Introduction
Longitudinal bone growth is a results of the
proliferation, hypertrophy and matrix production of cartilage cells
in growth plates (1). The growth
plate is a relatively thin layer of cartilage which is found in
growing long bones during longitudinal skeleton growth (2,3).
The growth of the long bone at the growth plate is regulated by an
endocrine signaling systems that involves glucocorticoids, growth
hormone (GH) and insulin-like growth factor-I (IGF-I) (4). If a child is afflicted by poor
nutrition, the levels of thyroid hormones and IGF-I decline, and
during puberty, malnutrition also causes the downregulation of sex
steroids. These endocrine changes decrease the elongation of bones
(4). In order to reproduce this
malnutrition, mice are fed a low-protein diet. Protein-energy
malnutrition (PEM) is an important form of undernourishment and is
a condition caused by an inequality between food intake (protein
and energy) and the amount that the body requires for optimal
growth and function (5).
GH is a pivotal factor in development and growth
(6). GH begins to exert its
effects when it binds to its receptor, GH receptor (GHR), which is
present on the cell surface (7).
After GH binds to GHR, several signaling cascades are activated,
including the Janus kinase 2 (JAK2)-signal transducer and activator
of transcription (STAT)5 signaling pathway, and the phospholipase
C/protein kinase C/Ca2+ pathway (8,9).
The most well characterized signaling cascade is the JAK2-STAT5
pathway, which mediates the effects of GH on gene transcription in
the liver (10,11). This pathway is responsible for the
transcription of the GH target gene IGF-I (12). IGF-I is considered essential for
longitudinal bone growth and plays an important role in cell growth
and survival. In general, the majority of plasma IGF-I is produced
in the liver (13–15).
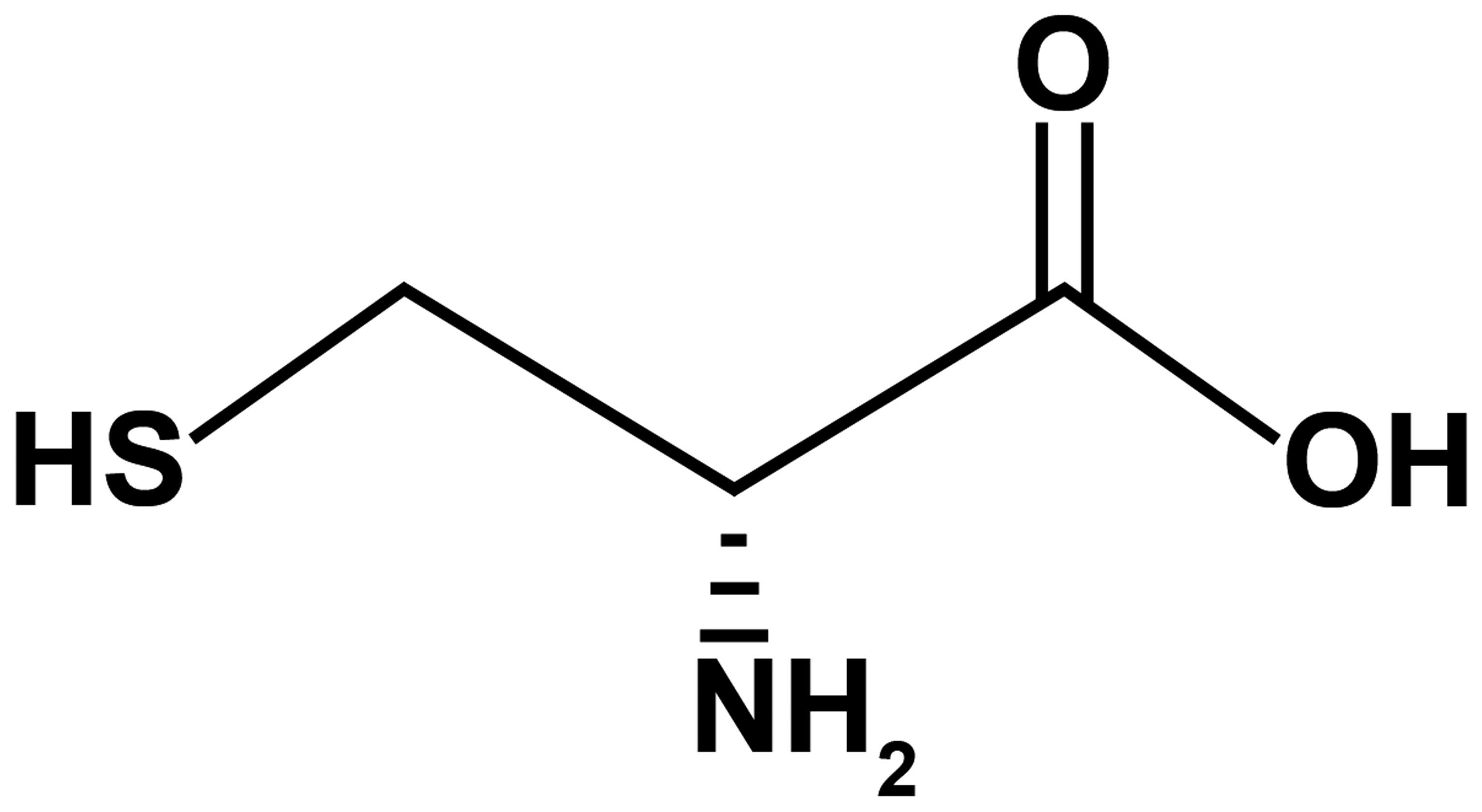
Cysteine (Cys) (Fig.
1) is a semi-essential amino acid that is known to exert
antioxidant, antipancreatitic and antidiabetic effects (16–18). However, the effects of Cys on bone
growth have not been elucidate to date. Thus, in this study, we
aimed to investigate the effects of Cys on longitudinal bone growth
and to elucidate the underlying mechansims.
Materials and methods
Reagents
Cys, avidin-peroxidase and bovine serum albumin were
purchased from Sigma Chemical Co. (St. Louis, MO, USA). Anti-mouse
IGF-I antibody (Cat. no. MAB791), biotinylated anti-mouse IGF-I
antibody (Cat. no. BAF791) and recombinant mouse IGF-I (Cat. no.
791-MG-050) were purchased from R&D Systems (Minneapolis, MN,
USA). The phosphorylated (p-)JAK2 (Cat. no. 44-426G) and STAT5
(Cat. no. 71-6900) antibodies were purchased from Invitrogen Life
Technologies (Camarillo, CA, USA). JAK2 (Cat. no. sc-294) and STAT5
(Cat. no. sc-836) antibodies were purchased from Santa Cruz
Biotechnology (Santa Cruz, CA, USA).
Cell culture and treatment
The human hepatocellular carcinoma cell line, HepG2
(from the Korean Cell Line Bank), was grown in DMEM (Gibco, Grand
Island, NY, USA) containing penicillin (100 units/ml; Gibco),
streptomycin (100 µg/ml; Gibco) and 10% fetal bovine serum
(HyClone, Logan, UT, USA) at 37°C in an incubator with 95%
humidified air/5% CO2. The cells were treated with
0.5-50 µg/ml of Cys for 12 h. The control group represented
cells treated with the vehicle (distilled water).
Animals and diet
Male Institute of Cancer Research (ICR) mice (4
weeks old) and food were purchased from Dae-Han Experimental Animal
Center (Eumsung, Korea). The mice were allowed to acclimatize for 7
days and were then randomly assigned to an adequate-protein diet
group [control (CON) group, 20% protein] or a low-protein diet
group (PEM group, 4% protein), as previously described (19). The protein source used was casein.
Aside from the protein content, the 2 diets were identical and
isocaloric (Table I). After 2
weeks, the mice were divided into 3 groups (5 mice per group) as
follows: i) the CON group: mice were fed an adequate-protein diet +
distilled water (DW); ii) the PEM group: mice were fed a
low-protein diet + DW; and iii) the Cys group: mice were fed a
low-protein diet and administered Cys. The mice were fed an
adequate-protein diet or a low-protein diet and were administered
orally with DW or Cys (50 mg/kg) using a stomach sonde twice a week
for 12 weeks, as previously described in the study by Farombi et
al (20). It is widely
accepted that bone growth is associated with a gain in body weight.
Thus, the mice were weighed at the end of the experiment. The mice
were housed in a laminar air-flow room, with a constant temperature
of 22±1°C and a relative humidity of 55±1%, throughout the study.
All epxeriments were conducted in accordance with internationally
accepted principles for laboratory animal use and care, as found in
US guidelines (NIH publication no. 85–23, revised in 1985).
 | Table IComposition of experimental
diets. |
Table I
Composition of experimental
diets.
| Ingredients | Standard chow diet
(g/kg diet) | PEM diet (g/kg
diet) |
|---|
| Casein (>85%
protein) | 200 | 40 |
| Sucrose | 100 | 100 |
| Fiber | 10 | 10 |
| Corn oil | 80 | 80 |
| Mineral
mixture | 40 | 40 |
| Vitamin
mixture | 10 | 10 |
| L-methionine | 1.5 | 1.5 |
| Choline
bitartrate | 2.5 | 2.5 |
| Cornstarch | 556.5 | 716.5 |
Enzyme-linked immunosorbent assay
(ELISA)
The levels of IGF-I were measured by a modified
ELISA, as previously described (21–24). A sandwich ELISA for IGF-I was
carried out in a duplicate in a 96-well ELISA plate. The plate was
coated with anti-mouse IGF-I antibody and incubated overnight at
4°C. The plate was washed in PBS containing 0.05% Tween-20 (Sigma)
and blocked with PBS containing 1% bovine serum albumin, 5% sucrose
and 0.05% NaN3 for 1 h. After additional washes, the
culture supernatant and IGF-I standards were added and incubated at
room temperature for 2 h. After 2 h incubation at room temperature,
the plate was washed and biotinylated anti-mouse IGF-I was then
added and again incubated at room temperature for 2 h. After
washing the wells, avidin-peroxidase was added and the plate was
incubated for 30 min at 37°C. The wells were again washed and TMB
substrate (Pharmingen, San Diego, CA, USA) was added. Color
development was measured at 405 nm using an automated microplate
ELISA reader. A standard curve was run on the plate using
recombinant mouse IGF-I in serial dilutions.
Quantitative polymerase chain reaction
(qPCR)
qPCR was performed using SYBR-Green Master Mix, and
the detection of mRNA was carried out using an ABI StepOne
real-time PCR system (Applied Biosystems, Foster City, CA, USA), as
previously described (25–27).
PCR was performed with the following primers: human IGF-I forward,
5′-TGC CCA AGA CCC AGA AGT-3′ and reverse, 5′-CTC CTG TCC CCT CCT
TCT GTT-3′; human GAPDH forward, 5′-TCG ACA GTC AGC CGC ATC TTC
TTT-3′ and reverse, 5′-ACC AAA TCC GTT GAC TCC GAC CTT-3′; mouse
IGF-I forward, 5′-CCG GAC CAG AGA CCC TTT G-3′ and reverse, 5′-CCT
GTG GGC TTG TTG AAG TAA AA-3′; and mouse GAPDH forward, 5′-GGC AAA
TTC AAC GGC ACA-3′ and reverse, 5′-GTT AGT GGG GTC TCG CTC CTG-3′.
Typical profile times used were an initial step, 95°C for 10 min
followed by a second step at 95°C for 15 sec and 60°C for 30 sec
for 40 cycles with a melting curve analysis. The level of target
mRNA was normalized to the level of GAPDH and compared with the
control. Data were analyzed using the ΔΔCT method.
Western blot analysis
The supernatants of the homogenized liver tissues
were prepared in a sample buffer [62.5 mM Tris hydrochloride
(Tris-HCl), pH 6.8, 2% sodium dodecyl sulphate (SDS), 20% glycerol
and 10% 2-mercaptoethanol], as previously described (28–30). The samples were heated at 95°C for
5 min and briefly cooled on ice. Following centrifugation at 15,000
x g for 5 min, the proteins in the lysates were then separated by
10% SDS-polyacrylamide gel electrophoresis and transferred onto a
nitrocellulose membrane. The membrane was blocked with 5% skim milk
in PBS-tween-20 for 1 h at room temperature and then incubated with
primary (p-JAK2, 1:500 dilution; p-STAT5, 1:500 dilution; JAK2,
1:1,000 dilution; STAT5, 1:1,000 dilution) and secondary (mouse
anti-rabbit IgG-HRP, 1:5,000 dilution, Santa Cruz Biotechnology,
Santa Cruz, CA, USA) antibodies. Finally, the protein bands were
visualized by an enhanced chemiluminesence solution purchased from
Amersham Co. (Newark, NJ, USA) following the manufacturer’s
instructions.
Micro-computed tomography (µCT)
The 2D and 3D information on bone geometry was
attained by high-resolution µCT, as previously described
(31,32). The mice were sacrificed; the
femora and tibiae were dissected, cleaned of soft tissue, and fixed
in 4% formaldehyde before storage. The source of the open tube type
and the minimum focal spot size was 8 µm. Reconstruction was
carried out using a modified Feldkamp algorithm using SkyScan
NRecon software (SkyScan, Kontich, Belgium). The x-ray source was
set at 75 kV and 100 mA. Four hundred projections were acquired
over an angle of 180°. The image slices were reconstructed using
cone-beam reconstruction software based on the Feldkamp algorithm
(Dataviewer; SkyScan). The trabecular bone was extracted by drawing
ellipsoid contours with CT analyzer software. Trabecular bone
volume (BV/TV; percentage) and trabecular number (Tb.N) of femur
epiphysis and proximal tibial metaphysis were calculated by the
mean intercept length method. Trabecular thickness (Tb.Th; mm) was
calculated according to the study of Ngueguim et al
(31). 3D parameters were based
on analysis of a Marching cubes-type model with a rendered surface.
To analyze 3D parameters in tibias, whole bone was scanned, and 600
slices of 8 µm in thickness were placed through the former
area. Bone mineral density (BMD), bone volume/tissue volume
(BV/TV), trabecular thickness (Tb.Th), trabecular number (Tb.N),
connectivity density (Conn.D), and total porosity were recorded.
The thickness of excised bone growth plate was determined using
SkyScan 1076 software (SkyScan) as described in a previously
published protocol (32).
Statistical analysis
Statistical analysis was performed using SPSS
software (version 14.0; SPSS Inc., Chicago, IL, USA). All results
are expressed as the means ± SEM. The statistical evaluation of the
results was performed by an independent t-test and an ANOVA with
Tukey’s post hoc test. P values <0.05 were considered to
indicate statistically significant differences.
Results
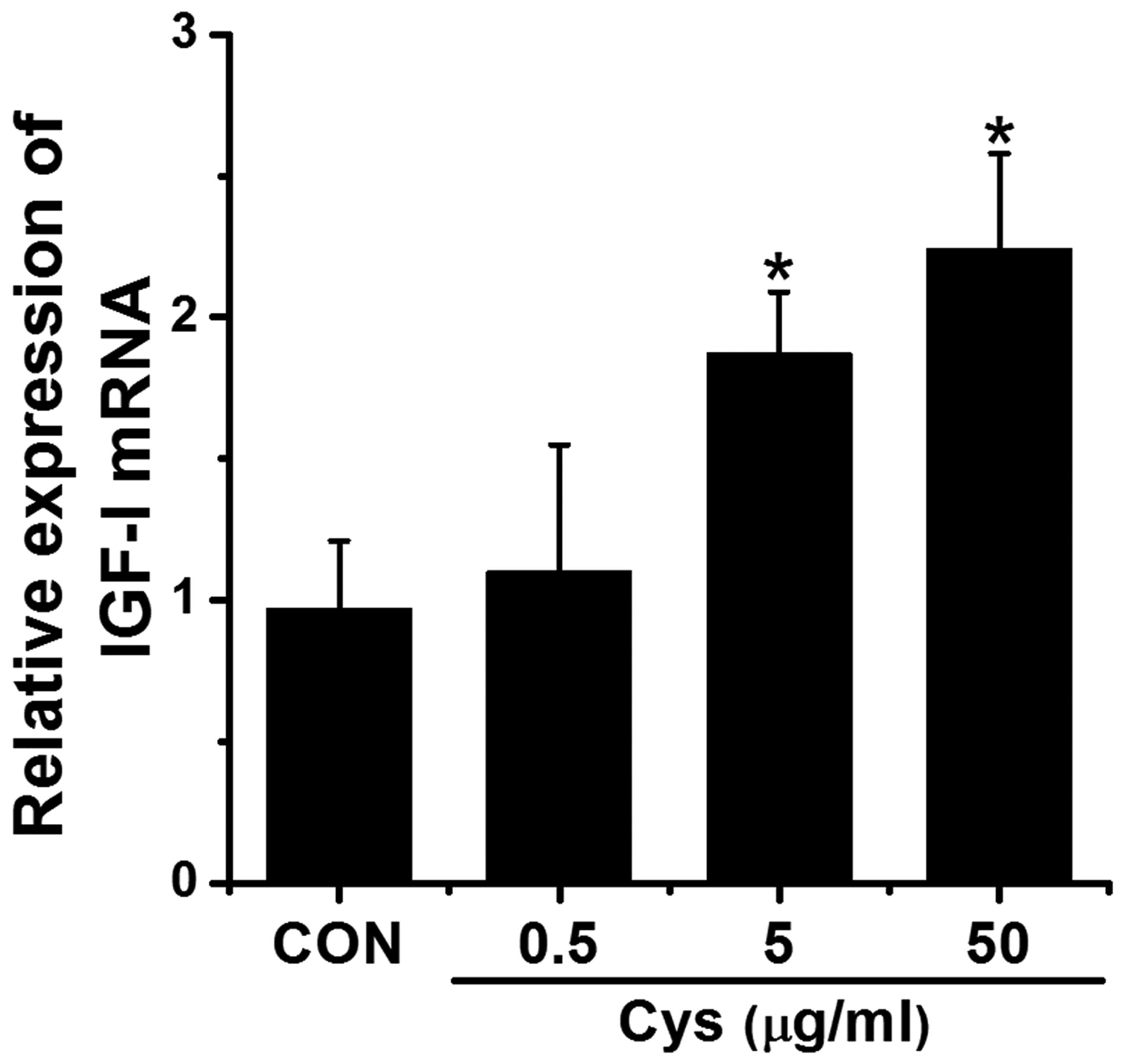
Effect of Cys on IGF-I mRNA in HepG2
cells
IGF-I plays an important role in linear bone growth
(33). Thus, to determine whether
Cys increases IGF-I mRNA expression in the liver, we used a
hepatocyte cell line, HepG2. When we treated the HepG2 cells with
Cys (0.5–50 mg/ml), IGF-I mRNA expression was elevated in a
dose-dependent manner (Fig. 2).
Treatment with 50 mg/ml of Cys had the most prominent effect, and
thus we evaluated the effects of treatment with 50 mg/kg of Cys in
the next set of experiments, i.e., in vivo mouse models.
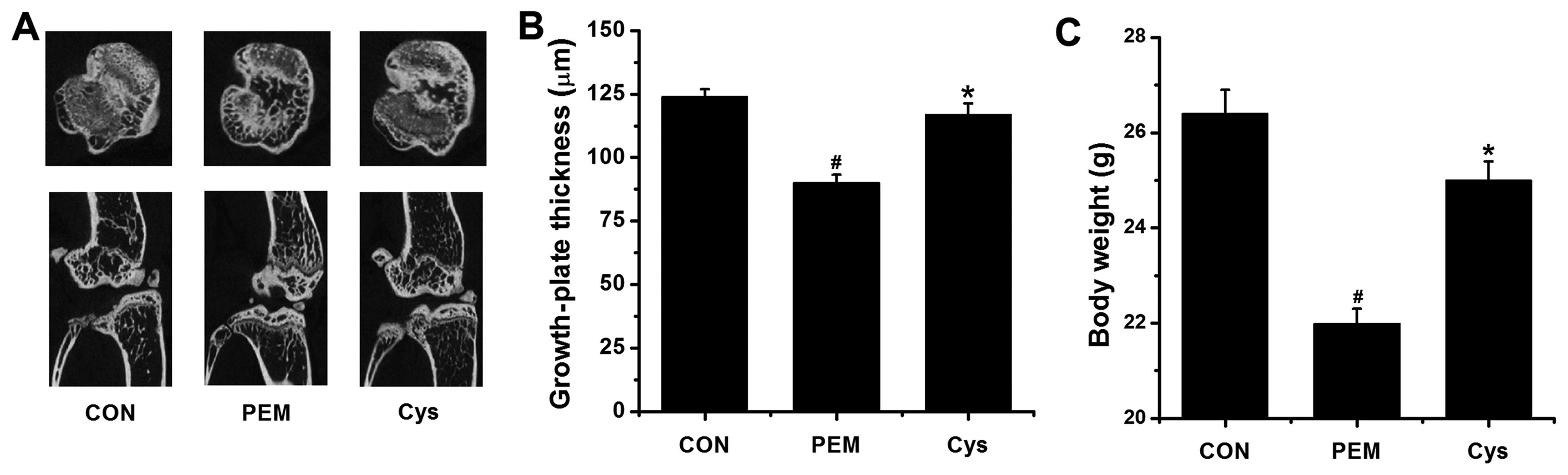
Effect of Cys on growth plates in
mice
The thickness of the growth plate in the proximal
tibia was assessed (Fig. 3A), and
we also investigated linear bone growth using µCT. The
thickness of the growth plates in the proximal tibias in the CON
and PEM groups were 124.0±2.9 and 90.1±3.1, respectively; the
growth-plate thickness in the Cys group was 117.0±4.3 (P<0.05;
Fig. 3B). Thus, it is clear that
Cys significantly increased linear bone growth, compared to the PEM
group. In addition, treatment with Cys significantly increased the
body weight of the mice compared to the PEM group.
Effect of Cys on trabecular bone
parameters
We used µCT measurements to determine whether
Cys improves longitudinal bone growth. The administration of Cys
increased BMD (Fig. 4A). The
results of the 3D µCT reconstruction of the trabecular bone
images, which were converted into parameters representing
trabecular connectivity, demonstrated that the BV/TV, Tb.Th, Tb.N
and Conn.D in the tibia were increased by treatment with Cys, and
that total porosity was decreased (Fig. 4B–F).
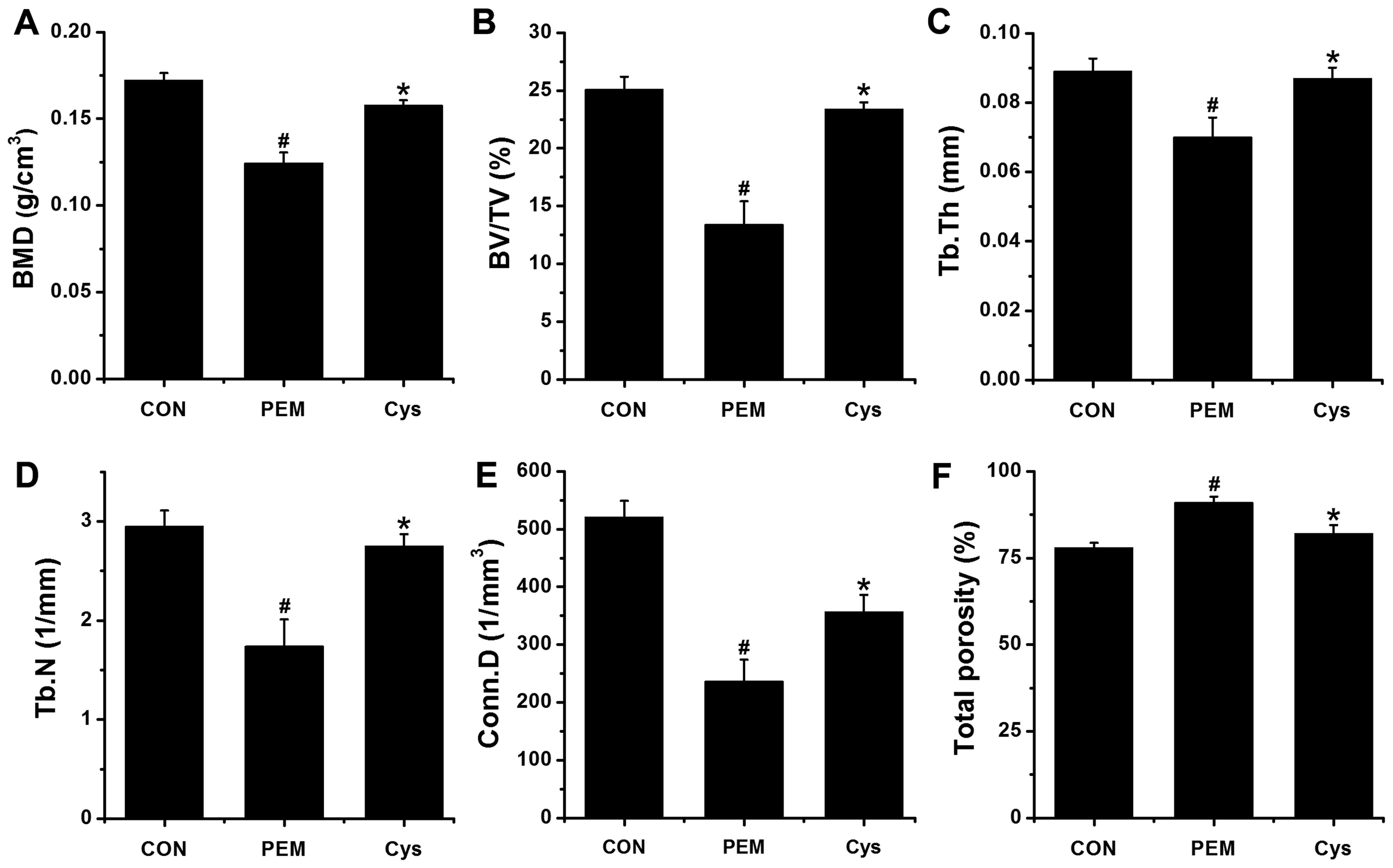 | Figure 4Effect of cysteine (Cys) on
trabecular bone parameters. Bone mineral density (BMD) and
trabecular parameters of excised bone at the tibia were measured.
(A) BMD levels were measured using 2D micro-computed tomography
(µCT). (B–F) Quantified parameters (BV/TV, Tb.Th, Tb.N,
Con.D and total porosity) of proximal tibia were measured using 3D
µCT. Data represent the means ± SEM of 3 independent
experiments. #P<0.05 denotes a significan difference
compared to the CON. *P<0.05 denotes a significant
difference compared to the protein-energy malnutrition (PEM) group.
BV/TV, bone volume/tissue volume; Tb.Th, trabecular thickness;
Tb.N, trabecular number; Conn.D, connectivity density; CON, group
administered adequate-protein diet + distilled water (DW); PEM,
group administered low-protein diet + DW; Cys, group administered
low-protein diet + Cys. |
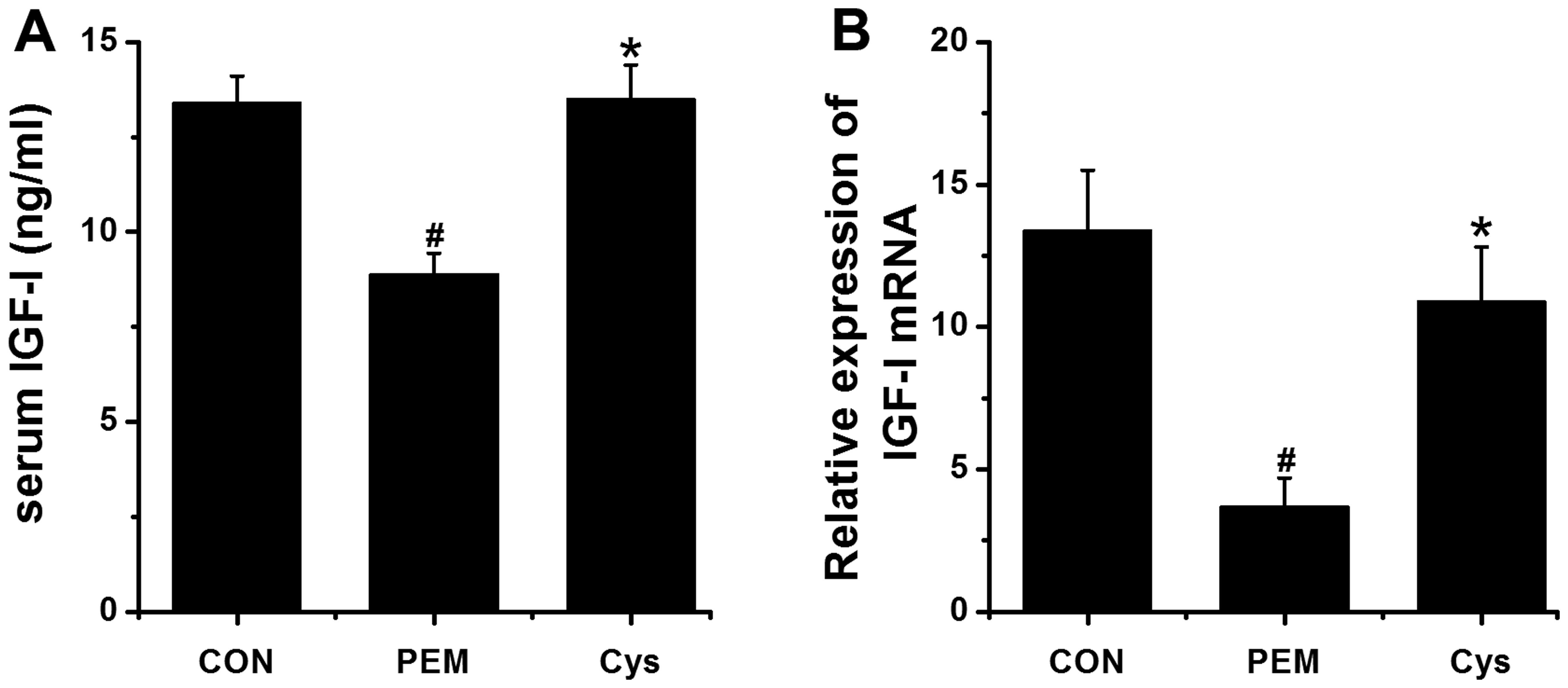
Effect of Cys on the regulation of
IGF-I
To investigate whether Cys increase serum IGF-I
levels, we measured the IGF-I levels by ELISA. The serum IGF-I
levels were significantly increased by Cys, compared to those of
the PEM group (P<0.05; Fig.
5A). We then examined the mRNA expression of IGF-I in the
liver, as the majority of plasma IGF-I is produced in the liver.
The mRNA expression of IGF-I in the liver was shown to be
significantly upregulated in the mice administered Cys, compared to
the mice in the PEM group (P<0.05; Fig. 5B).
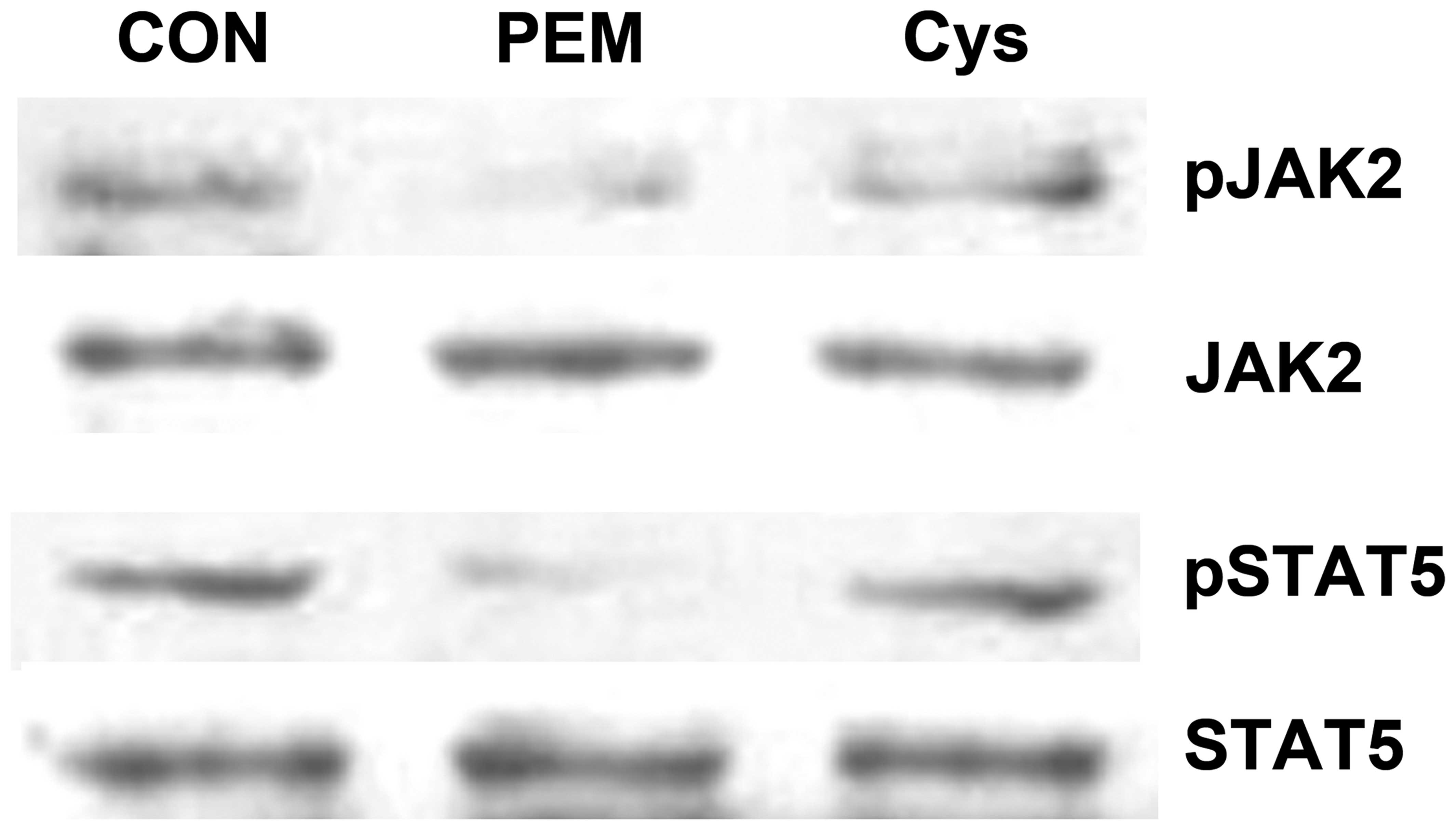
Effect of Cys on levels of p-JAK2 and
p-STAT5
To examine whether the linear bone growth caused by
Cys is regulated by JAK2-STAT5 phosphorylation, we performed
western blot analysis. As depicted in Fig. 6, the levels of p-JAK2 and p-STAT5
in the liver were decreased due to malnutrition. However, the
decrease in the level sof p-JAK2 and p-STAT5 was reversed by
treatment with Cys (Fig. 6).
Discussion
In the present study, we demonstrated that Cys
increased the mRNA expression of IGF-I, increased the thickness of
growth plates, and increased the BV/TV, Tb.Th, Tb.N and Conn.D; Cys
decreased total porosity. In addition, Cys increased the serum
IGF-I levels through JAK2-STAT5 phosphorylation.
In general, endochondral cell proliferation in the
growth plate results in bone growth. Thus, growth-plate thickness
is a direct indicator of linear bone growth (34). Our results demonstrated that Cys
increased growth-plate thickness (Fig. 3). From this result, it is
suggested that Cys may prove to be helpful to children whose growth
is retarded.
Cys is a sulfur-containing amino acid (35). A deficiency of sulfur amino acid
has been shown to decrease neonatal pig growth (36). McLean et al (37) reported that Cys restored growth
which was inhibited by paracetamol. In addition, Cys has been shown
to induce the growth of human peritoneal mesothelial cells
(38). In the present study, the
sulfur-containing amino acid, Cys, increased growth-plate thickness
in mice fed a PEM diet (Fig. 3).
Thus, we can presume that the effect of Cys on bone growth, in
part, is due to sulfur.
Baines et al (39) reported that subjects with low BMD
had a significantly lower plasma Cys concentration, suggesting that
there is an association between Cys and bone mass. In the present
study, we demonstrated that Cys elevated tibial BMD, in ways
consistent with a previous report (39). Furthermore, we noted that Cys
improved bone parameters (it increased BV/TV, Tb.Th, Tb.N, Conn.D
and decreased total porosity; Fig.
4B-F). Thus, Cys may have the potential to improve not only
growth retardation, but may also alleviate osteoporosis.
Growth retardation is a major concern in children
with chronic inflammatory diseases, uncontrolled juvenile
idiopathic arthritis and chronic kidney disease (40–42). Thus, many children are
administered the anti-inflammatory drug, dexamethasone. However,
Baron et al (43) reported
that dexamethasone acts locally to suppress linear bone growth in
rabbits. It is fortunate that IGF-1 possesses anti-apoptotic
properties and may, therefore, have the ability to prevent
dexa-methasone-induced chondrocyte apoptosis. Chrysis et al
(44) demonstrated that IGF-I
prevented dexamethasone-induced apoptosis, as well as the
suppression of chondrocyte proliferation. In the present study, Cys
elevated the level of serum IGF-I (Fig. 5). Thus, we can speculate that Cys
partially counteracts the dexamethasone-induced growth retardation
through the upregulation of IGF-I.
GH has long been known to stimulate linear growth
and regulate metabolism. The binding of GH to GHR promotes the
phosphorylation of JAK2 and STAT5 (45,46). GH-induced IGF-I gene transcription
is mediated by STAT5B (12). In
our study, we noted that Cys increased the levels of p-JAK2 and
p-STAT5 (Fig. 6). Thus, our
findings suggest that Cys promotes linear bone growth through the
phosphorylation of JAK2-STAT5.
In conclusion, in the present study, we demonsrated
that Cys increased growth-plate thickness and improved bone
parameters in mice. In addition, Cys upregulated the IGF-I levels
through the phosphorylation of JAK2-STAT5. Overall, these results
suggest that Cys may prove to be helpful in children whose growth
is retarded, as it induces longitudinal bone growth through the
upregulation of IGF-I.
Acknowledgments
This study was supported by the Ministry of Trade,
Industry and energy (MOTIE), KOREA, through the Education Support
program for Creative and Industrial Convergence.
References
|
1
|
Brito I, Gil-Peña H, Molinos I, Loredo V,
Henriques-Coelho T, Caldas-Afonso A and Santos F: Growth cartilage
expression of growth hormone/insulin-like growth factor I axis in
spontaneous and growth hormone induced catch-up growth. Growth Horm
IGF Res. 22:129–133. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Abad V, Meyers JL, Weise M, Gafni RI,
Barnes KM, Nilsson O, Bacher JD and Baron J: The role of the
resting zone in growth plate chondrogenesis. Endocrinology.
143:1851–1857. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Loqman MY, Bush PG, Farquharson C and Hall
AC: Suppression of mammalian bone growth by membrane transport
inhibitors. J Cell Biochem. 114:658–668. 2013. View Article : Google Scholar
|
|
4
|
Lui JC and Baron J: Effects of
glucocorticoids on the growth plate. Endocr Dev. 20:187–193. 2011.
View Article : Google Scholar
|
|
5
|
Fock RA, Rogero MM, Vinolo MA, Curi R,
Borges MC and Borelli P: Effects of protein-energy malnutrition on
NF-kappaB signalling in murine peritoneal macrophages.
Inflammation. 33:101–109. 2010. View Article : Google Scholar
|
|
6
|
de Vos AM, Ultsch M and Kossiakoff AA:
Human growth hormone and extracellular domain of its receptor:
crystal structure of the complex. Science. 255:306–312. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brooks AJ and Waters MJ: The growth
hormone receptor: mechanism of activation and clinical
implications. Nat Rev Endocrinol. 6:515–525. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Le Roith D, Bondy C, Yakar S, Liu JL and
Butler A: The somato-medin hypothesis: 2001. Endocr Rev. 22:53–74.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gevers EF, Hannah MJ, Waters MJ and
Robinson IC: Regulation of rapid signal transducer and activator of
transcription-5 phosphorylation in the resting cells of the growth
plate and in the liver by growth hormone and feeding.
Endocrinology. 150:3627–3636. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Davey HW, Park SH, Grattan DR, McLachlan
MJ and Waxman DJ: STAT5b-deficient mice are growth hormone
pulse-resistant. Role of STAT5b in sex-specific liver p450
expression. J Biol Chem. 274:35331–35336. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rabkin R, Sun DF, Chen Y, Tan J and
Schaefer F: Growth hormone resistance in uremia, a role for
impaired JAK/STAT signaling. Pediatr Nephrol. 20:313–318. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Woelfle J, Chia DJ and Rotwein P:
Mechanisms of growth hormone (GH) action. Identification of
conserved Stat5 binding sites that mediate GH-induced insulin-like
growth factor-I gene activation. J Biol Chem. 278:51261–51266.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang R, Ruan D and Zhang C: Effects of
TGF-beta1 and IGF-1 on proliferation of human nucleus pulposus
cells in medium with different serum concentrations. J Orthop Surg
Res. 1:92006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Weng CY, Kothary PC, Verkade AJ, Reed DM
and Del Monte MA: MAP kinase pathway is involved in
IGF-1-stimulated proliferation of human retinal pigment epithelial
cells (hRPE). Curr Eye Res. 34:867–876. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
King B, Jiang Y, Su X, Xu J, Xie L,
Standard J and Wang W: Weight control, endocrine hormones and
cancer prevention. Exp Biol Med. 238:502–508. 2013. View Article : Google Scholar
|
|
16
|
Tuncer PB, Bucak MN, Büyükleblebici S,
Sarıözkan S, Yeni D, Eken A, Akalın PP, Kinet H, Avdatek F, Fidan
AF and Gündoğan M: The effect of cysteine and glutathione on sperm
and oxidative stress parameters of post-thawed bull semen.
Cryobiology. 61:303–307. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Salman ZK, Refaat R, Selima E, El Sarha A
and Ismail MA: The combined effect of metformin and L-cysteine on
inflammation, oxidative stress and insulin resistance in
streptozotocin-induced type 2 diabetes in rats. Eur J Pharmacol.
714:448–455. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang LJ, Wan R, Shen JQ, Shen J and Wang
XP: Effect of L-cysteine on remote organ injury in rats with severe
acute pancreatitis induced by bile-pancreatic duct obstruction.
Hepatobiliary Pancreat Dis Int. 12:428–435. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Reeves PG, Nielsen FH and Fahey GC Jr:
AIN-93 purified diets for laboratory rodents: final report of the
American Institute of Nutrition ad hoc writing committee on the
reformulation of the AIN-76A rodent diet. J Nutr. 123:1939–1951.
1993.PubMed/NCBI
|
|
20
|
Farombi EO, Nwankwo JO and Emerole GO: The
effect of modulation of glutathione levels on markers for aflatoxin
B1-induced cell damage. Afr J Med Med Sci. 34:37–43.
2005.PubMed/NCBI
|
|
21
|
Choi IY, Moon PD, Koo HN, Myung NY, Kim
SJ, Lee JH, Han SH, Moon G, Seo SY and Sung HJ: Observations of
Forsythia koreana methanol extract on mast cell-mediated allergic
reactions in experimental models. In Vitro Cell Dev Biol Anim.
43:215–221. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Moon PD and Kim HM: Thymic stromal
lymphopoietin is expressed and produced by caspase-1/NF-κB pathway
in mast cells. Cytokine. 54:239–243. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Moon PD and Kim HM: Suppression of thymic
stromal lympho-poietin production by rutin in mast cells. Food
Chem. 133:76–81. 2012. View Article : Google Scholar
|
|
24
|
Moon PD, Jeong HJ and Kim HM:
Down-regulation of thymic stromal lymphopoietin by curcumin.
Pharmacol Rep. 65:525–531. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Han NR, Kim HM and Jeong HJ: Kanamycin
activates caspase-1 in NC/Nga mice. Exp Dermatol. 20:659–663. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Moon PD, Choi IH and Kim HM:
Epigallocatechin-3-O-gallate inhibits the production of thymic
stromal lymphopoietin by the blockade of caspase-1/NF-κB pathway in
mast cells. Amino Acids. 42:2513–2519. 2012a. View Article : Google Scholar
|
|
27
|
Moon PD, Jeong HJ, Kim SJ, An HJ, Lee HJ,
Yang WM, Park SK, Hong SH, Kim HM and Um JY: Use of
electroacupuncture at ST36 to inhibit anaphylactic and inflammatory
reaction in mice. Neuroimmunomodulation. 14:24–31. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Moon PD and Kim HM: The suppression of
thymic stromal lymphopoietin expression by selenium. Amino Acids.
43:999–1004. 2012. View Article : Google Scholar
|
|
29
|
Moon PD, Jeong HJ and Kim HM: Effects of
schizandrin on the expression of thymic stromal lymphopoietin in
human mast cell line HMC-1. Life Sci. 91:384–388. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Moon PD and Kim HM: Antiinflammatory
effects of traditional Korean medicine, JinPi-tang and its active
ingredient, hesperidin in HaCaT cells. Phytother Res. 26:657–662.
2012. View
Article : Google Scholar
|
|
31
|
Ngueguim FT, Khan MP, Donfack JH, Siddiqui
JA, Tewari D, Nagar GK, Tiwari SC, Theophile D, Maurya R and
Chattopadhyay N: Evaluation of Cameroonian plants towards
experimental bone regeneration. J Ethnopharmacol. 141:331–337.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sharan K, Mishra JS, Swarnkar G, Siddiqui
JA, Khan K, Kumari R, Rawat P, Maurya R, Sanyal S and Chattopadhyay
N: A novel quercetin analogue from a medicinal plant promotes peak
bone mass achievement and bone healing after injury and exerts an
anabolic effect on osteoporotic bone: the role of aryl hydrocarbon
receptor as a mediator of osteogenic action. J Bone Miner Res.
26:2096–2111. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Locatelli V and Bianchi VE: Effect of
GH/IGF-1 on bone metabolism and osteoporsosis. Int J Endocrinol.
Jul 23–2014. View Article : Google Scholar : Epub ahead of print.
PubMed/NCBI
|
|
34
|
Lee SH, Kim JY, Kim H, Park SK, Kim CY,
Chung SY and Chang GT: Amomum villosum induces longitudinal bone
growth in adolescent female rats. J Tradit Chin Med. 32:453–458.
2012. View Article : Google Scholar
|
|
35
|
Suzuki Y, Suda K, Matsuyama Y, Era S and
Soejima A: Close relationship between redox state of human serum
albumin and serum cysteine levels in non-diabetic CKD patients with
various degrees of renal function. Clin Nephrol. 82:320–325. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bauchart-Thevret C, Stoll B, Chacko S and
Burrin DG: Sulfur amino acid deficiency upregulates intestinal
methionine cycle activity and suppresses epithelial growth in
neonatal pigs. Am J Physiol Endocrinol Metab. 296:E1239–1250. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
McLean AE, Armstrong GR and Beales D:
Effect of D- or L-methionine and cysteine on the growth inhibitory
effects of feeding 1% paracetamol to rats. Biochem Pharmacol.
38:347–352. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bird SD, Legge M and Walker RJ: L-cysteine
improves growth of human peritoneal mesothelial cells in vitro.
Perit Dial Int. 16:599–606. 1996.PubMed/NCBI
|
|
39
|
Baines M, Kredan MB, Davison A, Higgins G,
West C, Fraser WD and Ranganath LR: The association between
cysteine, bone turnover, and low bone mass. Calcif Tissue Int.
81:450–454. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Schmeling H, Seliger E and Horneff G:
Growth reconstitution in juvenile idiopathic arthritis treated with
etanercept. Clin Exp Rheumatol. 21:779–784. 2003.
|
|
41
|
Marcovecchio ML, Mohn A and Chiarelli F:
Inflammatory cytokines and growth in childhood. Curr Opin
Endocrinol Diabetes Obes. 19:57–62. 2012. View Article : Google Scholar
|
|
42
|
Mehls O and Fine RN: Growth hormone
treatment after renal transplantation: a promising but underused
chance to improve growth. Pediatr Nephrol. 28:1–4. 2013. View Article : Google Scholar
|
|
43
|
Baron J, Huang Z, Oerter KE, Bacher JD and
Cutler GB Jr: Dexamethasone acts locally to inhibit longitudinal
bone growth in rabbits. Am J Physiol. 263:E489–E492.
1992.PubMed/NCBI
|
|
44
|
Chrysis D, Zaman F, Chagin AS, Takigawa M
and Sävendahl L: Dexamethasone induces apoptosis in proliferative
chondrocytes through activation of caspases and suppression of the
Akt-phosphatidylinositol 3′-kinase signaling pathway.
Endocrinology. 146:1391–1397. 2005. View Article : Google Scholar
|
|
45
|
Argetsinger LS and Carter-Su C: Mechanism
of signaling by growth hormone receptor. Physiol Rev. 76:1089–1107.
1996.PubMed/NCBI
|
|
46
|
Sandoval-Usme MC, Umaña-Pérez A, Guer ra
B, Hernández-Perera O, García-Castellano JM, Fernández-Pérez L and
Sánchez-Gómez M: Simvastatin impairs growth hormone-activated
signal transducer and activator of transcription (STAT) signaling
pathway in UMR-106 osteosarcoma cells. PLoS One. 9:e877692014.
View Article : Google Scholar : PubMed/NCBI
|