Introduction
Lumbar intervertebral disc (IVD) degeneration is a
major cause of lower back pain (1); it may reduce the quality of life of
individuals (2) and causes
significant socioeconomic problems (3). During the process of degeneration,
IVDs undergo morphological and biochemical changes (4,5). A
decrease in water content is one of the characteristics of lumbar
IVD degeneration (6).
Aquaporins (AQPs) are a family of small, integral
membrane proteins which regulate the influx and outflow of water
and small molecules (7). AQP3 is
a member of the AQP family, and it acts as a water channel to
promote glycerol permeability and water transport across cell
membranes (8). It has previously
been noted that AQP3 immunoreactivity significantly decreased in
the nucleus pulposus (NP) and annulus fibrosus (AF) of aged rats
compared with young rats (9), and
the expression of AQP3 is reduced in the degenerated tissue of
human lumbar IVDs (10). However,
little is known about the effect of AQP3 on the pathogenesis of IVD
degeneration.
Wnt/β-catenin signaling plays an important role in
the progression of IVD degeneration (11–13). The activation of Wnt/β-catenin
signaling triggers the process of IVD degeneration, suppresses the
proliferation of NP cells and induces cellular senescence (14–16).
In the present study, we performed an in
vitro study using human NP cells (hNPCs) to evaluate the effect
of AQP3 on the progression of IVD degeneration for the first time,
to the best of our knowledge. Furthermore, this experiment aimed to
determine whether Wnt/β-catenin signaling is involved in the effect
of AQP3 on IVD degeneration
Materials and methods
Tissue samples
The present study was approved by the Ethics
Committee of The Second Affiliated Hospital of Harbin Medical
University (Harbin, China). Following the provision of informed
consent, patients with lumbar IVD degeneration (n=12) and patients
with spinal trauma (n=8) who underwent surgery at the Department of
Orthopedic Surgery, The Second Affiliated Hospital of Harbin
Medical University were enrolled in this study. The human NP
samples were harvested and immediately frozen in liquid nitrogen
for RNA and protein extraction at a later date.
Cell culture
hNPCs (ScienCell Research Laboratories, San Diego,
CA, USA) were maintained in Dulbecco's modified Eagle's medium
(DMEM) supplemented with 10% fetal bovine serum (FBS) (both from
Gibco Life Technologies, Carlsbad, CA, USA) at 37°C in a humidified
atmosphere of 5% CO2. Lithium chloride (LiCl) was
purchased from Sigma-Aldrich (St. Louis, MO, USA) and dissolved in
water at a concentration of 20 mM, to treat hNPCs for 24 h.
Cell transfection
The cells were seeded at 2.5×105
cells/well in a 6-well plate. Following overnight adherence, the
cells were transfected with 1 µg AQP3-pcDNA3.1 plasmid,
empty pcDNA3.1 vector, AQP3 siRNA or control siRNA (all were
obtained from Fitgene Biological Technology Co., Ltd., Guangzhou,
China) per well using Lipofectamine 2000 (Invitrogen Life
Technologies, Carlsbad, CA, USA) according to the manufacturer's
instructions. Cells in the blank group were not transfected with
any plasmids or siRNA. Briefly, plasmid DNA/siRNA and Lipofectamine
2000 (Invitrogen Life Technologies) were diluted in two independent
250 µl volumes of Opti-MEM I (Invitrogen Life Technologies).
After a 5-min incubation at room temperature, the DNA and the
Lipofectamine 2000 in Opti-MEM I were combined and incubated for an
additional 20 min to allow for the formation of DNA-Lipofectamine
2000 complexes. The DNA-Lipofectamine 2000 complexes were then
added to each well. Six hours post-transfection, the medium was
replaced with 2 ml of complete growth medium.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the tissues using
TRIzol (Invitrogen Life Technologies). Subsequently, 1 µg of
total RNA was reverse transcribed to cDNA using M-MLV reverse
transcriptase (Promega Corp., Madison, WI, USA). The qPCR reactions
were performed using the SYBR-Green PCR Master Mix (Applied
Biosystems Life Technologies; Thermo Fisher Scientific, Waltham,
MA, USA). The PCR primers were as follows: AQP3 sense,
5′-ccttcttgggtgctggaata-3′ and antisense,
5′-acacacacgataagggaggc-3′; and β-actin sense,
5′-aagtactccgtgtggatcgg-3′ and antisense,
5′-caccttcaccgttccagttt-3′. The PCR cycles were as follows: initial
denaturation at 95°C for 5 min, followed by 40 cycles of 94°C for
20 sec, 60°C for 20 sec and 72°C for 5 min, and a final extension
at 72°C for 5 min. The PCR products were confirmed by
electrophoresis on 2% agarose gel. All reactions were performed in
duplicate on an ABI PRISM 7900HT Sequence Detection system (Applied
Biosystems Life Technologies). The expression of mRNA was defined
from the threshold cycle, and the final results were expressed as
fold-changes between the patients with lumbar IVD degeneration and
the patients with spinal trauma after normalization to the internal
control β-actin.
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay
In the present study, an MTT assay was carried out
using a cell proliferation assay kit (Boster Inc., Wuhan, China).
Briefly, 104 cells were plated on 96-well plates.
Following treatment, 20 µl MTT solution was added to each
well and incubated for 4 h at 37°C, followed by the addition of 100
µl dimethyl sulfoxide (DMSO; Sigma-Aldrich) to solubilize
the formazan. Finally, absorbance was recorded at 570 nm using a
Multiskan Ascent 354 microplate reader (Thermo Labsystems, Waltham,
MA, USA). The cell viability results of three independent
experiments were normalized to the cells treated with medium
only.
Western blot analysis
Proteins were prepared using a total protein
extraction kit (KeyGen Biotech, Nanjing, China) according to the
manufacturer's instructions. The proteins were resolved on a sodium
dodecyl sulfate (SDS) polyacrylamide gel and electrotransferred to
a nitrocellulose membrane (EMD Millipore, Billerica, MA, USA).
After blocking with 5% non-fat milk in TBST at 4°C overnight, the
membranes were incubated with the following specific antibodies:
rabbit polyclonal to aquaporin 3 (ab125219; 1:400); rabbit
polyclonal to aggrecan (ab36861; 1:500); rabbit polyclonal to a
disintegrin and metalloproteinase with thrombospondin motifs
(ADAMTS)4 (ab45038; 1:1,000); rabbit polyclonal to ADAMTS5
(ab41037; 1:400); rabbit polyclonal to Axin 2 (ab32197; 1:500) (all
from Abcam, Cambridge, MA, USA); rabbit polyclonal to β-catenin
(sc-7199; 1:800; Santa Cruz Biotechnology, Inc., Santa Cruz, CA,
USA); rabbit polyclonal to β-actin (RS-0061R; 1:1,000; Shanghai
Ruiqi Biological Technology Co., Ltd., Shanghai, China) followed
with goat anti-rabbit IgG polyclonal antibody (020002-G; 1:5,000;
Beijing CellChip Biotechnology, Beijing, China). Immunolabeling was
undertaken using an ECL western blotting kit (Pierce Biotechnology,
Inc., Rockford, IL, USA).
Statistical analysis
Data are presented as the means ± standard deviation
(SD) from at least three independent experiments conducted in
triplicate. The differences between two groups were analyzed by
Student's t-test. P-values <0.05 were considered to indicate a
statistically significant difference.
Results
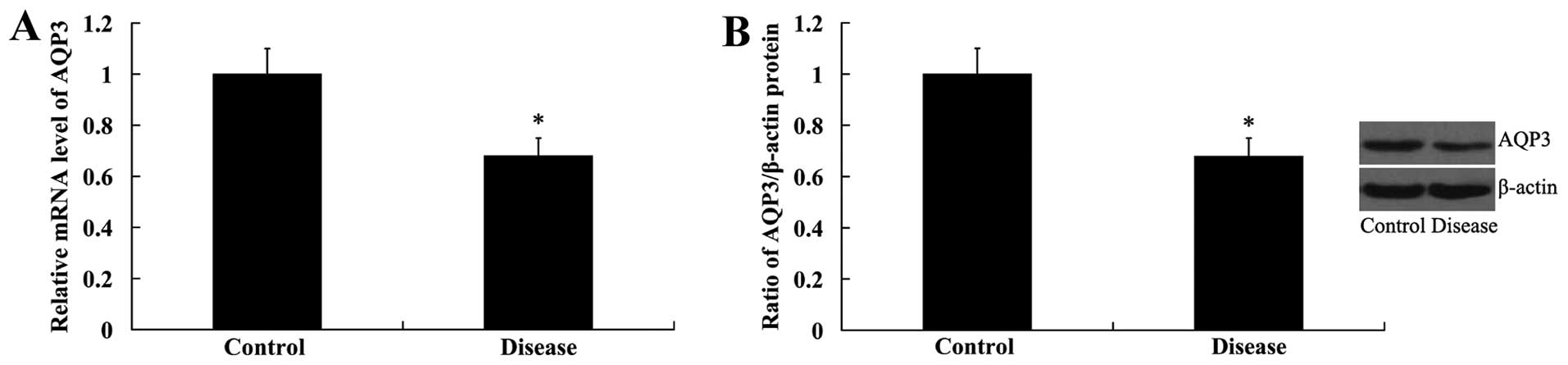
AQP3 expression in the NP tissue
samples
AQP3 expression in the NP tissues was examined at
the mRNA and protein levels. RT-qPCR results revealed that AQP3
mRNA expression was significantly decreased in the samples from
patients with IVD degenereration compared with that in the control
samples (the patients with spinal trauma). Similar to the mRNA
expression results, AQP3 protein expression was also lower in the
NP tissues from patients with IVD degeneration than those of
patients in the control group (the patients with spinal trauma)
(Fig. 1).
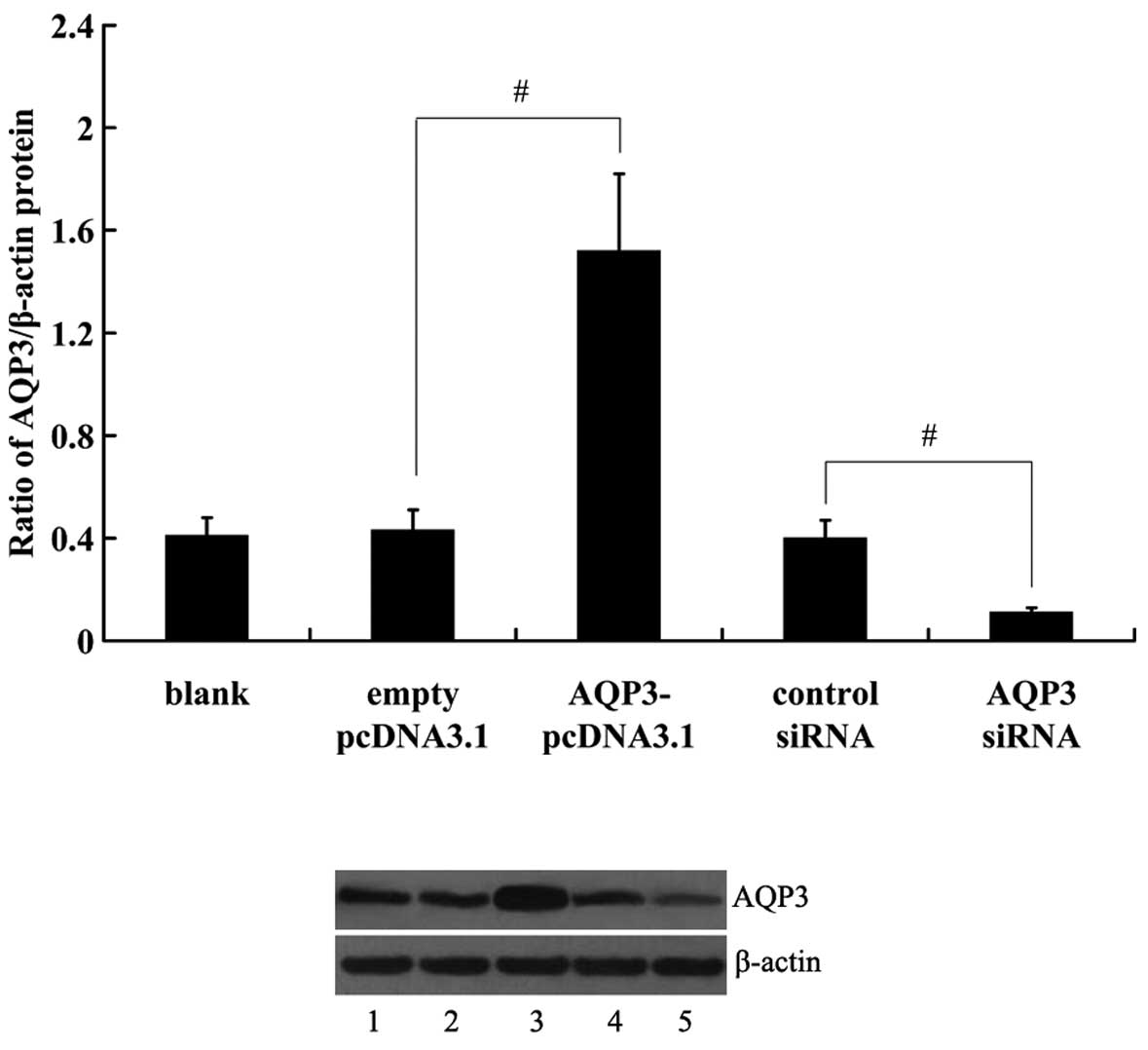
Expression of AQP3 in human NP cells
following transfection with the AQP3-pcDNA3.1 plasmid and AQP3
siRNA
AQP3-pcDNA3.1 plasmid or AQP3-siRNA were transfected
into hNPCs to cause overexpression or suppression of AQP3. Empty
pcDNA3.1 vector and control siRNA exerted no significant effects on
AQP3 expression (Fig. 2).
However, AQP3 protein levels in the hNPCs were significantly
increased by transfection with the AQP3-pcDNA3.1 plasmid.
Transfection with the AQP3 siRNA resulted in a significant decrease
in AQP3 expression.
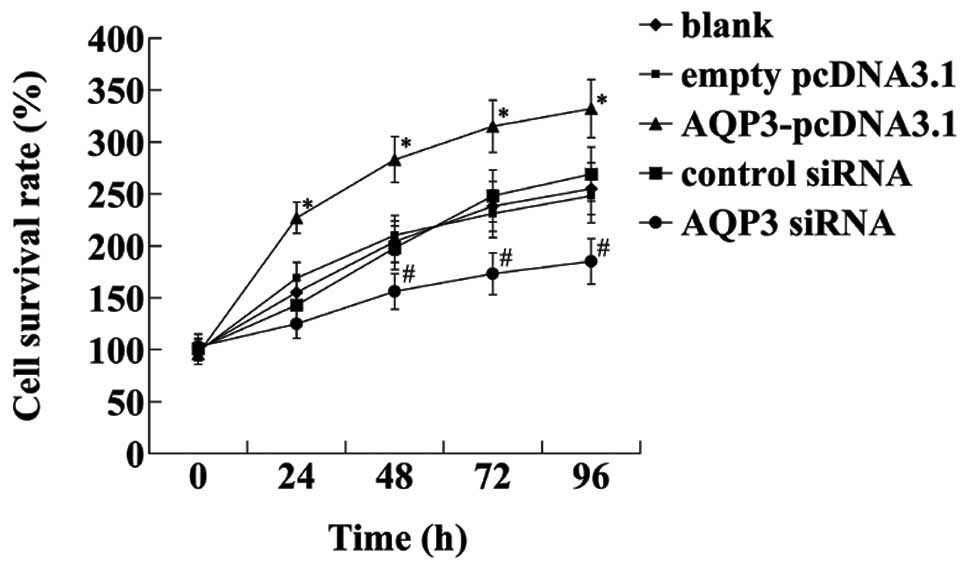
Effect of AQP3 expression on hNPC
proliferation
An MTT assay was performed to determine the effect
of AQP3 overexpression on hNPC proliferation. AQP3 overexpression
significantly increased hNPC proliferation (Fig. 3). In order to confirm the effect
of endogenous AQP3 on hNPC proliferation, we examined whether AQP3
knockdown by AQP3 siRNA affected the survival rate of hNPCs. It was
demonstrated that AQP3 knockdown exerted a significant inhibitory
effect on hNPC proliferation.
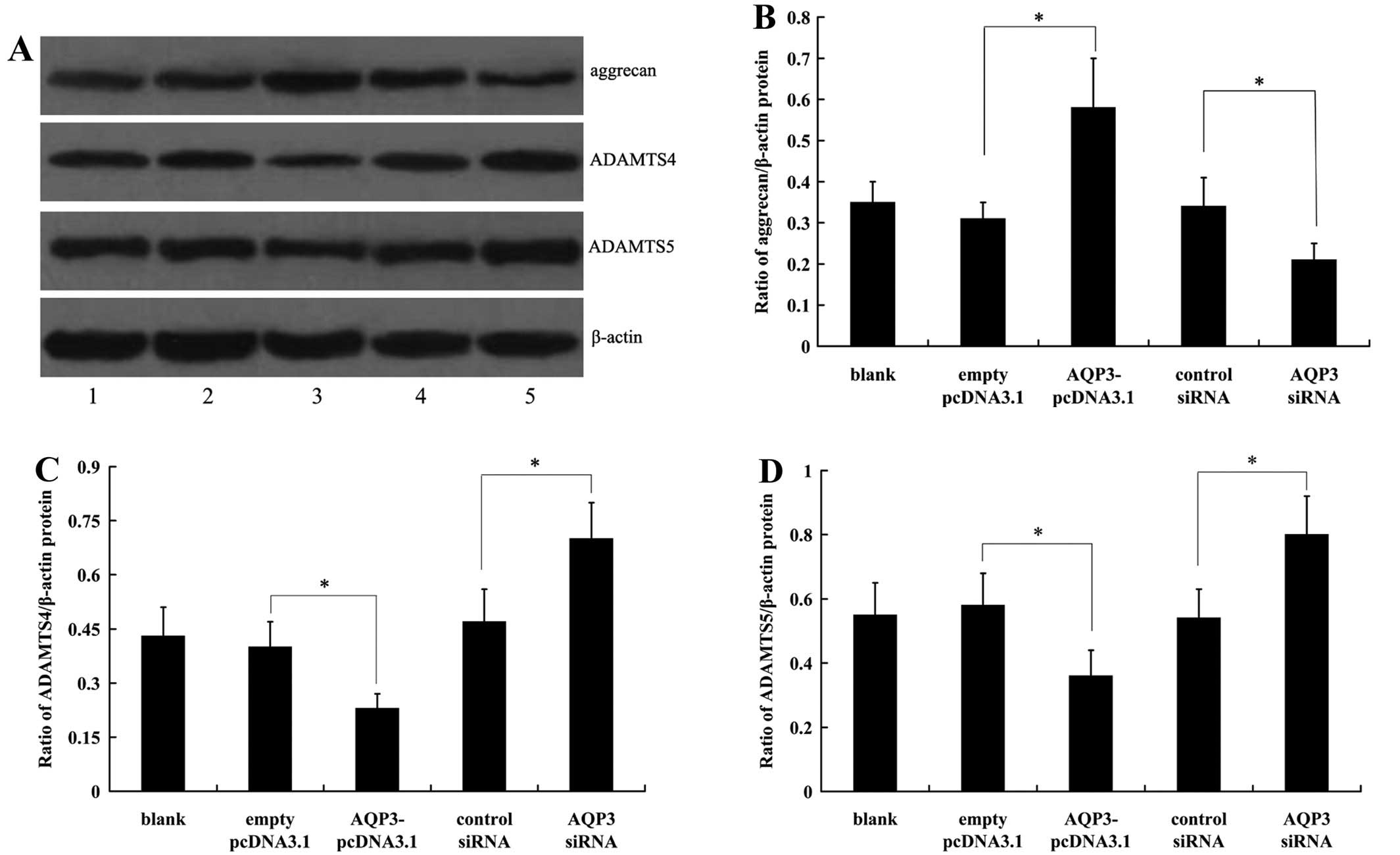
Effect of AQP3 expression on
extracellular matrix (ECM) degradation
The effect of AQP3 overexpression on ECM degradation
was investigated by western blot analysis. The results demonstrated
that aggrecan expression was increased in AQP3 overexpressing-hNPCs
(Fig. 4A and B). In addition, the
expression of the enzymes which are involved in the matrix
degrading process, ADAMTS4 and ADAMTS5, was significantly decreased
in hNPCs following overexpression of AQP3 (Fig. 4A, C and D).
The effect of endogenous AQP3 on ECM degradation was
also determined by detecting the expression of aggrecan, ADAMTS4
and ADAMTS5 in hNPCs. We demonstrated that the expression level of
aggrecan was significantly decreased in hNPCs following
transfection with the AQP3 siRNA (Fig. 4A and B). Moreover, knockdown of
AQP3 expression in hNPCs led to the increased expression of ADAMTS4
and ADAMTS5 (Fig. 4A, C and
D).
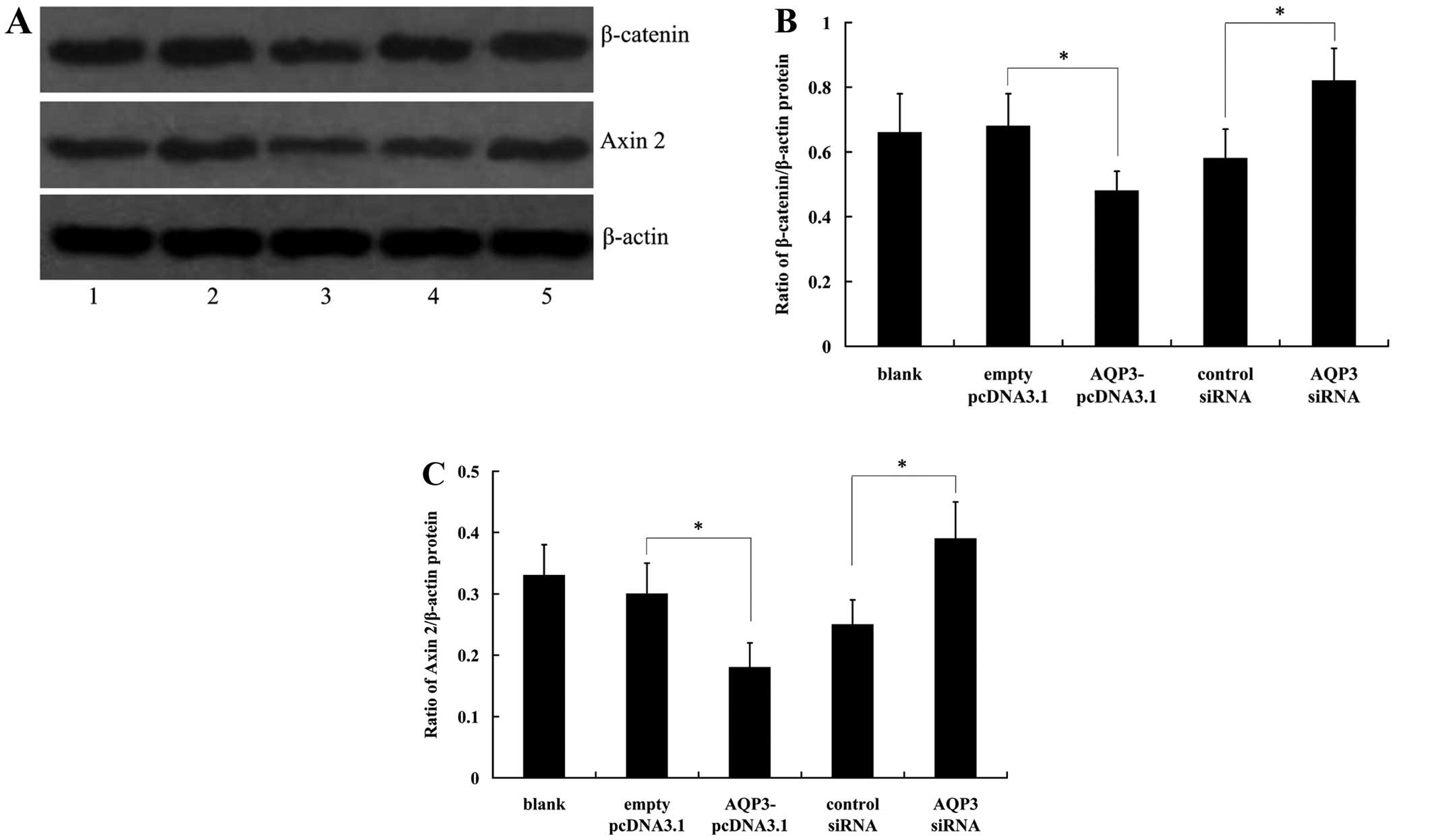
Effect of AQP3 on Wnt/β-catenin
signaling
To investigate the effect of AQP3 overexpression on
Wnt/β-catenin signaling, β-catenin and Axin 2 protein expression
was measured in hNPCs. The results from western blot analysis
revealed that AQP3 overexpression significantly decreased the
expression of β-catenin and Axin 2. Knockdown of AQP3 expression in
hNPCs induced a significant increase in β-catenin and Axin 2
protein levels (Fig. 5).
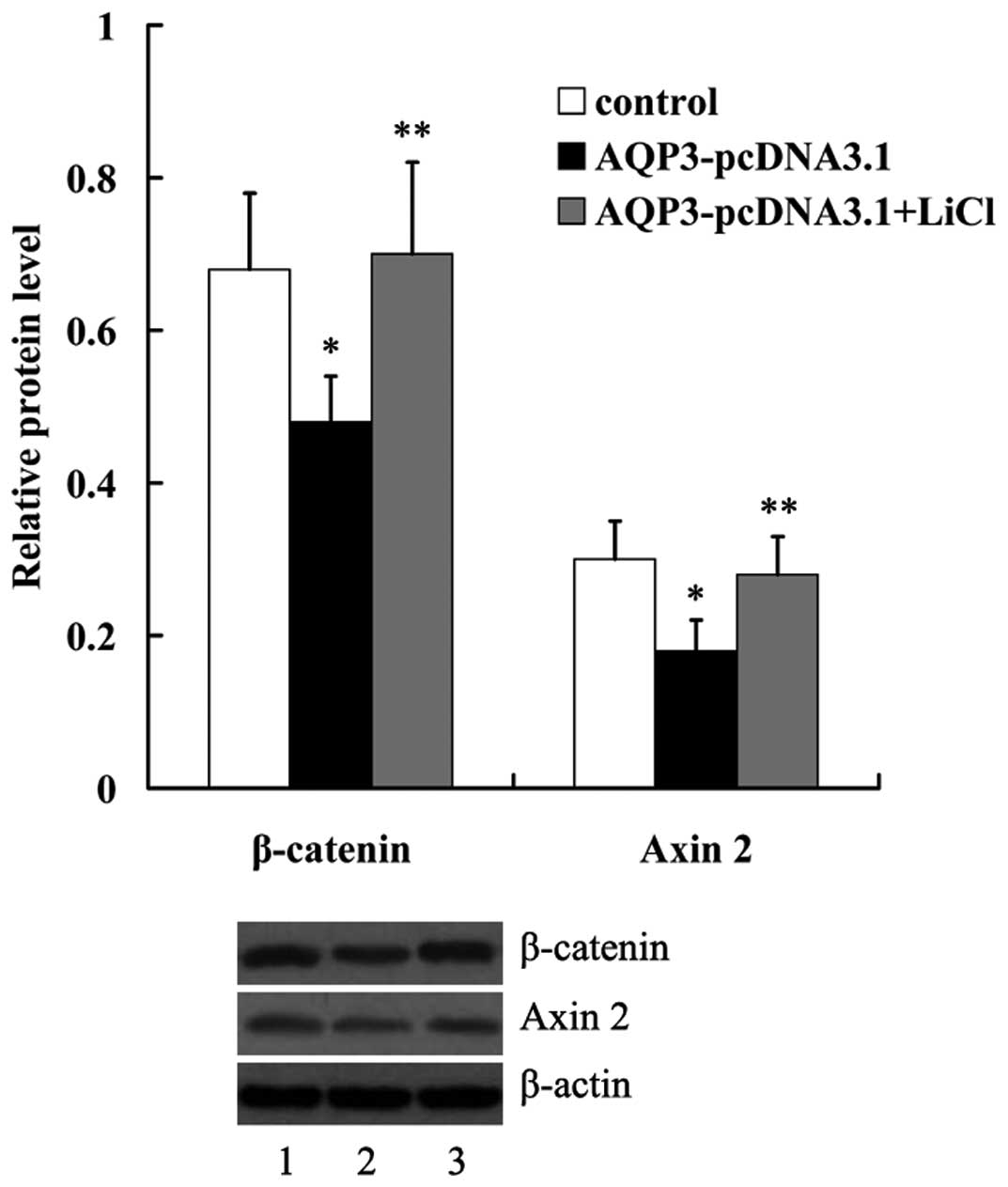
Wnt/β-catenin signaling activation
reverses the effect of AQP3 on hNPC proliferation and ECM
degradation
To investigate whether Wnt/β-catenin signaling was
involved in mediating the effect of AQP3 on hNPC hNPC proliferation
and ECM degration, hNPCs overexpressing AQP3 were treated with
LiCl, a known activator of Wnt signaling. As shown in Fig. 6, in hNPCs the protein levels of
β-catenin and Axin 2, which had been downregulated by AQP3
overexpression, were significantly increased by incubation with 20
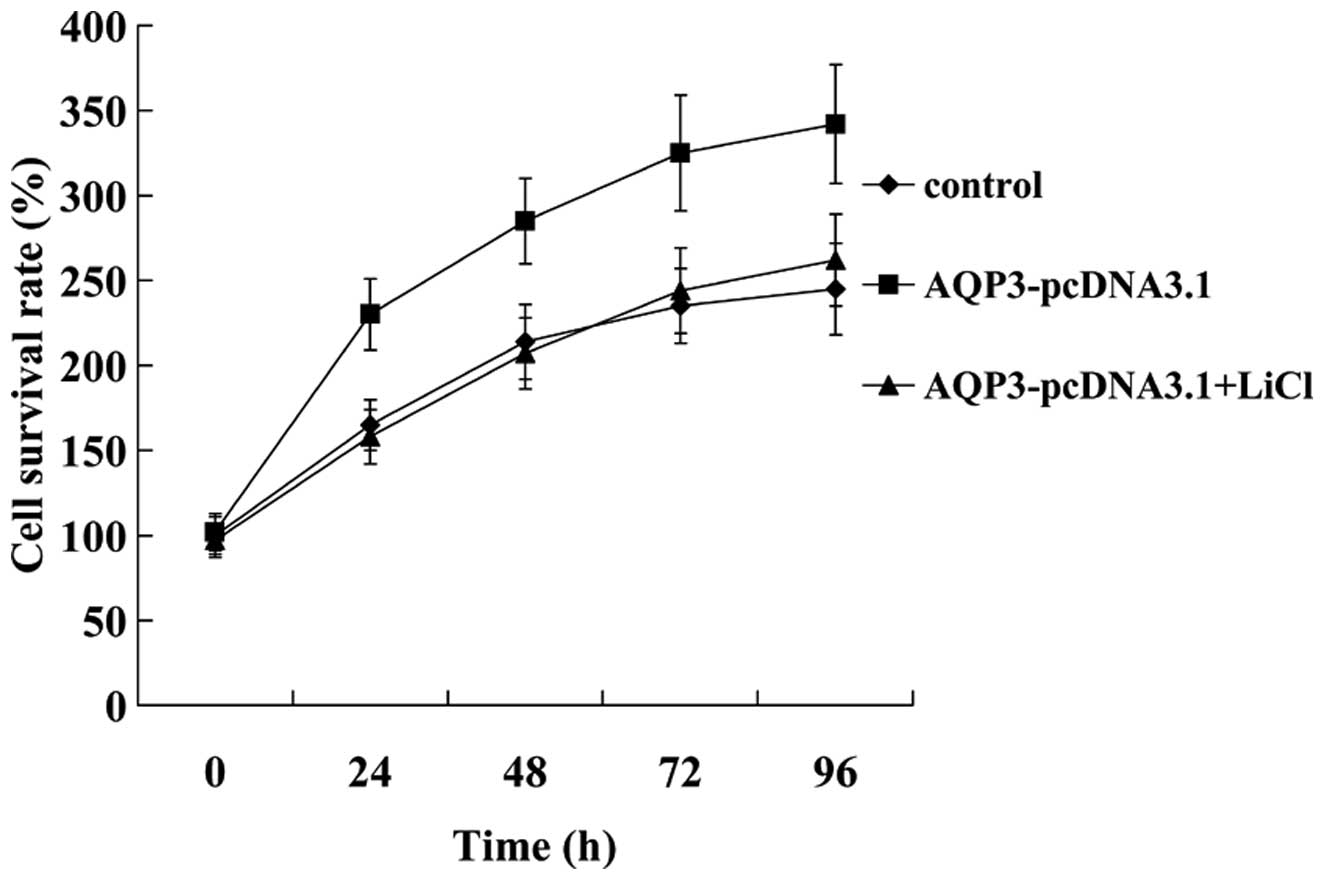
mM LiCl for 24 h. An MTT assay revealed that the effect of AQP3
overexpression on promoting hNPC proliferation was attenuated by
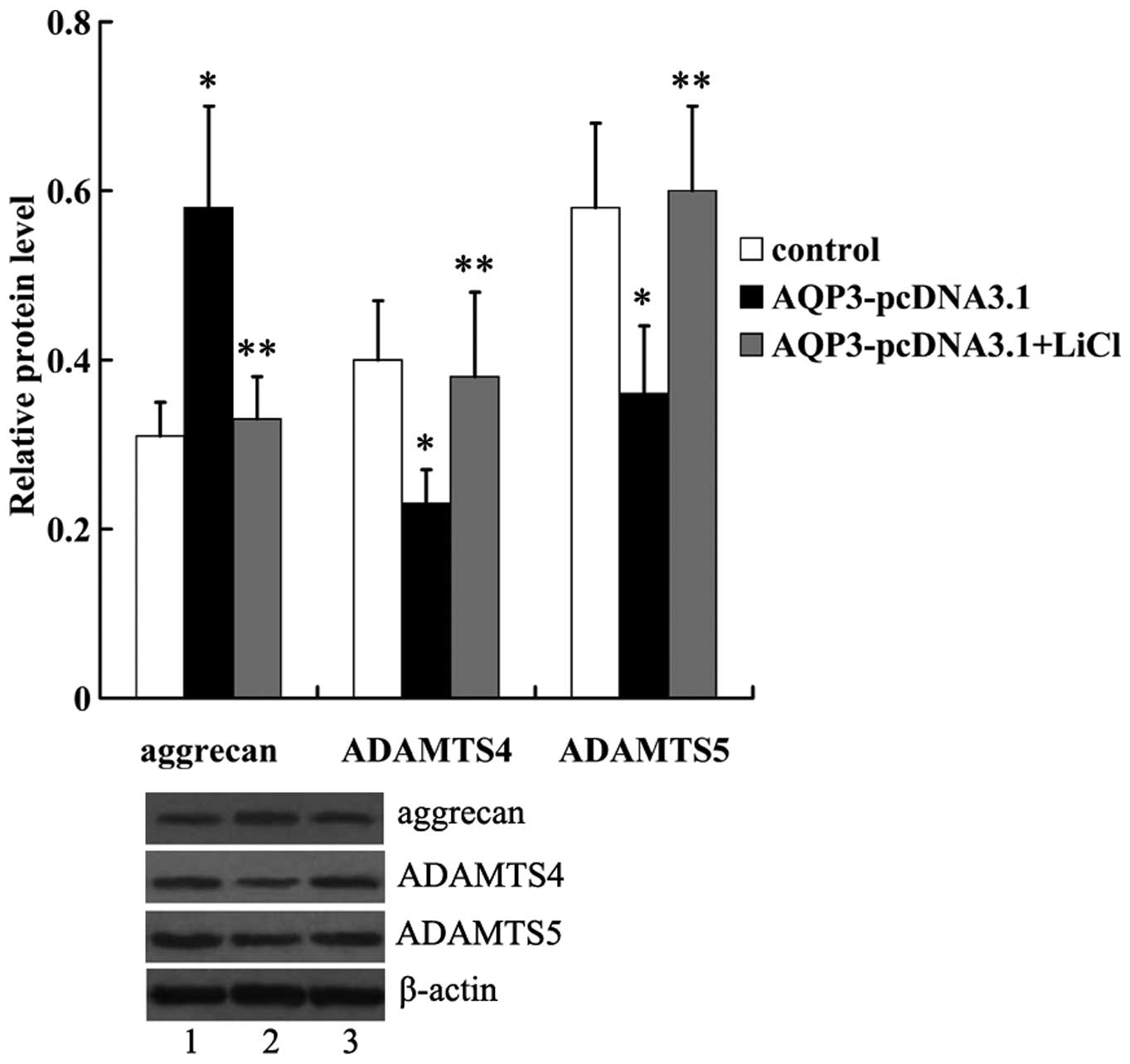
LiCl (Fig. 7). In addition, we
noted that the altered expression of aggrecan, ADAMTS4 and ADAMTS5
in AQP3-overexpressing hNPCs was reversed by LiCl (Fig. 8).
Discussion
AQP3 is expressed in multiple epithelial tissues,
knee articular cartilage, subchondral osteoblasts, synovium and
IVDs (17,18). In the present study, we found that
AQP3 was downregulated in the NP tissue samples of patients with
lumbar IVD degeneration. This finding is consistent with the report
by Ufuk et al which found that AQP-3 immunoreactivity
significantly decreased in the NP of aged rats compared with young
rats (9).
The abnormal expression of AQP3 has been linked to
various disease pathologies (19,20). In the present study, in
vitro gain-of-function and loss-of-function experiments were
performed to verify the effect of AQP3 on IVD degeneration. The IVD
consists of three distinct regions: the vertebral endplates (EPs),
the AF and the NP (21). NP cells
secrete a complex ECM which mainly consists of proteoglycan and
fibrillar collagen. The most important elements of disc
degeneration are the decline in the number of disc cells and the
extensive degradation of the ECM (22,23). Aggrecan is a type of proteoglycan,
and it is the major macromolecular component of ECM. The synthesis
and degradation of aggrecan are in dynamic equilibrium, maintaining
the normal structure and biomechanical function of the IVD
(24). ADAMTS4 and ADAMTS5 are
the two major aggrecanases, which have the ability to degrade
aggrecan (25). It has been
demonstrated that the expression of ADAMTSs is increased in the NP
of human degenerative disc disease (26).
Our data demonstrated that AQP3 promoted cell
proliferation, as determined by an MTT assay. In addition, we found
that AQP3 inhibited ECM degradation in hNPCs, as evidenced by the
increased expression of aggrecan, and the decreased expression of
ADAMTS4 and ADAMTS5. These results indicate that AQP3 exerts
protective effects against IVD degeneration.
Wnt/β-catenin signaling regulates the proliferation
and differentiation of various cell types; therefore, it is
involved in tissue degeneration and regeneration (27,28). Several research groups have
suggested that Wnt/β-catenin signaling contributes to the
pathogenesis of IVD degeneration. It has previously been
demonstrated that the levels of β-catenin-positive cells increased
as IVD degeneration progressed (29). In a previous study, it was noted
that treatment of NP cells with a Wnt/β-catenin activator resulted
in decreased cell proliferation (15). In addition, activation of
Wnt/β-catenin signaling induced the expression of matrix
metalloproteinases and TGF-β in NP cells, leading to an increased
breakdown of the matrix, thereby promoting IVD degeneration
(16). In the present study, we
noted that in hNPCs, Wnt/β-catenin signaling was regulated by AQP3.
Activation of the Wnt/β-catenin signaling pathway attenuated the
effect of AQP3 on cell proliferation and ECM degradation in hNPCs.
These findings suggest that AQP3 exerts protective effects against
IVD degeneration through the inhibition of Wnt/β-catenin
signaling.
In conclusion, we have provided in vitro
evidence that AQP3 exerts protective effects against IVD
degeneration, and these are effected, at least partially, through
the inhibition of Wnt/β-catenin signaling. This study has improved
our understanding of the mechanisms underlying the effects of AQP3
in the progress of IVD degeneration, and suggested a potential
strategy for the treatment of IVD degeneration.
References
|
1
|
Podichetty VK: The aging spine: the role
of inflammatory mediators in intervertebral disc degeneration. Cell
Mol Biol (Noisy-le-grand). 53:4–18. 2007.
|
|
2
|
Speed C: Low back pain. BMJ.
328:1119–1121. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Takahashi K, Aoki Y and Ohtori S:
Resolving discogenic pain. Eur Spine J. 17(Suppl 4): 428–431. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Weber KT, Jacobsen TD, Maidhof R,
Virojanapa J, Overby C, Bloom O, Quraishi S, Levine M and Chahine
NO: Developments in intervertebral disc disease research:
pathophysiology, mechanobiology, and therapeutics. Curr Rev
Musculoskelet Med. 8:18–31. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang SZ, Rui YF, Lu J and Wang C: Cell and
molecular biology of intervertebral disc degeneration: current
understanding and implications for potential therapeutic
strategies. Cell Prolif. 47:381–390. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Isherwood I, Prendergast DJ, Hickey DS and
Jenkins JP: Quantitative analysis of intervertebral disc structure.
Acta Radiol Suppl. 369:492–495. 1986.PubMed/NCBI
|
|
7
|
Agre P: The aquaporin water channels. Proc
Am Thorac Soc. 3:5–13. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zeuthen T and Klaerke DA: Transport of
water and glycerol in aquaporin 3 is gated by H+. J Biol
Chem. 274:21631–21636. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Taş U, Caylı S, Inanır A, Ozyurt B, Ocaklı
S, Karaca Zİ and Sarsılmaz M: Aquaporin-1 and aquaporin-3
expressions in the intervertebral disc of rats with aging. Balkan
Med J. 29:349–353. 2012.
|
|
10
|
Li SB, Yang KS and Zhang YT: Expression of
aquaporins 1 and 3 in degenerative tissue of the lumbar
intervertebral disc. Genet Mol Res. 13:8225–8233. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ye S, Wang J, Yang S, Xu W, Xie M, Han K,
Zhang B and Wu Z: Specific inhibitory protein Dkk-1 blocking
Wnt/β-catenin signaling pathway improve protectives effect on the
extracellular matrix. J Huazhong Univ Sci Technolog Med Sci.
31:657–662. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hiyama A, Yokoyama K, Nukaga T, Sakai D
and Mochida J: A complex interaction between Wnt signaling and
TNF-α in nucleus pulposus cells. Arthritis Res Ther. 15:R1892013.
View Article : Google Scholar
|
|
13
|
Kong J, Ma X, Wang T, Ma J, Tian P, Han C,
Zang J, Li P and Jiang H: Research progress of Wnt/beta-catenin and
nuclear factor-kappa B pathways and their relevance to
intervertebral disc degeneration. Zhongguo Xiu Fu Chong Jian Wai Ke
Za Zhi. 27:1523–1528. 2013.In Chinese.
|
|
14
|
Hiyama A, Sakai D, Arai F, Nakajima D,
Yokoyama K and Mochida J: Effects of a glycogen synthase kinase-3β
inhibitor (LiCl) on c-myc protein in intervertebral disc cells. J
Cell Biochem. 112:2974–2986. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hiyama A, Sakai D, Tanaka M, Arai F,
Nakajima D, Abe K and Mochida J: The relationship between the
Wnt/β-catenin and TGF-β/BMP signals in the intervertebral disc
cell. J Cell Physiol. 226:1139–1148. 2011. View Article : Google Scholar
|
|
16
|
Hiyama A, Sakai D, Risbud MV, Tanaka M,
Arai F, Abe K and Mochida J: Enhancement of intervertebral disc
cell senescence by WNT/β-catenin signaling-induced matrix
metalloproteinase expression. Arthritis Rheum. 62:3036–3047. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mobasheri A, Wray S and Marples D:
Distribution of AQP2 and AQP3 water channels in human tissue
microarrays. J Mol Histol. 36:1–14. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hagiwara K, Shinozaki T, Matsuzaki T,
Takata K and Takagishi K: Immunolocalization of water channel
aquaporins in human knee articular cartilage with intact and early
degenerative regions. Med Mol Morphol. 46:104–108. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Meng JH, Ma XC, Li ZM and Wu DC:
Aquaporin-1 and aquaporin-3 expressions in the temporo-mandibular
joint condylar cartilage after an experimentally induced
osteoarthritis. Chin Med J (Engl). 120:2191–2194. 2007.
|
|
20
|
Shen L, Zhu Z, Huang Y, Shu Y, Sun M, Xu
H, Zhang G, Guo R, Wei W and Wu W: Expression profile of multiple
aquaporins in human gastric carcinoma and its clinical
significance. Biomed Pharmacother. 64:313–318. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sive JI, Baird P, Jeziorsk M, Watkins A,
Hoyland JA and Freemont AJ: Expression of chondrocyte markers by
cells of normal and degenerate intervertebral discs. Mol Pathol.
55:91–97. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Roughley PJ, Alini M and Antoniou J: The
role of proteoglycans in aging, degeneration and repair of the
intervertebral disc. Biochem Soc Trans. 30:869–874. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
David G, Ciurea AV, Mitrica M and Mohan A:
Impact of changes in extracellular matrix in the lumbar
degenerative disc. J Med Life. 4:269–274. 2011.
|
|
24
|
Le Maitre CL, Pockert A, Buttle DJ,
Freemont AJ and Hoyland JA: Matrix synthesis and degradation in
human intervertebral disc degeneration. Biochem Soc Trans.
35:652–655. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tian Y, Yuan W, Fujita N, Wang J, Wang H,
Shapiro IM and Risbud MV: Inflammatory cytokines associated with
degenerative disc disease control aggrecanase-1 (ADAMTS-4)
expression in nucleus pulposus cells through MAPK and NF-κB. Am J
Pathol. 182:2310–2321. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huang M, Wang HQ, Zhang Q, Yan XD, Hao M
and Luo ZJ: Alterations of ADAMTSs and TIMP-3 in human nucleus
pulposus cells subjected to compressive load: Implications in the
pathogenesis of human intervertebral disc degeneration. J Orthop
Res. 30:267–273. 2012. View Article : Google Scholar
|
|
27
|
Smolders LA, Meij BP, Onis D, Riemers FM,
Bergknut N, Wubbolts R, Grinwis GC, Houweling M, Groot Koerkamp MJ,
van Leenen D, et al: Gene expression profiling of early
intervertebral disc degeneration reveals a down-regulation of
canonical Wnt signaling and caveolin-1 expression: implications for
development of regenerative strategies. Arthritis Res Ther.
15:R232013. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sanges D, Romo N, Simonte G, Di Vicino U,
Tahoces AD, Fernández E and Cosma MP: Wnt/β-catenin signaling
triggers neuron reprogramming and regeneration in the mouse retina.
Cell Rep. 4:271–286. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Iwata M, Aikawa T, Hakozaki T, Arai K,
Ochi H, Haro H, Tagawa M, Asou Y and Hara Y: Enhancement of Runx2
expression is potentially linked to β-catenin accumulation in
canine intervertebral disc degeneration. J Cell Physiol.
230:180–190. 2015. View Article : Google Scholar
|