1. Introduction
The α-Gal epitope (Galα1,3Galα1,4GlcNAc-R) is a
unique carbohydrate, which is naturally produced on glycolipids and
glycoprotein (1). Based on the
carbohydrate chains on glycol-conjugates, α-Gal epitope synthesis
is different in various species (2). In rabbits and cows, it is abundantly
expressed on red cell membrane glycolipids, on cell surface
glycoproteins of mouse Ehrlich ascites cells, lymphoma cells, and
on thyroglobulin of bovine, canine and porcine (3). In addition, the glycosylation enzyme
α1,3 galactosyltransferases (α1,3GT) can transfer galactose from
uridine diphosphate (UDP)-gal to N-acetyllactosamine, producing
oligosaccha-ride Galα1,3Galα1,4GlcNAc-R (termed α-Gal epitope)
(4). However, the α1,3GT was once
thought to be a single α1,3GT that exclusively synthesized the
important xenoepitope Gal(1,3)Gal. However, the iGb3 synthase and
α1,3GT in rats demonstrated two separate glycosylation pathways to
the synthesis of Gal(1,3)Gal, challenging what is known
regarding α1,3GT (5,6). As a matter of fact, the enzyme
α1,3GT is as universal as the α1,3 galactosyltransferase, and it is
encoded by the α1,3GT gene. In different species, the α1,3GT gene
is on a different locus, for example it resides on chromosome 2 of
Mus musculus (house mouse) (7),
chromosome 1 of Sus scrofa (pig) (8), chromosome 11 of Bos taurus
(cattle) (9), chromosome 9 of
Canis lupus familiaris (dog) (10), and chromosome 9 of the Homo
sapiens (human) pseudogene, as the specified gene sites for the
locus of the α1,3GT gene (11).
Comparing with the nucleotide sequence of the human α1,3GT
pseudogene with the corresponding different species sequences, and
considering the evolutionary tree of different species,
inactivation of α1,3GT genes in ancestral primates is caused by
several deletions on DNA sequences, which generates premature stop
codons and the truncation of the enzyme molecule (12). The expression of the α-Gal epitope
and the activity of α1,3GT demonstrate a striking difference
regarding their distribution in various species. Therefore,
although the α-Gal epitope is absent in humans, apes and Old World
monkeys, it is profusely generated in non-primate mammals,
prosimians and New World monkeys (13). A large quantity of the natural
anti-Gal antibody is produced in all humans. Since humans and Old
World primates lack the α-Gal epitope, they are not immunotolerant
to it, and therefore will produce anti-Gal antibodies (14,15). The anti-Gal antibody in humans is
encoded by several heavy-chain genes primarily of the VH3
immunoglobulin gene family (16).
Xenotransplantation is the transplantation from
animals, such as pigs, to humans. The α-Gal epitope on the
xeno-grafts will be specifically bound by the anti-Gal antibody,
and therefore the combination of the anti-Gal antibody with α-Gal
epitope contributes to the complement cascade (17,18). In detail, the complement cascade
would lead to the collapse of the xenograft vascular bed and
hyperacute rejection, which is the major obstacle in
xenotransplantation. In order to overcome xenografts rejection,
α1,3GT knockout mice (α1,3GT KO), lacking the ability to synthesize
α-Gal epitope, were generated, and immunotolerence of
xenotransplantation was induced (19). According to the success of α1,3GT
KO mouse experiment, the pig, as the major xenografts donor to
humans, has been applied to the same experiment. Currently, the
α1,3GT KO pigs, which proved to possess no hyperacute rejection,
are used in organs transplantation (20). In addition, regarding its
application in xenotransplantation, the α-Gal epitope can also be
used to increase the immunogenicity of the tumor cells, and it can
be developed for clinical use in cancer immunotherapy as well.
Furthermore, the immunogenicity of certain viral vaccines (21), including the flu vaccine (used in
the elderly), and human immunodeficiency virus (HIV) vaccine, is
deemed as suboptimal. The α-Gal epitope nanoparticles bind with the
anti-Gal, which will activate the complement system and will
recruit macrophages to induce tissue regeneration (22). Therefore, the application of α-gal
nanoparticles could accelerate wound healing (23,24). This therapy may be of further
significance in the regeneration of injured tissues, such as
ischemic myocardium and injured nerves.
2. α-Gal epitope and anti-Gal antibody
α-Gal epitope
The α-Gal epitope has a special terminal
carbohydrate structure in the form of Gala1, 3Galb1-4GlcNAc-R,
which is confirmed by the study of two structures of the major
glycolipids in rabbit red cell membranes: Ceramide trihexoside
(Galα1-4Galα1-4Glc-Cer) and ceramide pentahexoside
(Galα1-3Gaα1-4GlcNAcα1-3Galα1-4Glc-Cer) (3). The amount of α-Gal epitope exhibits
significant differences in different organs. High concentrations of
the α-Gal epitope were observed in all the hepatic cells. The
largest quantity of α-Gal epitope was observed in the proximal
convoluted tubules in the kidney and the α-Gal epitope were
presented in smaller quantities in the distal tubules and in the
glomeruli. No α-Gal epitope was identified in the collecting ducts.
In addition, the α-Gal epitope is expressed on numerous molecules
on platelets and it is predominantly found on fibrinogen, von
Willebrand's factor, α2 integrin and β3 integrin. On endothelial
cells, >20 glycoproteins carry α-Gal epitope, some of which have
been identified as von Willebrand's factor, DM-GRASP, and the α1,
αv, α3/α5, β1, β3 integrins (25). The expression of the α-Gal epitope
varies in different species; for example, it is expressed on the
red cell membrane glycolipids of rabbits and cows, the cell surface
glycoproteins of mouse Ehrlich ascites cells, lymphoma cells and
fibroblasts, on mouse laminin and on bovine, canine and porcine
thyroglobulin.
The α-Gal epitope is synthesized by the transferase
α1,3GT, and similar to numerous other glycosylating enzymes, the
α1,3GT resides within the Golgi of the cell (5). The α1,3GT is believed to be the only
enzyme to synthesize Gal(1,3)Gal. However, the rat iGb3 synthase is
also capable of synthesizing Gal(1,3)Gal
as the glycolipid structure iGb3, showing that the rat expresses
two distinct α1,3GT (26), α1,3GT
and iGb3 synthase. Therefore, in rats there exists two separate
glycosylation pathways to synthesize Gal(1,3)Gal. The α1,3GT is one of the α1,3GTs,
and this enzyme is active in the Golgi apparatus of cells and
transfers galactose from the sugar-donor UDP-gal to the acceptor
N-acetyllactosamine residue (Galα1-4GlcNAc-R) on carbohydrate
chains of glycolipids and glycoproteins, to form the α-Gal epitope
(27).
Additionally, the distributions of the α1,3GT gene
have a significant difference in various species (28). The α1,3GT gene of murine is
composed of nine exon sequences, and only six exons: Exons 4–9
(based on the numbering of mouse α1,3GT), are translated to α1,3GT
mRNA, which is ~1,500 base pairs (bp) (GenBank: M26925.1) (5). In Sus scrofa (pig), the
α1,3GT gene resides on chromosome 1, and α1,3-GT mRNA is 1,269 bp
(GenBank: L36152.1) (8). The
α1,3GT mRNA completed cds for bovine are 1,828 bp (GenBank:
J04989.1) (29). The coding
sequence of bovine α1,3GT predicates a membrane-bound protein with
three distinct structural features: A large, potentially
glycosylated COOH-terminal domain (346 amino acids), a single
transmembrane domain (16 amino acids), and a short NHz-terminal
domain (6 amino acids) (29). By
the analysis of the panel of human-rodent somatic cell hybrids, the
nonfunctional, processed pseudogene and the human homologue gene
represented by HGT-10 have been established to be located on human
chromosomes 12 and 9 (GenBank: J05421.1) (6,30).
The locus of α1,3GT mRNA for certain primates are about the same
locus as those of Gorilla gorilla (GenBank: M73304.1),
Pongo pygmaeus (GenBank: M73305.1) and Macaca mulatta
(GenBank: M73306.1) (31).
Anti-Gal antibody
The expression levels of the α-Gal epitope were
assessed by measuring the binding of the anti-Gal antibody with the
α-Gal epitope-specific lectin Bandeiraea simplicifolia IB4
in various cells. The distributions of α-Gal epitope, anti-Gal
antibody and the activities of α1,3GT in various species have a
significant difference. As shown in Table I, the α-Gal epitope is abundant on
red blood cells and nucleated cells of non-primate mammals,
prosimians and New World monkeys (such as marmoset, squirrel monkey
and spider monkey); however, it is absent in ancestral primates
such as humans, apes and Old World monkeys. Due to the continual
exposure of α-Gal epitope on the surface of entric bacteria and
other pathogens, and the lack of α-Gal epitope on humans and Old
World primates, they are not immunotolerant to the α-Gal epitope,
and therefore produce anti-α-Gal epitope antibody throughout the
whole life (32,33). As many as 1% of circulating B
cells are capable of generating the anti-α-Gal epitope antibody,
which is analogous to the mechanism by which the human ABO blood
group antibodies are produced. Anti-Gal antibody is not generated
at birth [except maternal-derived immunoglobulin G (IgG)]; however,
it is developed during the first few months of life. The anti-Gal
antibody isotypes of IgM, IgG, and IgA have been identified
(34–37), and the anti-Gal antibody in humans
is encoded by several heavy-chain genes, which are primarily within
the VH3 immunoglobulin gene family (38). When anti-Gal antibody specifically
binds with the α-Gal epitope, the binding site of anti-Gal antibody
resembles a 'pocket' for the α-Gal epitope with a size
corresponding with the free trisaccharide Gala1-3Galb1-4GlcNAc. The
affinity of the anti-Gal antibody to the free trisaccharide
Gala1-3Galb1-4GlcNAc is ~7-fold higher compared to the disaccharide
Gal1,3Gal. The similarity in binding of anti-Gal antibody to α-Gal
epitope on carbohydrate chains of glycoproteins (mannose in the
fourth position) and glycolipids (no mannose in the carbohydrate
chains) further implies that the pocket size is limited to a
trisaccharide length of the antigen.
 | Table IDistributions of α-Gal epitope,
anti-Gal antibody and the α1,3GT activity demonstrating significant
differences in various mammalian species. |
Table I
Distributions of α-Gal epitope,
anti-Gal antibody and the α1,3GT activity demonstrating significant
differences in various mammalian species.
| Mammals | α-Gal epitope
expression | α1,3-GT
activity | Anti-Gal antibody
expression |
|---|
| Non-primate
mammals | O | O | X |
| New world
monkeys | O | O | X |
| Prosimians | O | O | X |
| Old world
monkeys | X | X | O |
| Humans | X | X | O |
| Apes | X | X | O |
Evolution by the inactivation of α1,3GT
and anti-Gal antibody
According to the evolutionary tree of mammalian
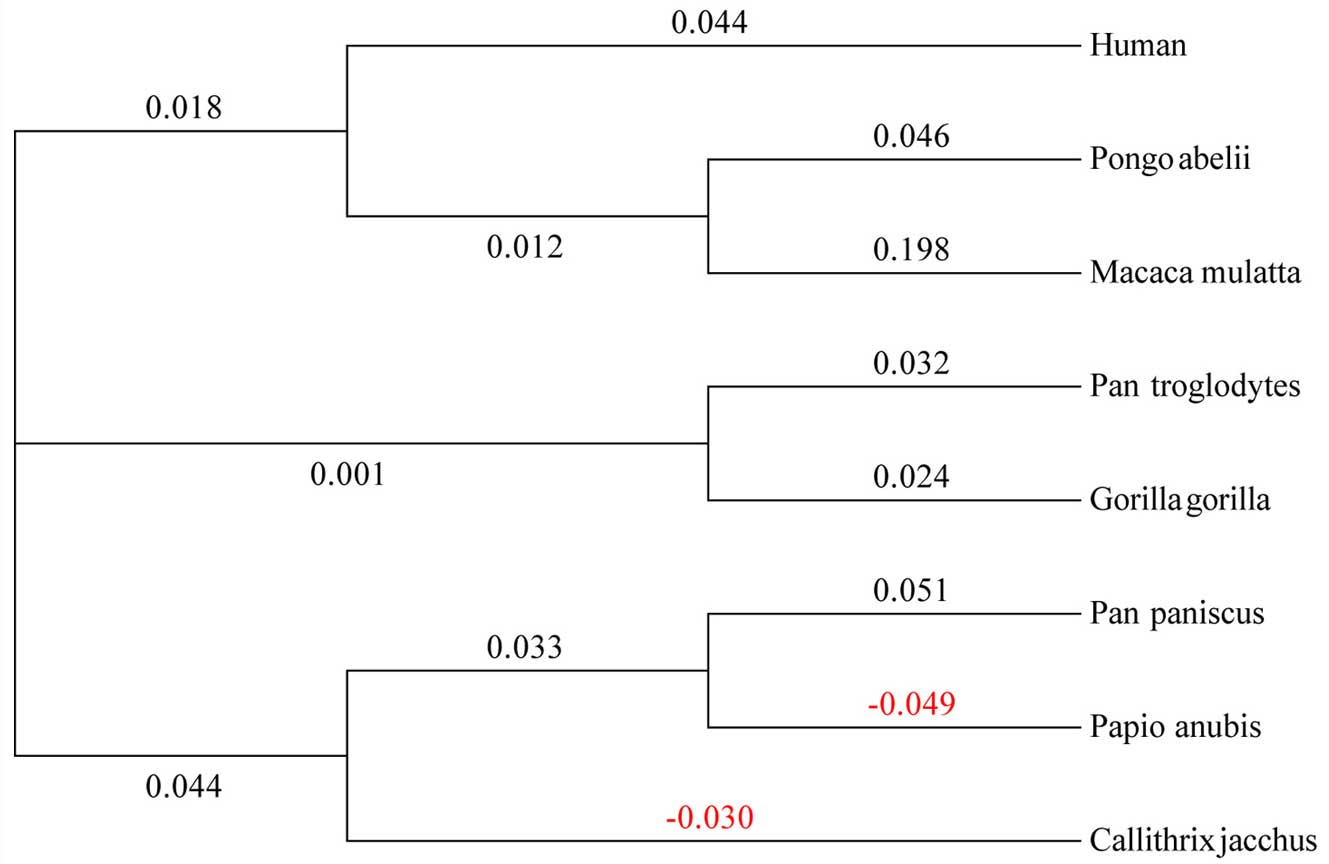
species (Fig. 1), it is clear
that α1,3GT and α-Gal epitope have been conserved in New World
monkeys (evolving on the South American continent, which was
separated from the African continent for >35 million years) and
in lemurs (prosimians residing in the Madagascar, a land mass
isolated from the African continent for >60 million years),
which suggests that α1,3GT appeared early in mammalian evolution,
and prior to the divergence of marsupials and placental mammals
(>125 million years ago) (39). However, following the geographical
separation between the South American and African continents,
ancestral Old World primates were subjected to a selective
evolutionary pressure, which resulted in the inactivation of the
α1,3GT gene. Northern blot results reveal that no transcripts of
the α1,3GT gene were identified in the higher primate species.
Notably, a variety of full-length transcripts were determined by
sensitive polymerase chain reaction in the tissues of rhesus
monkeys, orangutans and humans. Five crucial mutations were
delineated in the coding region of the human and rhesus, and three
crucial mutations were identified in the orangutan, any one of
which could be responsible for inactivation of the α1,3GT gene.
Therefore, the α1,3GT gene exhibits a unique evolutionary
characteristics.
Starting from the Old World primates, the α-Gal
epitope has been interrupted during the course of evolution.
Partial sequences of the α1,3GT gene, which governs the synthesis
of the α-Gal epitope, were detected in the human genome and were
found to be the corresponding pseudogenes. An experiment was
performed using the bovine α1,3GT cDNA as a probe to isolate two
non-overlapping clones (HGT-2 and HGT-10) from a human genomic DNA
library (40). HGT-2 contains a
putative coding region, containing multiple frame shift mutations
and nonsense codons in all three reading frames, which precludes
the synthesis of a functional enzyme. The coding sequences of
HGT-10, which is located on human chromosomes 12 and 9, contain
nonfunctional and processed pseudogene. Comparison of the
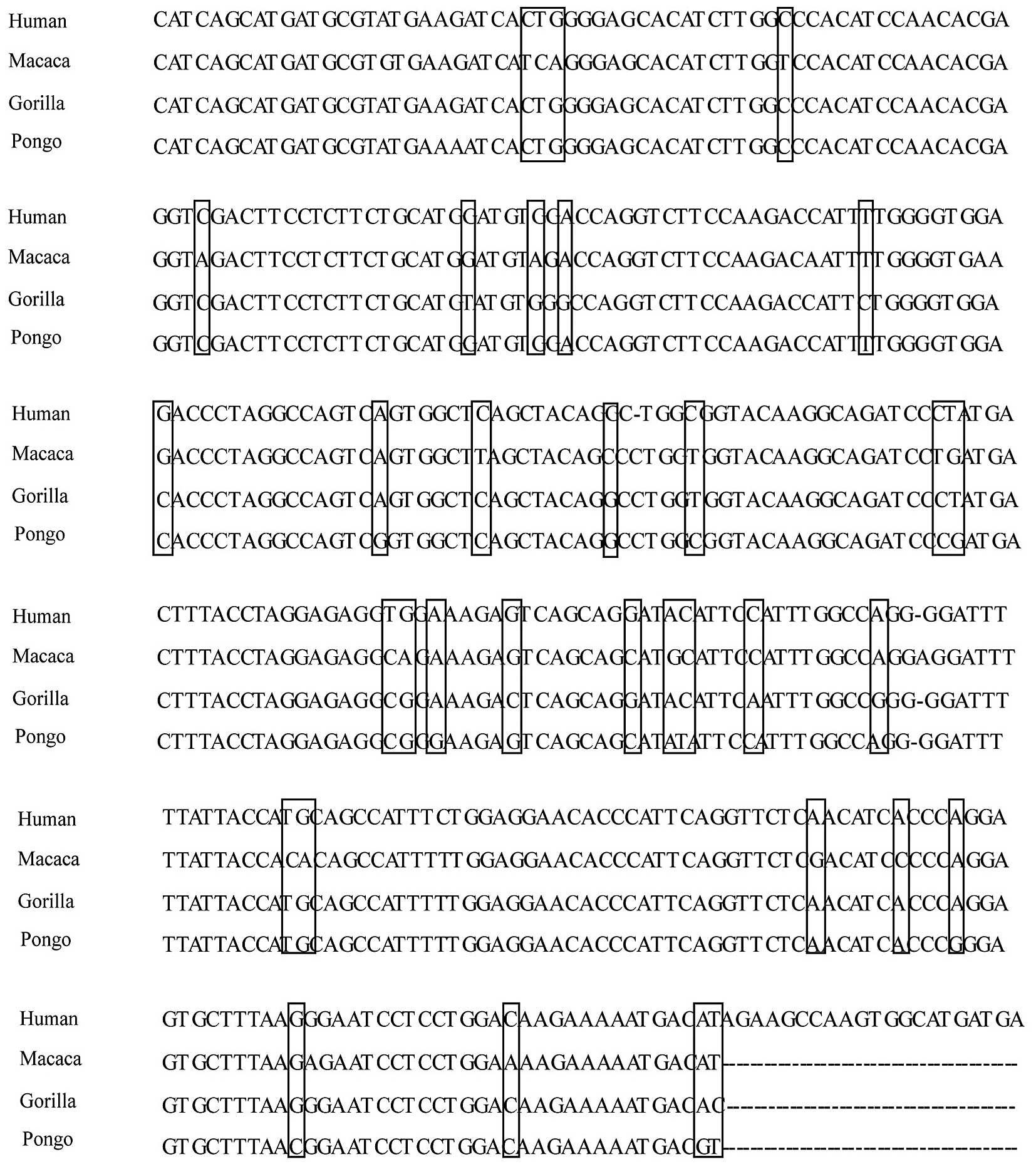
nucleotide sequence of the human α1,3GT pseudogene with the
corresponding different species (Fig.
2), the evolutionary loss of α-Gal epitope attributes to point
mutations in the coding region of the α1,3GT gene. The human α1,3GT
pseudogene resides on chromosome 9, and its sequence was found to
contain two point mutations, which results in a frame shift
mutation and a premature stop codon (13). Sequencing of this exon in the
chimpanzee shows the same two mutations, whereas the sequence from
gorilla only includes the first frame-shift mutation, which
generates a premature stop codon. The common mutations are
identified among the α1,3GT sequences of Old World monkey. These
findings in various primates suggest that the evolutionary
inactivation in ancestral Old World primates following the
divergence of apes and monkeys is estimated to have occurred ~20–25
million years ago. The evolutionary inactivation of the α1,3GT gene
resulted in the loss of immune tolerance to the α-Gal epitope and
the appearance of the anti-Gal antibody. Once the α-Gal epitope was
eliminated, primates could produce the anti-Gal antibody, possibly
by means of defending against pathogens, which express the α-Gal
epitope. Therefore, the inactivation of α1,3GT gene is generated by
several deletions, which ultimately encodes premature stop codons.
The observed inactivation of primate α1,3GT, along with distinct
geographical boundaries, suggests that it was due to an infectious
disease, which is endemic to the connected continents of the Old
World.
3. Attempts to overcome xenograft rejection
through the knockout of α1,3GT genes
A worldwide organs shortage for clinical
implantation causes 20–35% of patients, who need organ
transplantation, to succumb whilst on the waiting list. Therefore,
accompanied with increasing attention, the supply and demand for
the transplanted organs are the focuses of clinical
transplantation. With the application of xenotransplantation,
organs obtained from animal species provide a cure for the organ
shortage. Since non-human primates are the closest relatives to
humans, they were first considered as a potential organ source for
xenotransplantation (25).
Although chimpanzees were originally considered the best option, as
their organs are of similar size and they have good blood type
compatibility with humans, their organs were not widely used as
they are listed as an endangered species. Therefore, based on the
physiological, biological and ethical considerations, pigs appears
to be the most suitable donor for xenotransplantation. However,
when the pig organs were transplanted into Old World monkey, the
α-Gal epitope activates the complement system, creating C5a and C3b
and causing a cascade of further cleavage and activation events.
C3b binds to the surface of pathogens, leading to greater
internalization by phagocytic cells through opsonization. C5a is an
important chemotactic protein, helping the recruitment of
inflammatory cells, such as macrophages and neutrophils (41). Therefore, when the α-Gal epitope,
expressed on endothelial cells of the blood vessel walls in the
xenograft, binds with the anti-Gal antibody, it will result in the
activation and induction of platelet aggregation, and leads to the
collapse of the xenograft vascular bed and to the rapid hyperacute
rejection (42).
There are three major components to hyperacute
rejection: i) Antigen, ii) antibody and iii) complement (25). In order to overcome the hyperacute
rejection, actions can be taken to relieve the hyperacute rejection
by the inactivation of the α1,3GT gene. There are two commonly used
methods to relieve the hyperacute rejection. One strategy to the
inactivation of the α1,3GT gene would be using the transgenic
approach to add another transferase, which would deviate the
glycosylation pathway from α-Gal epitope expression and lead to the
production of another carbohydrate not recognized by natural
antibodies. The α(1,2) fucosyltransferase and α(2,3)
sialyltransferase or α(2,6) sialyltransferase are known to be
different glycosyltransferases, which compete for the same
substrate with each other. They can be applied to develop a
strategy aimed at blocking a specific carbohydrate epitope, an
appropriate enzyme to decrease the expression of Gal(1,3)Gal. According to a previous study, the
results of this method demonstrated that the amount of α-Gal
epitope is lesser than normal, but cannot be eliminated completely
(25). Another strategy is
accomplished by the disruption of the pig α1,3GT gene locus,
mediated by a pPL657 vector, targeting exon 9 (the location
encoding the catalytic domain of the α1,3GT gene) (38). Homozygous inactivation of these
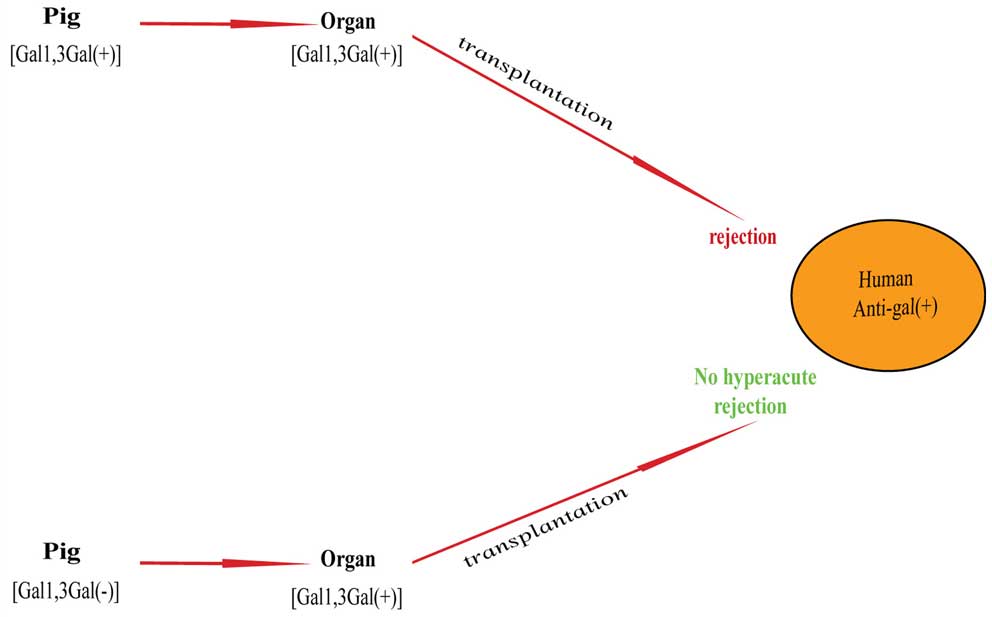
two alleles of the α1,3GT genes results in an inactive enzyme. It
is demonstrated that the natural antibodies of the α1,3GT KO pig
would appear to be similar to those antibodies in other species.
Therefore, α1,3GT cannot be encoded without the α1,3GT gene, and
the α-Gal epitope would not be synthesized. The strategy of
knocking out the gene of α1,3GT successfully avoids the hyperacute
rejection. In the same way, by studying the anti-Gal positive
B-cell in α1,3GT KO mice responding to α-Gal epitope of xenografts,
these mice are indicated to lack the α-Gal epitope due to the
disruption of the α1,3GT gene (43). The same experiment was carried out
on pigs, and organ transplantation from α1,3GT KO transgenic pigs
to humans did not result in complement-mediated hyperacute
rejection. Therefore in this way, rapid hyperacute rejection can be
eliminated, and the organ transplantation from pig to humans
becomes a clinically acceptable treatment (Fig. 3).
4. Clinical exploitation
Anti-Gal antibody, which is universal in human, can
effectively bind with the α-Gal epitope, and therefore activates
the complement cascade, which generates chemotactic cleavage
complement peptides, such as C5a and C3b. These chemotactic
peptides induce rapid and extensive migration of macrophages. Based
on the structure, mechanism and specificity of the anti-Gal
antibody, it can be exploited for a variety of clinical uses.
Firstly, the α1,3GT KO pigs can be used as organ donors from pig to
human in xenotransplantation. Secondly, the immunogenicity of tumor
vaccines and certain viral vaccines, including flu and HIV vaccine,
can be increased by the binding of anti-Gal antibody with α-Gal
epitope. In addition, wound healing, skin burns and tissue
regeneration can be induced by α-gal nanoparticles. The
regeneration of internal injuries, such as ischemic myocardium and
injured nerves, will be one of its applications in the future as
well.
Xenotransplantation
Xenotransplantation is one of the most important
methods to solve the problem of the organ shortage in humans. Those
cells, tissues or organs from different species are known as
xenografts, which are widely used for xenotransplantation (44). For example, the heart valve
substitutes and the patch materials are applied to a variety of
cardiovascular diseases (45).
However, the major barrier to the clinically successful
xenotrans-plantation is the lack of effective therapies aimed at
eliminating antibody and complement. Therefore, subsequent to
knocking out the α1,3GT gene, the α-Gal epitope cannot be produced,
and hyperacute rejection can be overcome (46).
As the whole organs of the heart is not the ideal
choice for heart xenotransplantation, heart valve substitutes and
patch materials are generally applied to a variety of
cardiovascular disease (45), and
the major application is the bioprosthetic heart valves (BHVs)
(47,48). Currently, the most commercially
available BHVs are made from porcine aortic valve or bovine
pericardium (49). They are
conventionally cross-linked with glutaraldehyde (GA) to impart
tissue stability, to reduce antigenicity and to maintain tissue
sterility (50).
However, GA-fixed xenografts are prone to dystrophic
calcification after long-term implantation in humans, which is one
of the limiting factors affecting the longevity of bioprostheses
(47). The mechanism of the
calcification of GA-fixed xenografts is complex and is not
completely understood. It is supposed that the degeneration of
bioprostheses may begin with the penetration of immunoglobulines
into the valve-matrix. Subsequently, macrophages are deposited on
the valve surface and erythrocytes penetrate into the valve.
Finally, collagen breakdown and calcification occur. Thus, the
deposition of immunoglobulins represents the initial immunological
trigger for calcification in bioprostheses (50). Another proposed mechanism
responsible for the calcification of xenografts is an immune
response to the xenografts (51).
According to the method of anti-calcification treatments, bovine
pericardial tissues were divided into eight groups. Prepared
tissues were subcutaneously implanted into the α-gal KO mice and
wild-type mice. The result demonstrated that except the high
concentration GA-treated group, calcium contents of
anticalcification-treated groups were significantly lower compared
to the control group (45). These
findings suggest a possible role of immune response in the
calcification of xenografts, and α-Gal epitope on the BHV
contributes to the immune response (52–54). The α-Gal epitope on BHV may be
removed completely using recombinant α-galactosidase and by
performing a decellularization procedure without modifying the
mechanical properties of the tissues (25,55,56). There is no difference in the
mechanical properties of BHV following the application of the
methods (57,58). In addition, a series of 7
heterotopic heart transplantations were performed from cluster of
differentiation 46-positive transgenic pigs to baboons using a
combination of therapeutic agents mainly targeted at controlling
the synthesis of anti-pig antibodies. This study reported that the
longest median survival of pig hearts transplanted into baboons was
96 days (59). However, there
also are numerous problems regarding BHV transplantation, in
pig-to-primate or human BHV. Although hyperacute rejection can be
prevented, the next rejection phenomenon unique to
xenotransplantation, known as delayed xenograft rejection or acute
vascular rejection, will occur (60). In addition, new immunological
problems arise, for example, except the α-Gal epitope, there exist
non-gal carbohydrate antigens and the identification of their
elusive antigens as neutrophils, macrophages, natural killer cells
and T cells (61).
Studies on the xenotransplantation of heart and
kidney from α1,3GT KO pigs to monkeys demonstrate that pig hearts
could live for ≤6 months, pig kidneys for 8–32 days, and pig
kidneys containing vascularized thymic tissue for up to 83 days
(62). Therefore, transplantation
of organs from α1,3GT KO pigs to humans becomes clinically
acceptable. However, in addition to the α-Gal epitope, the non-gal
carbohydrate antigen is another obstacle in kidney transplantation.
A study of total N-glycans from membrane glycoproteins, derived
from specific pathogen-free miniature pig kidneys, was carried out,
and the result showed that α-gal was the most abundant compound
among the neutral pig N-glycans. However, the various N-glycan
structures of Neu5Gc and Gala1-3Lex epitope as new non-gal
carbohydrate antigens were identified by mass spectrometry
(46,63). Therefore, non-gal carbohydrate
antigens may be the next complication in the transplantation from
animals to humans.
Cancer immunotherapy
Tumor cells express a variety of tumor-specific
antigens in animals and in humans. Some of the tumor antigens are
common to numerous types of tumors. Additionally, as certain
antigens may also be expressed on normal cells, the immune response
caused by these common tumor antigens is usually extremely low
(64). However, tumor antigens
are specific to the tumor type and to the patient. Therefore, the
autologous tumor cells could be considered a suitable source for
the production of the vaccine. In order to induce the expression of
α-Gal epitope by the tumor cells, chemical, biochemical and genetic
engineering can be applied (8).
As an example of the chemical and biochemical methods, autologous
intact tumor cells from hematological malignancies, or autologous
tumor cell membranes from solid tumors are processed to express
α-Gal epitope by the incubation with neuraminidase, recombinant
α1,3GT and uridine diphosphate galactose (9). Additionally, the expression of the
α-Gal epitope can also be achieved by the transduction of
recombinant adenovirus pAd-sGT, expressing pig α1,3GT genes, into
the tumor cells (65).
The adenoviral vector-mediated transduction of the
pig α1,3GT gene into human tumor cells, such as malignant melanoma
A375, stomach cancer SGC-7901, and lung cancer SPC-A-1, has been
reported. These studies demonstrated that α-Gal epitope did not
increase the sensitivity of human tumor cells to human
complement-mediated lysis (66).
Studies on the immune response to tumor vaccines have indicated
that, to achieve effective activation of tumor-specific naive T
cells, the vaccinating antigens must be effectively taken up by
antigen-presenting cells (APCs). Subsequently, the vaccinating
antigens should be transported from the vaccinating tumor membranes
to the regional lymph nodes, and the tumor-associated antigen (TAA)
peptides will be processed and presented by APCs (67–69). The autologous TAA peptides
activate tumor-specific T cells within the lymph nodes, eliciting
an immune response, which would eradicate the residual tumor cells
following completion of standard therapy (70).
In addition, the tumor cells can also express the
α-Gal epitope by the transfection of α1,3GT genes through gene
engineering. Following genetic manipulation, the binding ability of
the tumor cells with human IgG and IgM is increased. However, the
sensitivity of human tumor cells to human complement-mediated lysis
is not enhanced. In a cancer-related study, studies on the immune
response of tumor vaccines have indicated that a major prerequisite
for the formation of tumor vaccines is their effective uptake by
APCs and the transportation of these APCs to the regional lymph
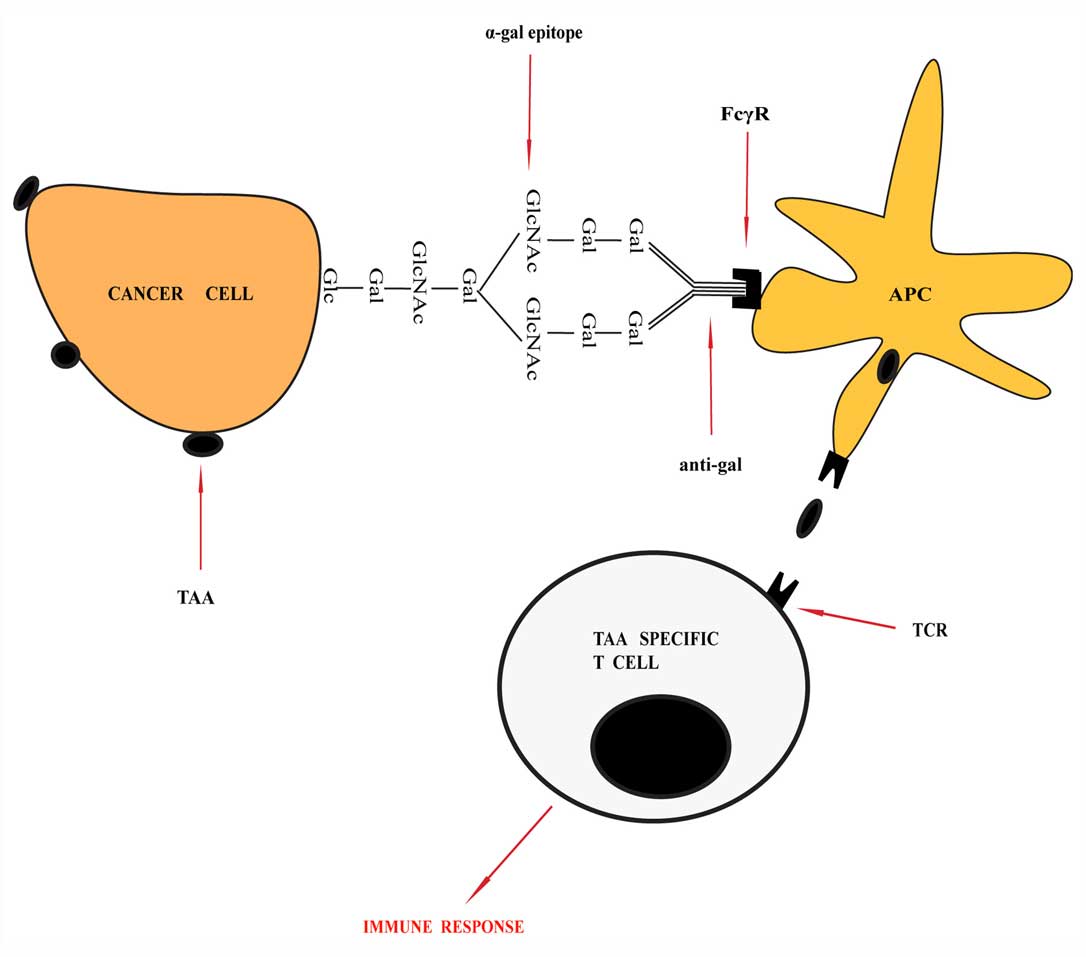
nodes where TAAs can be processed and presented (21,71). In detail, TAAs activate the immune
response, and the anti-Gal antibody would specifically bind to
α-Gal epitope. The Fc portion of the anti-Gal antibody binds with
the Fcα receptors on APCs and induce the effective uptake of the
vaccine by the APCs (Fig. 4)
(72). Subsequent to transporting
the vaccinating tumor membranes to the regional lymph nodes, TAA
peptides can be processed and presented. The autologous TAA
peptides activate tumor-specific T cells within the lymph nodes,
which causes an immune response and it will eradicate the residual
tumor cells that remain following the completion of standard
therapy (73).
With regard to the application of tumor vaccines,
MUC1 was overexpressed and aberrantly glycosylated in the majority
of human ductal pancreatic carcinoma (74). The α1,3GT KO mice used as the
model were pre-immunized with pig kidney and transplanted with
B16F10 melanoma cells, and were transfected with the MUC1
expressing vector. Compared with the mice vaccinated with MUC1
alone, the tumor growth was significantly inhibited and the
survival time was significantly improved in mice vaccinated with
pancreatic cancer cells expressing α-Gal epitope. Based on this
study, it will be a novel strategy for the treatment of pancreatic
cancer (75,76). Another study reported the
treatment of 11 patients with advanced solid tumors with
intratumoral injection of 0.1, 1 or 10 mg α-gal glycolipids. There
was no dose-limiting toxicity and no clinical or laboratory
evidence of autoimmunity, or any other toxicity. Few patients had
an unexpectedly long survival. Therefore, it is feasible and safe
for inducing a protective antitumor immune response (77). The mice vaccinated with irradiated
melanoma cells, which express a-Gal epitope, were followed by a
challenge with live parental melanoma cells, and resulted in
protection with vaccines for ≥2 months in one-third of the mice.
The proportion of survival mice protected with the vaccines doubled
when they were immunized twice with irradiated melanoma cells
expressing α-Gal epitope, and challenged with the life span of
those of the control mice. A further histological study on the
developing tumors in the challenged mice, which were immunized with
melanoma cells expresssing α-Gal epitope, demonstrated the
extensive infiltration of T lymphocytes and macrophages (73). Chiefly, anti-gal can be applied to
the cancer immunotherapy, and breast cancer and brain tumors may be
treated by autologous tumor vaccines in the future.
Viral vaccines
Viral vaccines include the flu and HIV vaccines.
Similar to the tumor vaccine, targeting the anti-gal-mediated
complex to the APCs is essential (78). The Flu vaccine is mainly comprised
of the viral envelope glycoprotein 'hemagglutinin' (HA), which may
be served as a useful example for the application of anti-Gal into
vaccines. In the majority of strains of flu virus, HA has 6–8
N-linked carbohydrate chains, the majority of which are of the
complex type. This study demonstrates a method for the expression
of α-Gal epitope on the influenza virus HA by recombinant α1,3GT
(79). To generate the flu
vaccine expressing α-Gal epitope, sialic acid is removed from viral
glycoproteins following its digestion by viral neuraminidase, and
the terminal of the carbohydrate chains of HA was residual
N-acetyllactosamine. Subsequently, α1,3GT transfers sugar from the
sugar donor UDP-gal to the residual N-acetyllactosamine (64). Therefore, α-Gal epitope could be
added to all carbohydrate chains of HA. In the study of the α1,3GT
KO mice, the control group of the KO mice were immunized with the
original inactivated flu virus lacking α-Gal epitope, and another
group of the KO mice were immunized with 1 µg of inactivated flu
virus vaccine expressing the α-Gal epitope. The result demonstrated
that the anti-flu virus antibody and T cell response in the group
that was immunized with the virus vaccine expressing α-Gal epitope
was ~100-fold higher compared to the control group. Furthermore,
the survival time of KO mice with the vaccine expressing α-Gal
epitope infected with live virus was ~9-fold longer compared to the
KO mice immunized with the vaccine lacking this epitope (21). Therefore, the immunogenicity of
flu vaccines is increased by targeting anti-Gal antibody mediated
complex into APCs.
In addition, the HIV vaccine is another notable
viral vaccine. The main glycoprotein on the surface of HIV is
gp120, which has 24 N-linked glycosylation sites of the
carbohydrate chains, and the terminal structure of the gp120
carbohydrate chains is SA-Galα1-4GlcNAc-R. Following the incubation
with neuraminidase, UDP-gal and α1,3GT, gp120 can be capped with
the α-Gal epitope. Studying the HIV vaccine on KO mice, it
demonstrated that the anti-gp120 antibody and T cell responses were
increased ~100-fold following the immunization of gp120 with α-Gal
epitope (80).
Chiefly, the immunogenicity of the flu and HIV
vaccines is increased by expressing the α-Gal epitope. This method
can be used for any microbial, soluble or particulate vaccine.
However, the recombination does not affect the immunogenic epitopes
of the vaccine.
Wound healing, skin burns and tissue
regeneration by α-gal nanoparticles
The interaction between the anti-Gal antibody and
α-Gal epitope activates complement system, which induces rapid
recruitment of macrophages. Wound healing requires the help of the
local recruitment and the activation of macrophages. The activated
macrophages generate cytokines, such as vascular endothelial growth
factor, interleukin-1, platelet-derived growth factor and
colony-stimulating factor. These cytokines promote the healing of
injuries. However, if the α-Gal epitope was expressed on cells with
inserted α-gal glycolipids, the binding of anti-Gal antibody with
α-Gal epitope may lead to cell lysis and a pro-inflammatory
response. To avoid this pro-inflammatory response, the α-Gal
epitope is expressed on cells in the form of nanoparticles and
subsequently there will be no insertion into the cells (38). These nanoparticles were generated
from phospholipids, cholesterol and α-gal glycolipids. An
experiment regarding full-thickness wounds (20×20 mm) with tattooed
borders was carried out on the back of α1,3GT KO pigs. Treated
wounds with α-gal nanoparticles exhibited more macrophages compared
to the control wounds (saline-treated) in the same pig and at the
same time, and it was observed that angiogenesis was increased in
the wounds with α-gal nanoparticles. No keloid formation or
increase in scar formation was observed in pigs treated by α-gal
nanoparticles (81,82). Another study demonstrated that in
contrast to the wild-type mice, the healing time of excisional skin
wounds treated with α-gal liposomes in α1,3GT KO mice was twice as
fast as that of control wounds. Additionally, the time of scar
formation in α-gal liposome-treated wounds is much less compared to
those wounds with physiological healing (83).
In addition, the anti-Gal antibody can be used on
the skin burns healing as well. Efficacy of treatment in
accelerating burn healing is demonstrated in pairs of burns in
α1,3GT KO mice. It has been demonstrated that 3 days later, the
α-gal liposomes treated burns contained 5-fold as many neutrophils
as those of the control burns, and macrophages were found only in
α-gal liposome-treated burns. Six days later, 50–100% of the
surface area of the α-gal liposome-treated burns were covered with
regenerating epidermis (re-epithelialization), and almost no
epidermis was identified in the control burns (84).
Therefore, the extensive recruitment of macrophages
by α-gal nanoparticles that have interacted with anti-Gal antibody
will effectively accelerate wound healing, skin burns and tissue
regeneration.
Regeneration on ischemic myocardium and
injured nerves
In the same way of accelerating wound healing, skin
burns healing and tissue regeneration, α-gal nanoparticles may also
be applied to the regeneration on ischemic myocardium and injured
nerves. Combined with the regeneration on ischemic myocardium and
injured nerves, the mechanisms of these treatments have numerous
similarities. Firstly, the mechanism of regeneration of ischemic
myocardium and injured nerves requires the recruitment of
macrophages, which will secrete cytokines/growth factors. One of
the differences between them is that the regeneration on ischemic
myocardium requires the recruitment of stem cells, which received
cues from the adjacent healthy cells, the microenvironment and the
extracellular matrix to differentiate into cardiomyocytes, which
regenerate the tissue and restore its physiological activity.
However, the regeneration of nerves requires re-growth of multiple
sprouts from injured axons. These sprouts will reconnect across the
lesion and grow into the distal axonal tube of the damaged neurons.
In addition, they have a default mechanism for tissue repair
simultaneously. When the recruitment of macrophages is delayed,
leading to no important cytokines/growth factors regeneration, the
useful technique would be removal, which is irreversible.
The treatments for regeneration of ischemic
myocardium and injured nerves by α-gal nanoparticles are
theoretical possibilities at present, and the experiment is
expectantly being carried out on mammalian animal models (38).
5. Conclusion
The α-Gal epitope has a unique carbohydrate
structure, and the distribution of the α-Gal epitope, anti-Gal
antibody and the α1,3GT activity has a big difference in various
mammalian species. Previous studies showed that the inactivation of
the α1,3GT in ancestral primates can explain the inactivation of
the primate α1,3GT along distinct geographical boundaries. The
divergence is ascribed to the pressure coming from natural
selective. With regards to the notable differences in the
distribution of α-Gal epitope expression, the activity of α1,3GT
and the expression of anti-Gal antibody among ancestral primates
and other species, the anti-Gal antibody is regarded as a barrier
in the transplantation from animals, such as pigs to humans, which
causes the immune response to xenografts. However, the α1,3GT KO
pig can aid with this problem of rejection. The anti-Gal antibody
can be exploited for clinical use by increasing the immunogenicity
of viral vaccines and autologous tumor vaccines, which can be
processed to express the α-Gal epitope. In addition, the α-Gal
epitope, expressed by the tumor cells, can be generated by certain
methods. In vivo, it can be produced by catalysis with
α1,3GT. In vitro, its synthesis can be completed through
chemical, biochemical and genetic engineering methods. By targeting
those vaccines to APCs at the vaccination site, the immune
responses induced will be potent to clear up the residual tumor
cells following standard chemical therapy. Therefore, this method
can be applied to the cancer immunotherapy. In addition, in the
same way as the formation of tumor vaccines, the formation of viral
vaccines, such as flu and HIV vaccines, will be feasible in the
future. In another way, accelerating wound healing is achieved by
the application of α-gal nanoparticles, which would bind with the
anti-Gal antibody by activating complement system, recruiting and
activating macrophages and therefore inducing tissue regeneration.
Thus, with the help of α-gal nanoparticles, the healing time of
wounds and injury can be reduced efficiently. The regeneration on
ischemic myocardium and injured nerves is a direction for the
future.
Acknowledgments
The present study was supported by the Natural
Science Foundation of Sichuan, China (no. 2010SZ0139) and the
Special Program for Youth Science and Technology Innovative
Research Scientist of Sichuan, China (no. 08ZQ026-028).
References
|
1
|
Boix E, Zhang Y, Swaminathan GJ, Brew K
and Acharya KR: Structural basis of ordered binding of donor and
acceptor substrates to the retaining glycosyltransferase,
alpha-1,3-galacto-syltransferase. J Biol Chem. 277:28310–28318.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shao Y, Yu Y, Pei CG, Qu Y, Gao GP, Yang
JL, Zhou Q, Yang L and Liu QP: The expression and distribution of
α-Gal gene in various species ocular surface tissue. Int J
Ophthalmol. 5:543–548. 2012.
|
|
3
|
Galili U: The alpha-gal epitope and the
anti-Gal antibody in xenotransplantation and in cancer
immunotherapy. Immunol Cell Biol. 83:674–686. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Blanken WM and Van den Eijnden DH:
Biosynthesis of terminal Gal alpha 1→3Gal beta 1→4GlcNAc-R
oligosaccharide sequences on glycoconjugates. Purification and
acceptor specificity of a UDP-Gal:N-acetyllactosaminide alpha
1→3-galactosyltrans-ferase from calf thymus. J Biol Chem.
260:12927–12934. 1985.PubMed/NCBI
|
|
5
|
Taylor SG, McKenzie IF and Sandrin MS:
Characterization of the rat alpha(1,3)galactosyltransferase:
evidence for two independent genes encoding glycosyltransferases
that synthesize Galalpha(1,3)Gal by two separate glycosylation
pathways. Glycobiology. 13:327–337. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Joziasse DH, Shaper JH, Jabs EW and Shaper
NL: Characterization of an alpha 1→3-galactosyltransferase
homologue on human chromosome 12 that is organized as a processed
pseudogene. J Biol Chem. 266:6991–6998. 1991.PubMed/NCBI
|
|
7
|
Church DM, Goodstadt L, Hillier LW, Zody
MC, Goldstein S, She X, Bult CJ, Agarwala R, Cherry JL, DiCuccio M,
et al: Mouse Genome Sequencing Consortium: Lineage-specific biology
revealed by a finished genome assembly of the mouse. PLoS Biol.
7:e10001122009. View Article : Google Scholar
|
|
8
|
Strahan KM, Gu F, Preece AF, Gustavsson I,
Andersson L and Gustafsson K: cDNA sequence and chromosome
localization of pig alpha 1,3 galactosyltransferase.
Immunogenetics. 41:101–105. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zimin AV, Delcher AL, Florea L, Kelley DR,
Schatz MC, Puiu D, Hanrahan F, Pertea G, Van Tassell CP, Sonstegard
TS, et al: A whole-genome assembly of the domestic cow, Bos taurus.
Genome Biol. 10:R422009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lindblad-Toh K, Wade CM, Mikkelsen TS,
Karlsson EK, Jaffe DB, Kamal M, Clamp M, Chang JL, Kulbokas EJ III,
Zody MC, et al: Genome sequence, comparative analysis and haplotype
structure of the domestic dog. Nature. 438:803–819. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lander ES, Linton LM, Birren B, Nusbaum C,
Zody MC, Baldwin J, Devon K, Dewar K, Doyle M, FitzHugh W, et al:
International Human Genome Sequencing Consortium: Initial
sequencing and analysis of the human genome. Nature. 409:860–921.
2001. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Galili U: Evolution and pathophysiology of
the human natural anti-alpha-galactosyl IgG (anti-Gal) antibody.
Springer Semin Immunopathol. 15:155–171. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Larsen RDR-MC, Rivera-Marrero CA, Ernst
LK, Cummings RD and Lowe JB: Frameshift and nonsense mutations in a
human genomic sequence homologous to a murine UDP-Gal:beta-D-G
al(1,4)-D-GlcNAc alpha(1,3)-galactosyltransferase cDNA. J Biol
Chem. 265:7055–7061. 1990.PubMed/NCBI
|
|
14
|
Rodriguez IA and Welsh RM: Possible role
of a cell surface carbohydrate in evolution of resistance to viral
infections in old world primates. J Virol. 87:8317–8326. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hamanova M, Chmelikova M, Nentwich I, Thon
V and Lokaj J: Anti-Gal IgM, IgA and IgG natural antibodies in
childhood. Immunol Lett. 164:40–43. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang L, Radic MZ and Galili U: Human
anti-Gal heavy chain genes. Preferential use of VH3 and the
presence of somatic mutations. J Immunol. 155:1276–1285.
1995.PubMed/NCBI
|
|
17
|
Commins SP and Platts-Mills TA: Delayed
anaphylaxis to red meat in patients with IgE specific for galactose
alpha-1,3-ga-lactose (alpha-gal). Curr Allergy Asthma Rep.
13:72–77. 2013. View Article : Google Scholar :
|
|
18
|
Galili U, LaTemple DC and Radic MZ: A
sensitive assay for measuring alpha-Gal epitope expression on cells
by a monoclonal anti-Gal antibody. Transplantation. 65:1129–1132.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
LaTemple DC and Galili U: Adult and
neonatal anti-Gal response in knock-out mice for
alpha1,3galactosyltransferase. Xenotransplantation. 5:191–196.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Park S, Kim WH, Choi SY and Kim YJ:
Removal of alpha-Gal epitopes from porcine aortic valve and
pericardium using recombinant human alpha galactosidase A. J Korean
Med Sci. 24:1126–1131. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Abdel-Motal UM, Guay HM, Wigglesworth K,
Welsh RM and Galili U: Immunogenicity of influenza virus vaccine is
increased by anti-gal-mediated targeting to antigen-presenting
cells. J Virol. 81:9131–9141. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gurtner GCWS, Werner S, Barrandon Y and
Longaker MT: Wound repair and regeneration. Nature. 453:314–321.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
LaTemple DC, Henion TR, Anaraki F and
Galili U: Synthesis of alpha-galactosyl epitopes by recombinant
alpha1,3galactosyl transferase for opsonization of human tumor cell
vaccines by anti-galactose. Cancer Res. 56:3069–3074.
1996.PubMed/NCBI
|
|
24
|
Singer AJ and Clark RA: Cutaneous wound
healing. N Engl J Med. 341:738–746. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sandrin MS, Osman N and McKenzie IF:
Transgenic approaches for the reduction of Galalpha(1,3)Gal for
xenotransplantation. Front Biosci. 2:e1–11. 1997.PubMed/NCBI
|
|
26
|
Osman N, McKenzie IF, Mouhtouris E and
Sandrin MS: Switching amino-terminal cytoplasmic domains of
alpha(1,2) fucosyltransferase and alpha(1,3)galactosyltransferase
alters the expression of H substance and Galalpha(1,3)Gal. J Biol
Chem. 271:33105–33109. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xing L, Xia GH, Fei J, Huang F and Guo LH:
Adenovirus-mediated expression of pig alpha(1,3)
galactosyltransferase reconstructs Gal alpha(1, 3) gal epitope on
the surface of human tumor cells. Cell Res. 11:116–124. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu M, Zhu SM, Zheng H, Wang Y, Wang Z,
Yang YJ, Wu YX, Zeng YZ and Wang YP: Cloning of splicing variants
of alpha1,3-galactosyltransferase cDNA of Chinese Banna Minipig
inbred line and its expression in human cells. Sichuan Da Xue Xue
Bao Yi Xue Ban. 43:145–150. 2012.In Chinese. PubMed/NCBI
|
|
29
|
Joziasse DH, Shaper JH, Van den Eijnden
DH, Van Tunen AJ and Shaper NL: Bovine alpha
1→3-galactosyltransferase: Isolation and characterization of a cDNA
clone. Identification of homologous sequences in human genomic DNA.
J Biol Chem. 264:14290–14297. 1989.PubMed/NCBI
|
|
30
|
Lantéri M, Giordanengo V, Vidal F, Gaudray
P and Lefebvre JC: A complete alpha1,3-galactosyltransferase gene
is present in the human genome and partially transcribed.
Glycobiology. 12:785–792. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Galili U and Swanson K: Gene sequences
suggest inactivation of alpha-1,3-galactosyltransferase in
catarrhines after the divergence of apes from monkeys. Proc Natl
Acad Sci USA. 88:7401–7404. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ma YH, Zhou XG, Hu JH, Fei J, Xia GH and
Guo LH: Human xenoreactivity is reduced in mice bearing porcine
antisense alpha(1,3) galactosyltransferase cDNA. Acta Pharmacol
Sin. 22:231–238. 2001.PubMed/NCBI
|
|
33
|
Galili U and Andrews P: Suppression of
a-galactosyl epitopes synthesis and production of the natural
anti-Gal antibody: A major evolutionary event in ancestral Old
World primates. J Hum Evol. 29:433–442. 1995. View Article : Google Scholar
|
|
34
|
Fang J, Walters A, Hara H, Long C, Yeh P,
Ayares D, Cooper DK and Bianchi J: Anti-gal antibodies in
α1,3-galactosyltransferase gene-knockout pigs. Xenotransplantation.
19:305–310. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Koike C, Fung JJ, Geller DA, Kannagi R,
Libert T, Luppi P, Nakashima I, Profozich J, Rudert W, Sharma SB,
et al: Molecular basis of evolutionary loss of the alpha
1,3-galac-tosyltransferase gene in higher primates. J Biol Chem.
277:10114–10120. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sandrin MS, Vaughan HA, Dabkowski PL and
McKenzie IF: Anti-pig IgM antibodies in human serum react
predominantly with Gal(alpha 1–3) Gal epitopes. Proc Natl Acad Sci
USA. 90:11391–11395. 1993. View Article : Google Scholar
|
|
37
|
Galili U, Rachmilewitz EA, Peleg A and
Flechner I: A unique natural human IgG antibody with
anti-a-galactosyl specificity. J Exp Med. 160:1519–1581. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Galili U: Anti-Gal: An abundant human
natural antibody of multiple pathogeneses and clinical benefits.
Immunology. 140:1–11. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Galili U: Significance of the evolutionary
α1,3-galacto-syltransferase (GGTA1) gene inactivation in preventing
extinction of apes and old world monkeys. J Mol Evol. 80:1–9. 2015.
View Article : Google Scholar
|
|
40
|
Ma YH1, Zhou XG, Hu JH, Fei J, Xia GH and
Guo LH: Human xeno-reactivity is reduced in mice bearing porcine
antisense alpha(1,3) galactosyltransferase cDNA. Acta Pharmacol
Sin. 22:231–238. 2001.PubMed/NCBI
|
|
41
|
Celis E, Abraham KG and Miller RW:
Modulation of the immunological response to hepatitis B virus by
antibodies. Hepatology. 7:563–568. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Byrne GW, Stalboerger PG, Du Z, Davis TR
and McGregor CG: Identification of new carbohydrate and membrane
protein antigens in cardiac xenotransplantation. Transplantation.
91:287–292. 2011. View Article : Google Scholar
|
|
43
|
Galili U: Interaction of the natural
anti-Gal antibody with alpha-galactosyl epitopes: A major obstacle
for xenotransplan-tation in humans. Immunol Today. 14:480–482.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Anderson M: Xenotransplantation: A
bioethical evaluation. J Med Ethics. 32:205–208. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lee C, Ahn H, Kim SH, Choi SY and Kim YJ:
Immune response to bovine pericardium implanted into
α1,3-galactosyltransferase knockout mice: feasibility as an animal
model for testing efficacy of anticalcification treatments of
xenografts. Eur J Cardiothorac Surg. 42:164–172. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kim YG, Gil GC, Harvey DJ and Kim BG:
Structural analysis of alpha-Gal and new non-Gal carbohydrate
epitopes from specific pathogen-free miniature pig kidney.
Proteomics. 8:2596–2610. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Park CS, Oh SS, Kim YE, Choi SY, Lim HG,
Ahn H and Kim YJ: Anti-alpha-Gal antibody response following
xenogeneic heart valve implantation in adults. J Heart Valve Dis.
22:222–229. 2013.PubMed/NCBI
|
|
48
|
Wilczek P, Lesiak A, Niemiec-Cyganek A,
Kubin B, Slomski R, Nozynski J, Wilczek G, Mzyk A and Gramatyka M:
Biomechanical properties of hybrid heart valve prosthesis utilizing
the pigs that do not express the galactose-α-1,3-galactose (α-Gal)
antigen derived tissue and tissue engineering technique. J Mater
Sci Mater Med. 26(5329)2015. View Article : Google Scholar
|
|
49
|
Choi SY, Jeong HJ, Lim HG, Park SS, Kim SH
and Kim YJ: Elimination of alpha-gal xenoreactive epitope:
Alpha-galactosidase treatment of porcine heart valves. J Heart
Valve Dis. 21:387–397. 2012.PubMed/NCBI
|
|
50
|
Konakci KZ, Bohle B, Blumer R,
Hoetzenecker W, Roth G, Moser B, Boltz-Nitulescu G, Gorlitzer M,
Klepetko W, Wolner E, et al: Alpha-Gal on bioprostheses: Xenograft
immune response in cardiac surgery. Eur J Clin Invest. 35:17–23.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Collins BH, Cotterell AH, McCurry KR,
Alvarado CG, Magee JC, Parker W and Platt JL: Cardiac xenografts
between primate species provide evidence for the importance of the
alpha-galactosyl determinant in hyperacute rejection. J Immunol.
154:5500–5510. 1995.PubMed/NCBI
|
|
52
|
Manji RA, Menkis AH, Ekser B and Cooper
DK: Porcine bioprosthetic heart valves: The next generation. Am
Heart J. 164:177–185. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Tazelaar HD, Byrne GW and McGregor CG:
Comparison of Gal and non-Gal-mediated cardiac xenograft rejection.
Transplantation. 91:968–975. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Lim HG, Choi SY, Yoon EJ, Kim SH and Kim
YJ: In vivo efficacy of alpha-galactosidase as possible promise for
prolonged durability of bioprosthetic heart valve using
alpha1,3-galactosyl-transferase knockout mouse. Tissue Eng Part A.
19:2339–2348. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
McGregor CG, Carpentier A, Lila N, Logan
JS and Byrne GW: Cardiac xenotransplantation technology provides
materials for improved bioprosthetic heart valves. J Thorac
Cardiovasc Surg. 141:269–275. 2011. View Article : Google Scholar
|
|
56
|
Mangold A, Szerafin T, Hoetzenecker K,
Hacker S, Lichtenauer M, Niederpold T, Nickl S, Dworschak M, Blumer
R, Auer J, et al: Alpha-Gal specific IgG immune response after
implantation of bioprostheses. Thorac Cardiovasc Surg. 57:191–195.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Nam J, Choi SY, Sung SC, Lim HG, Park SS,
Kim SH and Kim YJ: Changes of the structural and biomechanical
properties of the bovine pericardium after the removal of α-Gal
epitopes by decellularization and α-galactosidase treatment. Korean
J Thorac Cardiovasc Surg. 45:380–389. 2012. View Article : Google Scholar :
|
|
58
|
Kasimir MT, Rieder E, Seebacher G, Wolner
E, Weigel G and Simon P: Presence and elimination of the
xenoantigen gal (alpha1, 3) gal in tissue-engineered heart valves.
Tissue Eng. 11:1274–1280. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
McGregor CG, Davies WR, Oi K, Teotia SS,
Schirmer JM, Risdahl JM, Tazelaar HD, Kremers WK, Walker RC, Byrnew
GW, et al: Cardiac xenotransplantation: Recent preclinical progress
with 3-month median survival. J Thorac Cardiovasc Surg.
130:844–851. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Milland J, Christiansen D, Lazarus BD,
Taylor SG, Xing PX and Sandrin MS: The molecular basis for
galalpha(1,3)gal expression in animals with a deletion of the
alpha1,3galactosyltransferase gene. J Immunol. 176:2448–2454. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Puga Yung G, Schneider MK and Seebach JD:
Immune responses to alpha1,3 galactosyltransferase knockout pigs.
Curr Opin Organ Transplant. 14:154–160. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Stone KR, Abdel-Motal UM, Walgenbach AW,
Turek TJ and Galili U: Replacement of human anterior cruciate
ligaments with pig ligaments: A model for anti-non-gal antibody
response in long-term xenotransplantation. Transplantation.
83:211–219. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Park HM, Kim YW, Kim KJ, Kim YJ, Yang YH,
Jin JM, Kim YH, Kim BG, Shim H and Kim YG: Comparative N-linked
glycan analysis of wild-type and α1,3-galactosyltransferase gene
knock-out pig fibroblasts using mass spectrometry approaches. Mol
Cells. 38:65–74. 2015.
|
|
64
|
Macher BA and Galili U: The
Galalpha1,3Galbeta1,4GlcNAc-R (alpha-Gal) epitope: A carbohydrate
of unique evolution and clinical relevance. Biochim Biophys Acta.
1780:75–88. 2008. View Article : Google Scholar
|
|
65
|
Deriy L, Chen ZC, Gao GP and Galili U:
Expression of alpha-gal epitopes on HeLa cells transduced with
adenovirus containing alpha1,3galactosyltransferase cDNA.
Glycobiology. 12:135–144. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Naso F, Gandaglia A, Bottio T, Tarzia V,
Nottle MB, d'Apice AJ, Cowan PJ, Cozzi E, Galli C, Lagutina I, et
al: First quantification of alpha-Gal epitope in current
glutaraldehyde-fixed heart valve bioprostheses.
Xenotransplantation. 20:252–261. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Galili U: Conversion of tumors into
autologous vaccines by intra-tumoral injection of α-Gal glycolipids
that induce anti-Gal/α-Gal epitope interaction. Clin Dev Immunol.
2011(134020)2011. View Article : Google Scholar
|
|
68
|
Huang AY, Golumbek P, Ahmadzadeh M, Jaffee
E, Pardoll D and Levitsky H: Role of bone marrow-derived cells in
presenting MHC class I-restricted tumor antigens. Science.
264:961–965. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Manca F, Fenoglio D, Li Pira G, Kunkl A
and Celada F: Effect of antigen/antibody ratio on macrophage
uptake, processing, and presentation to T cells of antigen
complexed with polyclonal antibodies. J Exp Med. 173:37–48. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Abdel-Motal UM, Wigglesworth K and Galili
U: Intratumoral injection of alpha-gal glycolipids induces a
protective anti-tumor T cell response which overcomes Treg
activity. Cancer Immunol Immunother. 58:1545–1556. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Schweighoffer T, Schmidt W, Buschle M and
Birnstiel ML: Depletion of naive T cells of the peripheral lymph
nodes abrogates systemic antitumor protection conferred by IL-2
secreting cancer vaccines. Gene Ther. 3:819–824. 1996.PubMed/NCBI
|
|
72
|
Liu C1, Gosselin EJ and Guyre PM: Fc gamma
RII on human B cells can mediate enhanced antigen presentation.
Cell Immunol. 167:188–194. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
LaTemple DC, Abrams JT, Zhang SY and
Galili U: Increased immunogenicity of tumor vaccines complexed with
anti-Gal: studies in knockout mice for
alpha1,3galactosyltransferase. Cancer Res. 59:3417–3423.
1999.PubMed/NCBI
|
|
74
|
Tanida T, Tanemura M, Miyoshi E, Nagano H,
Furukawa K, Nonaka Y, Akita H, Hama N, Wada H, Kawamoto K, et al:
Pancreatic cancer immunotherapy using a tumor lysate vaccine,
engineered to express α-gal epitopes, targets pancreatic cancer
stem cells. Int J Oncol. 46:78–90. 2015.
|
|
75
|
Tanemura M, Miyoshi E, Nagano H, Eguchi H,
Taniyama K, Kamiike W, Mori M and Doki Y: Role of α-gal
epitope/anti-Gal antibody reaction in immunotherapy and its
clinical application in pancreatic cancer. Cancer Sci. 104:282–290.
2013. View Article : Google Scholar
|
|
76
|
Deguchi T, Tanemura M, Miyoshi E, Nagano
H, Machida T, Ohmura Y, Kobayashi S, Marubashi S, Eguchi H, Takeda
Y, et al: Increased immunogenicity of tumor-associated antigen,
mucin 1, engineered to express alpha-gal epitopes: A novel approach
to immunotherapy in pancreatic cancer. Cancer Res. 70:5259–5269.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Whalen GF, Sullivan M, Piperdi B, Wasseff
W and Galili U: Cancer immunotherapy by intratumoral injection of
α-gal glyco-lipids. Anticancer Res. 32:3861–3868. 2012.PubMed/NCBI
|
|
78
|
Galili U, Repik PM, Anaraki F,
Mozdzanowska K, Washko G and Gerhard W: Enhancement of antigen
presentation of influenza virus hemagglutinin by the natural human
anti-Gal antibody. Vaccine. 14:321–328. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Henion TR, Gerhard W, Anaraki F and Galili
U: Synthesis of alpha-gal epitopes on influenza virus vaccines, by
recombinant alpha 1,3galactosyltransferase, enables the formation
of immune complexes with the natural anti-Gal antibody. Vaccine.
15:1174–1182. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Abdel-Motal U, Wang S, Lu S, Wigglesworth
K and Galili U: Increased immunogenicity of human immunodeficiency
virus gp120 engineered to express Galalpha1-3Galbeta1-4GlcNAc-R
epitopes. J Virol. 80:6943–6951. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Hurwitz ZIR, Lalikos J and Galili U:
Accelerated porcine wound healing with a-Gal nanoparticles. Plast
Reconstr Surg. 129:242–251. 2012. View Article : Google Scholar
|
|
82
|
Galili U: Discovery of the natural
anti-Gal antibody and its past and future relevance to medicine.
Xenotransplantation. 20:138–147. 2013.PubMed/NCBI
|
|
83
|
Wigglesworth KM, Racki WJ, Mishra R,
Szomolanyi-Tsuda E, Greiner DL and Galili U: Rapid recruitment and
activation of macrophages by anti-Gal/α-Gal liposome interaction
accelerates wound healing. J Immunol. 186:4422–4432. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Galili U, Wigglesworth K and Abdel-Motal
UM: Accelerated healing of skin burns by anti-Gal/alpha-gal
liposomes interaction. Burns. 36:239–251. 2010. View Article : Google Scholar
|