Introduction
It has been recognized that under physiological and
pathological conditions, cells may release vesicles that may be
instrumental in intercellular communication as they carry
biologically active molecules (1–4).
The non-apoptotic vesicles have been classified based on their
biogenesis into exosomes derived from membranes of the endosomal
compartment and microvesicles that are formed from the budding of a
portion of the cell plasma membrane (5,6).
Given the heterogeneity of the released vesicles and the
overlapping features of molecules carried by exosomes and
microvesicles, it has been suggested that they be collectively
termed extracellular vesicles (EVs) (7). During vesiculation, nucleic acids,
including mRNAs, long non-coding RNAs (lncRNAs) and microRNAs
(miRNAs or miRs) are encapsulated within EVs and when released,
they are protected from degrading enzymes (8–10).
Therefore, EVs may carry genetic information that may be shared
among cells (11–14). Aliotta et al (15) demonstrated that lung-derived
vesicles carried RNA to marrow cells and altered the phenotype of
these cells both genetically and functionally. The horizontal
transfer of extracellular RNAs carried by EVs has been shown to be
able to reprogram hematopoietic progenitors (11) and to activate endothelial cells
(12). The EV-mediated transfer
of miRNAs is a particularly good candidate for the epigenetic
alterations observed in recipient cells. In fact, it has been shown
that EVs are enriched in miRNAs and that they can be transferred to
target cells (13,14). Gibbings et al (16) suggested that the constituents of
the miRNA effector complex, such as GW182 and Argonaute 2 (Ago2),
are involved in the packaging of miRNAs within multivesicular
bodies during exosome biogenesis. We previously found that EVs
derived from adult stem cells contain several ribonucloproteins
(14) known to be implicated in
RNA storage and/or stability, such as Staufen 1 and 2 (Stau1/2),
T-cell internal antigen-1 (TIA-1), TIA-1-related protein (TIAR),
the AU-rich element binding protein, human antigent R (HuR), and
Ago2 (17,18), which is involved in the transport
and processing of miRNAs (19).
Vesicles derived from adult human liver stem-like cells (HLSCs)
have also been shown to contain ribonucleoproteins and different
RNA species involved in hepatic regeneration (20). The efficacy of HLSCs in restoring
hepatic mass and liver functionality has been shown (20,21). HLSCs do not express hematopoietic
stem cell markers, whereas they express several mesenchymal markers
and display multiple differentiating abilities (22). Moreover, HLSCs express stem cells
and embryonic markers, including alpha-fetoprotein (AFP),
octamer-binding transcription factor 3/4 (Oct 3/4), Sox2, Musashi1,
nestin, nanog and pax2 (20,22). The expression of albumin and AFP
suggests a partial hepatic commitment (20,22).
Moreover, we found that HLSC-derived EVs were
enriched and transferred to tumor cells through antitumor miRNAs
that induced tumor regression and inhibited vascularization both
in vitro and in vivo (23). However, little is known about the
mechanisms of miRNA enrichment in HLSC-derived vesicles. Several
studies have indicated that Alix, an accessory protein of the
endosomal sorting complex required for transport (ESCRT), is
involved in the biogenesis of EVs (24–27) and has multifunctional activities
due to its domains, which provide multiple protein-binding sites
(24,28–33).
The aim of the present study was to evaluate whether
Alix is involved in the packaging of miRNAs within EVs released by
HLSCs. For this purpose, co-immunoprecipitation (Co-IP) experiments
of Alix with Ago2 were performed and the presence of selected miRNA
immuno precipitates was investigated. Furthermore, Alix was knocked
down in HLSCs in order to investigate its role in miRNA enrichment
within released EVs.
Materials and methods
Cell cultures
The HLSCs were obtained from normal human
cryopreserved hepatocytes (Lonza, Basel, Switzerland) and their
culture and characterization have been previously described
(20,22). Briefly, the HLSCs were cultured in
medium composed of 3 to 1 proportion of α-minimum essential medium
(α-MEM) and endothelial cell basal medium (EBM)-1 (both from Lonza,
Verviers, Belgium), supplemented with 10% EV-free fetal bovine
serum (FBS; Sigma-Aldrich, St. Louis, MO, USA), 2 mM L-glutamine
and 100 IU/ml penicillin and 100 µg/ml streptomycin. The
HLSCs were used at approximately 80% confluence and at passages 3
to 7. The HLSCs were characterized for the expression of the
mesenchymal stem cell (MSC) markers, CD29, CD44, CD73 and CD90,
several embryonic markers, such as Nanog, Nestin, Oct 3/4, Sox2,
Musashi1, AFP and Pax2, and the hepatic marker, albumin, as
previously described (20,22).
Human umbilical cord vein-derived endothelial cells (HUVECs) were
cultured in EBM supplemented with the EGM kit (Lonza) (12).
Isolation and characterization of
HLSC-derived EVs
EVs were obtained from supernatants of the HLSCs
cultured in RPMI serum-free medium supplemented with 100
µg/ml penicillin and 100 µg/ml streptomycin for 16 h.
Following centrifugation at 3k g for 20 min to remove debris,
cell-free supernatants were centrifuged at 10k g for 1 h at 4°C to
remove large vesicles, and the supernatant recovered was submitted
to a second ultracentrifugation at 100k g for 3 h at 4°C (Beckman
Coulter Optima L-90K ultracentrifuge; Beckman Coulter, Fullerton,
CA, USA). The protein content was quantified by the Bradford method
(Bio-Rad, Hercules, CA, USA) and the EV profile was analyzed and
counted by NanoSight (NanoSight Ltd., Minton Park, UK) equipped
with nanoparticle tracking analysis (NTA) 2.3 analytic software
(34).
Flow cytometric analyses of typical HLSC markers
were performed on the EVs as previously described (35). In selected experiments, the EVs
were labeled with PKH26 dye (Sigma-Aldrich), a red fluorescent
aliphatic chromophore (12,23), and with green fluorescent
SYTO® RNASelect (Molecular Probes, Inc., Eugene, OR,
USA) (23). After labeling, the
EVs were washed by ultracentrifugation at 100k g for 1 h at 4°C and
re-suspended in RPMI serum-free medium. To prove that the labeled
EVs were membrane vesicles and not aggregated proteins, the
vesicles were treated with 10% Triton X-100 (Bio-Rad).
As observed under a confocal microscope (Zeiss LSM 5
Pascal; serial no. 1206000125; Carl Zeiss, Oberkochen, Germany),
the EVs were stained with PKH26, which labeled membrane
phospholipids, indicating that particles detected in the 100 kg
ultracentrifugation fractions were not protein aggregates. This was
confirmed by treatment with Triton X that was capable of
solubilizing the vesicles, but not protein aggregates. Moreover,
the PKH26-labeled EVs co-stained with SYTO-RNA dye indicated the
presence of RNA within vesicles as previously described (14).
To evaluate EV internalization in the HUVECs, the
PKH26-labeled EVs were incubated at 37°C for different periods of
time and after cell washing the internalization was evaluated by
confocal microscopy and fluorescence-activated cell sorting (FACS)
analysis, as previously described (23).
Transmission electron microscopy was performed on
the EVs derived from the HLSCs suspended in phosphate buffered
saline (PBS) and loaded onto 200 mesh nickel Formvar carbon-coated
grids (Electron Microscopy Science, Hatfield, PA, USA) for 20 min.
Following fixation with 2.5% glutaraldehyde containing 2% sucrose
and repeated washings, the samples were negatively stained with
NanoVan (Nanoprobes, Yaphank, NY, USA) and observed using a
JEM-1010 electron microscope (Jeol, Tokyo, Japan).
Immunogold electron microscopy was performed as
previously described (14).
Briefly, the EVs derived from the HLSCs suspended in PBS were
loaded onto 200 mesh nickel Formvar carbon-coated grids. After 15
min of fixation in 3% paraformaldheyde (PAF) in PBS containing 2%
sucrose, the EVs were permeabilized with cold methanol (−20°C) for
10 min, extensively washed and blocked with 1% bovine serum albumin
(BSA) in PBS (pH 7.4). The EVs were then immunolabeled with Ago2
primary antibody (Cat. no. ab57113; Abcam, Cambridge Science Park,
Cambridge, UK) for 1 h at room temperature. As a control, the
primary antibody was omitted. After washing with 1% BSA in PBS (pH
8.2), the EVs were then incubated with EM goat anti-mouse IgG (H +
L) 5 nm Immunogold conjugate antibody (Cat. no. EM.GMHL5; British
Biocell International, Cardiff, UK) for 1 h in PBS/BSA (pH 8.2) at
room temperature. The EVs were post-fixed in 2.5% gluteraldehyde,
washed and embedded in a mixture of uranyl acetate (0.8%) and
methyl cellulose (0.13%) following silver enhancement (British
Biocell International). The samples were observed under a Jeol
JEM-1010 electron microscope.
Western blot analysis
The lysis of cells or EVs was performed in lysis
buffer [50 mmol/l Tris-HCl (pH 8.3), 1% Triton X-100, 10 mol/l
phenylmethylsulfonyl fluoride (PMSF), 10 mol/l leupeptin and 100
U/ml aprotinin] for 1 h at 4°C. Following centrifugation at 14,000
× g, the protein content in the supernatant was measured using the
Bradford assay. Protein lysates (50 µg) were subjected,
under reducing conditions, to 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and transferred onto
filters of nitrocellulose membranes. The membranes were incubated
with 5% non-fat milk in 20 mmol/l Tris-HCl (pH 7.5), 500 mmol/l
NaCl plus 0.1% Tween (TBS-T) and immunoblotted at 4°C for 12 h with
the appropriate concentration of relevant primary antibodies:
hepatocyte growth factor-regulated tyrosine kinase substrate (Hrs;
mouse monoclonal IgG; Cat. no. sc-101280), tumor susceptibility
gene (Tsg)101 (mouse monoclonal IgG; Cat. no. sc-101254), charged
multivesicular body protein (CHMP)4a (goat polyclonal IgG; Cat. no.
sc-49913), CHMP4b (rabbit polyclonal IgG; Cat. no. sc-82556),
CHMP4c (goat polyclonal IgG; Cat. no. sc-82560), Alix (mouse
monoclonal IgG; Cat. no. sc-271975), TIA-1 (goat polyclonal IgG;
Cat. no. sc-1751), TIAR (goat polyclonal IgG; Cat. no. sc-1749),
HuR (mouse monoclonal IgG; Cat. no. sc-5261), Stau1 (mouse
monoclonal IgG; Cat. no. sc-376123), Stau2 (mouse monoclonal IgG;
Cat. no. sc-101144), STAM (mouse monoclonal IgG; Cat. no.
sc-133092) (Santa Cruz Biotechnology, Santa Cruz, CA, USA),
anti-ribosomal protein S29 (RPS29; rabbit polyclonal IgG; Cat. no.
ab56224; Abcam) Ago2 (mouse monoclonal IgG; Cat. no. ab57113;
Abcam), CD63 and actin (Santa Cruz Biotechnology) for
normalization. The membranes were extensively washed in TBS-T and
incubated with peroxidase-conjugated isotype-specific secondary
antibodies [goat anti-rabbit IgG (Cat. no. 31462); goat anti-mouse
IgG (Cat. no. 31432); mouse anti-goat IgG (Cat. no. 31400); Thermo
Scientific Pierce, Rockford, IL, USA] for 1 h at room temperature.
After washing with TBS-T, the reaction was revealed using ECL
detection reagents (Clarity Western ECL Substrate, Bio-Rad) in a
ChemiDoc™ XRS+ system (Bio-Rad).
Co-IP assays on the cells and the EVs were performed
with Protein A/G PLUS-Agarose beads (Santa Cruz Biotechnology).
Briefly, the cells or EV preparations were lysed as previously
described (36). Pre-clear lysate
(100–500 µg total protein), according to the manufacturer's
instructions, were incubated in the presence of the primary mouse
antibodies (2 µg), Ago2 or Alix for 1 h at 4°C. To the
mixture, 20 µl of re-suspended volume of Protein A/G
PLUS-Agarose was added and incubated at 4°C on a rocker platform
overnight. Bound proteins were collected by centrifugation at 2,500
rpm and washed 4 times with 1.0 ml RIPA. After a final wash, the
pellets were re-suspended in boiling Laemmli buffer. Re-suspended
proteins were then subjected to western blot analysis as described
above.
Co-IP/protein-RNA interactions
The cell or EV preparations were lysed with lysis
buffer [20 mM Tris-HCl, 150 mM NaCl, 0.5% Nonidet P-40, 2 mM EDTA,
0.5 mM dithiothreitol (DTT), 1 mM NaF, 1 mM PMSF, 1% protease
inhibitor cocktail from Sigma (pH 7.5) and RNase inhibitor (800
U/ml)]. The immunoprecipitation assay was performed as previously
described (36). To evidence the
background RNA contamination, a normal mouse antibody was used.
After a final wash, the pellets were resuspended in buffer for RNA
isolation using an All-in-One purification kit (Norgen Biotek
Corp., Thorold, ON, Canada). RNA was spectrophotometrically
quantified (Nanodrop ND-1000 spectrophotometer; Nanodrop
Technologies, Wilmington, DE, USA).
The miRNAs associated with Alix/Ago2 Co-IP were
analyzed by reverse transcription-quantitative polymerase chain
reaction (RT-qPCR). cDNA obtained by reverse transcription of 200
ng of RNA using the miScript Reverse Transcription kit was then
subjected to RT-qPCR using the miScript SYBR Green PCR kit (both
from Qiagen, Valencia, CA, USA) to detect and quantify the miRNAs
of interest. Each reaction was run in triplicate using 4 ng of
cDNA. miRNA-specific primers to miR-24, miR-31, miR-125b, miR-99b,
miR-221, miR-16 and miR-21 were used. As positive controls, RNU6B
was used. RT-qPCR was accomplished using a 48-well StepOne
Real-Time System (Applied Biosystems, Foster City, CA, USA).
Negative cDNA controls (no cDNA) were cycled in parallel with each
run. Data are expressed as normalized values of miRNA mean
differences (2^−ΔΔCt ± SD).
miRNA transfer
The EV-mediated transfer of miRNAs to the HUVECs was
evaluated by RT-qPCR performed on total RNA extracted from the
HUVECs treated or not with HLSC-derived EVs and in the presence of
actinomycin D (ActD; Sigma) as previously described (14). As a control, HUVECs were treated
with 10 ng/ml of vascular endothelial growth factor (VEGF; PHC9394;
Thermo Fisher Scientific, Waltham, MA, USA) in the absence or
presence of ActD. Briefly, the HUVECs were treated for 30 min with
ActD (5 µg/ml), to inhibit RNA transcription. After washing,
the HUVECs were incubated with or without EVs (100 µg/ml) at
different time-points. Following reverse transcription of the RNA
(200 ng), the cDNA was subjected to RT-qPCR to quantify miRNAs as
described above.
HLSC transfection
The shRNA plasmids were provided by Santa Cruz
Biotechnology in a group of 3 to 5 lentiviral plasmids which act as
carriers. Each plasmid was coding for a specific 19–25 (plus
hairpins) shRNA designed for the reduction of gene expression.
shRNA sequences corresponding to the Alix siRNA gene were provided
as plasmid DNA ready for transfection. Following transfection, the
cells stably expressing shRNA were isolated by puromycin
selection.
The HLSCs were transfected using the Amaxa
Nucleofector technology (Amaxa, Koeln, Germany). The cells were
resuspended in a human MSC Nucleofector kit, according to the Amaxa
guidelines. Briefly, 100 µl of 4–5×105 cells
added with 2 µg cDNA were nucleofected with an Amaxa
Nucleofector apparatus using U-23 (for high transfection
efficiency) Nucleofector program. The cells were then transferred
to 6-well plates containing 37°C pre-warmed culture medium. The
positive control vector, pmaxGFP, which was rovided with the kit,
was used to establish the best Nucleofactor program. The cells were
observed under a Leica DMI3000 B microscope (Leica, Mannheim,
Germany). As a negative control, cells in Nucleofector solution
with DNA, but without application of the program and cells in
Nucleofector solution without DNA with application of the program
were used. Twenty-four hours following transfection, RNA was
extracted from the cells in order to assess Alix gene expression
decrease. Total RNA was extracted using a mirVana RNA isolation kit
(Ambion, Life Technologies, Carlsbad, CA, USA). cDNA was obtained
from total RNA using the High Capacity cDNA Reverse Transcription
kit (Applied Biosystems). For each cDNA synthesis, 200 ng mRNA, 2
µl RT buffer, 0.8 µl dNTP mixture, 2 µl RT
random primers, 1 µl MultiScribe reverse transcriptase and
4.2 µl nuclease-free water were used. A total of 100 nM of
each primer (Alix or 18S), were analyzed in the 48-well StepOne
Real-Time System (Applied Biosystems). In parallel to each run,
negative cDNA controls (no cDNA) were cycled. The following primers
were used: human Alix F1, 5′-GACGCTCCTGAGATATTATGA TCAGA-3′ and R1,
5′-ACACACAGCTCTTTTCATATCCTA AGC-3′; and human 18S F1,
5′-CCTGCCAGTAGCATATGC TTGTC-3′ and R1,
5′-AGCCATTCGCAGTTTCACTGTA-3′.
The decrease in Alix protein expression was
evaluated following puromycine selection (250 ng/ml). Alix
knockdown was evaluated in the HLSCs and the HLSC-derived EVs by
western blot analysis. Variations in the amount and profiles of the
released EVs were evaluated by NanoSight analysis.
Statistical analysis
The Kolmogorov-Smirnov test was used for the
evaluation of the normality of the distribution of data obtained by
FACS analysis. The SigmaPlot11.0 software was used for statistical
analysis and the Student's t-test was performed to evaluate the
differences between the different groups. Data are expressed as the
means ± SD. A value of P<0.05 was considered to indicate a
statistically significant difference.
Results
ESCRT marker expression in EVs released
from HLSCs
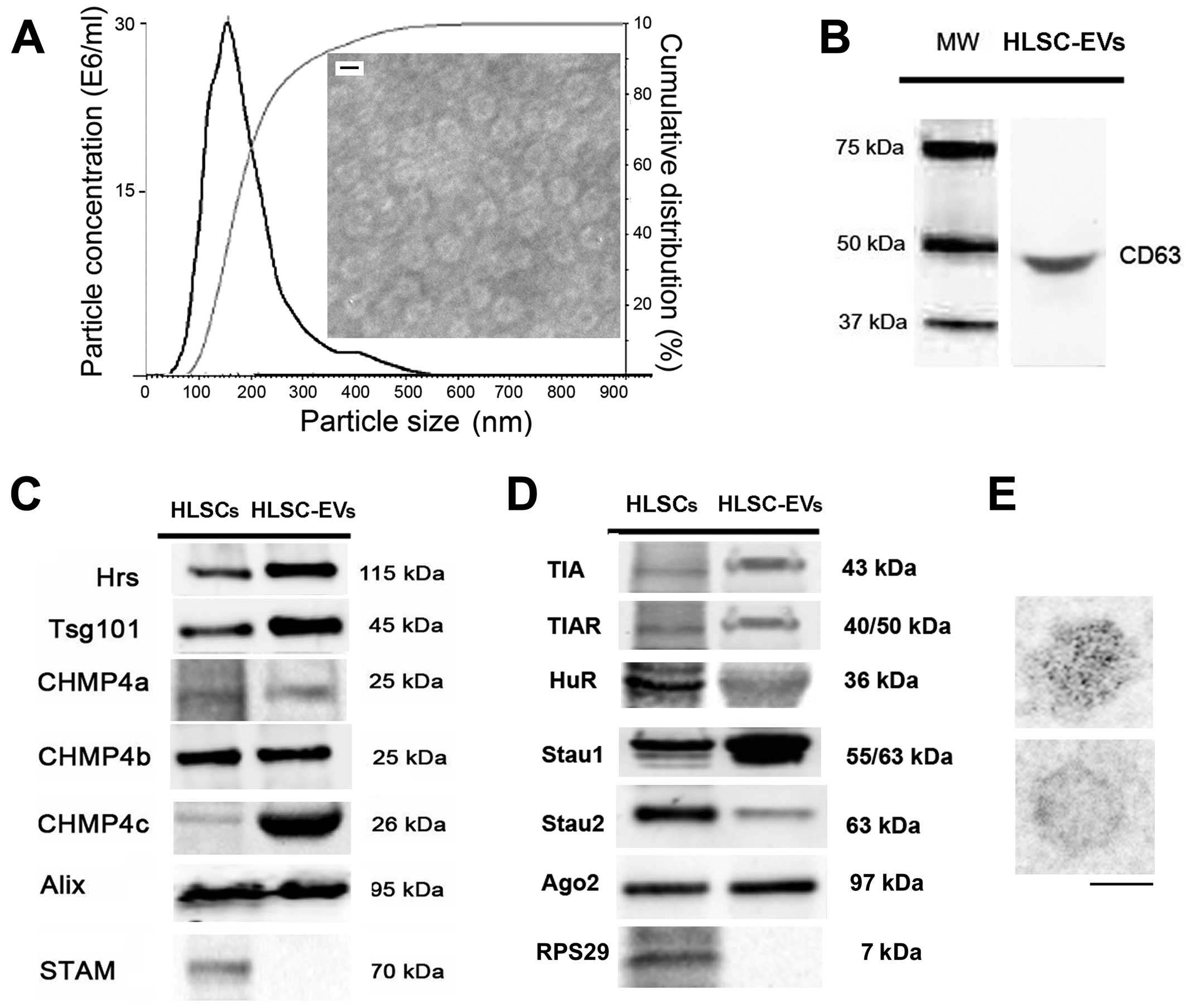
The 100k g purified fraction of the EVs revealed (by
NanoSight) a mean size of 164±30 nm (Fig. 1A). Using transmission electron
microscopy, the negatively stained EVs exhibited a round shape with
a dense core. Using electron microscopy, the size of the fixed EVs
was found to be approximately 100 nm, smaller than that observed
with NanoSight, which was influenced by the temperature.
FACS analysis revealed that the EVs expressed
several surface molecules of HLSCs, such as CD29, CD44, human
leukocyte antigen (HLA) I and integrin α5 (data not shown).
Moreover, the results of western blot analysis revealed that the
EVs expressed CD63 (Fig. 1B). The
EVs were obtained after 16 h of starvation and the amount of EVs
produced per single cell was 1321±256.
Western blot analysis revealed that the EVs
expressed, as did the cells of origin, several ESCRT proteins,
including Hrs, Tsg101, CHMP4a, CHMP4b, CHMP4c and Alix (Fig. 1C). When compared with the cells,
the expression of Hrs, Tsg101 and CHMP4c was increased in the EVs.
By contrast, STAM was detectable only in the cells and not in the
EVs. The positivity for ESCRT components in the EVs suggests their
transfer from HLSCs during vesiculation.
Moreover, the EVs expressed TIA-1, TIAR and HuR,
which are multifunctional ribonucleoproteins known to be expressed
in nuclei and stress granules (37) (Fig.
1D). In addition, they expressed Stau 1 and 2 proteins, which
are involved in the transport and stability of mRNAs (17), as well as Ago2, a protein of the
Argonaute family, which is involved in the transport and processing
of miRNAs (18) (Fig. 1D). The expression of Ago2 was
confirmed by immunogold electron microscopy (Fig. 1E). The presence within the
HLSC-derived EVs of selected ribonucleoproteins suggests a role in
mRNA and miRNA storage.
We have previously demonstrated an enrichment of
miRNAs within HLSC-derived EVs (14). Therefore, in this study, we
focused on the inter action between Alix, one of the main expressed
components of the ESCRT family in EVs, and Ago2 by performing Co-IP
experiments.
Alix and Ago2 interaction in HLSC-derived
EVs
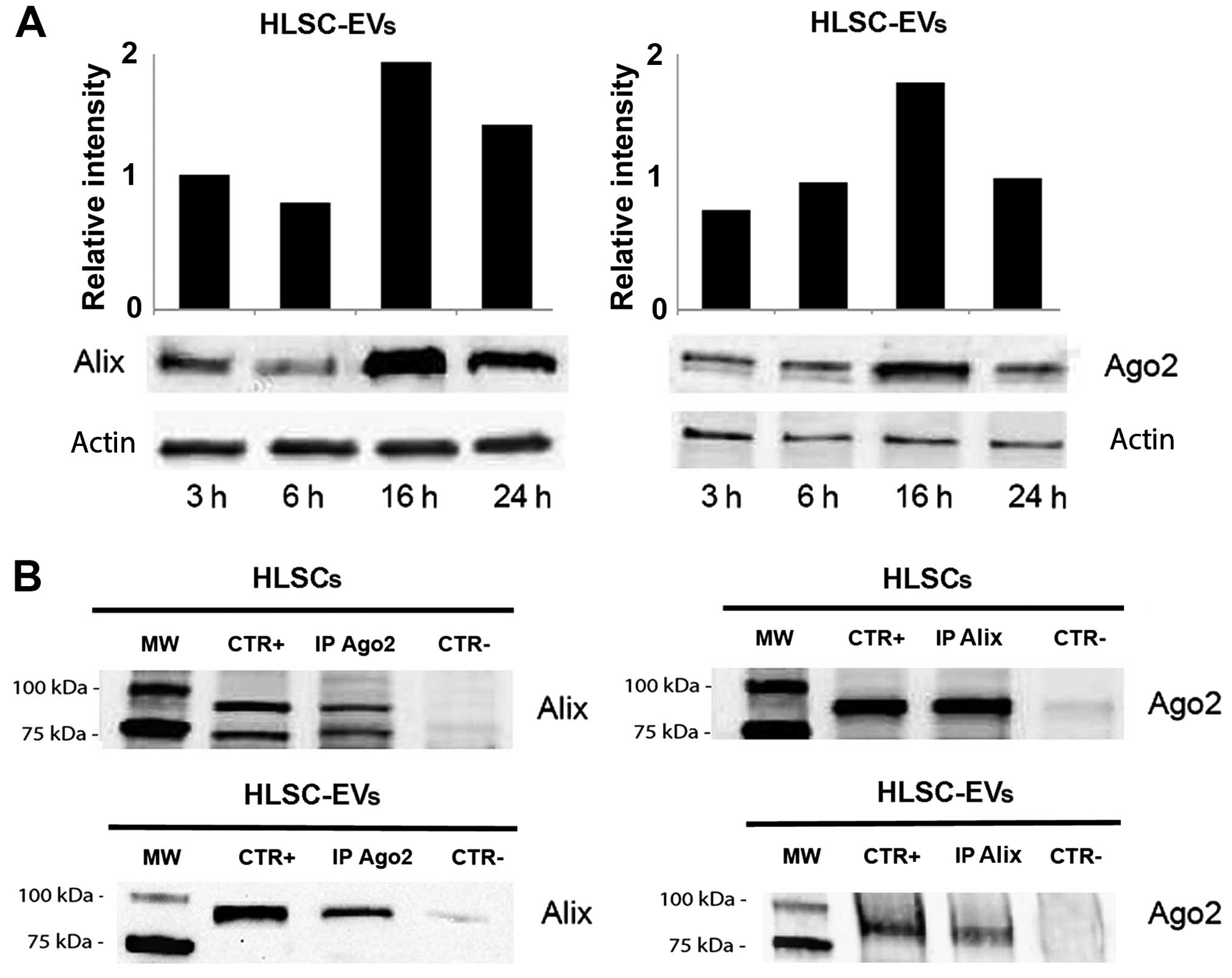
In kinetic experiments, we found that the expression
of Alix and Ago2 in the EVs was highest after 16 h of HLSC
starvation (Fig. 2A). As shown in
Fig. 2B, an interaction between
Alix and Ago2 was observed in the immunoprecipitates of the HLSCs
and the EVs. We therefore investigated whether the Alix/Ago2
complex was involved in the enrichment of miRNAs within the
HLSC-derived EVs.
miRNA association with the Alix/Ago2
complex
On the basis of previous experiments that identified
miRNAs with antitumor or regenerative activity in EVs derived from
HLSCs (23,38), we evaluated the enrichment of
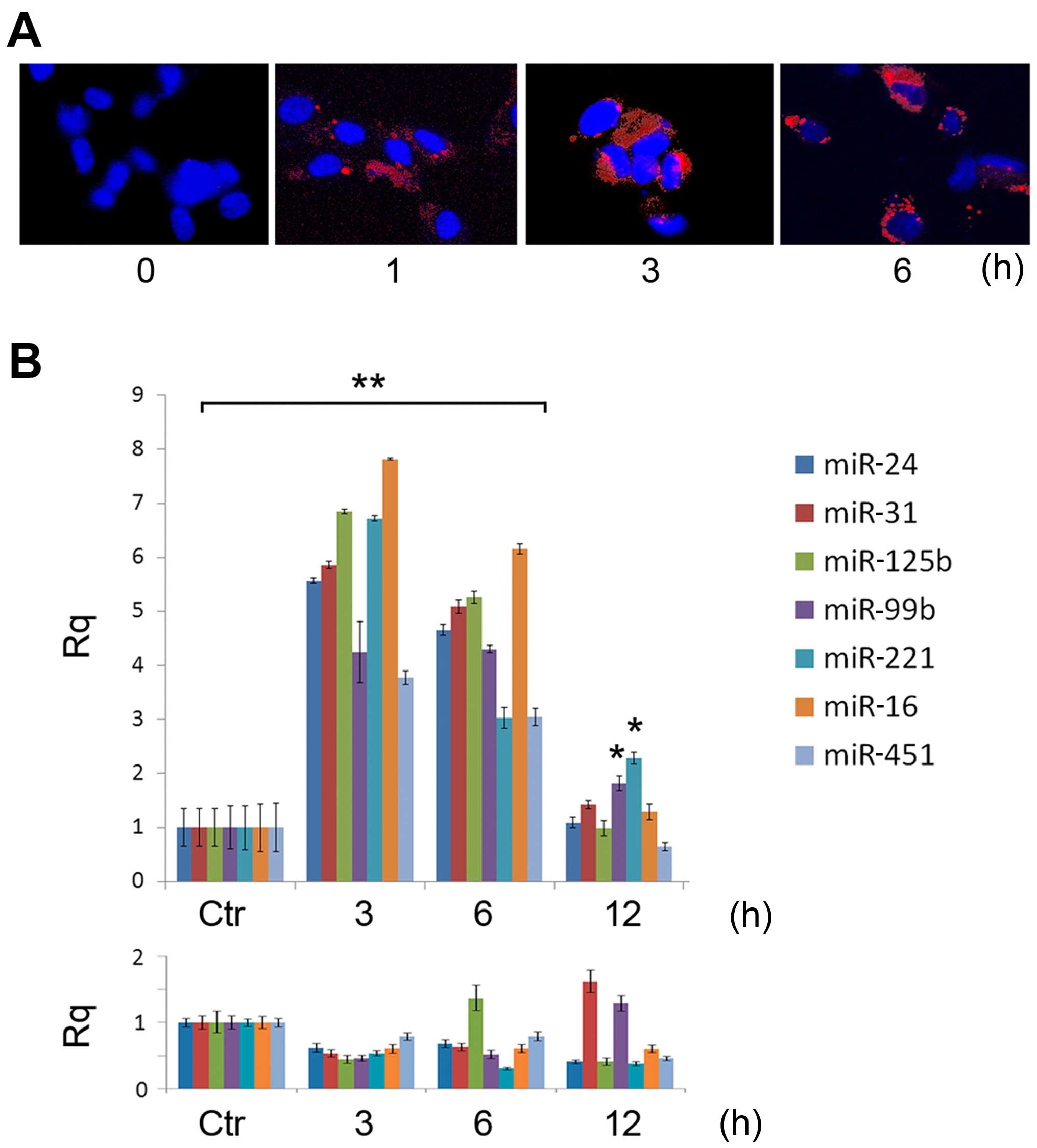
these miRNAs within the EVs. Fig.
3A shows the log-relative quantification of miR-24, miR-31,
miR-125b, miR-99b, miR-221, miR-16 and miR-451 in the EVs with
respect to the HLSCs evaluated by RT-qPCR. These results suggest
that during vesiculation, selected miRNAs are compartmentalized
within EVs.
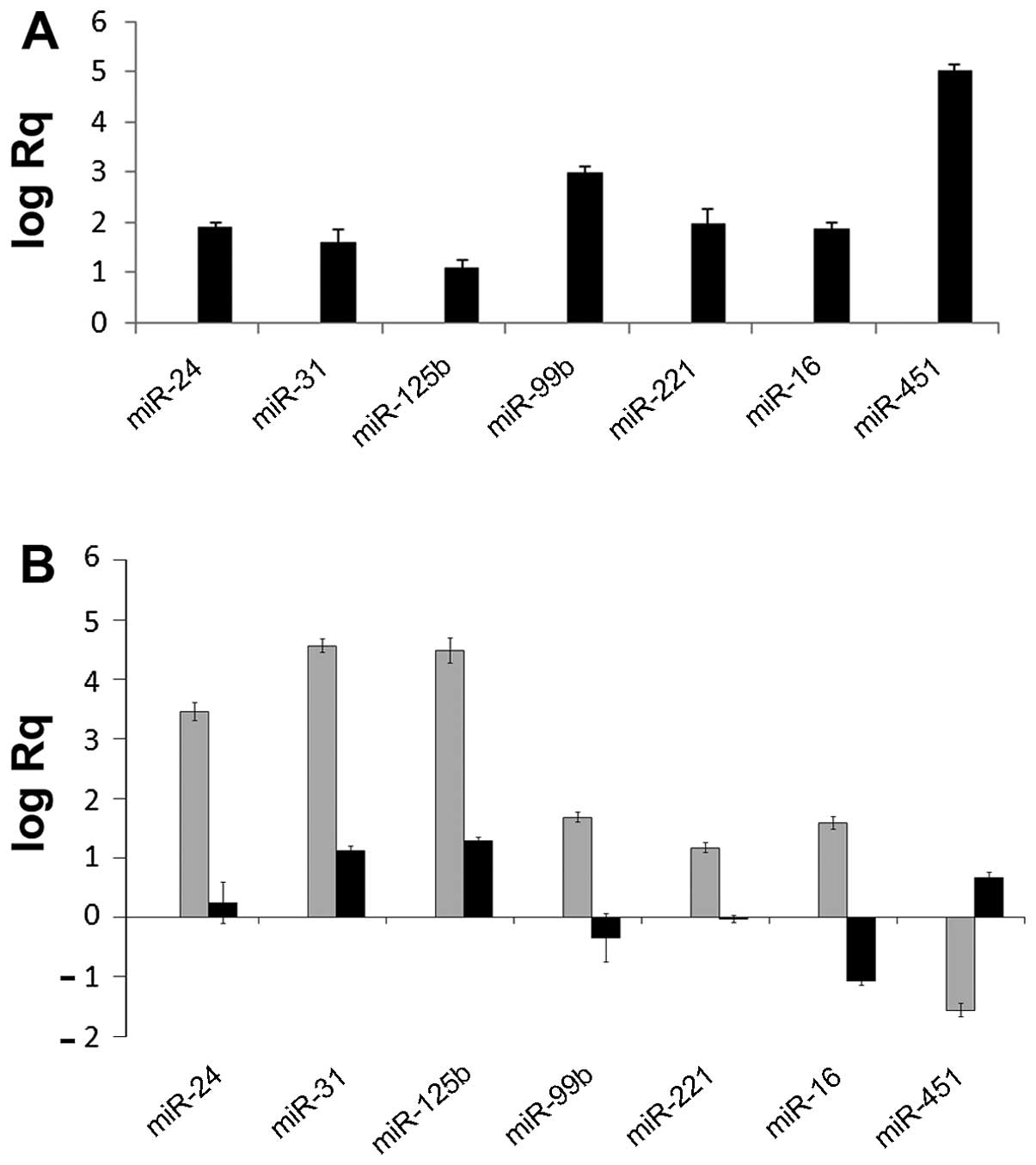 | Figure 3miRNA expression in human liver
stem-like cell (HLSC)-derived extracellular vesicles (EVs;
HLSC-EVs). (A) The expression of miR24, miR31, miR125b, miR99b,
miR221, miR16 and miR451 was evaluated by RT-qPCR in HLSC-derived
EVs with respect to HLSCs. Data are expressed as log of relative
quantification (Rq), normalized to RNU6B and to one for HLSC. These
miRNAs were enriched in EVs in respect to their relative HLSC. Data
are the means ± SD of 4 experiments. (B) The expression of miR24,
miR31, miR125b, miR99b, miR221, miR16 and miR451 was evaluated by
RT-qPCR in HLSC-derived EVs immunoprecipitated with Ago2 (gray
bars) and in HLSC-derived EVs immunoprecipitated with Alix (black
bars) with respect to 100k g purified EVs. Data are expressed as
the log of Rq, normalized to RNU6B and to one for 100k g purified
EVs. Data are the means ± SD of 4 experiments. |
Similarly, miRNA enrichment was observed in the
Alix/Ago2 immunoprecipitate complex (Fig. 3B), suggesting that the Alix/Ago2
complex was instrumental in miRNA accumulation within EVs.
To evaluate whether EVs are instrumental in
transferring miRNAs to target cells, we used HUVECs as recipient
cells. The extent of EV incorporation into the HUVECs was evaluated
by FACS analysis (data not shown) and confocal microscopy (Fig. 4A). The PKH26-labeled HLSC-derived
EVs were already detectable in the HUVECs following 1 h of
incubation, with a maximum internalization after 3–6 h (Fig. 4A). RT-qPCR was performed on RNA
from the HUVECs treated or not with HLSC-derived EVs in the
presence of ActD in order to block RNA transcription. The effective
block of RNA transcription by ActD was evaluated by monitoring
miR-126 and miR-31 expression in the HUVECs stimulated with VEGF
(10 µg/ml). The enhanced expression of these miRNAs was
inhibited by ActD (data not shown). As shown in Fig. 4B, the accumulation of miRNAs in
the HUVECs treated with EVs peaked after 3 h to progressively
decrease at 12 h. miRNA expression in the unstimulated HUVECs did
not significantly vary with time.
Alix knockdown experiments were performed by
plasmidic electroporation in HLSCs of shRNA against Alix to assess
its effects on the release of EVs and miRNA content. The
transfection efficiency evaluated as the green fluorescence protein
(GFP) signal ranged between 80–90% in the HLSCs after 24 h. At this
time, a significant decrease in the Alix mRNA levels was detected
by RT-qPCR. Moreover, the decrease in Alix protein expression was
detected up to 2 weeks following selection with puromycin (Fig. 5A). Decreased Alix expression was
also observed in the EVs derived from the HLSCs in which Alix had
been knocked down. Moreover, the expression of Ago2 was
significantly decreased in the EVs (Fig. 5B), but not in the HLSCs (data not
shown), suggesting that Alix is required for Ago2 localization
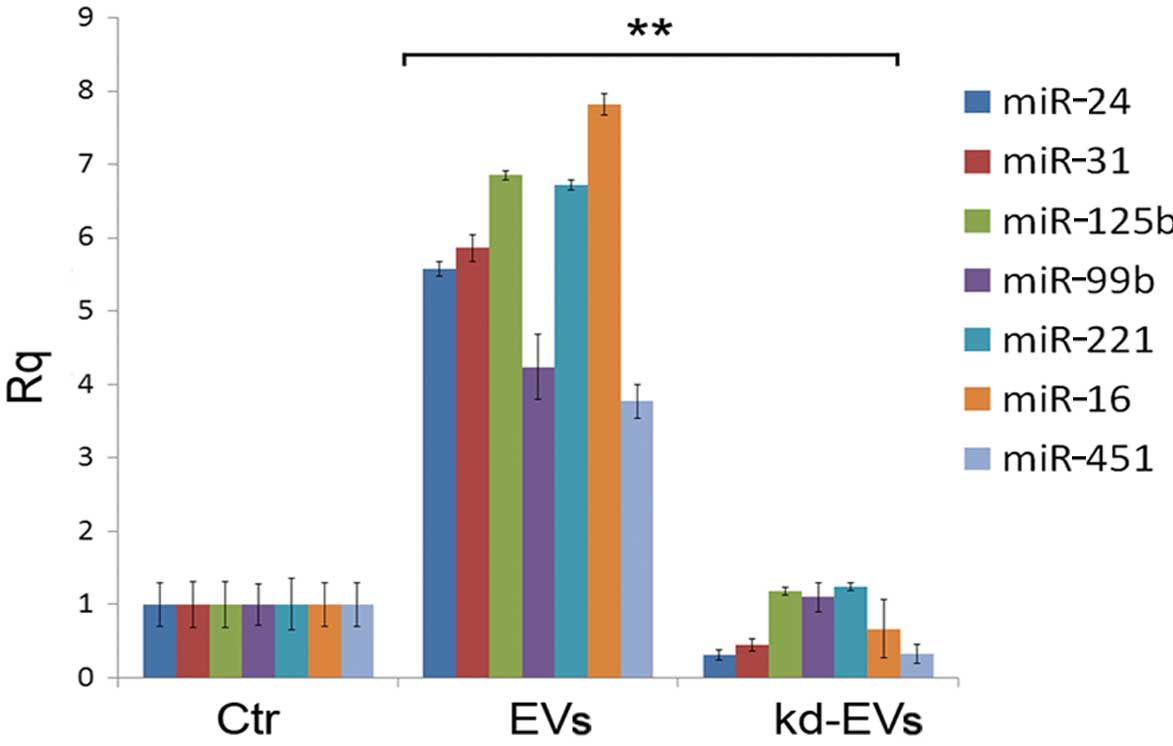
within EVs. As shown in Fig. 5C,
Alix knockdown in the HLSCs significantly reduced miRNA
compartmentalization in the EVs, confirming a critical role of the
Alix/Ago2 complex in driving miRNAs within EVs. The reduction in
the EV miRNA content resulted in a significant decrease in miRNA
transfer to the HUVECs (Fig. 6).
By contrast, Alix knockdown did not influence the amount of
vesicles produced by the HLSCs as detected by NanoSight (wild-type
HLSCs, 3.07±1.5 ×108 particles/ml; knockdown HLSCs,
3.68±1.5 ×108 particles/ml) suggesting that Alix was
instrumental in the composition of EVs rather than in their
secretion.
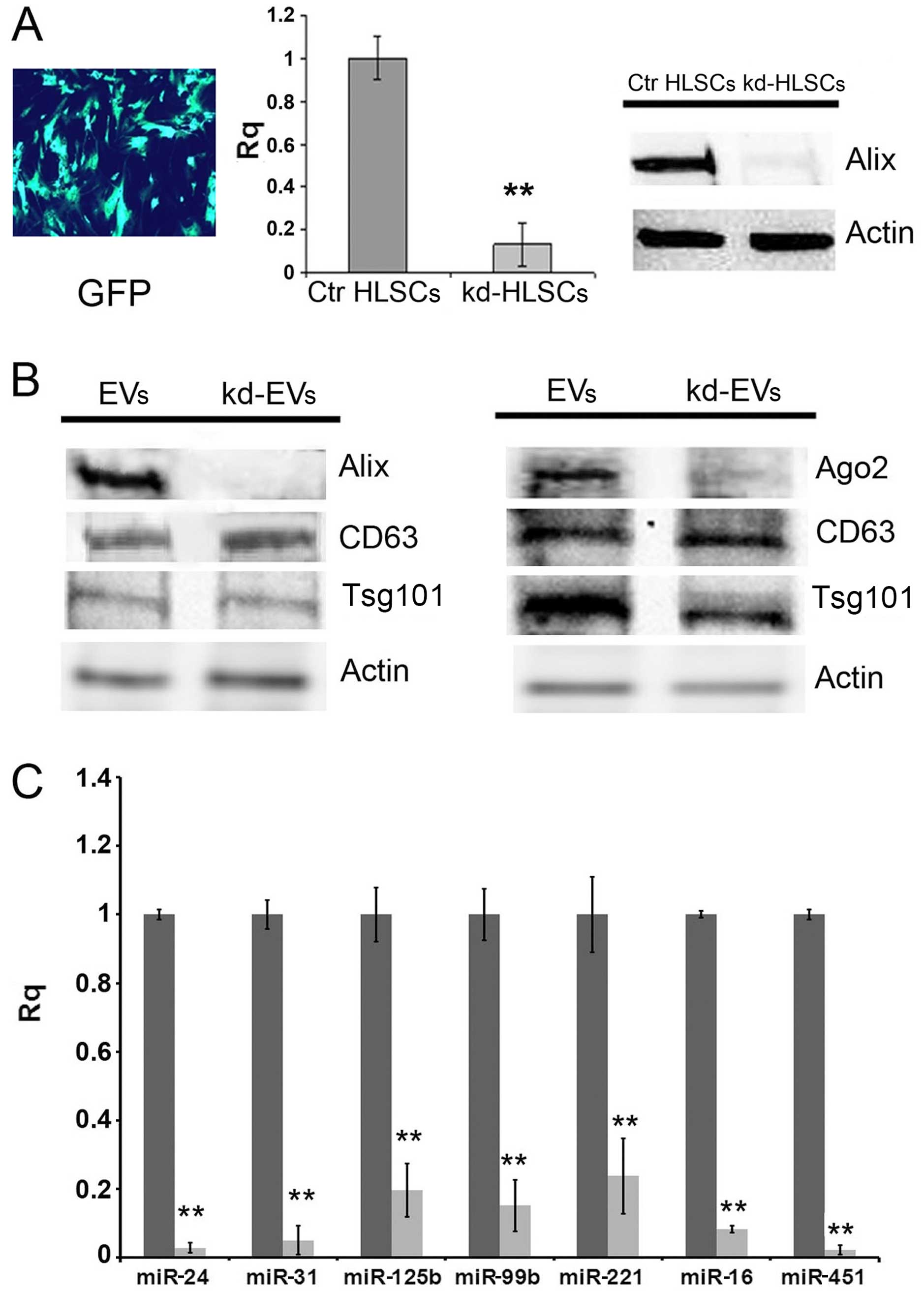 | Figure 5Effects of Alix knockdown in human
liver stem-like cells (HLSCs). (A) Representative image of HLSCs
transfected with the positive control vector, pmaxGFP, 24 h after
transfection (original magnification, ×100); Alix mRNA expression
in HLSCs in which Alix was knocked down (kd-HLSCs) with respect to
normal HLSCs (Ctr HLSCs) set to one, was evaluated by RT-qPCR. Data
are the means ± SD of 3 experiments expressed as relative
quantification (Rq), normalized to 18S. Representative western
blots of Alix protein expression in kd-HLSCs with respect to Ctr
HLSCs. Equal loading was confirmed by stripping the immunoblot and
reprobing it for actin. Three different experiments were performed
with similar results. (B) Representative western blots of Alix and
Ago2 protein expression in extracellular vesicles (EVs) released
from normal HLSCs (EVs) and in EVs released from HLSCs in which
Alix was knocked down (kd-EVs). Equal loading was confirmed by
stripping the immunoblot and reprobing it for CD63 and Tsg101.
Three different experiments were performed with similar results.
(C) The expression of miR-24, miR-31, miR-125b, miR-99b, miR-221,
miR-16 and miR-451 was evaluated by RT-qPCR in EVs derived from
HLSCs in which Alix was knocked down (gray bars) with respect to
EVs derived from normal HLSCs (black bars) set to one. Data are
expressed as Rq, normalized to RNU6B. Data are the means ± SD of 4
experiments. **P<0.001 vs. HLSC-derived EVs. |
Discussion
The results of the present study demonstrated that
the EVs released from the HLSCs are enriched in proteins involved
in ESCRT machinery, including Alix and miRNAs. Moreover, they
contain several RNA-binding proteins, including Ago2, a component
of the Argonaute family known to be involved in miRNA transport and
processing. Co-IP experiments demonstrated that Alix binds to Ago2
and miRNAs, suggesting that Alix plays a key role in the miRNA
enrichment during the biogenesis of EVs. We found that Alix was
involved in the molecular composition of vesicles released from
HLSCs, but not in their secretion. In fact, Alix knockdown did not
influence the number of EVs released by the HLSCs, but it
significantly decreased miRNA expression in the EVs and,
consequently, their transfer to the endothelium.
EVs, important mediators of intercellular
communication, have also recently emerged as new diagnostic
(38) and therapeutic (39) tools. EVs have been shown to be
involved in immune modulation (40), tumor biology (41) and tissue regeneration (42). The mechanisms of action of EVs are
associated with their ability to directly stimulate target cells
and/or to transfer proteins, biologically active lipids and nucleic
acids. In particular, the transfer of transcription factors, mRNAs
and non-coding RNAs has been shown to induce epigenetic
modifications in recipient cells, leading to changes in cell
function and phenotype (43). The
understanding of the mechanisms involved in the biogenesis
(6) and the loading of EV cargo
is crucial, not only for the definition of their physiological
role, but also for their possible engineering and exploitation in
therapy (42,44).
The compartment of biologically active molecules
within EVs is modified by environmental and metabolic conditions,
suggesting a tightly regulated mechanism which remains to be
elucidated.
Alix has been implicated in the sorting of exosomes
from infected and MCF7 cells during vesiculation (23–25). ESCRT machinery has been shown to
participate in processes of vesiculation and cargo sorting,
including multivesicular body biogenesis and the release of
enveloped viruses (45–47). In the present study, we found that
ESCRT proteins were expressed in HLSCs and in the released EVs,
suggesting a role in EV formation. Alix, an accessory protein of
the ESCRT complex, has multifunctional activities due to its
domains, which provide multiple protein-binding sites (24,28–33). In particular, it has been
demonstrated that Alix is involved in endocytic membrane
trafficking and cytoskeletal remodeling (27,29,33,48). The N-terminal 'Bro1 domain' allows
endosomal localization, whereas the C-terminal region may interact
with proteins that connect Alix with different cellular processes
(28–33). Several viruses exploit Alix in
order to bud from infected cells. An interaction between the
nucleocapsid (NC) domain of HIV and the Bro1 domain of Alix has
been demonstrated (32). In that
study, it was suggested that RNA may serve as a bridge for Bro1-NC
interactions (32). Of note, Shen
et al established a parallelism between EV biogenesis and
retrovirus budding, suggesting that plasma membrane anchors may
target highly oligomeric cytoplasmic proteins to EVs (49). They also demonstrated that
higher-order oligomerization was the main determinant of HIV Gag
budding/exosomal sorting (50,51).
It has been suggested that the enrichment of
extracellular RNA within EVs depends on its association with
RNA-binding proteins (14,52).
We found that the EVs released from the HLSCs contained several
RNA-binding proteins including Ago2, which is involved in miRNA
transport and processing (18).
Herein, we suggest that Alix, by forming a complex with Ago2,
drives it within EVs together with the associated miRNAs. The
observation that Alix knockdown in HLSCs prevented the accumulation
of Ago2 and miRNAs in EVs, as well as the consequent transfer of
miRNAs to target cells, further supports the relevance of this
protein in modulating the composition of EVs.
In the future, it would be interesting in the future
to evaluate whether the role of Alix in the enrichment of miRNA
species in EVs is a general mechanism involved in other cell types.
In conclusion, the results of the present study suggest a novel
function of Alix, demonstrating its involvement in the composition
of EVs released from HLSCs.
Acknowledgments
The present study was supported by the Associazione
Italiana per la Ricerca sul Cancro (AIRC) project IG 12890 and by
the National Center For Advancing Translational Sciences of the
National Institutes of Health under Award number UH2TR000880. The
content is solely the responsibility of the authors and does not
necessarily represent the official views of the National Institutes
of Health.
Abbreviations:
|
EVs
|
extracellular vesicles
|
|
HLSCs
|
human liver stem-like cells
|
|
HUVECs
|
human umbilical cord vein-derived
endothelial cells
|
|
ESCRT
|
endosomal sorting complex required for
transport
|
|
Ago2
|
argonaute 2
|
|
miRNAs
|
microRNAs
|
|
Stau 1
|
Staufen 1
|
|
Stau 2
|
Staufen 2
|
|
TIA-1
|
T-cell internal antigen-1
|
|
TIAR
|
TIA-1-related protein
|
|
HuR
|
human antigent R
|
|
Co-IP
|
co-immunoprecipitation
|
|
RT-qPCR
|
reverse transcriptase-quantitative
polymerase chain reaction
|
|
Rq
|
relative quantification
|
|
ActD
|
actinomycin D
|
References
|
1
|
Quesenberry PJ, Goldberg LR, Aliotta JM,
Dooner MS, Pereira MG, Wen S and Camussi G: Cellular phenotype and
extracellular vesicles: basic and clinical considerations. Stem
Cells Dev. 23:1429–1436. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ratajczak J, Wysoczynski M, Hayek F,
Janowska-Wieczorek A and Ratajczak MZ: Membrane-derived
microvesicles: important and underappreciated mediators of
cell-to-cell communication. Leukemia. 20:1487–1495. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Théry C, Ostrowski M and Segura E:
Membrane vesicles as conveyors of immune responses. Nat Rev
Immunol. 9:581–593. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lee TH, D'Asti E, Magnus N, Al-Nedawi K,
Meehan B and Rak J: Microvesicles as mediators of intercellular
communication in cancer - the emerging science of cellular
'debris'. Semin Immunopathol. 33:455–467. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cocucci E, Racchetti G and Meldolesi J:
Shedding microvesicles: artefacts no more. Trends Cell Biol.
19:43–51. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Colombo M, Raposo G and Théry C:
Biogenesis, secretion, and intercellular interactions of exosomes
and other extracellular vesicles. Annu Rev Cell Dev Biol.
30:255–289. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gould SJ and Raposo G: As we wait: coping
with an imperfect nomenclature for extracellular vesicles. J
Extracell Vesicles. 2:Feb 15–2013.Epub ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen X, Liang H, Zhang J, Zen K and Zhang
CY: Secreted microRNAs: a new form of intercellular communication.
Trends Cell Biol. 22:125–132. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang K, Zhang S, Weber J, Baxter D and
Galas DJ: Export of microRNAs and microRNA-protective protein by
mammalian cells. Nucleic Acids Res. 38:7248–7259. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gallo A, Tandon M, Alevizos I and Illei
GG: The majority of microRNAs detectable in serum and saliva is
concentrated in exosomes. PLoS One. 7:e306792012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ratajczak J, Miekus K, Kucia M, Zhang J,
Reca R, Dvorak P and Ratajczak MZ: Embryonic stem cell-derived
microvesicles reprogram hematopoietic progenitors: evidence for
horizontal transfer of mRNA and protein delivery. Leukemia.
20:847–856. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Deregibus MC, Cantaluppi V, Calogero R, Lo
Iacono M, Tetta C, Biancone L, Bruno S, Bussolati B and Camussi G:
Endothelial progenitor cell derived microvesicles activate an
angiogenic program in endothelial cells by a horizontal transfer of
mRNA. Blood. 110:2440–2448. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Valadi H, Ekström K, Bossios A, Sjöstrand
M, Lee JJ and Lötvall JO: Exosome-mediated transfer of mRNAs and
microRNAs is a novel mechanism of genetic exchange between cells.
Nat Cell Biol. 9:654–659. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Collino F, Deregibus MC, Bruno S, Sterpone
L, Aghemo G, Viltono L, Tetta C and Camussi G: Microvesicles
derived from adult human bone marrow and tissue specific
mesenchymal stem cells shuttle selected pattern of miRNAs. PLoS
One. 5:e118032010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Aliotta JM, Sanchez-Guijo FM, Dooner GJ,
Johnson KW, Dooner MS, Greer KA, Greer D, Pimentel J, Kolankiewicz
LM, Puente N, et al: Alteration of marrow cell gene expression,
protein production, and engraftment into lung by lung-derived
microvesicles: a novel mechanism for phenotype modulation. Stem
Cells. 25:2245–2256. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gibbings D, Leblanc P, Jay F, Pontier D,
Michel F, Schwab Y, Alais S, Lagrange T and Voinnet O: Human prion
protein binds Argonaute and promotes accumulation of microRNA
effector complexes. Nat Struct Mol Biol. 19:517–524. S12012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Laver JD, Li X, Ancevicius K, Westwood JT,
Smibert CA, Morris QD and Lipshitz HD: Genome-wide analysis of
Staufen-associated mRNAs identifies secondary structures that
confer target specificity. Nucleic Acids Res. 41:9438–9460. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Waris S, Wilce MC and Wilce JA: RNA
recognition and stress granule formation by TIA proteins. Int J Mol
Sci. 15:23377–23388. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Höck J and Meister G: The Argonaute
protein family. Genome Biol. 9(210)2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Herrera MB, Fonsato V, Bruno S, Grange C,
Gilbo N, Romagnoli R, Tetta C and Camussi G: Human liver stem cells
improve liver injury in a model of fulminant liver failure.
Hepatology. 57:311–319. 2013. View Article : Google Scholar
|
|
21
|
Herrera MB, Fonsato V, Gatti S, Deregibus
MC, Sordi A, Cantarella D, Calogero R, Bussolati B, Tetta C and
Camussi G: Human liver stem cell-derived microvesicles accelerate
hepatic regeneration in hepatectomized rats. J Cell Mol Med.
14:1605–1618. 2010. View Article : Google Scholar
|
|
22
|
Herrera MB, Bruno S, Buttiglieri S, Tetta
C, Gatti S, Deregibus MC, Bussolati B and Camussi G: Isolation and
characterization of a stem cell population from adult human liver.
Stem Cells. 24:2840–2850. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fonsato V, Collino F, Herrera MB,
Cavallari C, Deregibus MC, Cisterna B, Bruno S, Romagnoli R,
Salizzoni M, Tetta C and Camussi G: Human liver stem cell-derived
microvesicles inhibit hepatoma growth in SCID mice by delivering
antitumor microRNAs. Stem cells. 30:1985–1998. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Martin-Serrano J, Yarovoy A,
Perez-Caballero D and Bieniasz PD: Divergent retroviral
late-budding domains recruit vacuolar protein sorting factors by
using alternative adaptor proteins. Proc Natl Acad Sci USA.
100:12414–12419. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Baietti MF, Zhang Z, Mortier E, Melchior
A, Degeest G, Geeraerts A, Ivarsson Y, Depoortere F, Coomans C,
Vermeiren E, et al: Syndecan-syntenin-ALIX regulates the biogenesis
of exosomes. Nat Cell Biol. 14:677–685. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Roucourt B, Meeussen S, Bao J, Zimmermann
P and David G: Heparanase activates the syndecan-syntenin-ALIX
exosome pathway. Cell Res. 25:412–428. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ghossoub R, Lembo F, Rubio A, Gaillard CB,
Bouchet J, Vitale N, Slavík J, Machala M and Zimmermann P:
Syntenin-ALIX exosome biogenesis and budding into multivesicular
bodies are controlled by ARF6 and PLD2. Nat Commun. 5(3477)2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Missotten M, Nichols A, Rieger K and
Sadoul R: Alix, a novel mouse protein undergoing calcium-dependent
interaction with the apoptosis-linked-gene 2 (ALG-2) protein. Cell
Death Differ. 6:124–129. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Odorizzi G, Katzmann DJ, Babst M, Audhya A
and Emr SD: Bro1 is an endosome-associated protein that functions
in the MVB pathway in Saccharomyces cerevisiae. J Cell Sci.
116:1893–1903. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schmidt MH, Hoeller D, Yu J, Furnari FB,
Cavenee WK, Dikic I and Bögler O: Alix/AIP1 antagonizes epidermal
growth factor receptor downregulation by the Cbl-SETA/CIN85
complex. Mol Cell Biol. 24:8981–8993. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Strack B, Calistri A, Craig S, Popova E
and Göttlinger HG: AIP1/ALIX is a binding partner for HIV-1 p6 and
EIAV p9 functioning in virus budding. Cell. 114:689–699. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sette P, Dussupt V and Bouamr F:
Identification of the HIV-1 NC binding interface in Alix Bro1
reveals a role for RNA. J Virol. 86:11608–11615. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Morita E, Sandrin V, Chung HY, Morham SG,
Gygi SP, Rodesch CK and Sundquist WI: Human ESCRT and ALIX proteins
interact with proteins of the midbody and function in cytokinesis.
EMBO J. 26:4215–4227. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dragovic RA, Gardiner C, Brooks AS,
Tannetta DS, Ferguson DJ, Hole P, Carr B, Redman CW, Harris AL,
Dobson PJ, et al: Sizing and phenotyping of cellular vesicles using
Nanoparticle Tracking Analysis. Nanomedicine. 7:780–788. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lopatina T, Bruno S, Tetta C, Kalinina N,
Porta M and Camussi G: Platelet-derived growth factor regulates the
secretion of extracellular vesicles by adipose mesenchymal stem
cells and enhances their angiogenic potential. Cell Commun Signal.
12(26)2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Deregibus MC, Buttiglieri S, Russo S,
Bussolati B and Camussi G: CD40-dependent activation of
phosphatidylinositol 3-kinase/Akt pathway mediates endothelial cell
survival and in vitro angiogenesis. J Biol Chem. 278:18008–18014.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Anderson P and Kedersha N: RNA granules. J
Cell Biol. 172:803–808. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lin J, Li J, Huang B, Liu J, Chen X, Chen
XM, Xu YM, Huang LF and Wang XZ: Exosomes: novel biomarkers for
clinical diagnosis. Scientific World Journal. 2015(657086)2015.
View Article : Google Scholar
|
|
39
|
Katoh M: Therapeutics targeting
angiogenesis: genetics and epigenetics, extracellular miRNAs and
signaling networks (Review). Int J Mol Med. 32:763–767.
2013.PubMed/NCBI
|
|
40
|
Chaput N and Théry C: Exosomes: Immune
properties and potential clinical implementations. Semin
Immunopathol. 33:419–440. 2011. View Article : Google Scholar
|
|
41
|
Milane L, Singh A, Mattheolabakis G,
Suresh M and Amiji MM: Exosome mediated communication within the
tumor micro-environment. J Control Release. 219:278–294. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
De Jong OG, Van Balkom BW, Schiffelers RM,
Bouten CV and Verhaar MC: Extracellular vesicles: potential roles
in regenerative medicine. Front Immunol. 5(608)2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Quesenberry PJ, Aliotta J, Deregibus MC
and Camussi G: Role of extracellular RNA-carrying vesicles in cell
differentiation and reprogramming. Stem Cell Res Ther. 6(153)2015.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhang Y, Wu XH, Luo CL, Zhang JM, He BC
and Chen G: Interleukin-12-anchored exosomes increase cytotoxicity
of T lymphocytes by reversing the JAK/STAT pathway impaired by
tumor-derived exosomes. Int J Mol Med. 25:695–700. 2010.PubMed/NCBI
|
|
45
|
Morita E: Differential requirements of
mammalian ESCRTs in multivesicular body formation, virus budding
and cell division. FEBS J. 279:1399–1406. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Roxrud I, Stenmark H and Malerød L: ESCRT
& Co. Biol Cell. 102:293–318. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Votteler J and Sundquist WI: Virus budding
and the ESCRT pathway. Cell Host Microbe. 14:232–241. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Pan S, Wang R, Zhou X, He G, Koomen J,
Kobayashi R, Sun L, Corvera J, Gallick GE and Kuang J: Involvement
of the conserved adaptor protein Alix in actin cytoskeleton
assembly. J Biol Chem. 281:34640–34650. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Shen B, Wu N, Yang JM and Gould SJ:
Protein targeting to exosomes/microvesicles by plasma membrane
anchors. J Biol Chem. 286:14383–14395. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Fang Y, Wu N, Gan X, Yan W, Morrell JC and
Gould SJ: Higher-order oligomerization targets plasma membrane
proteins and HIV gag to exosomes. PLoS Biol. 5:e1582007. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Booth AM, Fang Y, Fallon JK, Yang JM,
Hildreth JE and Gould SJ: Exosomes and HIV Gag bud from
endosome-like domains of the T cell plasma membrane. J Cell Biol.
172:923–935. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Li L, Zhu D, Huang L, Zhang J, Bian Z,
Chen X, Liu Y, Zhang CY and Zen K: Argonaute 2 complexes
selectively protect the circulating microRNAs in cell-secreted
microvesicles. PLoS One. 7:e469572012. View Article : Google Scholar : PubMed/NCBI
|