Introduction
Hepatocellular carcinoma (HCC) is one of the most
common malignancies affecting the liver and the third and fifth
leading cause of cancer-related mortality worldwide in males and
females, respectively (1) The
incidence of HCC has increased in recent years; however,
satisfactory curative effects have not yet been achieved (2). Thus, it is important to elucidate
the precise molecular mechanisms responsible for the development of
HCC, and to identify novel therapeutic targets (2).
The heat shock proteins (HSPs) are highly conserved
molecular chaperones typically expressed in response to
environmental stress factors, including invading pathogens, toxins
and heat (3). Their principal
function is to prevent protein denaturation and misfolding. The
HSPs have been classified into the following 5 HSP families,
according to molecular mass and homology: small HSPs (sHSPs)
include HSP100, HSP90, HSP70 and HSPP60 (3,4).
HSP90 plays an important role in the metastasis, invasion and
vascularization of tumors, particularly in time course of cell
cycles, the proliferation of tumor cells and in maintaining the
function of a number of carcinogenic proteins involved in the
signal transduction pathway of cell apoptosis (5,6).
HSP90 has been found to be overexpressed in HCC (7). Yano et al (8) demonstrated that a high HSP90
expression significantly correlated with tumor size, lymph node
metastasis and a poor prognosis in breast cancer. In our previous
study, we demonstrated that a high HSP90 expression contributed to
the more aggressive phenotypes observed in colorectal cancer (CRC)
(9). However, the clinical
significance of HSP90 in predicting prognosis, as well as the
mechanisms responsible for the tumor-promoting effects of HSP90
remain unknown.
Hypoxia is a condition in which tissues are starved
of oxygen. It is a key characteristic of most tumor environments,
contributing to radioresistance, chemoresistance, metastasis,
angiogenesis, resistance to cell death and altered metabolism and
genomic instability (10).
Hypoxia-inducible factor (HIF), a transcription factor which plays
an important role in tumorigenesis, may induce angiogenesis and
drug-resistance (11). HIF-1 is a
heterodimer composed of an O2-regulated HIF-1α subunit
and a constitutively expressed HIF-1β subunit (11). It has been demonstrated that
HIF-1α regulates glycolysis, tumor angiogenesis and the
invasiveness of tumors (12–14). However, the mechanisms governing
HIF-1α protein stability remain poorly understood.
In the present study, we confirmed that HSP90 is an
independent prognostic factor for predicting the overall 3-year
survival of patients with HCC. HSP90 serves as a tumor promoter by
inhibiting apoptosis and promoting the growth of HCC cells. In HCC
tissues, HSP90 protein expression positively correlated with HIF-1α
protein expression. In addition, HSP90 interacted with HIF-1α and
inhibited HIF-1α ubiquitination, ultimately leading to HIF-1α
aggregation. Notably, the antitumor effects of HSP90 siRNA were
partially abrogated by the upregulation of HIF-1α expression in
vitro and in vivo. Our results demonstrate that HSP90
may target HIF-1α by inhibiting its ubiquitination and proteasomal
degradation, and inducing HCC cell growth and tumor
progression.
Materials and methods
Clinical samples, expression vectors and
cell lines
A total of 60 HCC tissue samples and paired normal
tumor-adjacent tissue samples (>1.5 cm distance from the margin
of the resection) were obtained and used after obtaining written
informed consent from patients at the Department of Hepatobiliary
Surgery at Zhejiang Provincial People's Hospital (Hangzhou, China)
between 2007 and 2010, with a median follow-up time of 32.5 months
(patients were followed-up for 3 years; however, as some patients
died before the 3-year period, the median follow-up time is 32.5
months). Prior to surgery, no patients had received any
radiotherapy, chemotherapy or radiofrequency ablation treatments.
The clinicopathological data and demographic characteristics of the
patients are presented in Table
I. The age of the patients ranged from 35–71 years (median, 52
years). Tumor tissue and matched normal tumor-adjacent tissue
specimens were collected and immediately stored in paraformaldehyde
for immunohistochemistry (IHC), as previousy described (15). The Ethics Committee of the
Provincial People's Hospital approved all the study protocols
according to the 1975 Declaration of Helsinki.
 | Table ICorrelation between heat shock
protein (HSP)90 expression and clinicopathological characteristics
in hepatocellular carcinoma (HCC). |
Table I
Correlation between heat shock
protein (HSP)90 expression and clinicopathological characteristics
in hepatocellular carcinoma (HCC).
| Clinicopathological
characteristics | Total no. of
patients n=60 | No. of patients
|
|---|
| HSP90-positive | HSP90-negative | P-value |
|---|
| Age (years) |
| <50 | 16 | 4 | 12 | 0.518 |
| ≥50 | 44 | 16 | 28 | |
| Gender |
| Male | 38 | 14 | 24 | 0.353 |
| Female | 22 | 6 | 16 | |
| HBV |
| Absent | 22 | 8 | 14 | 0.540 |
| Present | 38 | 12 | 26 | |
| Serum AFP level
(ng/ml) |
| <400 | 10 | 4 | 6 | 0.694 |
| ≥400 | 50 | 16 | 34 | |
| Tumor size
(cm) |
| <5 | 32 | 11 | 21 | 0.680 |
| ≥5 | 28 | 9 | 19 | |
| Cirrhosis |
| Absent | 11 | 7 | 4 | 0.201 |
| Present | 49 | 13 | 36 | |
| PVTT |
| Absent | 18 | 11 | 7 | 0.004a |
| Present | 42 | 9 | 33 | |
| Edmondson-Steiner
grading |
| I+II | 18 | 6 | 12 | 0.006a |
| III+IV | 42 | 30 | 12 | |
| TNM tumor
stage |
| I+II | 36 | 17 | 19 | 0.010a |
| III+IV | 24 | 3 | 21 | |
The cell lines, Hep3B, HepG2 and 293T cells were all
obtained from the Institute of Biochemistry and Cell Biology,
Chinese Academy of Sciences (Shanghai, China). All the cells were
maintained in Dulbecco's modified Eagle's medium (DMEM; Gibco,
Grand Island, NY, USA) supplemented with 10% fetal bovine serum
(FBS; Gibco) with 100 units/ml penicillin and 100 µg/ml
streptomycin (Sigma, St. Louis, MO, USA) and cultured in a
humidified 5% CO2 incubator at 37°C, as previously
described (15).
Control siRNA (sc-37007) and siRNA against HSP90α/β
(HSP90α/β siRNA, sc-35610) were obtained from Santa Cruz
Biotechnology (Santa Cruz, CA, USA). The retroviral vectors,
pMMP-Flag-HSP90 and pMMP-HA-HIF-1α, were generated by inserting the
respective cDNA into pMMP (Addgene). All constructs were confirmed
by western blot analysis and sequencing analysis. The day prior to
transfection, 5–6×106 293T cells were seeded in 100-mm
dishes, as previously described (15). Three plasmids, 1.5 µg
pMD.MLV, 0.5 µg pVSV.G and 2 µg of the relevant
retroviral vectors, were transfected into the cells using Effectene
Transfection reagent (Qiagen, Valencia, CA, USA), as previously
described (15). The medium
containing the retroviruses was collected 72 h following
transfection. Viral transduction was performed by incubating the
cells with the viral supernatant (25%) supplemented with Polybrene
(8 µg/ml; Santa Cruz Biotechnology) overnight at 37°C, as
previously described (15).
Further experiments were performed 72 h after viral transduction,
as previously described (15).
Immunohistochemical analysis
IHC was performed on 5-µm-thick sections from
formalin-fixed, paraffin-embedded tissue samples applied to coated
slides, as described in a previous study (16). The following antibodies were used
together with a streptavidin-peroxidase (SP) conjugate (SP-IHC
method): HSP90 (ab13492; 1:150) and HIF-1α (ab85886; 1:200) (both
from Abcam, Cambridge, MA, USA). IHC was performed as previously
described (17). The percentage
of positive tumor cells or hepatocytes was graded as per the
following criteria: 0, <10%; 1, 10–30%; 2, 30–50%; 3, >50%,
as previously described (18).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
The following primers were used: HSP90 sense,
5′-ATGGCAGCAAAGAAACAC-3′ and antisense, 5′-GTATC
ATCAGCAGTAGGGTCA-3′; and HIF-1α sense, 5′-GAACC TGATGCTTTAACT-3′
and antisense, 5′-CAACTGAT CGAAGGAACG-3′. The PCR amplification for
the quantification of the HSP90 and HIF-1α mRNA was performed using
a SYBR® Premix Ex Taq™ II (Perfect Real-Time) kit
(Takara Bio, Otsu, Japan) and an ABI PRISM 7300 Sequence Detection
system (Applied Biosystems, Foster City, CA, USA), as previously
described (19).
Western blot analysis
The following primary antibodies were used for
western blot analysis: HSP90 (ab13492; 1:1,000), HIF-1α (ab85886;
1:1,000) (both from Abcam), Akt (sc-5298; 1:1,000), CDK4 (sc-260;
1:1,000), ubiquitin (sc-8017; 1:500) and β-actin (sc-47778;
1:1,000) (all from Santa Cruz Biotechnology). Horseradish
peroxidase-conjugated goat anti-rabbit or anti-mouse secondary
antibodies (1721019 and 1708242; Bio-Rad, Hercules, CA, USA) were
used at a 1:1,000–1:5,000 dilution and the results were detected
using Western Blotting Luminol reagent (sc-2048; Santa Cruz
Biotechnology).
Immunofluorescence (IF) staining
The HCC cells were fixed with 3% paraformaldehyde
and then permeabilized with 0.2% Triton X-100, as previously
described (15). The fixed cells
were subsequently incubated with the HIF-1α (1:500) primary
antibody. An Alexa Fluor-conjugated IgG (Invitrogen, Carlsbad, CA,
USA) was used as the secondary antibody. A LSM 5 Pascal laser
scanning microscope (Zeiss, Oberkochen, Germany) was used to
capture fluorescence confocal images with a x40 lens and LSM 5
PASCAL software (version 4.2 SP1; Zeiss) was then used to scan the
images, as described in a previous study (15).
Co-immunoprecipitation (co-IP)
Flag (F1804; Sigma) and hemagglutinin (HA) (12CA5;
Roche, Indianapolis, IN, USA) antibodies were used in the co-IP
assays. Immunoprecipitation buffer was used to obtain total protein
lysate. The Bio-Rad DC™ protein assay reagent A/B/S (Bio-Rad) was
used to quantify the total protein concentration of the
supernatants. Total protein (500 µg) was mixed with 1
µg of the primary antibody, or IgG as previously described
(15), and the mixture was shaken
for 4 h at 4°C. The beads (Protein G Sepharose 4 Fast Flow; GE
Healthcare Life Sciences, Piscataway, NJ, USA) were then added to
the mixture and shaken at 4°C for 2 h. Subsequently, the beads were
collected by centrifugation (500 × g, 4°C) and washed 3 times using
immunoprecipitation buffer, as previously described (15). Sample loading buffer (5X) was
mixed with the beads and boiled for 10 min. The supernatant was
used for western blot analysis.
CHX chase assays were also performed to analyze the
HSP90-mediated downregulation of HIF-1α in the Hep3B cells in which
HSP90 was knocked down and in the controls. Cycloheximide (CHX;
ab120093), a protein synthesis inhibitor, was obtained from Abcam.
Furthermore, MG132 (a proteosome inhibitor; ab141003; Abcam) was
used to determine whether it can prevent the downregulation of
HIF-1α.
Cell viability and proliferation
assays
In the present study, 3-(4,
5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT;
Roche) assay was used to assess cell viability at 24, 48 and 72 h.
In addition, the HCC cells were seeded into 96-well plates at
3,000–5,000 cells/well for 24 h and 5-bromodeoxy-uridine (BrdU)
assay (chemiluminescent) (Roche) was used to assess cell
proliferation, as previously described (17).
Cell apoptosis
The level of apoptosis was analyzed using an Annexin
V-FLUOS staining kit (Roche), as previously described (20). Caspase-3/7 activity was analyzed
using an Apo-ONE® Homogeneous Caspase-3/7 assay
(Promega, Madison, WI, USA), as described in the study by Zheng
et al (21).
Flow cytometry
We analyzed cell apoptosis using the Annexin V-FLUOS
staining kit (obtained from Roche) after a 48-h transfection.
Briefy, the samples were analyzed using a BD FACSCanto II flow
cytometer (Becton-Dickinson, Franklin Lakes, NJ, USA). Three
independent repeated experiments were performed.
In vivo experiments
We used 4–6 week-old female BALB/c nude mice (n=18;
Centre of Laboratory Animals at Zhejiang Provincial People's
Hospital, Hangzhou, China) to establish a xenograft tumor model.
The mice (2 animals/cage) were housed in sterilized cages at a
constant humidity and temperature and the mice were fed a regular
autoclaved chow diet with water ad libitum, as previously
described (21). As described in
American Type Culture Collection (ATCC), HepG2 is not a tumorigenic
cell line; we thus inoculated 4–5×106 Hep3B cells
subcutaneously into the flank of each nude mouse. The tumor volume
was determined by measuring two dimensions and was calculated as
follows: tumor volume = length × width × widt h/2, as previousy
described (15). After 3 weeks,
we used a terminal deoxynucleotidyl transferase-mediated dUTP
nick-end labeling (TUNEL) assay kit (4810-30-K; R&D Systems,
Minneapolis, MN, USA) to detect the amount of apoptosis in the
tumor tissues according to the manufacturer's instructions.
Furthermore, all animal protocols were approved by the
Institutional Animal Care and Use Committee of Zhejiang Provincial
People's Hospital.
Statistical analysis
All data are presented as the means ± SEM. SPSS
software (SPSS, Inc., Chicago, IL, USA) was used for the
multivariate Cox regression analysis and the Pearson Chi-square
tests. GraphPad Prism 5 software (GraphPad Software, Inc., San
Diego, CA, USA) was used to evaluate statistical significance. A
value of P<0.05 was considered to indicate a statistically
significant difference.
Results
Clinical significance of increased HSP90
expression in HCC tissues
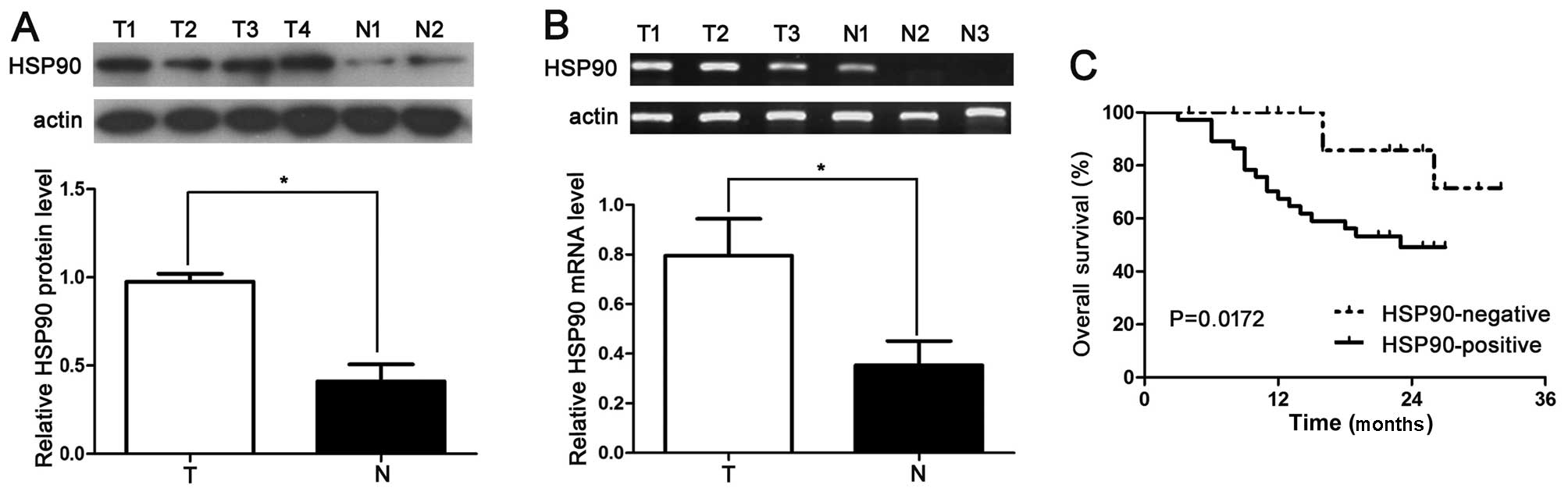
The protein expression of the HSP90 was determined
by the immunostaining of 60 pairs of cancerous and matched
para-carcinoma tissue samples, in order to investigate the clinical
significance of HSP90 in HCC. HSP90 immunoreactivity was considered
as either positive (scores 1–3) or negative (score 0). In these
tissues, HSP90 expression was detected in 20 (33.3%) of the HCC
specimens; however, only 8 (13. 3%) of the normal tumor-adjacent
tissues exhibited a positive HSP90 signal (P<0.05). In addition,
20 samples were subjected to RT-qPCR and western blot analysis for
HSP90 expression. We found that the HSP90 protein level in the HCC
tissues was significantly higher than that in the normal tissues
(P<0.05; Fig. 1A and B). The
results of the Pearson Chi-square test indicated that the increased
HSP90 expression in HCC tissues was significantly associated with
venous infiltration (P=0.004), an advanced tumor stage
[tumor-node-metastasis (TNM) stage III + IV; P=0.010] and a high
histological grade (Edmondson-Steiner grade III + IV; P=0.006)
(Table I).
Negative expression of HSP90 correlates
with an improved 3-year survival for patients with HCC
To confirm the role of HSP90 in evaluating the
prognosis of patients with HCC, the immunohistostaining of HSP90
was performed to determine the correlation between HSP90 expression
and the 3-year patient survival. We used the overall 3-year patient
survival data to analyze cases with negative and positive HSP90
staining by constructing Kaplan-Meier survival curves. Our data
indicated that the overall 3-year survival in the HSP90-negative
expression group was 63.75%. In comparison, the overall 3-year
survival in the HSP90-positive expression group was 45.46%. The
patients in the HSP90-positive expression group (n=20) had a
markedly poorer prognosis than those in the HSP90-negative
expression group (n=40; P=0.0172; Fig. 1C). These data suggest that in HCC,
HSP90 may act as a potential prognostic marker. Additionally, HSP90
expression is an independent factor for predicting the 3-year
overall survival in patients with HCC (P = 0. 0172; Table II).
| Table IIUnivariate and multivariate analysis
of factors associated with 3-year overall survival. |
Table II
Univariate and multivariate analysis
of factors associated with 3-year overall survival.
| Parameter | HR | P-value |
|---|
| Univariate
analysis |
| Tumor
size(cm) | 6.041 | 0.016a |
| Edmondson-Steiner
grading | ©.032 | 0.006a |
| TNM stage | 75.634 | 0.040a |
| HSP90 (high vs.
lower) | 22.298 | 0.0172a |
| Multivariate
analysis |
| Edmondson-Steiner
grading | 18.669 | 0.000a |
| TNM stage | 3.576 | 0.017a |
| HSP90 (high vs.
lower) | 4.230 | 0.040a |
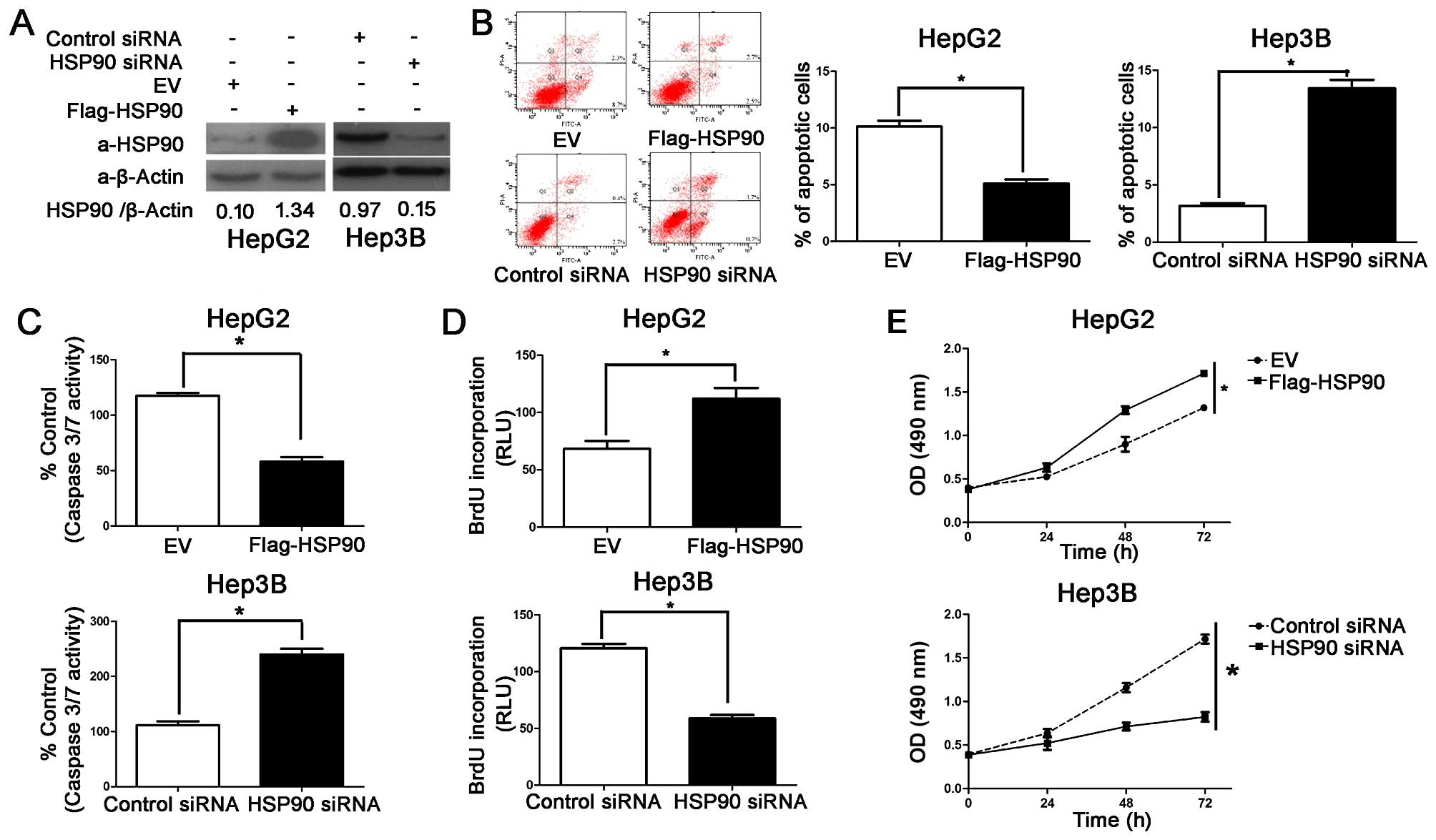
HSP90 induces the proliferation and
inhibits the apoptosis of HCC cells
In our previous study, we demonstrated that the
protein expression of HSP90 may be associated with the metastasis,
development and invasion of human CRC, and that its synergistic
effects may play a role in the development of CRC (9). In the present study, we aimed to
determine whether HSP90 may serve as a cancer-promoting gene in HCC
by promoting cell proliferation and inhibiting apoptosis. HSP90
protein expression was downregulatd by HSP90 siRNA or it was
increased by Flag-HSP90 (ectopically expressing a flag-tagged
HSP90), in the Hep3B and HepG2 cells, respectively (Fig. 2A). As determined by caspase-3/7
activity assays, apoptosis assays and flow cytometry, the
overexpression of HSP90 prevented the HepG2 cells from undergoing
apoptosis and the knockdown of HSP90 induced the apoptosis of the
Hep3B cells (P<0.05, respectively; Fig. 2B and C). MTT and BrdU assays were
also used to determine the effects of HSP90 on cancer cell
viability and proliferation, respectively. As expected, the
knockdown of HSP90 decreased the viability and proliferation of the
Hep3B cells and HSP90 overexpression enhanced the viability and
proliferation of the HepG2 cells (P<0.05; Fig. 2D and E). The HepG2 cells exhibited
a lower basal expression level of HSP90 than the Hep3B cells. Thus,
our data indicated that the Hep3B cells had more baseline apoptosis
and less proliferative ability than the HepG2 cells (as indicated
by our preliminary experiments; data not shown). Thus, HSP90 exerts
promotes the development of HCC effect by inhibiting the apoptosis
and promoting the growth of cancer cells.
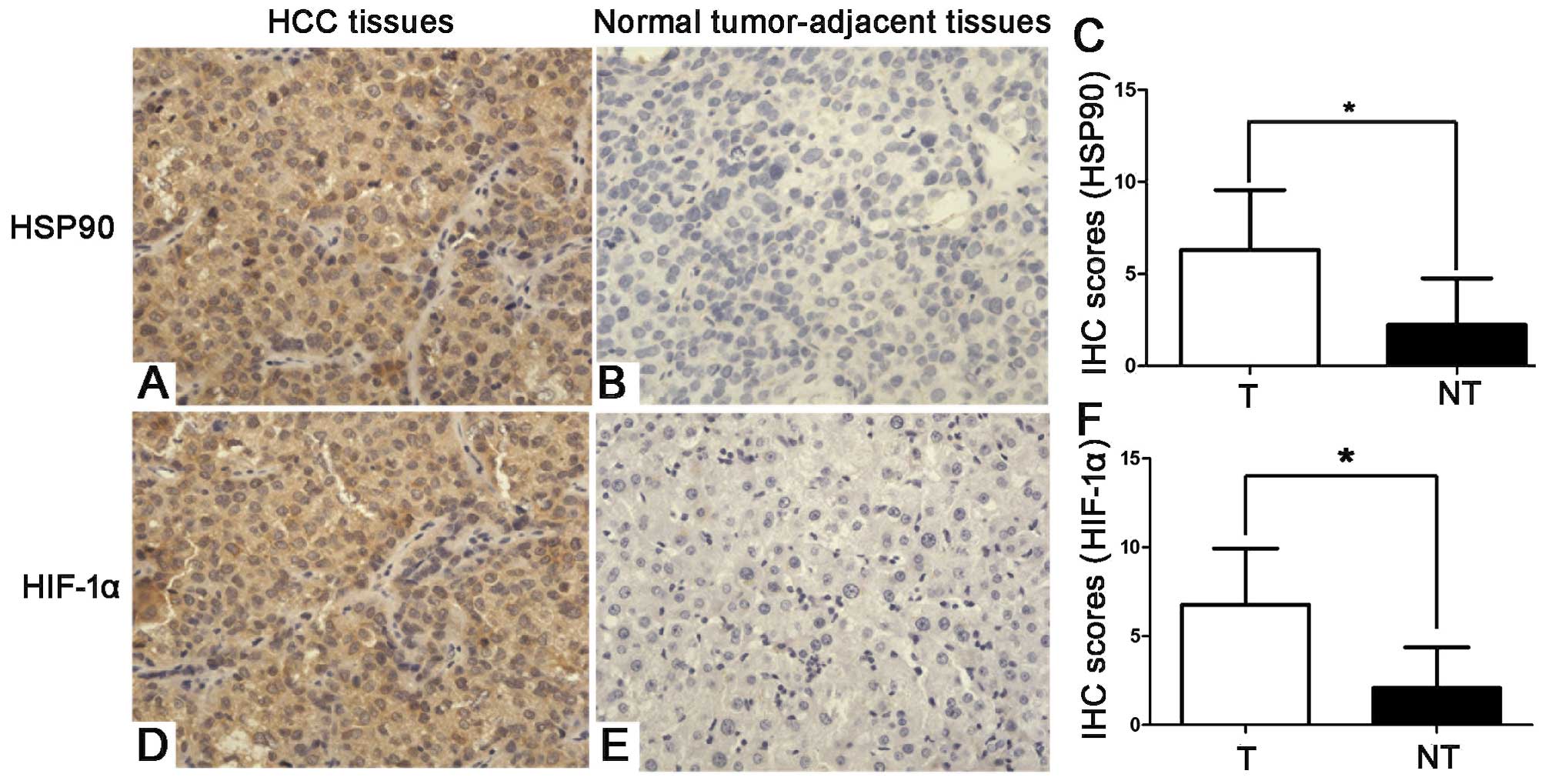
HSP90 positively correlates with HIF-1α
protein expression in HCC tissues
Since HI F-1α overexpression has been reported in
HCC (22), we examined the
correlation between HSP90 and HIF-1α in 60 HCC tissue samples using
IHC. HSP90 and HIF-1α immunoreactivity was considered as either
positive (scores 1–3) or negative (score 0). The protein expression
of HSP90 and HIF-1α in the cancer tissues was higher than that in
the paired para-carcinoma tissues (P<0.05). In addition, the IHC
scores that were used for the semi-quantitative analysis of HSP90
and HIF-1α expression revealed a strong positive correlation
between HSP90 and HIF-1α in the HCC tissues (r= 0.420; P<0.05;
Fig. 3).
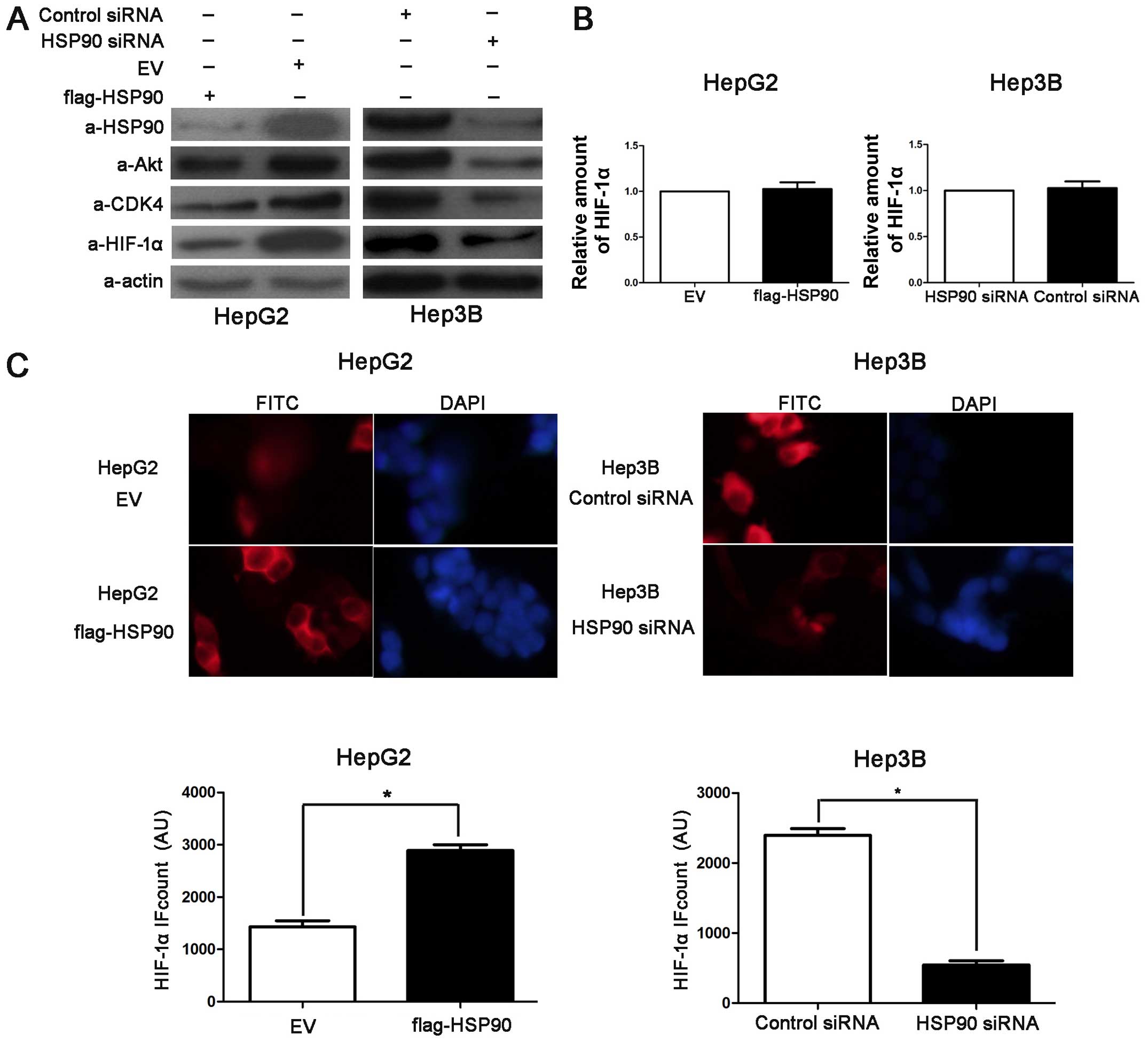
HSP90 regulates HIF-1α abundance in HCC
cells
To examine the downstream target genes of HSP90 in
HCC, the Hep3B and HepG2 cells were transfected with HSP90 siRNA
and Flag-HSP90, respectively. Western blot analysis was performed
to detect Akt, CDK4 and HIF-1α. Akt and CDK4 are confirmed target
proteins of HSP90 (23,24). HIF-1α acts as an activating
transcription factor involved in the regulation of apoptosis,
proliferation and cell growth (25). The overexpression of HSP90 led to
the accumulation of Akt and CDK4 in the HepG2 cells and
transfection with HSP90 siRNA decreased the levels of both proteins
in the Hep3B cells (Fig. 4A).
Furthermore, we found that the knockdown of HSP90 decreased the
protein level of HIF-1α in the Hep3B cells and that the
overexpression of HSP90 increased the HIF-1α protein levels in the
HepG2 cells (Fig. 4A), whereas
the HIF-1α mRNA levels were only either slightly increased or
slightly decreased (Fig. 4B). In
addition, IF staining for HIF-1α revealed that the average level of
HIF-1α in the Flag-HSP90-transfected HepG2 cells was significantly
higher than that in the control cells (P<0.05; Fig. 4C). However, the average level of
HIF-1α in the HSP90 siRNA-transfected Hep3B cells was significantly
lower than the HIF-1α levels in the control cells (P<0.05;
Fig. 4C). These data suggest that
HSP90 increases HIF-1α protein levels in HCC cells.
HSP90 inhibits the ubiquitination and
proteasomal degradation of HIF-1α
A previous study reported the identification of
receptor for activated protein C kinasex1 (RACK1) as a novel
HIF-1α-interacting protein (26).
It was demonstrated that RACK1 promotes the O2/prolyl
hydroxylase domain (PHD)/von Hippel-Lindau (VHL)-independent and
proteasome-dependent degradation of HIF-1α (26). RACK1 competes with HSP90 for
binding to the PAS-A domain of HIF-1α. RACK1 activity is required
for the mechanisms of action of the HSP90 inhibitor,
17-allylaminogeldanamycin, to induce HIF-1α degradation (26). In this study, to determine whether
HSP90 binds to HIF-1α and inhibits its ubiquitination and
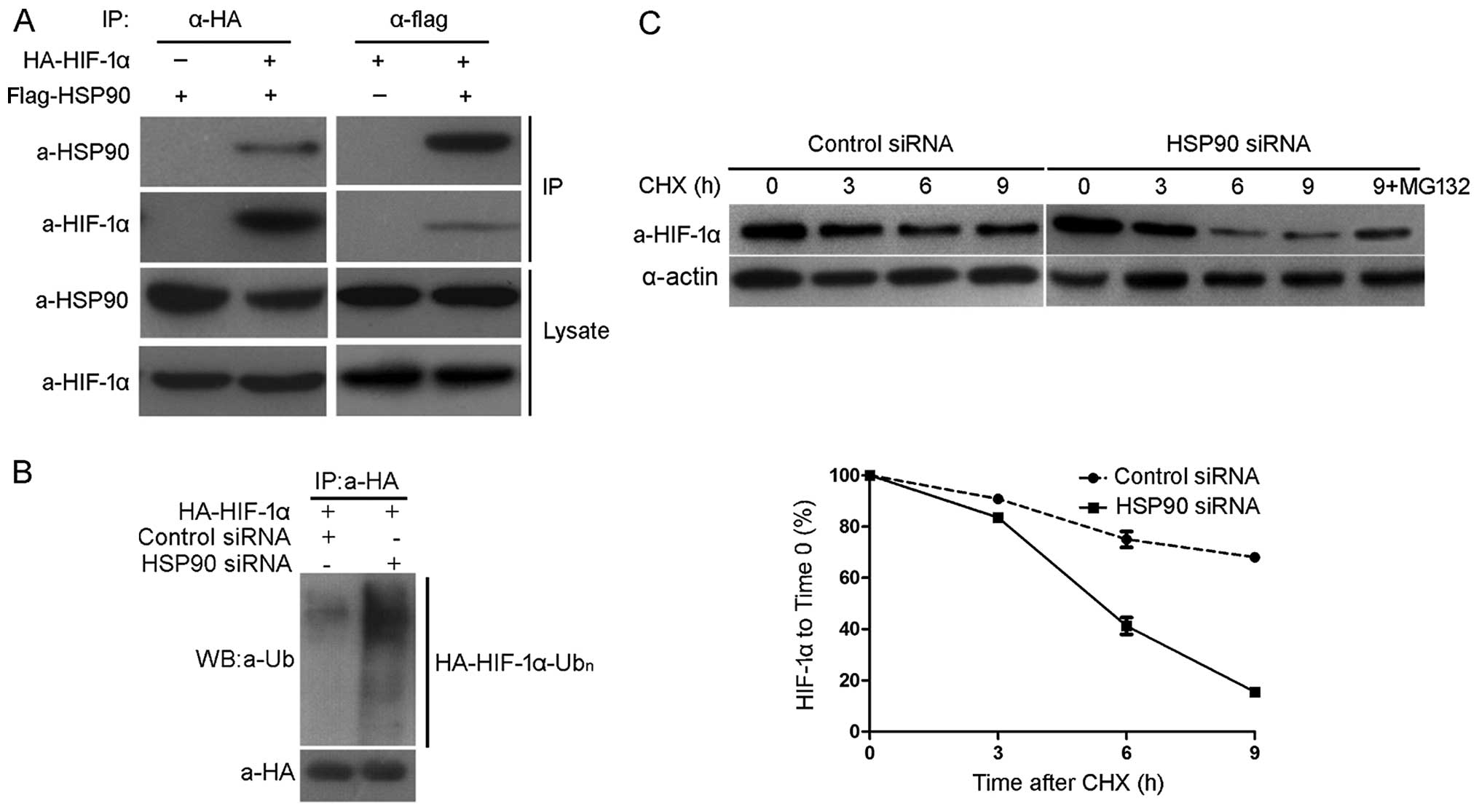
proteasomal degradation, we confirmed the interaction between HSP90
and HIF-1α in 293 cells using co-IP of HA-HIF-1α and Flag-HSP90
(Fig. 5A). In the next experiment
investigating the precipitation of H A- H I F-1α from HepG2 cells
expressing HA-HIF-1α, H I F-1α ubiquitination was detected by
western blot analysis using an ubiquitin antibody. As shown in
Fig. 5B, HSP90 knockdown
increased the ubiquitination of HIF-1α in the Hep3B cells. CHX is a
protein synthesis inhibitor and CHX chase assays were performed in
order to analyze the HSP90-mediated down-regulation of HIF-1α in
the Hep3B cells in which HSP90 was knocked down and the controls.
Compared with that observed in the control cells, the half-life of
HIF-1α was decreased in the cells in which HSP90 was knocked down
(4.9 vs. 12.4 h; Fig. 5C; 12.4 is
the average half-life of HIF-1α). Furthermore, MG132 (a proteasome
inhibitor) was able to prevent the downregulation of HIF-1α
(Fig. 5C). These data suggest
that HSP90 binds to H I F-1α and thereby inhibits the
ubiquitination and proteasomal degradation of HIF-1α.
HSP90 promotesthe proliferation and
inhibits the apoptosis of HCC cells by inhibiting the degradation
of HIF-1α
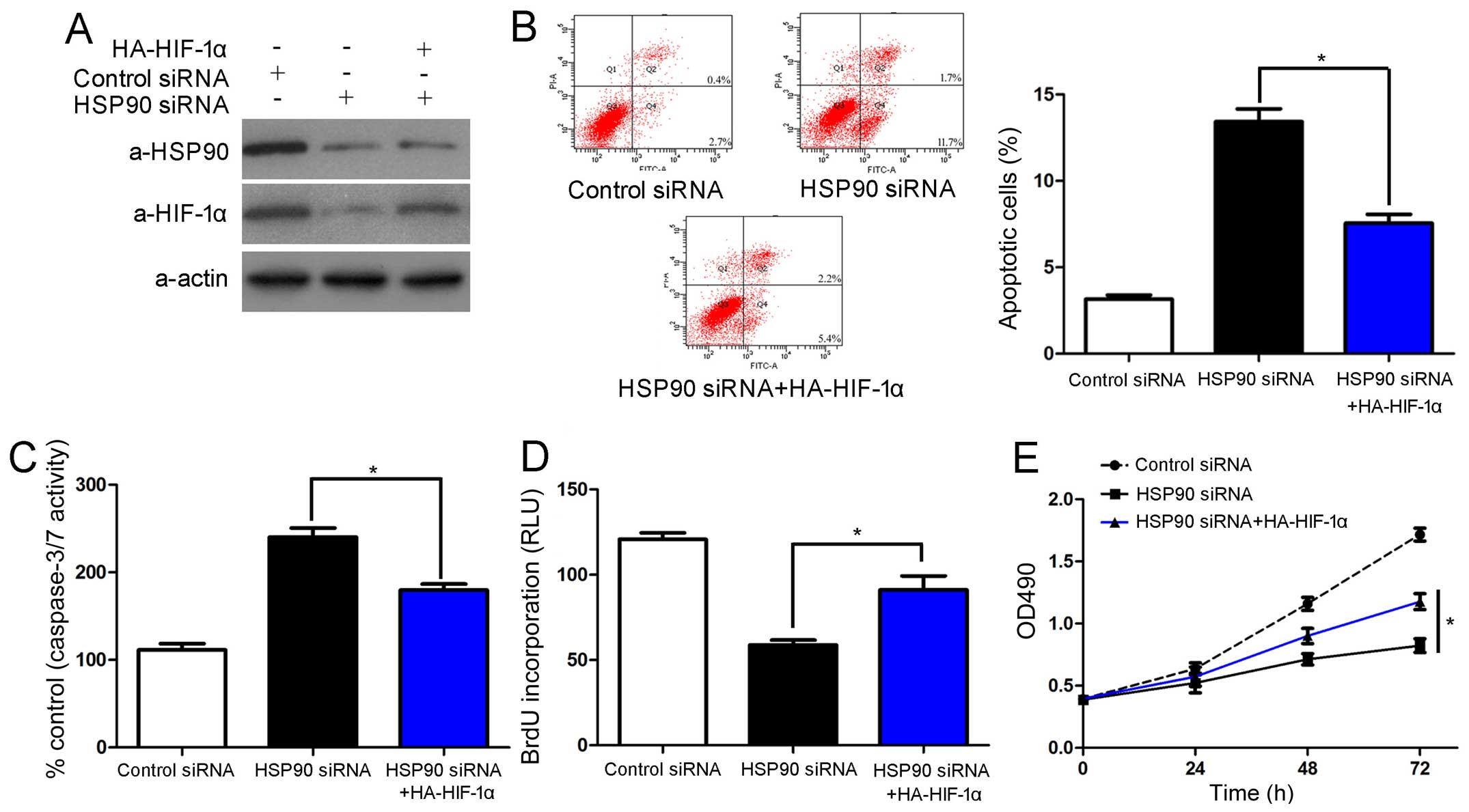
To determine whether HIF-1α protein is involved in
the HSP90 siRNA-induced growth arrest and apoptosis of HCC cells,
the Hep3B cells in which HSP90 was knocked down were transfected
with HA-HIF-1α. Unsurprisingly, the restoration of HIF-1α
expression in the Hep3B cells reversed the effects of HSP90
knockdown, which led to a significant decrease in the number of
apoptotic cells and increased cell viability and proliferation
(P<0.05, respectively; Fig.
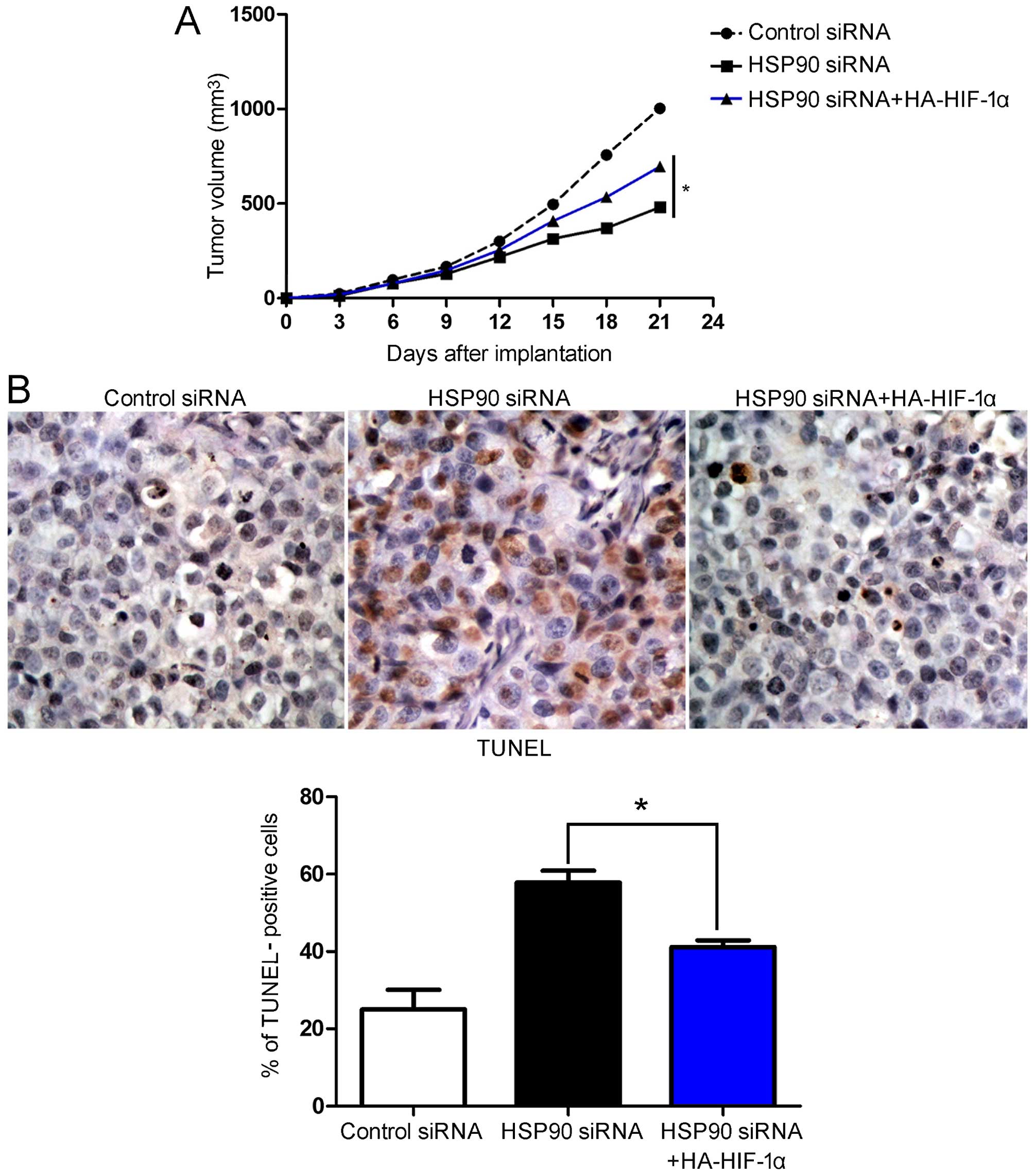
6). To determine whether HSP90 siRNA affected tumor growth by
inhibiting HIF-1α, we used a mouse tumor model of Hep3B
subcutaneous tumors. Firstly, the Hep3B cells were infected with
different retroviruses, and these Hep3B cells were then implanted
into nude mice by subcutaneous injection. After 21 days, tumor
growth curves suggested that HSP90 siRNA attenuated Hep3B tumor
growth in the mice. The restoration of HIF-1α expression in the
Hep3B cells in which HSP90 was knocked down partially restored
tumor growth (P<0.05; Fig.
7A). We performed IHC and TUNEL assays in the xenografted
tissues. As expected, HSP90 siRNA induced apoptosis in vivo.
The restoration of HIF-1α expression partially abolished the
inhibitory effects of HSP90 siRNA on HCC cell growth; it led to a
significant reduction in the number of apoptotic cells (P<0.05,
respectively; Fig. 7B). In
conclusion, these data indicate that HIF-1α may act as a downstream
factor in the HSP90-induced inhibition of apoptosis and the
promotion of cell growth in HCC.
Discussion
The occurrence, metastasis, development and invasion
of neoplasms result from interactions among multiple factors,
polygenes and multi-stages (9).
HSPs, also known as molecular chaperones, are essential for
regulating intracellular protein balance with a highly conserved
amino acid sequence (9). HSPs are
involved in the final activation of several regulatory proteins
(9). Based on the molecular
weight, HSPs have been classified into several groups, including
HSP70, HSP90 and HSP110 (27).
HSP90 is mainly comprised of HSP90β and HSP90α. Furthermore, it has
a molecular weight of 90 kDa, the largest part of which exists in
glucose-regulated protein 94 (GRP94) of the endoplasmic reticulum
and TNF receptor-associated protein (TRAP)1 of the mitochondria
(28). Under physiological
conditions, HSP90 accounts for 1–2% total proteins in cells;
however, in a state of stress, its content increases by
approximately 2–10-fold. The protein and mRNA expression and levels
of HSP90 have been shown to be upregulated in various types of
cancers (9,29–32). In this study, we first examined
the protein expression of HSP90 in samples from 60 patients with
HCC using IF staining and western blot analysis, and our data
demonstrated that HSP90 expression was significantly higher in the
HCC tissues compared with that in matched normal tissues. In
addition, HSP90 expression significantly correlated with venous
infiltration, TNM tumor staging and Edmondson-Steiner grading,
which is consistent with the results of our previous study
(9). Furthermore, our data
indicated that the negative expression of HSP90 significantly
correlated with an improved 3-year patient survival for HCC
patients, which is consistent with the results of previous studies
on breast cancer, gastrointesinal stromal tumors and mesenchymal
tumors (33,34). Importantly, multivariate Cox
repression analysis indicated that HSP90 is an independent factor
for predicting overall 3-year survival in patients with HCC. These
data suggested that HSP90 is crucial for the prognosis of patients
with HCC. A previous study demonstrated that HSP90 acts as a tumor
promoter which may be involved in the proliferation and apoptosis
of HCC cells (35). The present
study, further confirmed that HSP90 promotes tumor progression by
accelerating tumor growth and inhibiting the apoptosis of HCC
cells.
The prey or target proteins of HSP90 include
urokinase and matrix metalloproteinase-2 (MMP-2) which are involved
in the invasion and metastasis of tumor cells (9). It has been demonstrated that HIF is
capable of maintaining revascularization (28). Furthermore, to inhibit the
apoptosis of tumor cells, HIF-1α regulates the function of tumor
necrosis factor receptor (TNFR), protein kinase B (Akt) and nuclear
factor-κB (NF-κB) (28).
Importantly, in our previous study, we reported that the HSP90
levels positively correlated with HIF-1α in HCC tissues (9).
In the present study, we confirmed that HSP90 and
HIF-1α expression in the HCC tissues were higher compared with the
expression levels in normal tumor-adjacent tissues. Furthermore,
HSP90 was positively associated with HIF-1α protein expression in
the HCC tissues. In vitro experiments demonstrated that
HSP90 positively regulated Akt and CDK4 abundance in the HCC cells.
Notably, HSP90 overexpression induced a significant accumulation of
HIF-1α in the HepG2 cells. Conversely, the knockdown of HSP90 by
siRNA in the Hep3B cells decreased the expression of the target
genes including Akt, CDK4 and HIF-1α. However, the HIF-1α mRNA
levels were not altered with HSP90 regulation, which suggested that
HSP90 only regulated the level of the HIF-1α protein in the HCC
cells. The present study also revealed the positive interaction
between HSP90 and HIF-1α in 293 cells using co-IP. Furthermore,
western blot analysis and co-IP indicated that HSP90 siRNA promoted
HIF-1α ubiquitination and shortened the half-life of HIF-1α in the
Hep3B cells, and that treatment with MG132 (a proteasome inhibitor)
inhibited the downregulation of HIF-1α by HSP90 siRNA. The data we
present herein suggest that HSP90 inhibits the ubiquitination of
HIF-1α and its subsequent proteasomal degradation. However, further
studies are required to confirm whether HIF-1α is a target of
HSP90, and we aim to address this issue using proteomic data with
searches for HIF-1α with HSP90 in the future.
In the present study, we demosntrated that the
knockdown of HSP90 using siRNA led to the downregulation of HIF-1α
in vivo and in vitro. Furthermore, the observed
increase in growth arrest and apoptosis which the knockdown of
HSP90 resulted in, was partially reversed by the overexpression of
HIF-1α. However, only less than half of the inhibitory effects of
HSP90 siRNA on HCC cells were abolished, in spite of achieving more
than half the restoration in the levels of HIF-1α. These data
suggest that HIF-1α is not the only downstream gene of HSP90 which
inhibites apoptosis and promotes growth in HCC. As HSP90 targets
many oncoproteins and subsequently regulates cell proliferation,
the cell cycle and apoptosis, we suggest that HSP90 inhibits
apoptosis and promotes growth by regulating HIF-1α. HSP90
suppresses the degradation of HIF-1α and inhibits apoptosis and
promotes growth in HCC.
In conclusion, in the present study, we have proven
that increased levels of HSP90 are associated with poor
clinicopathological characteristics in tissue samples of patients
with HCC. In patients with HCC, the positive expression of HSP90 is
an independent factor for predicting poor prognosis. HSP90 promotes
growth and prevents apoptosis by inhibiting the ubiquitination and
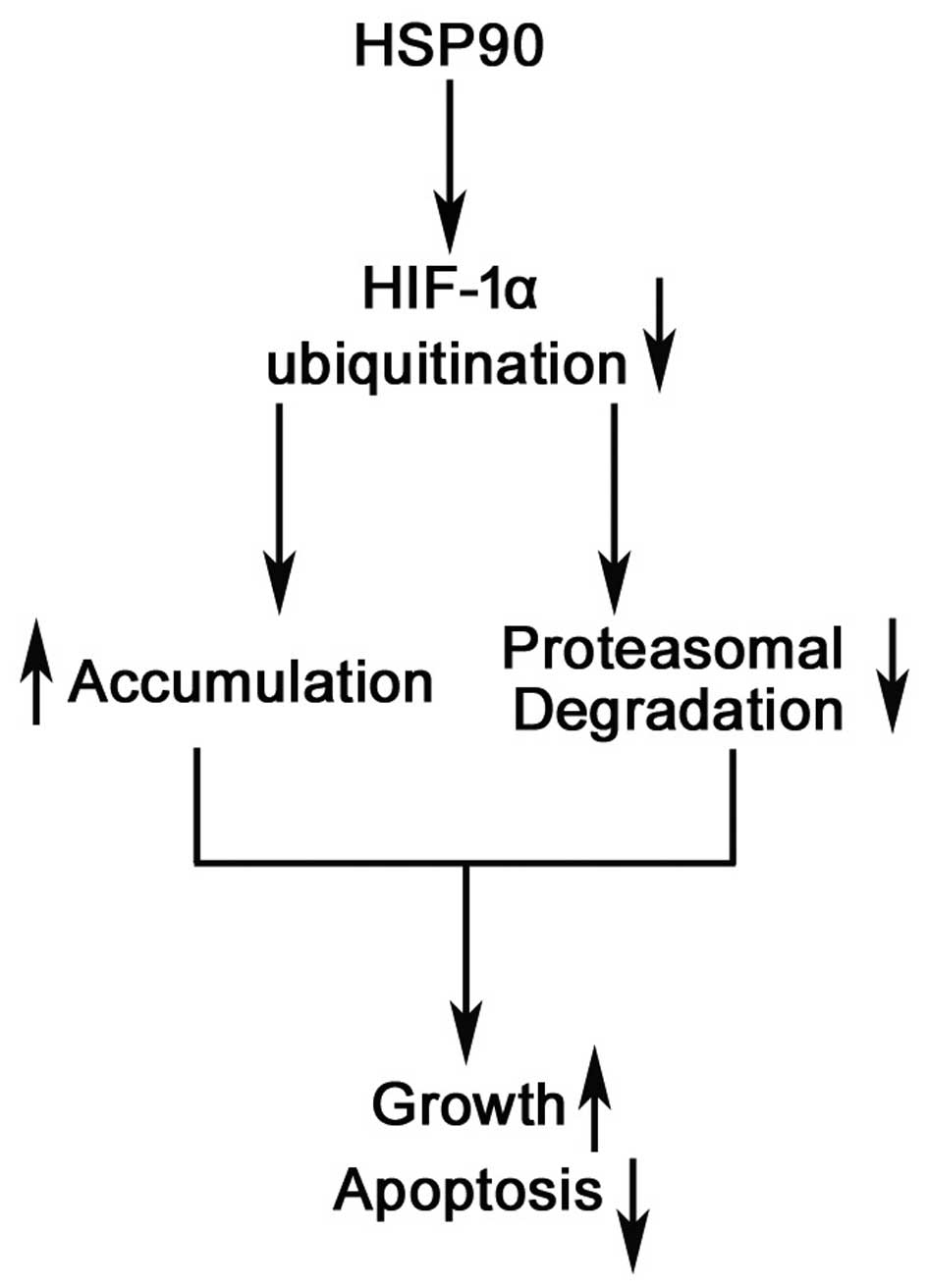
proteasomal degradation of the HIF-1α protein (Fig. 8). Thus, we suggest that a gain of
HSP90 function leads to hepatocarcinogenesis, partly through the
accumulation of HIF-1α. We have identified HSP90 as a potential
therapeutic target in HCC.
In conclusion, the findings of the present study
demonstrated that the expression of HSP90 and HIF-1α was markedly
higher in the cancer tissues compared with the expression levels in
the non-cancerous tissues, and that increased HSP90 levels
correlated with poor clinicopathological characteristics in HCC.
Furthermore, we have proven that HSP90 is an independent factor for
predicting the overall 3-year survival in patients with HCC. In
vitro experiments found that HSP90 promoted HCC cell growth by
inhibiting apoptosis and promoting growth. HIF-1α expression is
positively associated with HSP90. Furthermore, our data indicate
that HSP90 regulates HIF-1α protein abundance by inhibiting the
ubiquination and proteasomal degradation of HIF-1α in HCC cells.
Notably, the restoration of HIF-1α partially abolished the
inhibitory effects of HSP90 siRNA on HCC cell growth, which
suggests that HSP90 may promote tumor growth by regulating the
abundance of HIF-1α protein. The data of our study reveal that
HSP90 may have the potential to serve as a clinical biomarker and
as a target for gene therapy in HCC.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant no. 81502092), the
Zhejiang Provincial Natural Science Foundation of China (grant nos.
LY16H160043 and LY13H150009) and the general project funds from the
Health Department of Zhejiang province (grant nos. 2016KYA022 and
2015KYB033).
References
|
1
|
Buonaguro L, Petrizzo A, Tagliamonte M,
Tornesello ML and Buonaguro FM: Challenges in cancer vaccine
development for hepatocellular carcinoma. J Hepatol. 59:897–903.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Xu Q, Liu X, Zheng X, Yao Y, Wang M and
Liu Q: The transcriptional activity of Gli1 is negatively regulated
by AMPK through Hedgehog partial agonism in hepatocellular
carcinoma. Int J Mol Med. 34:733–741. 2014.PubMed/NCBI
|
|
3
|
Kiang JG and Tsokos GC: Heat shock protein
70 kDa: molecular biology, biochemistry, and physiology. Pharmacol
Ther. 80:183–201. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Welch WJ: Mammalian stress response: cell
physiology, structure/function of stress proteins, and implications
for medicine and disease. Physiol Rev. 72:1063–1081.
1992.PubMed/NCBI
|
|
5
|
Milicevic Z, Bogojevic D, Mihailovic M,
Petrovic M and Krivokapic Z: Molecular characterization of hsp90
isoforms in colorectal cancer cells and its association with tumour
progression. Int J Oncol. 32:1169–1178. 2008.PubMed/NCBI
|
|
6
|
Nimmanapalli R, O'Bryan E and Bhalla K:
Geldanamycin and its analogue
17-allylamino-17-demethoxygeldanamycin lowers Bcr-Abl levels and
induces apoptosis and differentiation of Bcr-Abl-positive human
leukemic blasts. Cancer Res. 61:1799–1804. 2001.PubMed/NCBI
|
|
7
|
Pascale RM, Simile MM, Calvisi DF, Frau M,
Muroni MR, Seddaiu MA, Daino L, Muntoni MD, De Miglio MR,
Thorgeirsson SS and Feo F: Role of HSP90, CDC37, and CRM1 as
modulators of P16(INK4A) activity in rat liver carcinogenesis and
human liver cancer. Hepatology. 42:1310–1319. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yano M, Naito Z, Tanaka S and Asano G:
Expression and roles of heat shock proteins in human breast cancer.
Jpn J Cancer Res. 87:908–915. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xu QR, Liu X, Yao YM and Liu QG:
Expression of HSP90 and HIF-1α in human colorectal cancer tissue
and its significance. Asian Pac J Trop Med. 7:720–724. 2014.
View Article : Google Scholar
|
|
10
|
Palsson-McDermott EM, Curtis AM, Goel G,
Lauterbach MA, Sheedy FJ, Gleeson LE, van den Bosch MW, Quinn SR,
Domingo-Fernandez R, Johnston DG, et al: Pyruvate kinase M2
regulates Hif-1α activity and IL-1β induction and is a critical
determinant of the Warburg effect in LPS-activated macrophages.
Cell Metabolism. 21:65–80. 2015. View Article : Google Scholar
|
|
11
|
Wang GL, Jiang B-H, Rue EA and Semenza GL:
Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS
heterodimer regulated by cellular O2 tension. Proc Natl
Acad Sci USA. 92:5510–5514. 1995. View Article : Google Scholar
|
|
12
|
Krishnamachary B, Berg-Dixon S, Kelly B,
Agani F, Feldser D, Ferreira G, Iyer N, LaRusch J, Pak B, Taghavi P
and Semenza GL: Regulation of colon carcinoma cell invasion by
hypoxia-inducible factor 1. Cancer Res. 63:1138–1143.
2003.PubMed/NCBI
|
|
13
|
Mizukami Y, Li J, Zhang X, Zimmer MA,
Iliopoulos O and Chung DC: Hypoxia-inducible factor-1-independent
regulation of vascular endothelial growth factor by hypoxia in
colon cancer. Cancer Res. 64:1765–1772. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cao D, Hou M, Guan YS, Jiang M, Yang Y and
Gou HF: Expression of HIF-1alpha and VEGF in colorectal cancer:
association with clinical outcomes and prognostic implications. BMC
Cancer. 9:4322009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tu K, Yang W, Li C, Zheng X, Lu Z, Guo C,
Yao Y and Liu Q: Fbxw7 is an independent prognostic marker and
induces apoptosis and growth arrest by regulating YAP abundance in
hepatocellular carcinoma. Mol Cancer. 13:110. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Riis ML, Lüders T, Nesbakken AJ, Vollan
HS, Kristensen V and Bukholm IR: Expression of BMI-1 and Mel-18 in
breast tissue-a diagnostic marker in patients with breast cancer.
BMC Cancer. 10:6862010. View Article : Google Scholar
|
|
17
|
Tu K, Zheng X, Zan X, Han S, Yao Y and Liu
Q: Evaluation of Fbxw7 expression and its correlation with the
expression of c-Myc, cyclin E and p53 in human hepatocellular
carcinoma. Hepatol Res. 42:904–910. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xu MZ, Yao TJ, Lee NP, Ng IO, Chan YT,
Zender L, Lowe SW, Poon RT and Luk JM: Yes-associated protein is an
independent prognostic marker in hepatocellular carcinoma. Cancer.
115:4576–4585. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xu Q, Liu X, Zheng X, Yao Y and Liu Q:
PKM2 regulates Gli1 expression in hepatocellular carcinoma. Oncol
Lett. 8:1973–1979. 2014.PubMed/NCBI
|
|
20
|
Tu K, Zheng X, Zhou Z, Li C, Zhang J, Gao
J, Yao Y and Liu Q: Recombinant human adenovirus-p53 injection
induced apoptosis in hepatocellular carcinoma cell lines mediated
by p53-Fbxw7 pathway, which controls c-Myc and cyclin E. PLoS One.
8:e685742013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zheng X, Vittar NB, Gai X,
Fernandez-Barrena MG, Moser CD, Hu C, Almada LL, McCleary-Wheeler
AL, Elsawa SF, Vrabel AM and Shire AM: The transcription factor
GLI1 mediates TGFβ1 driven EMT in hepatocellular carcinoma via a
SNAI1-dependent mechanism. PLoS One. 7:e495812012. View Article : Google Scholar
|
|
22
|
Cao S, Yang S, Wu C, Wang Y, Jiang J and
Lu Z: Protein expression of hypoxia-inducible factor-1 alpha and
hepatocellular carcinoma: a systematic review with meta-analysis.
Clin Res Hepatol Gastroenterol. 38:598–603. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xue N, Jin J, Liu D, Yan R, Zhang S, Yu X
and Chen X: Antiproliferative effect of HSP90 inhibitor Y306zh
against pancreatic cancer is mediated by interruption of AKT and
MAPK signaling pathways. Curr Cancer Drug Targets. 14:671–683.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Haarberg HE, Paraiso KH, Wood E, Rebecca
VW, Sondak VK, Koomen JM and Smalley KS: Inhibition of Wee1, AKT,
and CDK4 underlies the efficacy of the HSP90 inhibitor XL888 in an
in vivo model of NRAS-mutant melanoma. Mol Cancer Ther. 12:901–912.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lu XG, Xing CG, Feng YZ, Chen J and Deng
C: Clinical significance of immunohistochemical expression of
hypoxia-inducible factor-1alpha as a prognostic marker in rectal
adenocarcinoma. Clin Colorectal Cancer. 5:350–353. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu YV and Semenza GL: RACK1 vs. HSP90:
competition for HIF-1 alpha degradation vs. stabilization. Cell
Cycle. 6:656–659. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bagatell R and Whitesell L: Altered Hsp90
function in cancer: a unique therapeutic opportunity. Mol Cancer
Ther. 3:1021–1030. 2004.PubMed/NCBI
|
|
28
|
Goetz MP, Toft DO, Ames MM and Erlichman
C: The Hsp90 chaperone complex as a novel target for cancer
therapy. Ann Oncol. 14:1169–1176. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Whitesell L and Lindquist SL: HSP90 and
the chaperoning of cancer. Nat Rev Cancer. 5:761–772. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Workman P, Burrows F, Neckers L and Rosen
N: Drugging the cancer chaperone HSP90: Combinatorial therapeutic
exploitation of oncogene addiction and tumor stress. Ann NY Acad
Sci. 1113:202–216. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Qi XL, Bo AH, Yue SC, Wang CY and Liu HB:
Heat shock protein 90 mrna expressed in adenocarcinoma and adenoma
of significance. Proceed Military Academy Medical Sci. 30:351–353.
2011.
|
|
32
|
Bo AH, Dai J, Li SG, Zuo DS, Chen XL, Yan
YJ, et al: HSP90 mRNA expressed in stomach and large intestine
cancer research. Chin J Histochem Cytochem. 14:213–216. 2011.
|
|
33
|
Pick E, Kluger Y, Giltnane JM, Moeder C,
Camp RL, Rimm DL and Kluger HM: High HSP90 expression is associated
with decreased survival in breast cancer. Cancer Res. 67:2932–2937.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kang GH, Lee EJ, Jang KT, Kim KM, Park CK,
Lee CS, Kang DY, Lee SH, Sohn TS and Kim S: Expression of HSP90 in
gastrointestinal stromal tumours and mesenchymal tumours.
Histopathology. 56:694–701. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhao S, Li H, Jiang C, Ma T, Wu C, Huo Q
and Liu H: 17-Demethoxy-reblastatin, an Hsp90 inhibitor, induces
mitochondria-mediated apoptosis through downregulation of Mcl-1 in
human hepatocel-lular carcinoma cells. J Bioenerg Biomembr.
47:373–381. 2015. View Article : Google Scholar : PubMed/NCBI
|