Introduction
Red seaweeds have attracted increasing attention in
recent years through research aimed to develop new medicines and
healthy diets from bioactive compounds (1). Porphyra yezoensis (P.
yezoensis) is a critical alga that is mainly consumed in Korea,
China and Japan. This alga is an important source of
physiologically active substances, such as sulfated
polysaccharides, polyphenols and peptides, with biological effects
including antitumor (2),
anti-inflammatory (3),
antioxidant (4) and blood
pressure effects (5). The
majority of studies on the biological activities of P.
yezoensis have been conducted in vivo. In the present
study, the induction of proliferation was examined in intestinal
epithelial cells by a peptide from P. yezoensis.
Activation of the insulin-like growth factor-I
receptor (IGF-IR) via autocrine, paracrine and endocrine mechanisms
appears to play an important role in regulating cell growth,
proliferation and differentiation (6). The insulin receptor substrate (IRS)
family (IRS-1 to IRS-4) and Shc are the best characterized
substrates of IGF-IR. The IRS proteins are docking proteins that
potentially bind to IGF-IR and recruit several effector proteins
through Src homology 2 (SH2) domain interactions (7,8).
Shc is a substrate of tyrosine kinase receptors, non-receptor
kinases and certain phosphatases (9,10).
Upon stimulation with IGF-I, tyrosine phosphorylation sites in the
IRS proteins bind to phosphotyrosine-containing binding motifs
(YXXM) within SH2 domains in several downstream signaling
molecules, including growth factor receptor-bound protein 2,
SH2-containing protein-tyrosine phosphatase 2 and the p85
regulatory subunit of phosphatidylinositol 3-kinase (PI3K)
(11). Receptor activation leads
to activation of various signaling pathways, including
mitogen-activated protein kinase (MAPK) cascades (12). PI3K is a critical regulatory
protein involved in intracellular signal transduction processes and
controlling major cellular functions (13). Activated PI3K catalyzes the
phosphorylation of the membrane phospholipid phosphatidylinositol
4,5-bisphosphate to generate phosphatidylinositol
3,4,5-trisphosphate, thereby producing a lipid-binding site on the
cell membrane for the serine/threonine kinase Akt (14). Akt is activated by phosphorylation
at Thr308 and Ser473 residues by two
phosphoinositide-dependent protein kinases, PDK1 and PDK2 (15). Akt controls cell survival,
proliferation, growth and motility (16). In mammalian cells, MAPK cascades
constitute a large kinase network that regulates a variety of
biological processes, including cell growth, proliferation,
differentiation and inflammatory responses (17). The p42/p44 MAPK [extracellular
signal-regulated kinase (ERK)] signal transduction pathway is
activated by various mitogens. By contrast, c-Jun N-terminal kinase
(JNK) and p38 pathways are mainly activated by cellular stress and
inflammatory cytokines. Notably, ERK is associated with cell
proliferation and growth (18,19).
Activator protein-1 (AP-1) comprises homodimers and
heterodimers composed of basic-region leucine zipper proteins that
belong to the Jun (c-Jun, v-Jun and Jun D) and Fos (c-Fos, v-Fos
and Fos B) families, as well as the associated activating
transcription factors (ATF2, B-ATF and ATF3/LRF1) (20,21). AP-1 regulation is induced by
various stimuli, including growth factors, cytokines and
ultraviolet (UV) irradiation. In addition, three different types of
MAPKs (ERK, JNK and fos-regulating kinase) contribute to induction
of AP-1 activity in response to a diverse array of extracellular
stimuli (22). Jun-Fos
(heterodimeric) and Jun-Jun (homodimeric) complexes preferentially
bind to the 12-O-tetredecanoylphorbol-13-acetate-responsive
element (23). The AP-1 complex
mediates responses to cellular signals by binding to DNA and
inducing gene transcription changes leading to physiological
activity in the cell. AP-1 thereby regulates numerous cellular
processes, including cell proliferation, differentiation and stress
responses (24).
In the present study, a peptide from P.
yezoensis (known as PY-PE) is shown to have proliferative
effects on IEC-6 intestinal epithelial cells. The intracellular
mechanism of PY-PE was determined, focusing on the IGF-IR signaling
pathway, which is involved in the regulation of cellular
proliferation and differentiation.
Materials and methods
Preparation of PY-PE
PY-PE (A-L-E-G-G-K-S-S-G-G-G-E-A-T-R-D-P-E-P-T) was
synthesized by Peptron, Inc. (Daejeon, Korea). Purification of
PY-PE was performed using the Shimadzu Prominence HPLC apparatus
and a C18 column (Capcell Pak; Shiseido, Tokyo, Japan) in 0.1%
trifluoroacetic acid (TFA)/water and a gradient of 10–70%
acetonitrile (0% acetonitrile in 2 min, 0–30% acetonitrile in 10
min, 30–90% acetonitrile in 2 min) in 0.1% TFA, with a flow rate of
1 ml/min and UV detection at 220 nm, controlled using the software
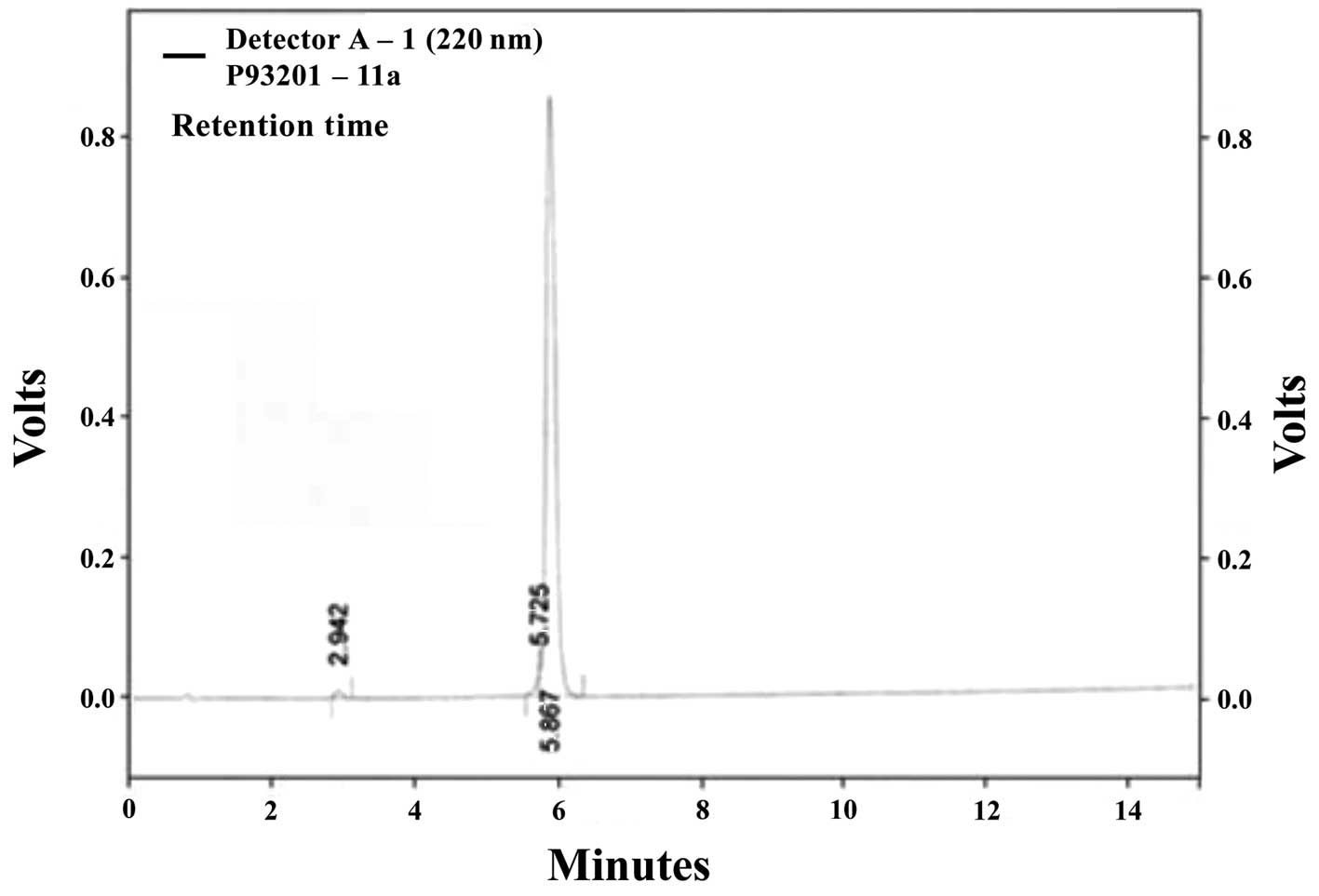
package Class-VP, 6.14 (Kyoto, Japan). The molecular weight of
PY-PE was determined to be 1,916 kDa (Fig. 1) using a mass spectrometer (HP
1100 Series LC/MSD; Agilent Technologies, Santa Clara, CA,
USA).
Cell culture
IEC-6 rat small intestinal epithelial cells (ATCC
CRL-1592) were obtained from the American Type Culture Collection
(Rockville, MD, USA). Cells were maintained in a humidified 5%
CO2 incubator at 37°C in Dulbecco’s modified Eagle’s
medium supplemented with 10% fetal bovine serum (HyClone, Inc.,
South Logan, UT, USA), 100 U/ml penicillin and 100 mg/ml
streptomycin. The medium was replaced every 2 days.
Cell proliferation assay
Cell proliferation was estimated using a CellTiter
96® aqueous non-radioactive cell proliferation assay
(Promega, Madison, WI, USA), which is based on the cleavage of
3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfonyl)-2H-tetrazolium
(MTS) into a formazan product that is soluble in tissue culture
medium. Cells were seeded in 96-well plates at 1×104
cells/well in 100 μl medium and attached for 24 h. Attached
cells were maintained in serum-free medium (SFM) for 4 h and were
subsequently treated with PY-PE (125–1,000 ng/ml) for 24 h. The
cells were incubated with 10 μl MTS solution for 30 min, and
the absorbance of each well was measured at 490 nm using a
SpectraMax 340PC microplate reader (Molecular Devices, Sunnyvale,
CA, USA).
Extraction of whole-cell protein
lysates
IEC-6 cells were plated in 100-mm dishes at a
density of 2×104 cells/ml and cultured to 60% confluence
at 37°C. The cells were subsequently incubated for 24 h in SFM
containing 0, 125, 250, 500 or 1,000 ng/ml PY-PE. Cells were washed
with phosphate-buffered saline and suspended on ice in lysis buffer
[50 mM Tris, 5 mM EDTA, 150 mM NaCl and 1% Triton X-100 (pH 7.5)]
containing protease inhibitors (1 mg/ml aprotinin, 1 mg/ml
leupeptin, 1 mg/ml pepstatin A, 200 mM
Na3VO4, 500 mM NaF and 100 mM PMSF). The
extracts were centrifuged at 14,000 rpm (10,770 × g) for 10 min,
and the supernatant was used for western blot analysis.
Extraction of nuclear lysates
Cells were treated and harvested as described above,
lysed with hypotonic lysis buffer [10 mM
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid)(HEPES) (pH
7.9), 10 mM KCl and 1.5 mM MgCl2], and incubated for 15
min at 4°C. Cells were lysed further by the addition of 2.5% NP-40
and incubated for 10 min at 4°C. After 10 min, nuclei were
collected by centrifugation at 5,000 rpm (1,593 × g) for 5 min at
4°C. Nuclear proteins were resuspended in extraction buffer [10 mM
HEPES (pH 7.9), 100 mM NaCl, 1.5 mM MgCl2, 0.1 mM EDTA
and 0.1 mM dithiothreitol] and incubated for 20 min at 4°C. The
extracts were centrifuged at 14,000 rpm (10,770 × g) for 10 min and
the supernatant was used for western blot analysis.
Western blot analysis
Protein extracts (30 μg) were separated by
7.5–12.5% sodium dodecyl sulfate-polyacrylamide gel electrophoresis
and transferred to polyvinylidene fluoride membranes (Millipore,
Billerica, MA, USA). Membranes were blocked with 1% bovine serum
albumin (BSA) in TBS-T [10 mM Tris-HCl, 150 mM NaCl (pH 7.5) and
0.1% Tween-20] and incubated overnight with the indicated primary
antibodies (diluted 1:500, 1:1,000 or 1:2,000; Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) in TBS-T containing 1%
BSA with gentle agitation at 4°C. The secondary antibody was a
horseradish peroxidase-conjugated goat anti-mouse or anti-rabbit
antibody (A90-116P; diluted 1:10,000; Bethyl Laboratories, Inc.,
Montgomery, TX, USA). The signals were detected using an enhanced
chemiluminescence Western blotting kit (Thermo Fisher Scientific,
Inc., Rockford, IL, USA).
Reverse transcription-polymerase chain
reaction (RT-PCR)
The mRNA expression levels of specific genes were
evaluated by RT-PCR. IEC-6 cells were seeded in 100-mm dishes at a
density of 2×104 cells/well and were cultured for 24 h,
after which the medium was replaced with SFM containing PY-PE (125,
250, 500 or 1,000 ng/ml) for 24 h. RNA was extracted from cells
using the TRIzol reagent (Invitrogen Life Technologies, Carlsbad,
CA, USA) and quantified using oligo(dT) primers (Intron
Biotechnology Co., Ltd., Seongnam, Korea). cDNA was reverse
transcribed from the RNA and subjected to amplification using a PCR
kit (dNTP mix, 10X Ex Taq Buffer and Ex Taq; Takara Bio, Inc.,
Shiga, Japan) with primers (Table
I) in 0.1% diethylpyrocarbonate water. PCR products were
resolved on 1% agarose gels. Gels were stained with 10 mg/ml
ethidium bromide to visualize the amplification products.
 | Table IOligonucleotide sequences of the
primers used in RT-PCR. |
Table I
Oligonucleotide sequences of the
primers used in RT-PCR.
| Gene name | Primer sequence
(5′-3′) |
|---|
| IGF-IR | F:
AAA-TGT-GCC-CGA-GCG-TGT-G
R: TGC-CCT-TGA-AGA-TGG-TGC-ATC |
| IRS-1 | F:
ACT-TGA-GCT-ATG-ACA-CGG-CT
R: GGT-TGG-AGC-AAC-TGG-ATG-AA |
| PI3K | F:
GCC-GAA-CAC-CTT-TTT-GAG-TC
R: AGG-AGC-GGT-ACA-GCA-AAG-AA |
| PDK1 | F:
AAG-GGT-ACG-GGC-CTC-TCA-AA
R: CCC-ACG-TGA-TGG-ACT-GAA-AGA |
| Akt | F:
CAA-CTT-CTC-TGR-GGC-GCA-GTG
R: GAC-AGG-TGG-AAG-AAC-AGC-TCG |
| c-Jun | F:
TCA-AAA-TGT-TTG-CAA-CTG-CTG-CG
R: ATG-ACT-GCA-AAG-ATG-GAA-ACG |
| c-Fos | F:
GGA-GAA-TCC-GAA-GGA-AGG
R: GCT-TGG-GCT-CAG-GGT-CAT-TG |
| GAPDH | F:
CAG-CCG-AGC-CAC-ATC-G
R: TGA-GGC-TGT-TGT-CAT-ACT-TCT-C |
Statistical analysis
Significant differences among multiple mean values
were assessed by analysis of variance using SPSS version 10.0 (SPSS
Inc., Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Proliferative effect of PY-PE in IEC-6
cells
Through mass spectrometry, a 1,916-kDa compound was
identified from P. yezoensis, which was designated as PY-PE
(Fig. 1). The proliferative
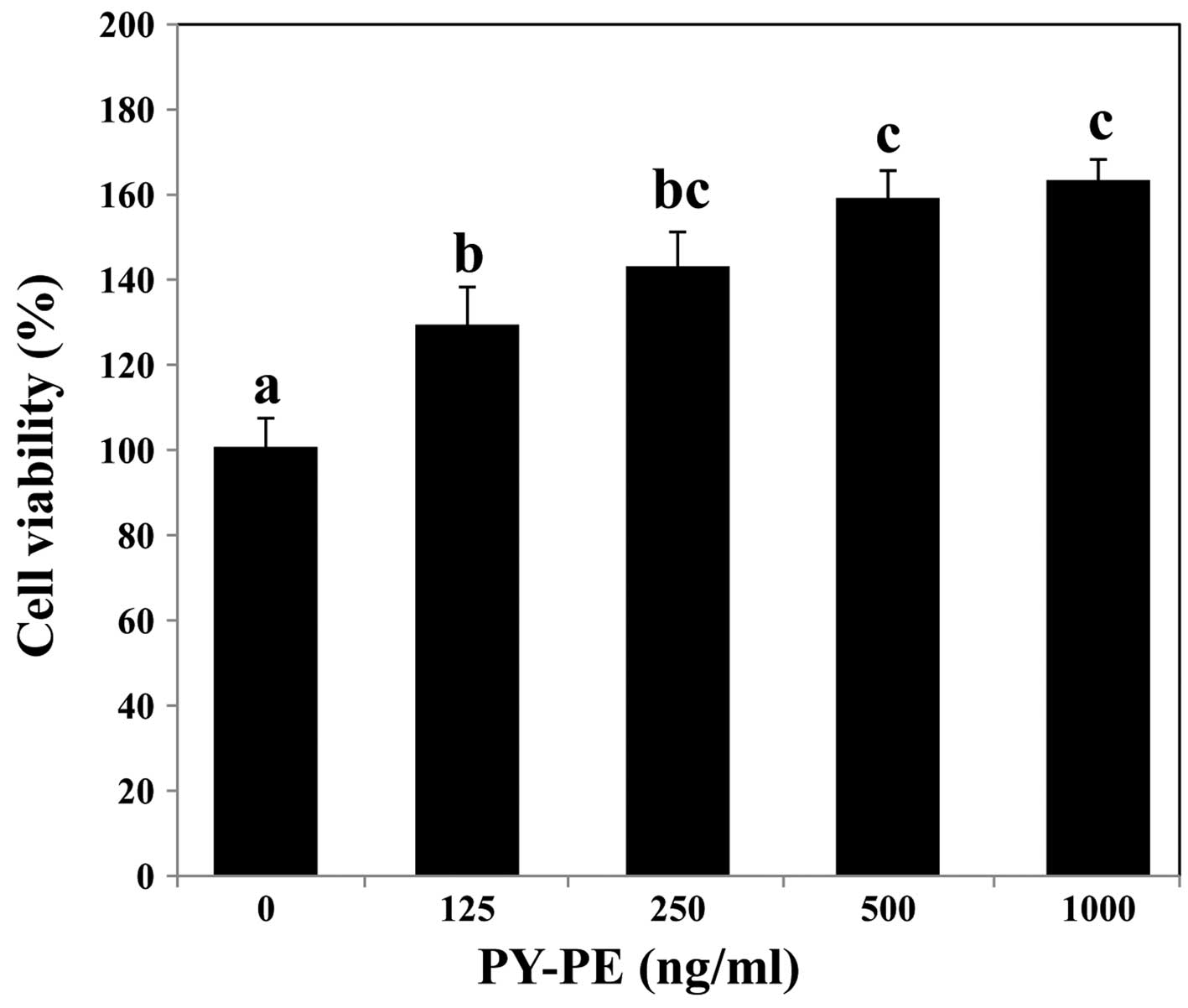
effect of PY-PE on IEC-6 cells was confirmed using MTS assay.
Treatment with PY-PE for 24 h increased cell viability in a
dose-dependent manner (Fig.
2).
Effect of PY-PE treatment on the
expression of IGF-IR-related proteins
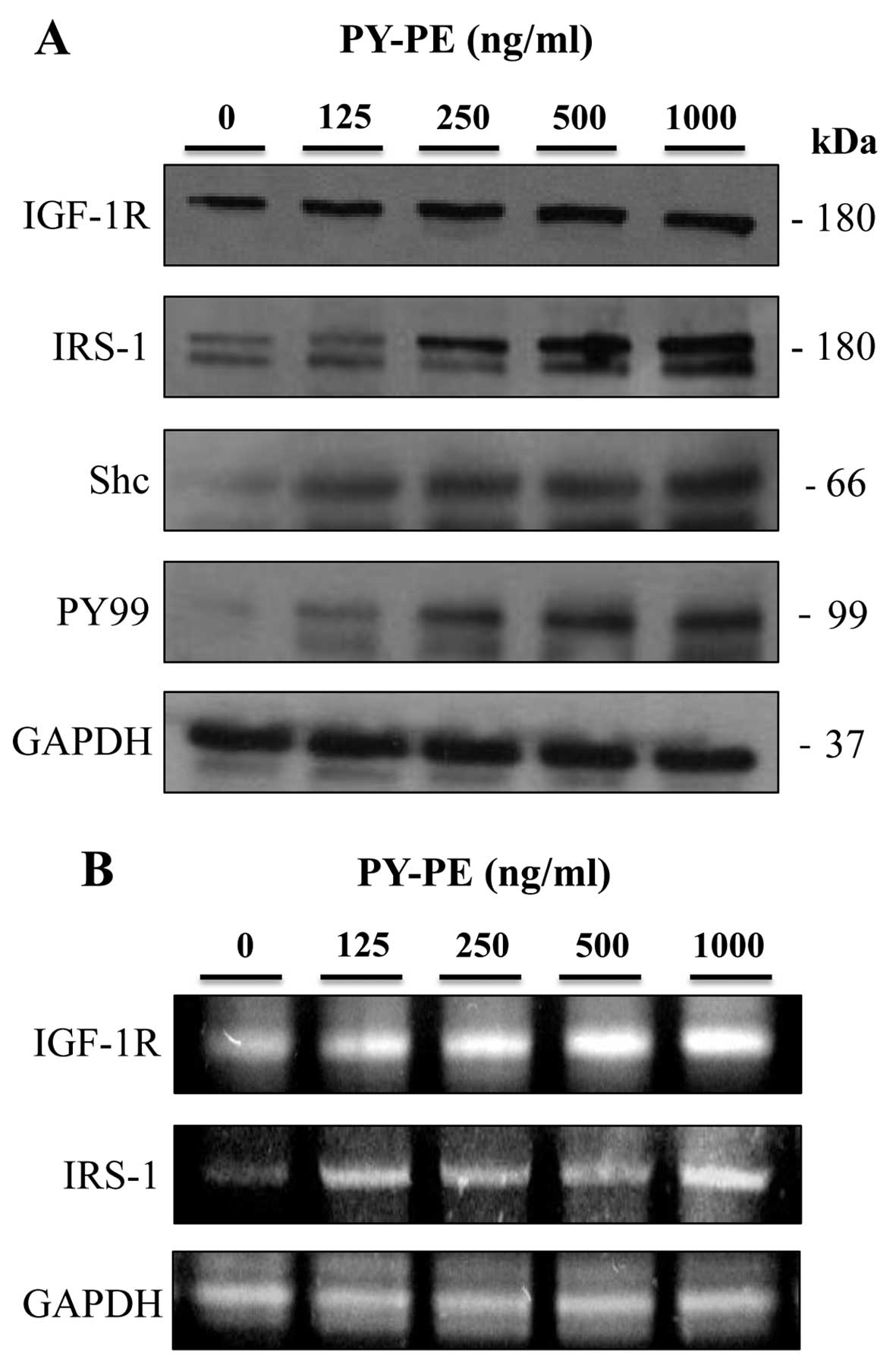
To confirm the mechanism of PY-PE-induced
proliferation in IEC-6 cells, the effects of PY-PE were examined on
IGF-IR signaling pathway-related proteins. The protein and mRNA
expression levels of IGF-IR, IRS-1, Shc and PY-99 in IEC-6 cells
treated with PY-PE (125, 250, 500 and 1,000 ng/ml) for 24 h were
determined by western blotting and RT-PCR. Treatment with PY-PE
dose-dependently upregulated the protein (Fig. 3A) and mRNA (Fig. 3B) expression levels of IGF-IR,
IRS-1, Shc and PY-99. IGF-IR stimulates the proliferation of
various cell types and inhibits apoptosis (25).
Effect of PY-PE treatment on the
expression of MAPK signaling pathway proteins
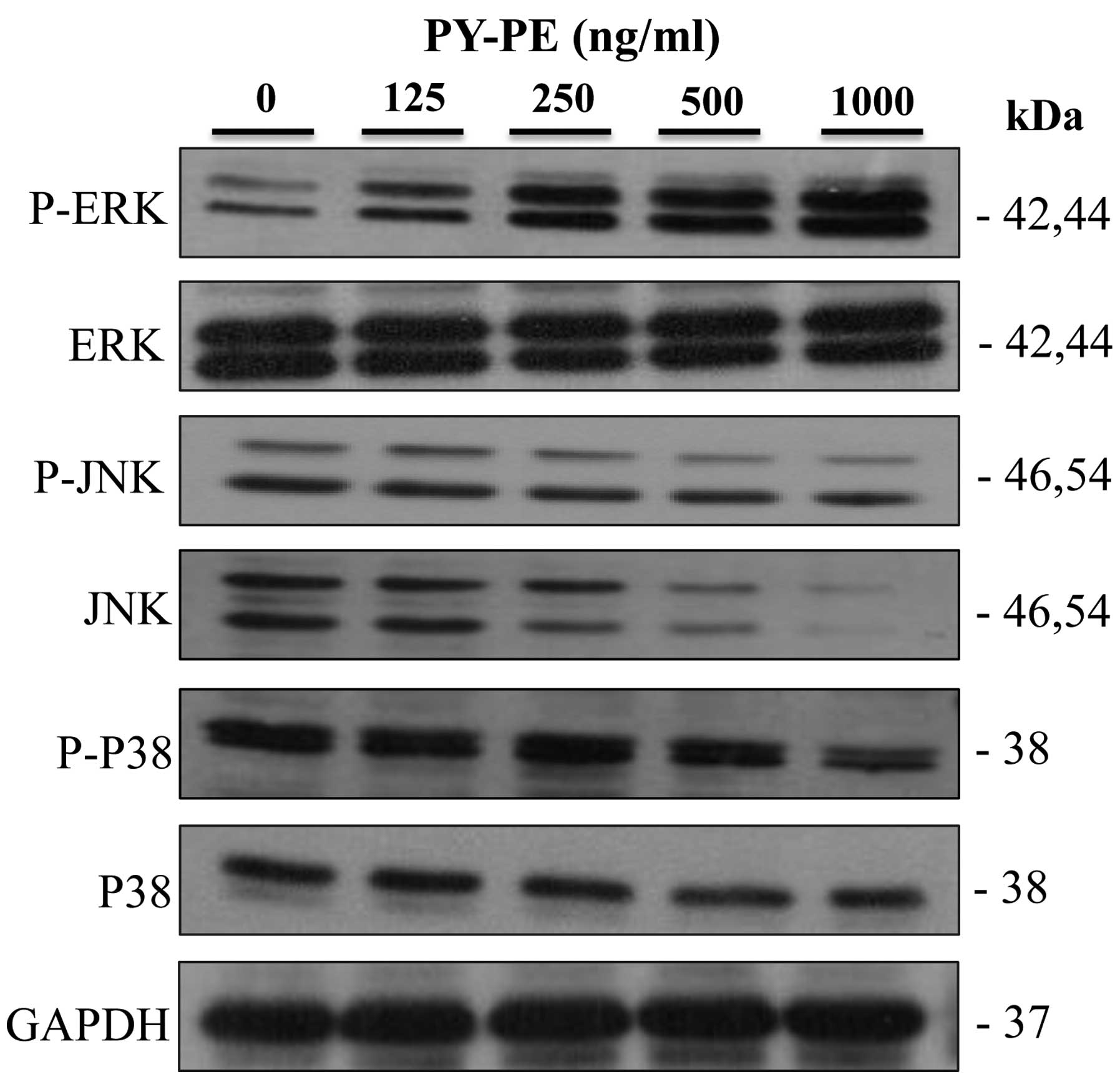
To further investigate the downstream signals
regulated by IGF-IR activation, the expression levels of the MAPK
family proteins (ERK1/2, JNK and P38) in IEC-6 cells treated with
PY-PE (125, 250, 500 and 1,000 ng/ml) for 24 h were determined by
western blot analysis. Treatment with PY-PE dose-dependently
increased the protein expression level of ERK1/2. By contrast,
PY-PE treatment inhibited the activation of JNK and p38 in
dose-dependent manners (Fig. 4).
These results suggest that ERK1/2 plays an important role in the
proliferation of IEC-6 cells.
Effect of PY-PE treatment on the
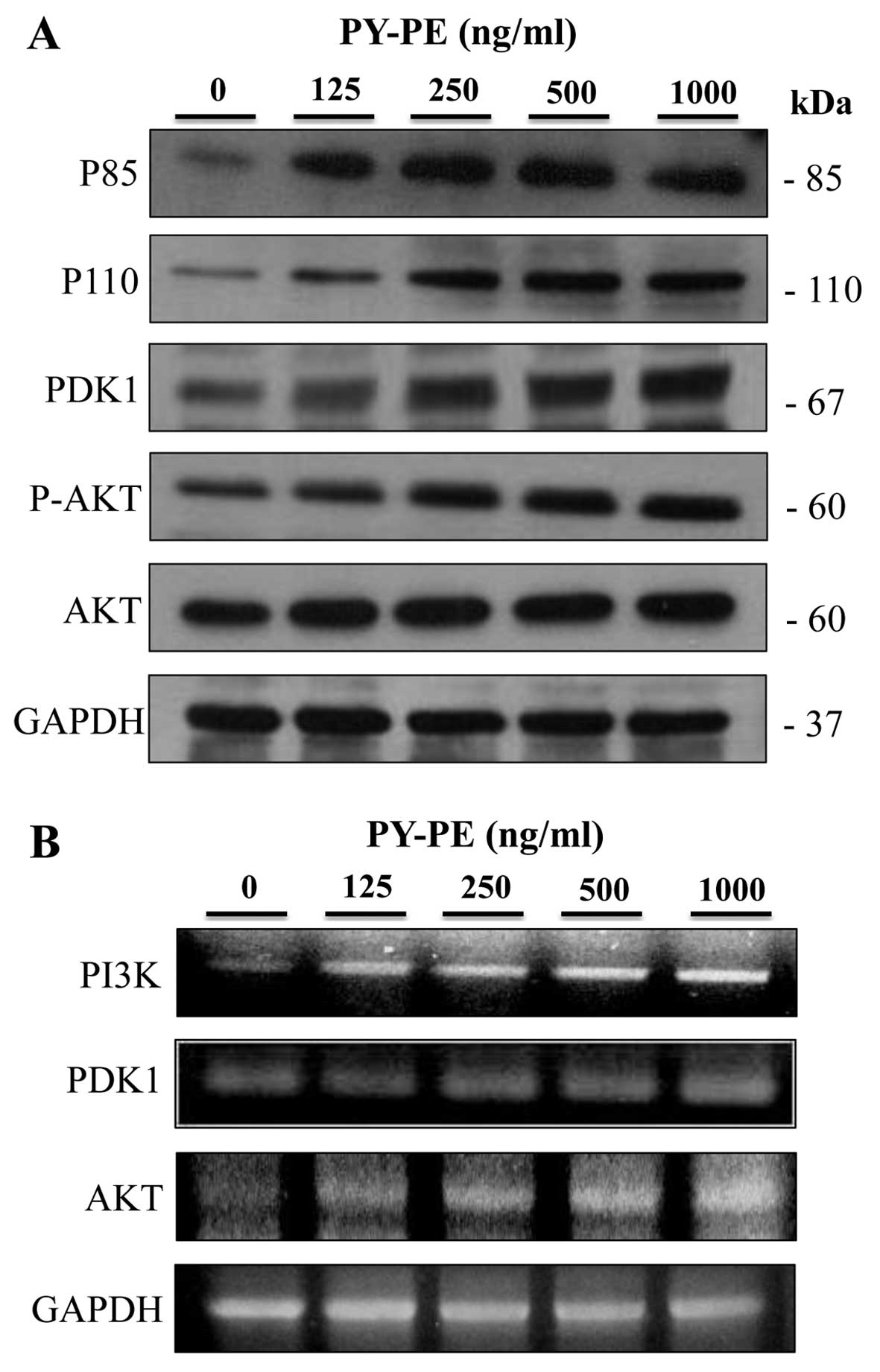
expression of PI3K-Akt signaling pathway proteins
The expression levels of the PI3K-Akt signaling
pathway intermediates were examined by western blotting and RT-PCR.
PY-PE treatment for 24 h resulted in increased protein and mRNA
expression levels of p85, p110, PDK1 and p-Akt compared to the
controls (Fig. 5).
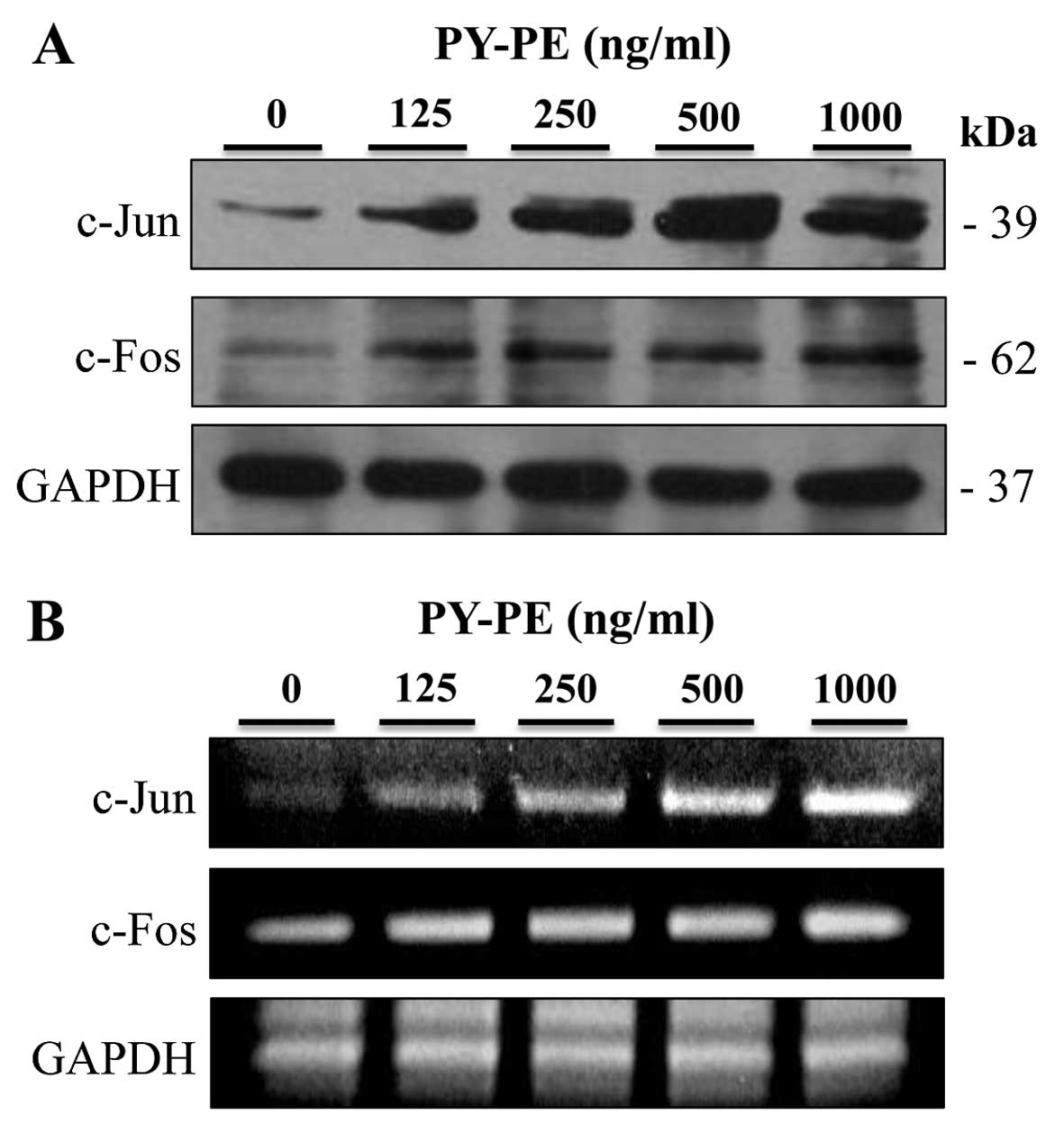
Effect of PY-PE treatment on the
expression of AP-1
IGF-IR activates the PI3K/Akt and p42/p44 MAPK
pathways, which regulate the activation of transcription factors,
such as AP-1 (26). AP-1
regulates cell proliferation and survival. Therefore, AP-1 protein
and mRNA expression levels were examined by western blot analysis
and RT-PCR, respectively. PY-PE treatment of IEC-6 cells
upregulated the protein and mRNA expression levels of c-Jun and
c-fos compared to the controls (Fig.
6).
Discussion
Numerous types of seaweed are important sources of
physiologically active substances, such as peptides and
polysaccharides, and have shown medicinal benefits, including
anti-ulcer, antitumor, antibacterial, antioxidant and antiviral
activities (27–29).
The purpose of the present study was to determine
whether PY-PE induced proliferation of IEC-6 cells and to identify
the associated signals. In an MTS assay, exposure to PY-PE for 24 h
had dose-dependent effects on the proliferation of IEC-6 cells
(Fig. 2). The mechanism
associated with this proliferative effect was subsequently
examined. The regulation of cell proliferation is dependent on
various signaling pathways. In the present study, the IGF-IR
signaling pathway was the main focus.
IGF-IR regulates the biological activity of IGF-I
and plays critical roles in cell growth, differentiation and
proliferation. Additionally, adaptor proteins, such as IRS-1, Shc
and PY-99, have all been indicated in sending signals to the cell
nucleus (7,30). The results of the present study
showed that PY-PE increased the protein and mRNA expression levels
of IGF-IR, IRS-1, Shc and PY-99 (Fig.
3). Following IGF-IR activation, downstream signaling pathways,
including the PI3K and p42/p44 MAPK pathways, are activated
(31). The MAPK family in
mammalian cells includes ERK-1/2, JNK, SAPK and p38 kinase
(32). ERK1/2 is activated
primarily in response to mitogens and growth factors. This pathway
plays an important role in cell proliferation, growth and survival
(33–35). By contrast, JNK and p38 are mostly
activated by cellular stress and by cytokines (36). In the present study, in accordance
with PY-PE-induced cell proliferation, ERK1/2, a critical mediator
that regulates cell growth and proliferation, was activated by
exposure to PY-PE. By contrast, PY-PE treatment did not induce
phosphorylation of JNK or p38 (Fig.
4).
The present results showed that IGF-IR activation
also contributes to PY-PE-induced cell proliferation through
activation of the PI3K/Akt pathway, a key intracellular signaling
pathway that regulates multiple cellular processes, including cell
survival, growth and proliferation (37). PI3K is a heterodimeric protein
composed of two subunits: A p85 regulatory subunit (85 kDa) and a
p110 catalytic subunit (110 kDa) (38,39). Once the p85 regulatory subunit is
positioned appropriately, the p110 catalytic subunit of PI3K
generates phosphatidylinositol 3,4,5-triphosphate, which in turn
activates various downstream targets, including the
serine/threonine kinase Akt (40,41). In the present study, PY-PE
increased the protein and mRNA expression levels of p85, p110 and
PDK1, as well as Akt phosphorylation (Fig. 5).
In a wide range of systems, it has been well
established that growth factor ligation leads to activation of the
p42/p44 MAPK and PI3K/Akt pathways, resulting in the activation of
various transcription factors (42). For instance, platelet-derived
growth factor activates the p42/p44 MAPK pathway, which modulates
AP-1 activation, in NIH 3T3 mouse fibroblasts (43,44). Therefore, PY-PE-induced cell
proliferation was assumed to involve AP-1. These results show that
PY-PE increased the protein and mRNA expression levels of c-Jun and
c-fos (Fig. 6).
In conclusion, PY-PE stimulated the proliferation of
IEC-6 normal intestinal epithelial cells. This effect was also
associated with the IGF-IR signaling pathway. Therefore, PY-PE may
be a potential component of bio-functional foods with a
proliferative effect on intestinal epithelial cells.
Acknowledgments
The present study was supported by the Basic Science
Research Program through the National Research Foundation of Korea
(NRF) funded by the Ministry of Education (grant no.
2012R1A6A1028677).
References
|
1
|
Chopin T, Yarish C, Wilkes R, Belyea E, Lu
S and Mathieson A: Developing Porphyra/salmon integrated
aquaculture or bioremediation and diversification of the
aquaculture industry. J Appl Phycol. 11:463–472. 1999. View Article : Google Scholar
|
|
2
|
Kwon MJ and Nam TJ: Porphyran induces
apoptosis related signal pathway in AGS gastric cancer cell lines.
Life Sci. 79:1956–1962. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shin ES, Hwang HJ, Kim IH and Nam TJ: A
glycoprotein from Porphyra yezoensis produces anti-inflammatory
effects in liposaccharide-stimulated macrophages via the TLR4
signaling pathway. Int J Mol Med. 28:809–815. 2011.PubMed/NCBI
|
|
4
|
Liu F, Liu J, Zhang L, Shen W, Guo T, Liu
C and He P: Ex vivo anti-oxidation activity of polysaccharides from
the red alga Porphyra yezoensis. Res J Pharmacol. 1:61–66.
2007.
|
|
5
|
Suetsuna K: Purification and
identification of angiotensin I-converting enzyme inhibitors from
the red alga Porphyra yezoensis. J Mar Biotechnol. 6:163–167.
1998.PubMed/NCBI
|
|
6
|
Lee AV and Yee D: Insulin-like growth
factor and breast cancer. Biomed Pharmacother. 49:415–421. 1995.
View Article : Google Scholar
|
|
7
|
Butler AA, Yakar S, Gewolb IH, Karas M,
Okubo Y and LeRoith D: Insulin-like growth factor-I receptor signal
transduction: at the interface between physiology and cell biology.
Comp Biochem Physiol B Biochem Mol Biol. 121:19–26. 1998.
View Article : Google Scholar
|
|
8
|
Guvakova MA and Surmacz E: Tamoxifen
interferes with the insulin-like growth factor I receptor (IGF-IR)
signaling pathway in breast cancer cells. Cancer Res. 57:2606–2610.
1997.PubMed/NCBI
|
|
9
|
Pelicci G, Lanfrancone L, Salcini AE, et
al: Constitutive phosphorylation of Shc proteins in human tumors.
Oncogene. 11:899–907. 1995.PubMed/NCBI
|
|
10
|
Habib T, Herrera R and Decker SJ:
Activators of protein kinase C stimulate association of Shc and the
PEST tyrosine phosphatase. J Biol Chem. 269:25243–25246.
1994.PubMed/NCBI
|
|
11
|
Rhodes CJ and White MF: Molecular insights
into insulin action and secretion. Eur J Clin Invest. 32:3–13.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Annunziata M, Cranata R and Ghigo E: The
IGF system. Acta Diabetol. 48:1–9. 2011. View Article : Google Scholar
|
|
13
|
Krasilnikov MA: Phosphatidylinositol-3
kinase dependent pathways: the role in control of cell growth,
survival, and malignant transformation. Biochemistry (Mosc).
65:59–67. 2000.
|
|
14
|
Latres E, Amini AR, Amini AA, et al:
Insulin-like growth factor-1 (IGF-1) inversely regulates
atrophy-induced genes via the phosphatidylinositol
3-kinase/Akt/mammalian target of rapamycin (PI3K/Akt/mTOR) pathway.
J Biol Chem. 280:2737–2744. 2005. View Article : Google Scholar
|
|
15
|
Guha M and Mackman N: The
phosphatidylinositol 3-kinase-Akt pathway limits lipopolysaccharide
activation of signaling pathways and expression of inflammatory
mediators in human monocytic cells. J Biol Chem. 277:32124–32132.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cantley LC: The phosphoinositide 3-kinase
pathway. Science. 296:1655–1657. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Junttila MR, Li SP and Westermarck J:
Phosphatase-mediated crosstalk between MAPK signaling pathways in
the regulation of cell survival. FASEB J. 22:954–965. 2008.
View Article : Google Scholar
|
|
18
|
Roux PP and Blenis J: ERK and p38
MAPK-activated protein kinases: a family of protein kinases with
diverse biological functions. Microbiol Mol Biol Rev. 68:320–344.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Weston GR and Davis RJ: The JNK signal
transduction pathway. Curr Opin Cell Biol. 19:142–149. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Karin M, Liu ZG and Zandi E: AP-1 function
and regulation. Curr Opin Cell Biol. 9:240–246. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Karin M: The regulation of AP-1 activity
by mitogen-activated protein kinases. J Biol Chem. 270:16483–16486.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shaulian E and Karin M: AP-1 in cell
proliferation and survival. Oncogene. 20:2390–2400. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Marais R, Wynne J and Treisman R: The SRF
accessory protein Elk-1 contains a growth factor regulated
transcription activation domain. Cell. 73:381–393. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gopalakrishnan A, Xu CJ, Nair SS, Chen C,
Hebbar V and Kong AN: Modulation of activator protein-1 (AP-1) and
MAPK pathway by flavonoids in human prostate cancer PC3 cells. Arch
Pharm Res. 29:633–644. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Song SH, Kim IH and Nam TJ: Effect of a
hot water extract of Chlorella vulgaris on proliferation of IEC-6
cells. Int J Mol Med. 29:741–746. 2012.PubMed/NCBI
|
|
26
|
Bancroft C, Chen Z, Yeh J, et al: Effects
of pharmacologic antagonists of epidermal growth factor receptor,
PI3K and MEK signal kinases on NF-kappaB and AP-1 activation and
IL-8 and VEGF expression in human head and neck squamous cell
carcinoma lines. Int J Cancer. 99:538–548. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gómez-Ordóñez E, Jiménez-Escrig A and
Rupérez P: Effect of the red seaweed Mastocarpus stellatus intake
on lipid metabolism and antioxidant status in healthy Wistar rats.
Food Chem. 135:806–811. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ronghai H, Haile M and Xinzheng L:
Ultrasonic extraction of polysaccharides from Porphyra yezoensis
and their inhibition effects in U937 cell growth. Trans CSAE.
21:165–168. 2005.
|
|
29
|
Costa LS, Fidelis GPL, Cordeiro SL, et al:
Biological activities of sulfated polysaccharides from tropical
seaweeds. Biomed Pharmacother. 64:21–28. 2010. View Article : Google Scholar
|
|
30
|
Wang Y, Hailey J, Williams D, et al:
Inhibition of insulin-like growth factor-I receptor (IGF-IR)
signaling and tumor cell growth by a fully human neutralizing
anti-IGF-IR antibody. Mol Cancer Ther. 4:1214–1221. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Starkman B, Cravero J, Delcarlo M and
Loeser R: IGF-I stimulation of proteoglycan synthesis by
chondrocytes requires activation of the PI3-Kinase pathway but not
ERK MAPK. Biochem J. 389:723–729. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang W and Liu HT: MAPK signal pathways
in the regulation of cell proliferation in mammalian cells. Cell
Res. 12:9–18. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ballif BA and Blenis J: Molecular
mechanisms mediating mammalian mitogen-activated protein kinase
(MAPK) kinase (MEK)-MAPK cell survival signals. Cell Growth Differ.
12:397–408. 2001.PubMed/NCBI
|
|
34
|
Rubinfeld H and Seger R: The ERK cascade:
a prototype of MAPK signaling. Mol Biotechnol. 31:151–174. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Meloche S and Pouysségur J: The ERK1/2
mitogen-activated protein kinase pathway as a master regulator of
the G1- to S-phase transition. Oncogene. 26:3227–3239. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Davis RJ: Signal transduction by the JNK
group of MAP kinases. Cell. 103:239–252. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Go H, Hwang HJ and Nam TJ: Glycoprotein
extraction from Laminaria japonica promotes IEC-6 cell
proliferation. Int J Mol Med. 24:819–824. 2009.PubMed/NCBI
|
|
38
|
Kapeller R and Cantley LC:
Phosphatidylinositol 3-kinase. Bioessays. 16:565–576. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Carpenter CL, Duckworth B, Auger K, Cohen
B, Schaffhausen B and Cantley LC: Purification and characterization
of phosphoinositide 3-kinase from rat liver. J Biol Chem.
265:19704–19711. 1990.PubMed/NCBI
|
|
40
|
Morgan S and Grandis JR: ErbB receptors in
the biology and pathology of the aerodigestive tract. Exp Cell Res.
315:572–582. 2009. View Article : Google Scholar :
|
|
41
|
Jiang BH, Aoki M, Zheng JZ, Li J and Vogt
PK: Myogenic signaling of phosphatidylinositol 3-kinase requires
the serine-threonine kinase Akt/protein kinase B. Proc Natl Acad
Sci USA. 96:2077–2081. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Qiu Q, Yang M, Tsang BK and Gruslin A:
EGF-induced trophoblast secretion of MMP-9 and TIMP-1 involves
activation of both PI3K and MAPK signalling pathways. Reproduction.
128:355–363. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lavarone C, Catania A, Marinissen MJ,
Visconti R, Acunzo M, Tarantio C, et al: The platelet-derived
growth factor controls c-myc expression through a JNK-and
AP-1-dependent signaling pathway. J Biol Chem. 278:50024–50030.
2003. View Article : Google Scholar
|
|
44
|
Monje P, Marinissen MJ and Gutkind JS:
Phosphorylation of the carboxyl-terminal transactivation domain of
c-Fos by extracellular signal-regulated kinase mediates the
transcriptional activation of AP-1 and cellular transformation
induced by platelet-derived growth factor. Mol Cell Biol.
23:7030–7043. 2003. View Article : Google Scholar : PubMed/NCBI
|