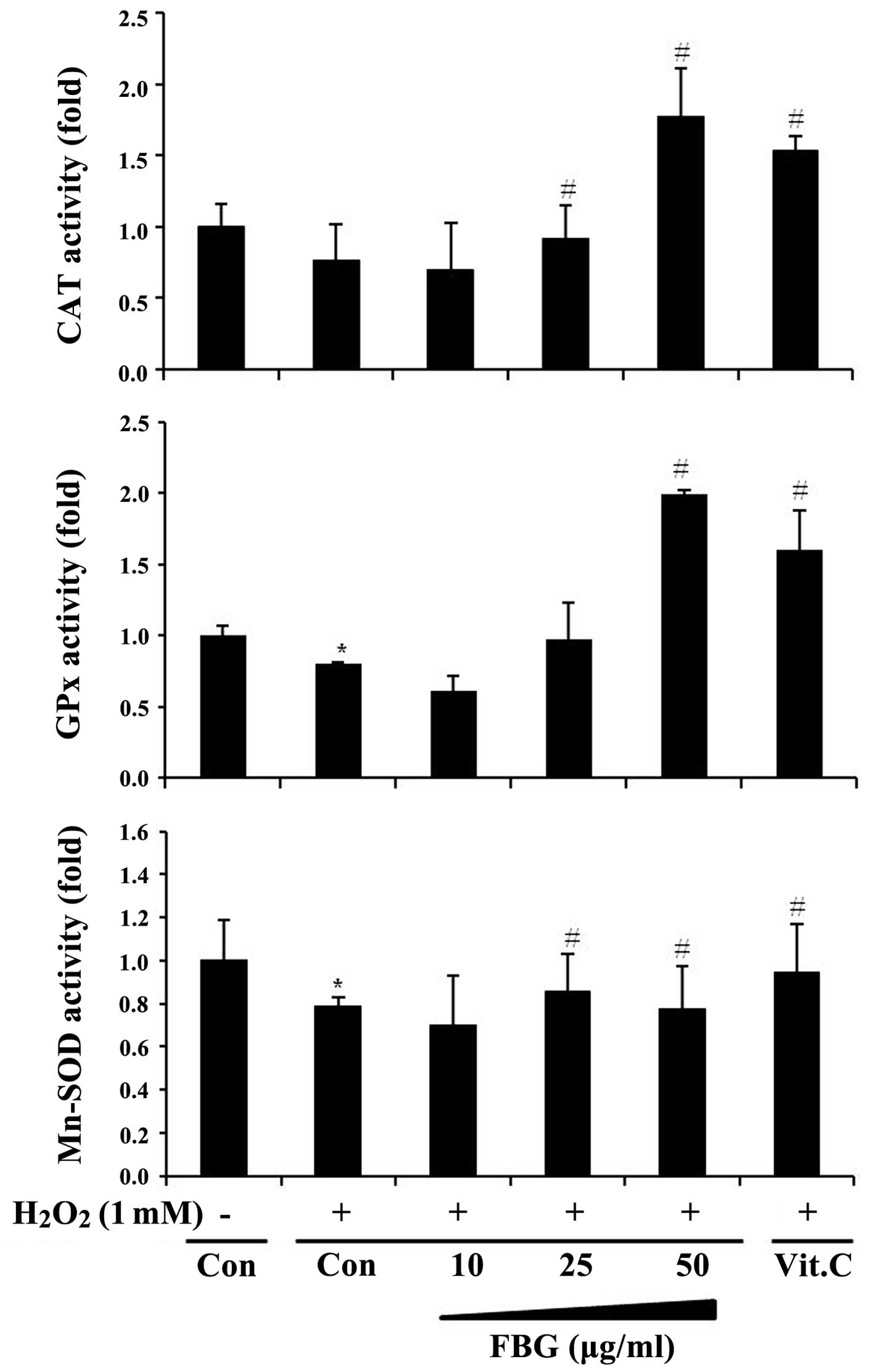
Effects of FBG on the activity of
antioxidant enzymes
In order to investigate whether the antioxidant
activity of FBG is mediated by its ability to increase the activity
of cellular antioxidant enzymes, we measured the activity of
antioxidant enzymes, including CAT, GPx and Mn-SOD in the
H2O2-treated HepG2 cells. When the cells were
treated with H2O2 (1 mM) alone, the activity
of CAT, GPx and SOD-2 significantly decreased below the basal level
of the vehicle-treated controls (Fig.
3). However, treatment with FBG increased the activity of these
enzymes; the increased activity levels of CAT and GPx following
treatment with 50 μg/ml FBG were much higher than the basal control
(DW) levels and were even higher than the levels observed following
treatment with vitamin C (positive control; 100 μg/ml), a well
known antioxidant.
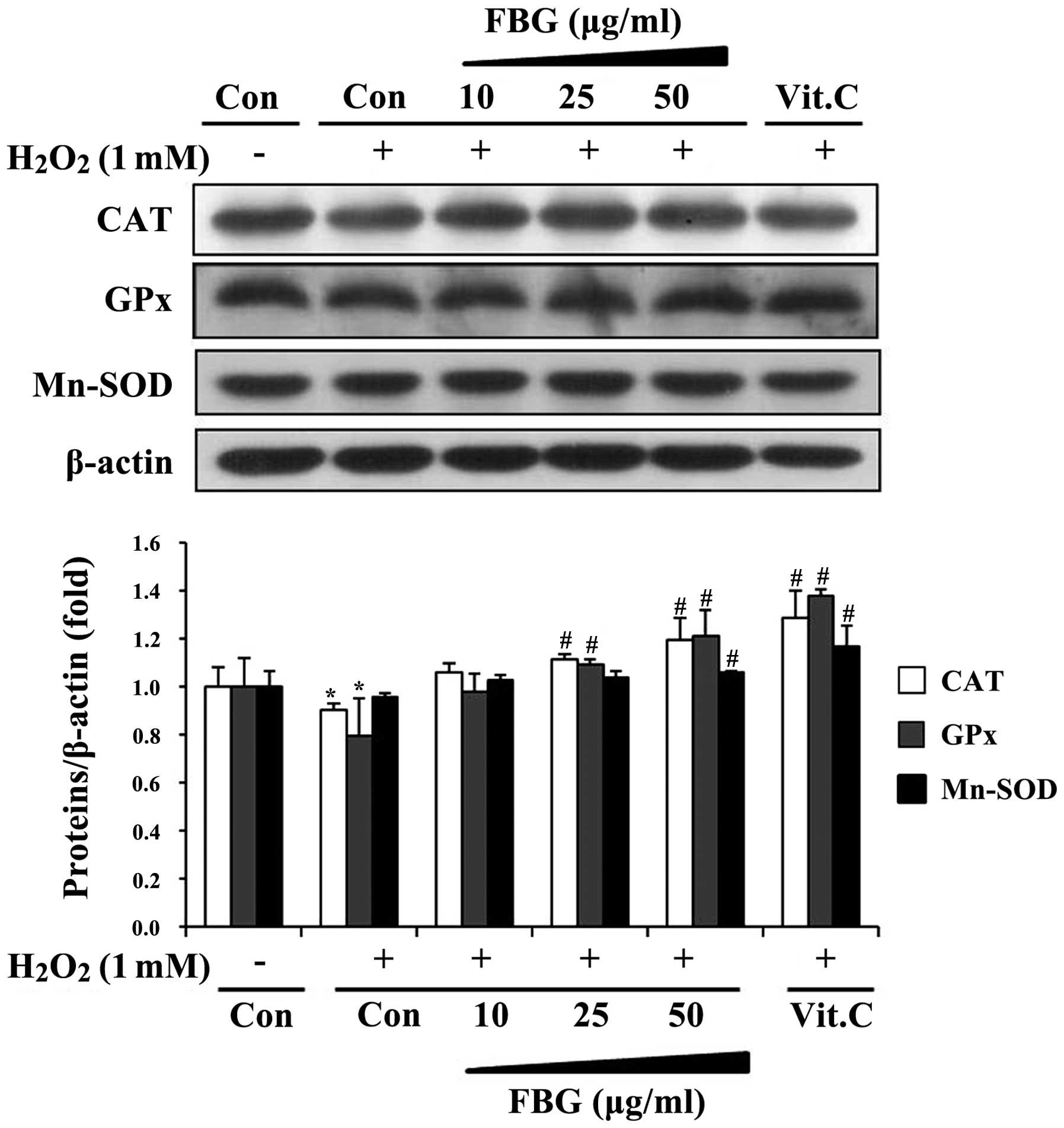
Effects of FBG on the expression of
antioxidant enzymes
In addition to its stimulatory effects on enzyme
activity, we determined the effects of FBG on the protein
expression of these antioxidant enzymes in HepG2 cells. When the
cells were treated with H2O2 alone, CAT, GPx
and Mn-SOD protein expression levels were diminished compared with
the vehicle-treated controls. However, treatment with FBG restored
and upregulated the protein expression of these enzymes in a
dose-dependent manner and vitamin C significantly induced the
protein level of antioxidant enzymes compared to the
H2O2-treated group (Fig. 4).
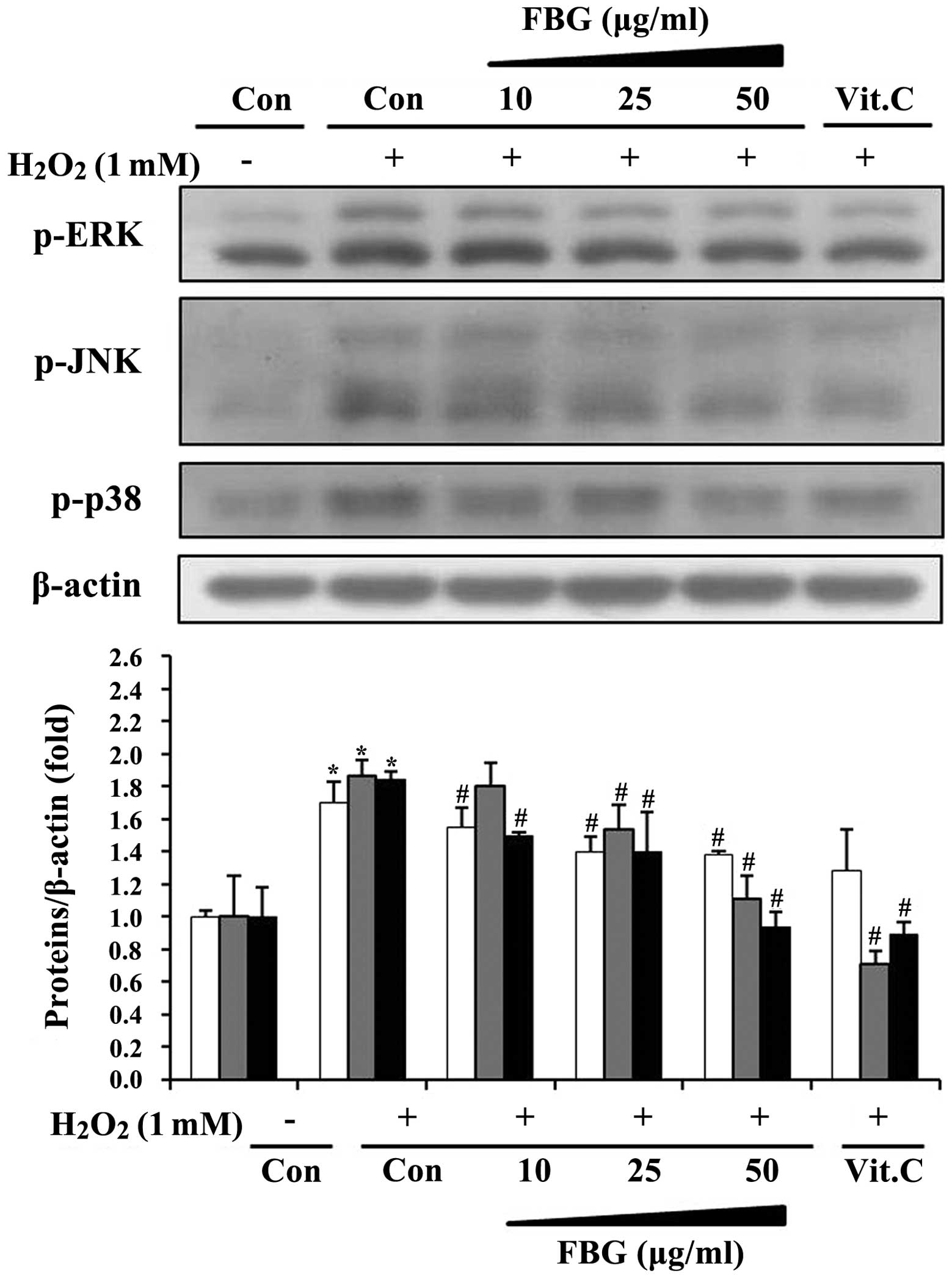
Effects of FBG on the phosphorylation of
MAPKs
In order to determine whether antioxidant enzyme
expression induced by FBG was associated with the MAPK pathway, we
examined the phosphorylation levels of MAPK subfamilies, such as
ERK, JNK and p38. The results revealed that
H2O2 stimulated the phosphorylation of all
MAPKs (Fig. 5). The
H2O2-stimulated phosphorylation of MAPKs was
decreased following treatment with FBG in a dose-dependent manner
and vitamin C inhibited the phosphorylation levels of MAPKs in the
presence of H2O2. This confirmed that
protective effects of vitamin C against H2O2
by the induction of antioxidant enzymes through the inhibition of
MAPK phosphorylation.
Discussion
The induction of antioxidant enzyme activity may be
considered as a frontline defense strategy to protect human health
against various oxidative stress-related diseases. Accordingly,
numerous bioactive plant materials have been investigated for their
antioxidant potential (4,6–8).
In the present study, to the best of our knowledge, we demonstrate
for the first time that FBG protects HepG2 cells against
H2O2-induced oxidative stress through the
regulation of ROS production and antioxidant enzymes, and signaling
pathways including MAPKs.
ROS are known to play a central role in mediating
various metabolic disorders related to several diseases. Thus,
inhibiting ROS production and enhancing the scavenging ability of
antioxidants may prove to be a useful strategy in the treatment of
diseases related to oxidative stress (24). ROS-induced oxidative DNA damage
has been implicated in mutagenesis and carcinogenesis and has
attracted extensive attention in recent years. In addition,
H2O2 is a major component of ROS produced
intracellularly during a number of physiological and pathological
processes, and causes oxidative damage (25,26). For this reason,
H2O2 has often been used as an experimental
model to investigate the mechanisms of cell injury induced by
oxidative stress (27–29). In the present study, when HepG2
cells pre-treated with FBG were challenged with
H2O2, ROS formation decreased. A previous
study demonstrated that Korean red ginseng extract exerted
antioxidant and chemopreventive effects by decreasing ROS
production in HepG2 cells treated with arachidonic acid and iron
(30). These results suggest that
ginsengs may have an antioxidant capacity by directly scavenging
radicals. Maintaining the balance between free radicals (and/or
ROS) and antioxidants is an essential part of biological
homeostasis (31).
Antioxidant enzymes, including SOD, CAT and GPx are
regarded as the firstline of the antioxidant defense system against
ROS generation during oxidative stress. Recently, white ginseng has
been reported to prevent oxidative stress by enhancing the
intracellular activity of antioxidant enzymes and decreasing ROS
formation (32). In addition, red
ginseng has been reported to exhibit a variety of antioxidant and
hepatoprotective effects on ethanol-induced oxidative injury in rat
liver and TIB-73 cells (33,34). Jun and Chang (35), reported that red ginseng extract
increased SOD, CAT and GPx activity after ICR male mice were
γ-irradiated. Furthermore, a ginseng extract has been shown to
induce hepatic SOD, CAT and GPx activity in Sprague-Dawley rats
(36). Our results are in
accordance with those of these studies, suggesting that FBG may
prove useful against oxidate stress by reducing ROS levels and
increasing antioxidant enzymes activity and expression. According
to previous studies, a variety of ginsengs has shown antioxidant
properties in maintaining cellular function against free radicals
in vivo, as well as in vitro (4,7,37).
In addition, the activity of intracellular antioxidant enzymes,
such as SOD, CAT and GPx plays an important role in protecting
cells against oxidative stress. Since the changes occurring in the
activities of these enzymes can be considered a biomarker of
antioxidant response under conditions of oxidative stress, the
increased activity of these enzymes in FBG-treated HepG2 cells
strongly suggests that FBG has antioxidant properties that function
by simulating the activity of antioxidant enzymes in addition to
directly scavenging ROS/free radicals. Lee et al (38), revealed that black ginseng has a
protective effect on ethanol-induced teratogenesis through the
augmentation of antioxidant activity in embryos.
Heat processing has been reported to increase the
free radical scavenging activity of ginseng and stimulate the
protective effects of ginseng against oxidative damage caused by
oxidative stress (20,39). Black ginseng is known to contain
different ginsenosides (Rg3, Rg4, Rg6, Rk3, Rs3 and Rs4) which are
not present in white ginseng (40), and exhibits more potent
pharmacological activities than white ginseng and red ginseng
(41,42). Black ginseng is prepared by
steaming at 85°C for 8 h and then drying until the water content
decreases below 20%. This steaming and drying process is repeated 9
times. This process makes white ginseng black, and from this point
on it is known as ‘black ginseng’. For the preparation of FBG,
black ginseng is grinded and extracted with distilled water at 80°C
for 72 h. Subsequently, this water extract is fermented with
Saccharomyces cerevisiae at 35°C for 24 h.
The activity and expression of antioxidant enzymes
may be modulated by upstream protein MAPKs, such as JNK, ERK and
p38. The phosphorylation of these proteins has been shown to be
mediated through H2O2-induced oxidative
stress (11). Although FBG
inhibits the phosphorylation of MAPK, as observed in this study,
its effects on the downstream targets, such as antioxidant enzymes
may be selective, at least in the current model system. Dong et
al (30), indicated that red
ginseng extract attenuates oxidative stress by reducing ROS
formation through the LKB1-AMPK pathway in the HepG2 cell line, but
not through the MAPK signaling pathway. It has been reported that
the effects of dietary compounds on antioxidant enzyme expression
and activity are mediated by the modification of several different
signal transduction pathways (43). In addition, different agents can
play one or more roles at different targets, and the cellular
events may depend on the types and concentrations of the agents, as
well as on the cell or tissue types. For example, Fan et al
(44), revealed that ginseng
pectin exerts protective effects against H2O2
through the ERK1/2 and Akt pathway in U87 neuronal cells. In
addition, red ginseng and its primary ginsenosides inhibit
ethanol-induced oxidative injury by reducing ROS production and
lipid peroxidation through MAPK pathways in TIB-73 mouse
hepatocytes (34). To the best of
our knowledge, in this study, we report for the first time the
modulatory role of FBG on upstream MAPKs. However, further studies
are required using animals in order to determine the optimal dose,
duration and method of administration.
In conclusion, FBG has the ability to protect cells
against oxidative damage by scavenging ROS and inducing both the
activity and the expression of cellular antioxidant enzymes
possibly through the inhibition of MAPK signaling pathways.
Therefore, FBG may be a potential natural agent for cellular
defense, at least in liver cells. However, further in vivo
studies using FBG are warranted.
Acknowledgements
This study was supported by the research fund of
Dankook University in 2012.
References
|
1
|
Ray PD, Huang BW and Tsuji Y: Reactive
oxygen species (ROS) homeostasis and redox regulation in cellular
signaling. Cell Signal. 24:981–990. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Valko M, Leibfritz D, Moncol J, Cronin MT,
Mazur M and Telser J: Free radicals and antioxidants in normal
physiological functions and human disease. Int J Biochem Cell Biol.
39:44–84. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Matés JM: Effects of antioxidant enzymes
in the molecular control of reactive oxygen species toxicology.
Toxicology. 153:83–104. 2000.PubMed/NCBI
|
|
4
|
Bak MJ, Jun M and Jeong WS: Antioxidant
and hepatoprotective effects of the red ginseng essential oil in
H2O2-treated HepG2 cells and
CCl4-treated mice. Int J Mol Sci. 13:2314–2330. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schnabel D, Salas-Vidal E, Narváez V, et
al: Expression and regulation of antioxidant enzymes in the
developing limb support an function of ROS in interdigital cell
death. Dev Biol. 291:291–299. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Naik SR and Panda VS: Antioxidant and
hepatoprotective effects of Ginkgo biloba phytosomes in
carbon tetrachloride-induced liver injury in rodents. Liver Int.
27:393–399. 2007.
|
|
7
|
Jung CH, Seog HM, Choi IW, Choi HD and Cho
HY: Effects of wild ginseng (Panax ginseng C.A. Meyer)
leaves on lipid peroxidation levels and antixodant enzyme
activities in streptozotocin diabetic rats. J Ethnopharmacol.
98:245–250. 2005.
|
|
8
|
Bak MJ, Jeong JH, Kang HS, Jin KS, Jun M
and Jeong WS: Stimulation of activity and expression of antioxidant
enzymes by solvent fractions and isolated compound from Cedrela
sinensis leaves in HepG2 cells. J Med Food. 14:405–412. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Luna-Vázquez FJ, Ibarra-Alvarado C,
Rojas-Molina A, et al: Nutraceutical value of black cherry
Prunus serotina Ehrh. fruits: antioxidant and
antihypertensive properties. Molecules. 18:14597–14612. 2013.
|
|
10
|
Son Y, Cheong YK, Kim NH, Chung HT, Kang
DG and Pae HO: Mitogen-activated protein kinases and reactive
oxygen species: How can ROS activate MAPK pathways? J Signal
Transduct. 2011:7926392011.PubMed/NCBI
|
|
11
|
Chen C and Kong AN: Dietary
cancer-chemopreventive compounds: from signaling and gene
expression to pharmacological effects. Trends Pharmacol Sci.
26:318–326. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yu R, Jiao JJ, Duh JL, Gudehithlu K, Tan
TH and Kong AN: Activation of mitogen-activated protein kinases by
green tea polyphenols: potential signaling pathways in the
regulation of antioxidant-responsive element-mediated phase II
enzyme gene expression. Carcinogenesis. 18:451–456. 1997.
View Article : Google Scholar
|
|
13
|
Shin MH, Rhie GE, Kim YK, et al:
H2O2 accumulation by catalase reduction
changes MAP kinase signaling in aged human skin in vivo. J Invest
Dermatol. 125:221–229. 2005.
|
|
14
|
Kang JH, Song KH, Woo JK, et al:
Ginsenoside Rp1 from Panax ginseng exhibits anti-cancer
activity by down-regulation of the IGF-1R/Akt pathway in breast
cancer cells. Plant Foods Hum Nutr. 66:298–305. 2011.
|
|
15
|
Lee HS, Kim MR, Park Y, et al: Fermenting
red ginseng enhances its safety and efficacy as a novel skin care
anti-aging ingredient: in vitro and animal study. J Med Food.
15:1015–1023. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yuan HD, Quan HY, Jung MS, et al:
Anti-diabetic effect of pectinase-processed ginseng radix (GINST)
in high rat diet-fed ICR mice. J Ginseng Res. 35:308–314. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim Y, Choi EH, Doo M, et al: Anti-stress
effects of ginseng via down-regulation of tyrosine hydroxylase (TH)
and dopamine β-hydroxylase (DBH) gene expression in
immobilization-stressed rats and PC12 cells. Nutr Res Pract.
4:270–275. 2010.PubMed/NCBI
|
|
18
|
Demir I, Kiymaz N, Gudu BO, et al: Study
of the neuroprotective effect of ginseng on superoxide dismutase
(SOD) and glutathione peroxidase (GSH-Px) levels in experimental
diffuse head trauma. Acta Neurochir (Wien). 155:913–922. 2013.
View Article : Google Scholar
|
|
19
|
Lee MR, Yun BS, In OH and Sung CK:
Comparative study of korean white, red, and black ginseng extract
on cholinesterase inhibitory activity and cholinergic function. J
Ginseng Res. 35:421–428. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kang KS, Yamabe N, Kim HY, Okamoto T, Sei
Y and Yokozawa T: Increase in the free radical scavenging
activities of American ginseng by heat processing and its safety
evaluation. J Ethnopharmacol. 113:225–232. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Oyanagui Y: Reevaluation of assay methods
and establishment of kit for superoxide dismutase activity. Anal
Biochem. 142:290–296. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Carrillo MC, Kanai S, Nokubo M and Kitani
K: (−) deprenyl induces activities of both superoxide dismutase and
catalase but not of glutathione peroxidase in the striatum of young
male rats. Life Sci. 48:517–521. 1991.
|
|
23
|
Bogdanska JJ, Korneti P and Todorova B:
Erythrocyte superoxide dismutase, glutathione peroxidase and
catalase activities in healthy male subjects in Republic of
Macedonia. Bratisl Lek Listy. 104:108–114. 2003.PubMed/NCBI
|
|
24
|
Kirkinezos IG and Moraes CT: Reactive
oxygen species and mitochondrial diseases. Semin Cell Dev Biol.
12:449–457. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ziech D, Franco R, Pappa A and
Panayiotidis MI: Reactive oxygen species (ROS)-induced genetic and
epigenetic alterations in human carcinogenesis. Mutat Res.
711:167–173. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Panayiotidis M: Reactive oxygen species
(ROS) in multistage carcinogenesis. Cancer Lett. 266:3–5. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Campos J, Schmeda-Hirschmann G, Leiva E,
et al: Lemon grass (Cymbopogon citratus (D.C) Stapf)
polyphenols protect human umbilical vein endothelial cell (HUVECs)
from oxidative damage induced by high glucose, hydrogen peroxide
and oxidised low-density lipoprotein. Food Chem. 151:175–181.
2014.
|
|
28
|
Suchaoin W and Chanvorachote P: Caveolin-1
attenuates hydrogen peroxide-induced oxidative damage to lung
carcinoma cells. Anticancer Res. 32:483–490. 2012.PubMed/NCBI
|
|
29
|
Meehan WJ, Spencer JP, Rannels DE, Welch
DR, Knobbe ET and Ostrander GK: Hydrogen peroxide induces oxidative
DNA damage in rat type II pulmonary epithelial cells. Environ Mol
Mutagen. 33:273–278. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dong GZ, Jang EJ, Kang SH, et al: Red
ginseng abrogates oxidative stress via mitochondria protection
mediated by LKB1-AMPK pathway. BMC Complement Altern Med.
13:642013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jaruga P and Oliński R: Activity of
antioxidant enzymes in cancer diseases. Postepy Hig Med Dosw.
48:443–455. 1994.(In Polish).
|
|
32
|
Sohn SH, Kim SK, Kim YO, et al: A
comparison of antioxidant activity of Korean White and Red Ginsengs
on H2O2-induced oxidative stress in HepG2
hepatoma cells. J Ginseng Res. 37:442–450. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Seo SJ, Cho JY, Jeong YH and Choi YS:
Effect of Korean red ginseng extract on liver damage induced by
short-term and long-term ethanol treatment in rats. J Ginseng Res.
37:194–200. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Park HM, Kim SJ, Mun AR, et al: Korean red
ginseng and its primary ginsenosides inhibit ethanol-induced
oxidative injury by suppression of the MAPK pathway in TIB-73
cells. J Ethnopharmacol. 141:1071–1076. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jun C and Chang CC: The effect of red
ginseng extracts on the superoxide dismutase, peroxidase and
catalase activities in the liver of gamma ray irradiated mice.
Korean J Ginseng Sci. 17:29–34. 1993.
|
|
36
|
Lee DW, Shon HO, Lim HB and Lee YG:
Antioxidation of Panax ginseng. Korean J Ginseng Sci.
19:31–38. 1995.
|
|
37
|
Kim GN, Lee JS, Song JH, Oh CH, Kwon YI
and Jang HD: Heat processing decreases Amadori products and
increases total phenolic content and antioxidant activity of Korean
red ginseng. J Med Food. 13:1478–1484. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lee SR, Kim MR, Yon JM, et al: Black
ginseng inhibits ethanol-induced teratogenesis in cultured mouse
embryos through its effects on antioxidant activity. Toxicol in
Vitro. 23:47–52. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kang KS, Kim HY, Pyo JS and Yokozawa T:
Increase in the free radical scavenging activity of ginseng by
heat-processing. Biol Pharm Bull. 29:750–754. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sun BS, Gu LJ, Fang ZM, et al:
Simultaneous quantification of 19 ginsenosides in black ginseng
developed from Panax ginseng by HPLC-ELSD. J Pharm Biomed
Anal. 50:15–22. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lee MR, Kim BC, Kim R, et al: Anti-obesity
effects of black gineng extract in high fat diet-fed mice. J
Ginseng Res. 37:308–349. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Park HJ, Shim HS, Kim KS and Shim I: The
protective effect of black ginseng against transient focal
ischemia-induced neuronal damage in rats. Korean J Physiol
Pharmacol. 15:333–338. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ramiro-Puig E, Urpí-Sardà M, Pérez-Cano
FJ, et al: Cocoa-enriched diet enhances antioxidant enzyme activity
and modulates lymphocyte composition in thymus from young rats. J
Agric Food Chem. 55:6431–6438. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Fan Y, Sun C, Gao X, et al: Neuroprotetive
effects of ginseng pectin through the activation of ERK/MAPK and
Akt survival signaling pathways. Mol Med Rep. 5:1185–1190.
2012.PubMed/NCBI
|