Introduction
It is well known that the efficiency of the wound
healing process is reduced with aging, and the skin becomes more
fragile and susceptible to trauma (1). Estrogen deprivation in
post-menopausal women is considered responsible for a number of
issues associated with aging, including poor wound healing
(2–4). Notably, in females, estrogen
replacement therapy (ERT) has been proven to reverse delayed wound
healing which is related to aging, and this effect is mediated by
at least two basic mechanisms: i) the downregulation of the
expression of macrophage migration inhibitory factor (5), a key player in skin biology and
wound healing (6); and ii) the
increase of transforming growth factor-β1 (TGF-β1) production by
dermal fibroblasts (7).
Cells with low-level differentiation potential have
the ability to stimulate tissue renewal (8), which is of great significance in
tumor growth and/or tissue repair (9,10).
Keratinocytes proliferate and migrate over the wound to create a
barrier between the outer and inner environments (11), through re-epithelialization. The
level of keratinocyte differentiation can change during the process
of epithelialization, determined by assessing the presence of
distinct keratins (12). For
example, the expression of keratin-10 is restricted to
differentiated keratinocytes located in the suprabasal epidermal
layer (13,14). By contrast, keratin-14 positivity
is considered a marker of proliferating, non-terminally
differentiated keratinocytes located in the basal layer of the
epidermis (13,15). In addition, the expression of
keratin-19 is confined to cells of hair follicles (16), a characteristic which exemplifies
the stem cell-like character of keratinocytes (16,17).
As regards routes of biological information
transfer, increasing attention has been paid to glycans attached to
proteins and lipids. Notably, sugar-encoded information of
glyco-conjugates is translated into cellular responses by
endogenous lectins (18–20). Members of the family of
adhesion/growth-regulatory galectins are known to be involved in
these responses, and their expression is stringently controlled,
e.g., during differentiation (21–23). Since galectins play an important
regulatory role in cell proliferation, migration and extracellular
matrix formation (24–26) and are expressed in tumors (cell
lines and clinical specimens) as detected by hemagglutination and
purification by affinity chromatography (27,28), it has been postulated that they
are biorelevant modulators of wound/tumor microenvironments
(29). For example, the
extracellular matrix (ECM), which is rich in fibronectin and
galectin-1, serves as an active substratum when feeder cells are
substituted for keratinocytes (26). Galectin-1, a multifunctional
effector in various compartments (30,31), is upregulated during the early
phases of healing (25,32), and is known to have
anti-inflammatory properties (33).
As keratinocytes are known to express estrogen
receptors (ERs) (34), we can
posit that the regeneration of the epidermis may be modulated
through this route. In this context, it has been previously
demonstrated that the administration of exogenous estrogen to
ovariectomized ER-β knockout mice delays wound healing and that the
beneficial effects of ERT are mediated through epidermal ER-β
(35,36). However, little is known about the
underlying mechanisms of estrogen regeneration, in particular the
cell type-specific role of the two nuclear ERs, ER-α and ER-β.
Therefore, in the present study, we aimed to assess the effects of
two ER agonists on the expression of certain protein markers
(Ki-67, keratins-10, -14 and -19, and galectin-1) in HaCaT
kerati-nocytes in an attempt to better understand the mechanisms of
the ER-β-mediated acceleration of re-epithelialization, which has
been previously identified (36).
In addition, the in vivo effects of the selective ER
agonists were investigated using an open wound healing model with
ovariectomized Sprague-Dawley rats.
Materials and methods
Drug preparation
4,4′,4″-(4-propyl [1H]
pyrazole-1,3,5-triyl)-trisphenol (PPT), a selective ER-α agonist
and 2,3-bis(4-hydroxyphenyl)-propionitrile (DPN), a selective ER-β
agonist, were purchased from Tocris Bioscience (Bristol, UK) and
dissolved in dimethyl sulfoxide (DMSO; Sigma-Aldrich, St. Louis,
MO, USA).
Human keratinocyte cell line, HaCaT
Cells of the HaCaT line (37) were obtained from CLS Cell Lines
Service (Eppelheim, Germany). The cells were cultured in Dulbecco's
modified Eagle's medium (DMEM) supplemented with 10% fetal bovine
serum (FBS) and antibiotics (streptomycin and penicillin) (all from
Biochrom, Berlin, Germany). The cells were seeded on coverslips at
a density of 5,000 cells/cm2 and cultured for 24 h. The
ER agonists, PPT and DPN, were added to the medium to reach a final
concentration of 10 nM, as previously described (38), and the cells were then cultured
for 4 days.
HaCaT immunocytochemical analysis
The HaCaT cells were washed with phosphate-buffered
saline (PBS) and fixed in 2% paraformaldehyde (pH 7.2).
Non-specific binding of the secondary antibody was blocked by
pre-incubation with normal swine serum (Dako, Glostrup, Denmark)
diluted in PBS for 30 min. Details of the commercial antibodies
used in the present study are presented in Table I; the anti-galectin-1 antibody was
made in our laboratory, and we tested it to ensure that it was free
of cross-reactivity against human galectins-2, -3, -4, -7, -8 and
-9 by western blot analysis and enzyme-linked immunosorbent assays
(ELISAs), as previously described (39). We controlled antigen-dependent
specificity was by replacing the first-step antibody with an
antibody of the same isotype directed against an antigen not
present in the cells, or omitting the incubation stage with the
antibody. The nuclei of the cells were counterstained with
4′,6-diamidino-2-phenylindole (DAPI; Sigma-Aldrich), which
specifically recognizes DNA.
 | Table ICommercial reagents used in
immunocytochemical analysis. |
Table I
Commercial reagents used in
immunocytochemical analysis.
| Primary
antibody | Abbreviation | Host | Produced by | Secondary
antibody | Produced by | Channel |
|---|
| Ki-67 | Ki-67 | Mouse
monoclonal | DakoCytomation | Goat
anti-mouse | Sigma-Aldrich | TRITC-red |
|
High-molecular-weight keratin | HMWK | Mouse
monoclonal | DakoCytomation | Goat
anti-mouse | Sigma-Aldrich | TRITC-red |
| Keratin-10 | K10 | Mouse
monoclonal | DakoCytomation | Goat
anti-mouse | Sigma-Aldrich | TRITC-red |
| Keratin-14 | K14 | Mouse
monoclonal | Sigma-Aldrich | Goat
anti-mouse | Sigma-Aldrich | TRITC-red |
| Keratin-19 | K19 | Mouse
monoclonal | Dakopatts | Goat
anti-mouse | Sigma-Aldrich | TRITC-red |
| Wide-spectrum
keratin | WSK | Rabbit
polyclonal | Abcam | Swine
anti-rabbit | Santa Cruz
Biotechnology | FITC-green |
| Sox-2 | SOX-2 | Rabbit
polyclonal | Abcam | Swine
anti-rabbit | Santa Cruz
Biotechnology | FITC-green |
| Estrogen
receptor-α | ER-α | Rabbit
polyclonal | Sigma-Aldrich | Swine
anti-rabbit | Santa Cruz
Biotechnology | FITC-green |
| Estrogen
receptor-β | ER-β | Rabbit
polyclonal | Sigma-Aldrich | Swine
anti-rabbit | Santa Cruz
Biotechnology | FITC-green |
RNA isolation, cDNA preparation by
reverse transcription and ER-specific mRNA amplification by
real-time (quantitative) PCR
Total RNA was extracted from the HaCaT cells using
TRIzol reagent (Life Technologies, Woburn, MA, USA), according to
the manufacturer's instructions. RT-PCR was carried out according
to the instructions provided by Qiagen with the One-Step RT-PCR kit
(Qiagen, Hilden, Germany). Briefly, for each sample, 150 ng of
total RNA was added to a solution with RT-PCR buffer,
deoxynucleotide triphosphate (dNTP) mix (10 mM of each dNTP),
primers (10 µM each) and enzyme mix. The following primers were
used: for ER-α detection forward, 5′-GGA GGG CAG GGG TGA A-3′ and
reverse, 5′-GGC CAG GCT GTT CTT CTT CTT AG-3′; for ER-β detection
forward, 5′-AGA GTC CCT GGT GTG AAG CAA-3′ and reverse, 5′-GAC AGC
GCA GAA GTG AGC ATC-3′; and for β-actin detection forward, 5′-ACC
AAC TGG GAC GAC ATG GAG AA-3′ and reverse, 5′-GTA GCC GCG CTC GGT
GAG GAT CT-3′. SYBR-Green Supermix (Bio-Rad iQ™; Bio-Rad
Laboratories, Inc., Hercules, CA, USA) was used with the following
thermal cycling steps: 30 min at 50°C for reverse transcription,
and 5 min at 95°C for initial PCR activation using the LightCycler
Carousel-Based system (Roche, Basel, Switzerland). PCR cycles were
run as follows: 60 cycles of 15 sec at 95°C, 30 sec at 55°C, and 15
sec at 72°C (cooling to 37°C).
Acquisition of microphotographs and
processing of data
Microphotographs of processed samples labeled with
fluorochromes were recorded with identical settings using an
Eclipse 90i fluorescence microscope (Nikon, Tokyo, Japan) equipped
with filterblocks for fluorescein isothiocyanate (FITC),
tetramethylrhodamine (TRITC) and DAPI, and a Cool-1300Q CCD camera
(Vosskühler, Osnabrück, Germany). Data were processed and analyzed
with a LUCIA 5.1 computer-assisted image analysis system
(Laboratory Imaging, Prague, Czech Republic).
Animal model
The experimental conditions complied with European
rules on animal treatment and welfare. Our study was approved by
the Ethics Committee of the Faculty of Medicine of Pavol Jozef
Šafárik University in Košice and by the State Veterinary and Food
Administration of the Slovak Republic.
Female Sprague-Dawley rats (n=20) at 4 months of
age, were used in the present study. The rats were randomly divided
into 4 groups with 5 animals in each group: i) the control group:
sham-operated rats, treated with the vehicle (NOV-C group); ii)
ovariectomized rats treated with the vehicle (OVX-C group); iii)
ovariectomized rats treated with the selective ER-α agonist, PPT
(OVX-PPT group); and iv) ovariectomized rats treated with the
selective ER-β agonist, DPN (OVX-DPN group).
All surgical interventions were performed under
general anesthetic induced by the administration of 33 mg/kg of
ketamine (Narkamon a.u.v.; Spofa a.s., Prague, Czech Republic), 11
mg/kg xylazine (Rometar a.u.v.; Spofa a.s.) and 5 mg/kg tramadol
(Tramadol-K; Krka, Novo Mesto, Slovenia).
Twelve weeks prior to beginning the wound-healing
experiment, as previously described (40), rats from all the OVX groups
underwent ovariectomies, whereas the rats from the control group
were sham-operated.
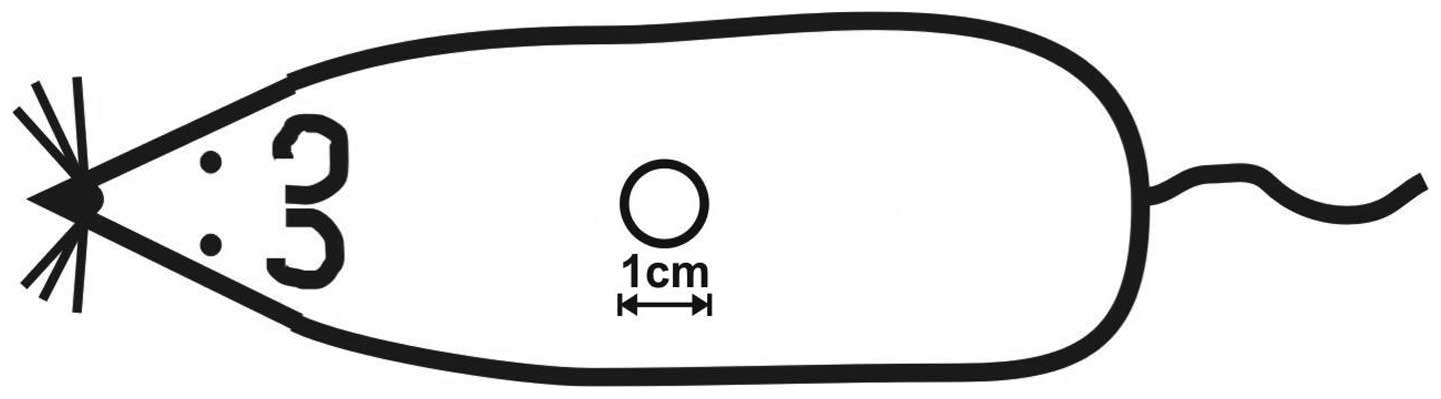
One round full-thickness skin wound, 1 cm in
diameter, was inflicted under aseptic conditions on the back of
each rat (Fig. 1). After
wounding, rats from the OVX-PPT and OVX-DPN groups were treated
daily (during the first 7 days after surgery) with 1 mg/kg of PPT
and DPN subcutaneously, while the other rats received the vehicle
(1% DMSO), as previously described (41,42). On day 21 post-surgery, 5 animals
from each group were sacrificed by ether inhalation, and the wound
tissues were removed for further processing.
Histological analysis of skin wounds and
semi-quantitative analysis of histological sections
Tissue specimens were processed routinely for light
microscopy [fixed in 4% buffered formaldehyde, dehydrated, embedded
in paraffin, sectioned and stained with hematoxylin and eosin
(H&E)]. The stained sections were evaluated in a blinded manner
(without knowing which section belonged with which rat group) using
an Olympus BX51 microscope equipped with a DP50 CCD camera
(Olympus, Tokyo, Japan).
A semi-quantitative method, which has been
previously described (43), was
used to monitor the re-epithelialization of the epidermis and the
presence of inflammatory cells [polymorphonuclear leukocytes
(PMNLs), fibroblasts, vessels and new collagen]. The sections were
evaluated in a blinded manner according on a scale of 0 to 4
(Table II).
| Table IIDefinition of scale in the
semi-quantitative evaluation of the histological sections. |
Table II
Definition of scale in the
semi-quantitative evaluation of the histological sections.
| Scale | Epithelization | PMNL | Fibroblasts | Luminized
vessels |
|---|
| 0 | Thickness of cut
edges | Absent | Absent | Absent |
| 1 | Migration of cells
(<50%) | Mild ST | Mild ST | Mild SCT |
| 2 | Migration of cells
(≥50%) | Mild DL/GT | Mild GT | Mild GT |
| 3 | Bridging the
excision | Moderate DL/GT | Moderate GT | Moderate GT |
| 4 | Keratinization | Marked DL/GT | Marked GT | Marked GT |
Statistical analysis
One-way analysis of variance (ANOVA) followed by the
Tukey-Kramer post hoc test was used to compare the differences in
the number (percentages) of Ki-67-, keratin-10-, keratin-14-,
keratin-19- and galectin-1-positive cells (data are presented as
the means ± standard deviation). Data from the semi-quantitative
analysis are presented as median and were compared using the
Kruskal-Wallis non-parametric test. A P-value <0.05 was
considered to indicate a statistically significant difference.
Results
ER detection in HaCaT cells
We noted that the HaCaT keratinocytes expressed both
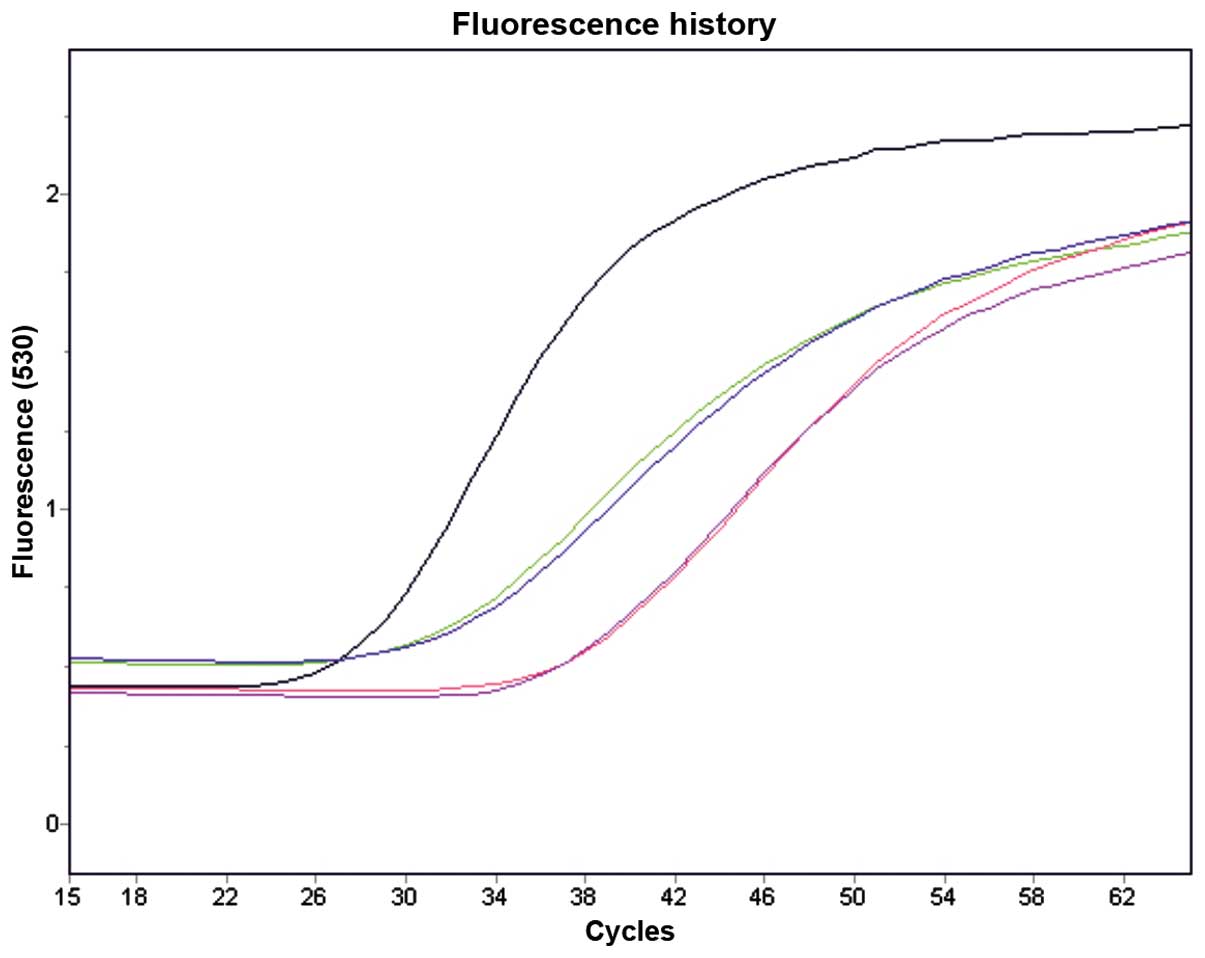
ER-α and -β, mainly in the cell nuclei (Fig. 2a). ERs were detected by RT-qPCR,
and the gene transcription of the ER-β receptor was slightly higher
when compared to that of the ER-α receptor (Fig. 3). Cytochemically, in comparison to
the control and PPT-treated cells, treatment with the ER-β agonist
(DPN) increased the percentage of Ki-67-positive cells (Fig. 2b, panels A1-A3). It should be
noted that targeting ER-β abolished galectin-1 expression, whereas
the control and ER-α agonist-treated cells were positive for
galectin-1 (Fig. 2b, panesl
E1-E3).
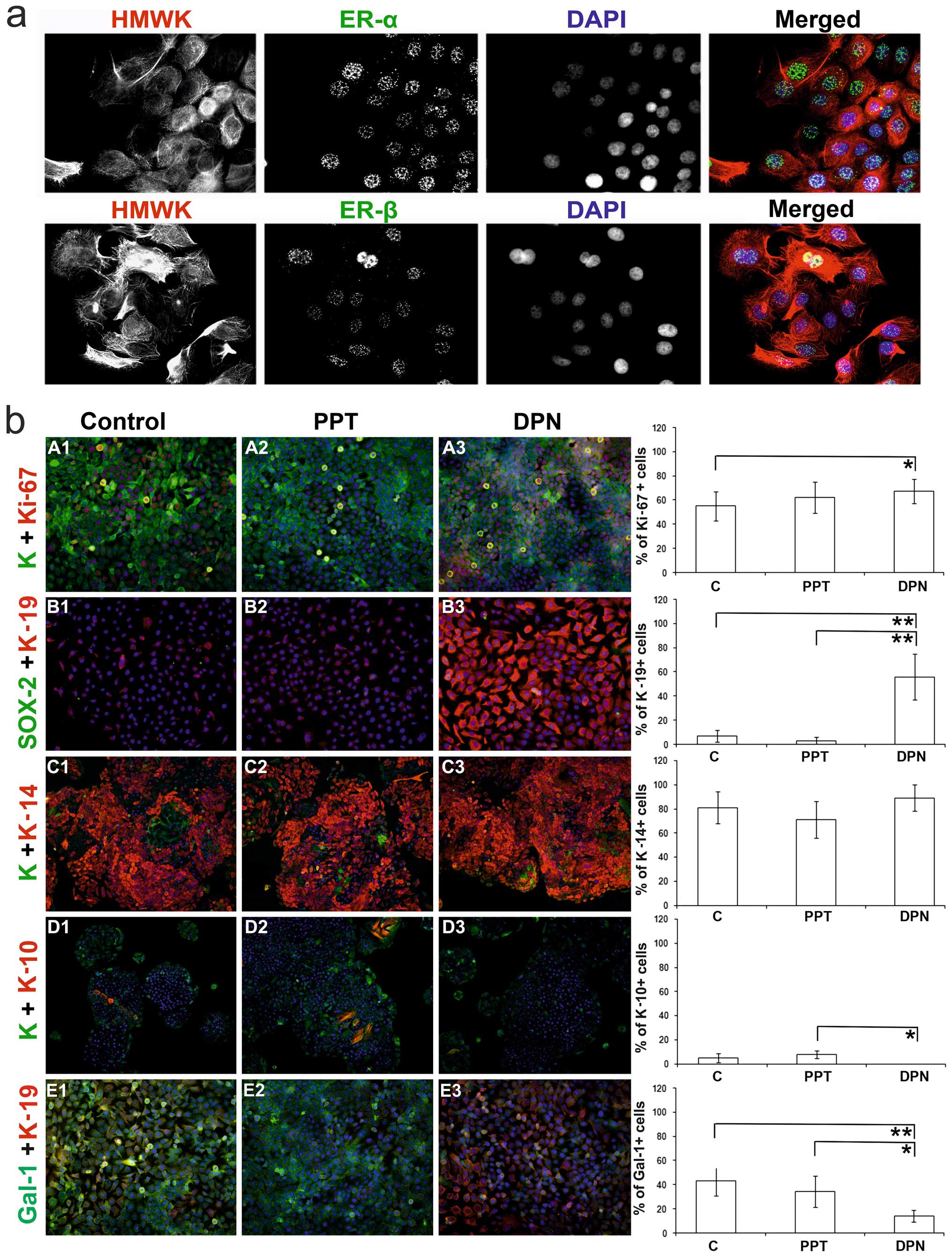 | Figure 2(a) Culture of HaCaT keratinocytes
(magnification, ×600), which express both estrogen receptor (ER)-α
and -β mainly in the cell nuclei; cytoskeleton is stained with
high-molecular-weight keratins (HMWK), cell nuclei are visualized
by DAPI. (b) HaCaT keratinocytes cultured under the influence of
selective ER agonists; first line of horizontal panels (A1-A3,
magnification, ×200): detection of the proliferation marker Ki-67
and wide-spectrum keratin; second horizontal panel (B1-B3,
magnification, ×200): presence of keratin-19 (marker of poorly
differentiated keratinocytes with stem-like phenotype) and Sox-2
(stem cels marker); third horizontal panel (C1-C3, magnification,
×100): positivity for keratin-14 (marker of poorly differentiated
keratinocytes) and wide-spectrum keratin; fourth horizontal panel
(D1-D3, magnification, ×100): expression of keratin-10 (marker of
differentiated keratinocytes) and wide-spectrum keratin; fifth
horizontal panel (E1-E3, magnification, ×100): presence of
keratin-19 and galectin-1 (Gal-1). PPT-, 4,4′,4″-(4-propyl [1H]
pyrazole-1,3,5-triyl)-trisphenol, a selective ER-α agonist; DPN,
2,3-bis(4-hydroxyphenyl)-propionitrile, a selective ER-β agonist;
*P<0.05 and **P<0.01. C, control; K,
wide-spectrum keratin. |
The majority (81±13%) of the HaCaT control cells
expressed keratin-14 (Fig. 2b,
panels C1-C3). Only a small percentage of the control cells
expressed keratin-10 (5±4%; Fig.
2b, panels D1-D3) and keratin-19 (7±5%; Fig. 2b, panels B1-B3). The cells
stimulated with the ER-α agonist (PPT) had similar percentages of
keratin-based phenotypes (K10, 8±3%; K14, 71±15%; K19, 3±3%;
Fig. 2b, panels B2, C2 and D2)
compared to the untreated controls (Fig. 2b, panels B1, C1 and D1). The cells
stimulated with the ER-β agonist (DPN) had less differentiated
phenotypes, with a marked positivity of keratin-19 (64±19%) and the
absence of keratin-10 (0±0%) (Fig.
2b, panels B3, C3 and D3). Of note, the level of keratin-14
following treatment with DPN remained relatively consistent (K14,
89±11%). In all the groups, no effects on the expression level of
Sox-2 (0±0%) were observed (Fig.
2b, panels B1-B3).
Skin wounds
During the post-surgical period, all animals
remained healthy and did not exhibit any clinical symptoms of
infection. Of note, the inflammatory phase passed in all groups
with no presence of PMNLs noted and only very minor occurrences of
tissue macrophages at the sites of injury. The results of
semi-quantitative analysis of the histological sections are
summarized in Table III.
| Table IIISemi-quantitative analysis of
histological structures/changes 21 days post-surgery (data are
presented as the median). |
Table III
Semi-quantitative analysis of
histological structures/changes 21 days post-surgery (data are
presented as the median).
| Group |
Epithelialization | PMNLs | Fibroblasts | Luminized
vessels |
|---|
| NOV-C | 4 | 0 | 3 | 2 |
| OVX-C | 4 | 0 | 3 | 2 |
| OVX-PPT | 4 | 0 | 3 | 3 |
| OVX-DPN | 4 | 0 | 3 | 3 |
On day 21 after wounding, a thin keratin layer was
present in all wounds (Fig. 4),
demonstrating that a normal keratinocyte differentiation had
occurred. However, differences were noted in the process of
epidermal regeneration. In the rats from the OVX-C and OVX-PPT
groups, we noted that hair follicle regeneration and epidermal
thickening were both delayed (Fig.
4). Treatment with the ER-β agonist (Fig. 4; OVX-DPN) resulted in a normalized
process of epidermal regeneration, comparable to that of the
sham-operated animals (NOV-C). In comparison to both control groups
(OVX-C and NOV-C), the number of luminized vessels slightly
increased upon treatment with the estrogen agonists treatment
(OVX-PPT and OVX-DPN). A moderate number of fibroblasts in the
granulation tissues of all wounds was observed, reflecting the
progression of in tissue fibrosis. Of note, no significant
differences were observed between the groups in terms of the
presence of luminized vessels or of fibroblasts.
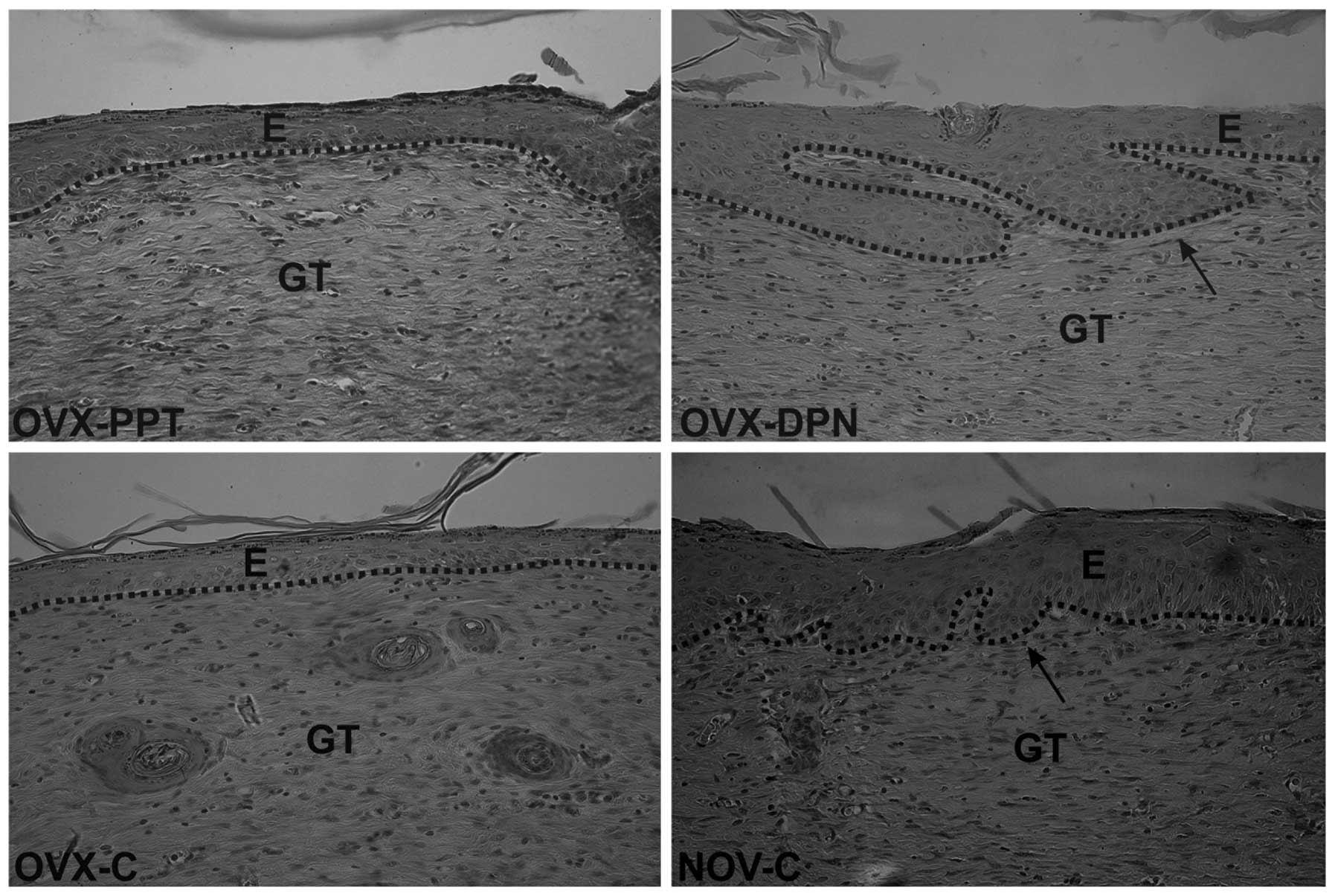 | Figure 4Healing of skin wounds 21 days
post-surgery (magnification, ×400). The maturation phase of healing
was noted in all groups, and differences were observed in epidermal
regeneration, which was impaired in the ovariectomized rats treated
with the vehicle (1% DMSO; OVX-C) and ovariectomized rats treated
with the ER-α agonist, PPT (OVX-PPT. Black arrows indicate growing
hair follicles in sham-operated vehicle-treated rats (NOV-C) and
ovariectomized rats treated with the ER-β agonist, DPN (OVX-DPN).
The dotted line distinguishes the epidermis from the granulation
tissue; staining was done with H&E. E, epidermis; GT,
granulation tissue; PPT-, 4,4′,4″-(4-propyl [1H]
pyrazole-1,3,5-triyl)-trisphenol, a selective ER-α agonist; DPN,
2,3-bis(4-hydroxyphenyl)-propionitrile, a selective ER-β
agonist. |
Discussion
Monitoring markers for differentiation revealed that
the ER-β agonist, DPN, decreases the expression of galectin-1 in
HaCaT cells. The expression of this lectin is known to be
sterol-sensitive (please see below), and this observation provides
direction to further assessments of its impact on other members of
the galectin network and also on glycosylation, making cells
susceptible to galectins. These lectins can have site-specific
additive or antagonistic effects, and when co-expressed they form a
network, as in tumors (44–46). Therefore, their regulation may
alter the clinical course of a tumor, and in this context, the
phytoestrogen, genistein, and its potential chemopreventive effects
on breast cancer also deserve attention (47,48). Following initial
immunohistochemical detection of this class of tumors, galectin-1
has been shown to be upregulated in invasive breast carcinoma with
a positive correlation with the TNM staging system (49,50). Fittingly, as previously
demonstrated, the silencing of galectin-1 in a breast carcinoma
model overcame breast cancer-associated immunosuppression,
inhibited tumor growth and prevented metastatic disease (51). Of note, in our previous studies,
we found that galectin-1 was upregulated during the early phases of
wound healing (25,32). Since long-term estradiol
deprivation enhances estrogen sensitivity (52), by upregulating ER-α expression,
further studies focusing on this receptor are warranted in order to
reduce tumor cell proliferation (53). Antagonizing ER-α, together with
agonizing ER-β, may ameliorate galectin-1-induced immunosuppression
in breast cancer; however, further consideration based on detailed
network and glycosylation studies is necessary.
Although estrogen is considered a key regulator of
wound healing, an incomplete understanding of the molecular
mechanisms of action of estrogen, as well as the well-documented
adverse effects of estrogens during menopause in clinical trials,
preclude the common clinical use of ERT as a wound-healing
treatment. One example of the negative effects of estrogens is that
the activation of ER-α leads to a decrease in the tensile strength
of wounds during the proliferation phase of healing (41,54), whereas the activation of ER-α
and/or -β significantly increases this parameter during the early
maturation phase (54). The ER-α
agonist, TGF-β1-dependently increases fibroblast migration and
keratinocyte proliferation. By contrast, the ER-β agonist does not
affect cell migration (55,56) and increases keratinocyte
proliferation in a TGF-β1-independent manner (57). Accordingly, in vivo
experiments have shown that targeting ER-β, but not ER-α leads to
accelerated re-epithelialization in mice (36) and rats (54). Furthermore, ER-α has been proven
to be responsible for impaired wound healing in male mice (58). In the present study, we
demonstrated that the pharmacological activation of ER-β, but not
that of ER-α, led to a significant alteration in the pattern of
differentiation and the proliferation activity of keratinocytes. In
relation to markers, previous research has demonstrated that the
ER-β agonist does not induce Sox-2 expression, a characteristic of
stem-like properties (59), in
keratin-19-positive cells.
In conclusion, our data suggest that marker-based
cytochemical monitoring provides new information on ER-modulated
keratinocyte differentiation and proliferation. The stimulation of
epidermal regeneration may ensue after treating wounds with an ER-β
agonist. In order to activate the TGF-β1 pathway to this end, ER-α
should be targeted (57).
However, the nature of the animal model and restrictions on
extrapolations must be taken into consideration, as we did in our
study, by combining a human in vitro model with in
vivo data on rats.
Acknowledgments
We would like to thank Iva Burdová, Ladislava
Kratinová and Marcela Blažeková for providing technical assistance.
The present study was supported in part by the Grant Agency of
Ministry of the Education, Science, Research and Sport of the
Slovak Republic (VEGA 1/0299/13, VEGA 1/0404/15, and VEGA
1/0048/15), the Agency for Science and Research under the contract
no. APVV-0408-12 and APVV-14-0731, the Charles University in Prague
(project for support of specific university student research,
project UNCE 204013 and project PRVOUK 27), as well as by the
project BIOCEV (Biotechnology and Biomedicine Centre of the Academy
of Sciences and Charles University in Vestec -
CZ.1.05/1.1.00/02.0109, from the European Regional Development
Fund) and by EC funding for GlycoHIT (contract no. 260600) and the
Marie Curie ITN network GLYCOPHARM (contract no. 317297).
References
|
1
|
Calleja-Agius J and Brincat M: The effect
of menopause on the skin and other connective tissues. Gynecol
Endocrinol. 28:273–277. 2012. View Article : Google Scholar
|
|
2
|
Hall G and Phillips TJ: Estrogen and skin:
the effects of estrogen, menopause, and hormone replacement therapy
on the skin. J Am Acad Dermatol. 53:555–568; quiz 569–572. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kloepper JE, Tiede S, Brinckmann J,
Reinhardt DP, Meyer W, Faessler R and Paus R: Immunophenotyping of
the human bulge region: the quest to define useful in situ markers
for human epithelial hair follicle stem cells and their niche. Exp
Dermatol. 17:592–609. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Archer DF: Postmenopausal skin and
estrogen. Gynecol Endocrinol. 28(Suppl 2): 2–6. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ashcroft GS, Mills SJ, Lei K, Gibbons L,
Jeong MJ, Taniguchi M, Burow M, Horan MA, Wahl SM and Nakayama T:
Estrogen modulates cutaneous wound healing by downregulating
macrophage migration inhibitory factor. J Clin Invest.
111:1309–1318. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gilliver SC, Emmerson E, Bernhagen J and
Hardman MJ: MIF: a key player in cutaneous biology and wound
healing. Exp Dermatol. 20:1–6. 2011. View Article : Google Scholar
|
|
7
|
Ashcroft GS, Dodsworth J, van Boxtel E,
Tarnuzzer RW, Horan MA, Schultz GS and Ferguson MW: Estrogen
accelerates cutaneous wound healing associated with an increase in
TGF-beta1 levels. Nat Med. 3:1209–1215. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li X, Bai J, Ji X, Li R, Xuan Y and Wang
Y: Comprehensive char-acterization of four different populations of
human mesenchymal stem cells as regards their immune properties,
proliferation and differentiation. Int J Mol Med. 34:695–704.
2014.PubMed/NCBI
|
|
9
|
Zhu T, Park HC, Son KM, Kwon JH, Park JC
and Yang HC: Effects of thymosin β4 on wound healing of rat palatal
mucosa. Int J Mol Med. 34:816–821. 2014.PubMed/NCBI
|
|
10
|
Liu H, Du L, Wen Z, Yang Y, Li J, Dong Z,
Zheng G, Wang L, Zhang X and Wang C: Sex determining region Y-box 2
inhibits the proliferation of colorectal adenocarcinoma cells
through the mTOR signaling pathway. Int J Mol Med. 32:59–66.
2013.PubMed/NCBI
|
|
11
|
Törmä H, Lindberg M and Berne B: Skin
barrier disruption by sodium lauryl sulfate-exposure alters the
expressions of involucrin, transglutaminase 1, profilaggrin, and
kallikreins during the repair phase in human skin in vivo. J Invest
Dermatol. 128:1212–1219. 2008. View Article : Google Scholar
|
|
12
|
Freedberg IM, Tomic-Canic M, Komine M and
Blumenberg M: Keratins and the keratinocyte activation cycle. J
Invest Dermatol. 116:633–640. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Reichelt J, Büssow H, Grund C and Magin
TM: Formation of a normal epidermis supported by increased
stability of keratins 5 and 14 in keratin 10 null mice. Mol Biol
Cell. 12:1557–1568. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Carter CA, Jolly DG, Worden CE Sr, Hendren
DG and Kane CJ: Platelet-rich plasma gel promotes differentiation
and regeneration during equine wound healing. Exp Mol Pathol.
74:244–255. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Peryassu MA, Cotta-Pereira G,
Ramos-e-Silva M and Filgueira AL: Expression of keratins 14, 10 and
16 in marginal keratoderma of the palms. Acta Dermatovenerol Croat.
13:206–211. 2005.PubMed/NCBI
|
|
16
|
Michel M, Török N, Godbout MJ, Lussier M,
Gaudreau P, Royal A and Germain L: Keratin 19 as a biochemical
marker of skin stem cells in vivo and in vitro: keratin 19
expressing cells are differentially localized in function of
anatomic sites, and their number varies with donor age and culture
stage. J Cell Sci. 109:1017–1028. 1996.PubMed/NCBI
|
|
17
|
Dvoránková B, Smetana K Jr, Chovanec M,
Lacina L, Stork J, Plzáková Z, Galovicová M and Gabius HJ:
Transient expression of keratin 19 is induced in originally
negative interfollicular epidermal cells by adhesion of suspended
cells. Int J Mol Med. 16:525–531. 2005.PubMed/NCBI
|
|
18
|
Gabius HJ, André S, Jiménez-Barbero J,
Romero A and Solís D: From lectin structure to functional
glycomics: principles of the sugar code. Trends Biochem Sci.
36:298–313. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
André S, Kaltner H, Manning JC, Murphy PV
and Gabius HJ: Lectins: getting familiar with translators of the
sugar code. Molecules. 20:1788–1823. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Solís D, Bovin NV, Davis AP,
Jiménez-Barbero J, Romero A, Roy R, Smetana K Jr and Gabius HJ: A
guide into glycosciences: How chemistry, biochemistry and biology
cooperate to crack the sugar code. Biochim Biophys Acta.
1850:186–235. 2015. View Article : Google Scholar
|
|
21
|
Villalobo A, Nogales-Gonzalez A and Gabius
HJ: A guide to signaling pathways connecting protein-glycan
interaction with the emerging versatile effector functionality of
mammalian lectins. Trends Glycosci Glyc. 18:1–37. 2006. View Article : Google Scholar
|
|
22
|
Kaltner H and Gabius HJ: A toolbox of
lectins for translating the sugar code: the galectin network in
phylogenesis and tumors. Histol Histopathol. 27:397–416.
2012.PubMed/NCBI
|
|
23
|
Katzenmaier EM, André S, Kopitz J and
Gabius HJ: Impact of sodium butyrate on the network of
adhesion/growth-regulatory galectins in human colon cancer in
vitro. Anticancer Res. 34:5429–5438. 2014.PubMed/NCBI
|
|
24
|
Nagy N, Bronckart Y, Camby I, Legendre H,
Lahm H, Kaltner H, Hadari Y, Van Ham P, Yeaton P, Pector JC, et al:
Galectin-8 expression decreases in cancer compared with normal and
dysplastic human colon tissue and acts significantly on human colon
cancer cell migration as a suppressor. Gut. 50:392–401. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Klíma J, Lacina L, Dvoránková B, Herrmann
D, Carnwath JW, Niemann H, Kaltner H, André S, Motlík J, Gabius HJ
and Smetana K Jr: Differential regulation of galectin
expression/reactivity during wound healing in porcine skin and in
cultures of epidermal cells with functional impact on migration.
Physiol Res. 58:873–884. 2009.
|
|
26
|
Dvořánková B, Szabo P, Lacina L, Gal P,
Uhrova J, Zima T, Kaltner H, André S, Gabius HJ, Sykova E, et al:
Human galectins induce conversion of dermal fibroblasts into
myofibroblasts and production of extracellular matrix: potential
application in tissue engineering and wound repair. Cells Tissues
Organs. 194:469–480. 2011. View Article : Google Scholar
|
|
27
|
Teichberg VI, Silman I, Beitsch DD and
Resheff G: A beta-D-galactoside binding protein from electric organ
tissue of Electrophorus electricus. Proc Natl Acad Sci USA.
72:1383–1387. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gabius HJ, Engelhardt R, Cramer F, Bätge R
and Nagel GA: Pattern of endogenous lectins in a human epithelial
tumor. Cancer Res. 45:253–257. 1985.PubMed/NCBI
|
|
29
|
Smetana K Jr, Szabo P, Gál P, André S,
Gabius HJ, Kodet O and Dvořánková B: Emerging role of tissue
lectins as microenviron-mental effectors in tumors and wounds.
Histol Histopathol. 30:293–309. 2015.
|
|
30
|
Smetana K Jr, André S, Kaltner H, Kopitz J
and Gabius HJ: Context-dependent multifunctionality of galectin-1:
a challenge for defining the lectin as therapeutic target. Expert
Opin Ther Targets. 17:379–392. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang S, Moussodia RO, Murzeau C, Sun HJ,
Klein ML, Vértesy S, André S, Roy R, Gabius HJ and Percec V:
Dissecting molecular aspects of cell interactions using
glycodendrimer-somes with programmable glycan presentation and
engineered human lectins. Angew Chem Int Ed Engl. 54:4036–4040.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gál P, Vasilenko T, Kostelníková M,
Jakubco J, Kovác I, Sabol F, André S, Kaltner H, Gabius HJ and
Smetana K Jr: Open wound healing in vivo: Monitoring binding and
presence of adhesion/growth-regulatory galectins in rat skin during
the course of complete re-epithelialization. Acta Histochem
Cytochem. 44:191–199. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cooper D, Norling LV and Perretti M: Novel
insights into the inhibitory effects of Galectin-1 on neutrophil
recruitment under flow. J Leukoc Biol. 83:1459–1466. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Emmerson E and Hardman MJ: The role of
estrogen deficiency in skin ageing and wound healing.
Biogerontology. 13:3–20. 2012. View Article : Google Scholar
|
|
35
|
Krahn-Bertil E, Dos Santos M, Damour O,
Andre V and Bolzinger MA: Expression of estrogen-related receptor
beta (ERRβ) in human skin. Eur J Dermatol. 20:719–723.
2010.PubMed/NCBI
|
|
36
|
Campbell L, Emmerson E, Davies F, Gilliver
SC, Krust A, Chambon P, Ashcroft GS and Hardman MJ: Estrogen
promotes cutaneous wound healing via estrogen receptor β
independent of its antiinflammatory activities. J Exp Med.
207:1825–1833. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Boukamp P, Petrussevska RT, Breitkreutz D,
Hornung J, Markham A and Fusenig NE: Normal keratinization in a
spontaneously immortalized aneuploid human keratinocyte cell line.
J Cell Biol. 106:761–771. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Guo J, Duckles SP, Weiss JH, Li X and
Krause DN: 17β-Estradiol prevents cell death and mitochondrial
dysfunction by an estrogen receptor-dependent mechanism in
astrocytes after oxygen-glucose deprivation/reperfusion. Free Radic
Biol Med. 52:2151–2160. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kaltner H, Seyrek K, Heck A, Sinowatz F
and Gabius HJ: Galectin-1 and galectin-3 in fetal development of
bovine respiratory and digestive tracts. Comparison of cell
type-specific expression profiles and subcellular localization.
Cell Tissue Res. 307:35–46. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gál P, Kilík R, Mokrý M, Vidinský B,
Vasilenko T, Mozeš S, Bobrov N, Tomori Z, Bober J and Lenhardt L:
Simple method of open skin wound healing model in
corticosteroid-treated and diabetic rats: standardization of
semi-quantitative and quantitative histological assessments. Vet
Med. 53:652–659. 2008.
|
|
41
|
Gál P, Novotný M, Vasilenko T, Depta F,
Šulla I and Tomori Z: Decrease in wound tensile strength following
post-surgical estrogen replacement therapy in ovariectomized rats
during the early phase of healing is mediated via ER-alpha rather
than ER-beta: a preliminary report. J Surg Res. 159:e25–e28. 2010.
View Article : Google Scholar
|
|
42
|
Wegorzewska IN, Walters K, Weiser MJ,
Cruthirds DF, Ewell E, Larco DO, Handa RJ and Wu TJ:
Postovariectomy weight gain in female rats is reversed by estrogen
receptor alpha agonist, propylpyrazoletriol. Am J Obstet Gynecol.
199:67.e1–67.e5. 2008. View Article : Google Scholar
|
|
43
|
Gál P, Toporcer T, Vidinský B, Mokrý M,
Grendel T, Novotný M, Sokolský J, Bobrov N, Toporcerová S, Sabo J
and Mozes S: Postsurgical administration of estradiol benzoate
decreases tensile strength of healing skin wounds in ovariectomized
rats. J Surg Res. 147:117–122. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sanchez-Ruderisch H, Fischer C, Detjen KM,
Welzel M, Wimmel A, Manning JC, André S and Gabius HJ: Tumor
suppressor p16 INK4a: Downregulation of galectin-3, an endogenous
competitor of the pro-anoikis effector galectin-1, in a pancreatic
carcinoma model. FEBS J. 277:3552–3563. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Amano M, Eriksson H, Manning JC, Detjen
KM, André S, Nishimura S, Lehtiö J and Gabius HJ: Tumour suppressor
p16(INK4a) - anoikis-favouring decrease in N/O-glycan/cell surface
sialylation by down-regulation of enzymes in sialic acid
biosynthesis in tandem in a pancreatic carcinoma model. FEBS J.
279:4062–4080. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Dawson H, André S, Karamitopoulou E,
Zlobec I and Gabius HJ: The growing galectin network in colon
cancer and clinical relevance of cytoplasmic galectin-3 reactivity.
Anticancer Res. 33:3053–3059. 2013.PubMed/NCBI
|
|
47
|
Shon YH, Park SD and Nam KS: Effective
chemopreventive activity of genistein against human breast cancer
cells. J Biochem Mol Biol. 39:448–451. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
DeSantis C, Ma J, Bryan L and Jemal A:
Breast cancer statistics, 2013. CA Cancer J Clin. 64:52–62. 2014.
View Article : Google Scholar
|
|
49
|
Gabius HJ, Brehler R, Schauer A and Cramer
F: Localization of endogenous lectins in normal human breast,
benign breast lesions and mammary carcinomas. Virchows Arch B Cell
Pathol Incl Mol Pathol. 52:107–115. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Jung EJ, Moon HG, Cho BI, Jeong CY, Joo
YT, Lee YJ, Hong SC, Choi SK, Ha WS, Kim JW, et al: Galectin-1
expression in cancer-associated stromal cells correlates tumor
invasiveness and tumor progression in breast cancer. Int J Cancer.
120:2331–2338. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Dalotto-Moreno T, Croci DO, Cerliani JP,
Martinez-Allo VC, Dergan-Dylon S, Méndez-Huergo SP, Stupirski JC,
Mazal D, Osinaga E, Toscano MA, et al: Targeting galectin-1
overcomes breast cancer-associated immunosuppression and prevents
metastatic disease. Cancer Res. 73:1107–1117. 2013. View Article : Google Scholar
|
|
52
|
Santen RJ, Song RX, Zhang Z, Kumar R, Jeng
MH, Masamura A, Lawrence J Jr, Berstein L and Yue W: Long-term
estradiol deprivation in breast cancer cells up-regulates growth
factor signaling and enhances estrogen sensitivity. Endocr Relat
Cancer. 12(Suppl 1): S61–S73. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Anbalagan M and Rowan BG: Estrogen
receptor alpha phosphorylation and its functional impact in human
breast cancer. Mol Cell Endocrinol. Jan 15–2015.Epub ahead of
print. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Novotný M, Vasilenko T, Varinská L,
Smetana K Jr, Szabo P, Sarišský M, Dvořánková B, Mojžiš J, Bobrov
N, Toporcerová S, et al: ER-α agonist induces conversion of
fibroblasts into myofibroblasts, while ER-β agonist increases ECM
production and wound tensile strength of healing skin wounds in
ovariectomised rats. Exp Dermatol. 20:703–708. 2011. View Article : Google Scholar
|
|
55
|
Stevenson S, Nelson LD, Sharpe DT and
Thornton MJ: 17beta-estradiol regulates the secretion of TGF-beta
by cultured human dermal fibroblasts. J Biomater Sci Polym Ed. 19.
pp. 1097–1109. 2008, View Article : Google Scholar
|
|
56
|
Stevenson S, Sharpe DT and Thornton MJ:
Effects of oestrogen agonists on human dermal fibroblasts in an in
vitro wounding assay. Exp Dermatol. 18:988–990. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Merlo S, Frasca G, Canonico PL and Sortino
MA: Differential involvement of estrogen receptor alpha and
estrogen receptor beta in the healing promoting effect of estrogen
in human keratinocytes. J Endocrinol. 200:189–197. 2009. View Article : Google Scholar
|
|
58
|
Gilliver SC, Emmerson E, Campbell L,
Chambon P, Hardman MJ and Ashcroft GS: 17beta-estradiol inhibits
wound healing in male mice via estrogen receptor-alpha. Am J
Pathol. 176:2707–2721. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Grinnell KL and Bickenbach JR: Skin
keratinocytes pre-treated with embryonic stem cell-conditioned
medium or BMP4 can be directed to an alternative cell lineage. Cell
Prolif. 40:685–705. 2007. View Article : Google Scholar : PubMed/NCBI
|