Introduction
Colorectal cancer is the third most frequent cause
of mortality in males and females, when the genders are considered
separately and the second most frequent cause when they are
considered together. In fact, colorectal cancer-related mortality
accounts for approximately 10% of all cancer-related deaths
(1). Although surgical resection
is considered the most effective and curative treatment for various
types of cancer, the majority of patients with colon cancer require
neoadjuvant or adjuvant chemotherapy before and after surgery.
Thus, chemotherapy is the most widely used form of treatment for
colon cancer, whether used as monotherapy or as adjuvant therapy
(2–4).
Colon cancer cells often become resistant during
chemotherapy, which limits treatment efficacy and worsens patient
prognosis (5,6). Researchers are actively engaged in
elucidating the mechanisms responsible for resistance to treatment
in aim to identify and develop strategies to overcome it, not only
in colon cancer, but also in other types of malignant disease
(7).
A major mechanism of cancer cell chemoresistance is
thought to be autophagy (8).
Autophagy is an otherwise normal process in which dysfunctional
cytoplasmic proteins and organelles are sequestered in
intracellular autophagosomes that fuse with lysosomes and are
degraded. Autophagy normally occurs as an adaptive response to
cellular stresses, such as nutrient starvation or metabolic stress.
Autophagosome formation is triggered by class III phosphoinositide
3-kinase and Beclin-1, also known as the mammalian homologue of
yeast autophagy-related gene 6 (9). Consistent with its critical role in
autophagosome formation, Beclin-1 expression increases during
autophagy (10). During this
process, microtubule-associated protein light chain 3 (LC3) is
converted from the cytosolic form, LC3-I, to the autophagic
membrane-bound form, LC3-II; relative amounts of LC3-I and II serve
as an index of autophagy (11).
Autophagy is thought to promote chemoresistance by promoting
cellular energy production. Consistent with this hypothesis, the
inhibition of autophagy using 3-methyladenine and chloroquine has
been shown to enhance the chemosensitivity of hepatocellular
carcinoma and supraglottic laryngeal cancer cells (12–14). These data suggest that the
inhibition of autophagy may reduce the risk or level of
chemoresistance in colon cancer cells.
It may be possible to regulate autophagy by
manipulating the levels of microRNAs (miRNAs or miRs), since these
post-transcriptional regulators help to control numerous cellular
processes (15). These short
RNAs, 18–24 nt in length, bind to the 3′-untranslated region (UTR)
of target mRNAs, destabilizing them and thereby inhibiting protein
translation (16). Several miRNAs
have been associated with tumorigenesis, response to treatment and
patient survival (17–20). However, whether miRNAs play a role
in the chemoresistance of cancer cells, including colon cancer
cells, remains largely unknown.
The miRNA miR-409-3p has been shown to be
significantly downregulated in fibrosarcoma, bladder and gastric
cancer (21–23). Thus, we wished to determine
whether miR-409-3p plays a role in chemoresistance in colon cancer.
In the present study, we demonstrate firstly using bioinformatics
analysis, and then using a dualluciferase assay in LoVo human colon
cancer cells, that miR-409-3p binds to the 3′-UTR of Beclin-1 mRNA
and downregulates its expression. We also demonstrate that
miR-409-3p expression is downregulated, whereas Beclin-1 expression
and autophagic activity are markedly upregulated, in chemoresistant
LoVo cell cultures. The overexpression of miR-409-3p in the same
cell line or treatment of the cells with miR-409-3p mimic in the
medium, significantly suppressed Beclin-1 expression and autophagic
activity, which rendered the cell lines sensitive to oxaliplatin
therapy. We concluded that increasing miR-409-3p expression may
inhibit chemotherapy-induced autophagy and thereby provide a
mechanism through which to enhance the sensitivity of colon cancer
cells to chemotherapy.
Materials and methods
Cell culture, human tissue samples and
mice
The human 293 cells, the human colon cancer cell
lines (LoVo, HCT 116, DLD-1, SW480, HT-29 and RKO) and normal colon
epithelial cell lines (FHC and CCD-18Co) were obtained from the
Chinese Academy of Sciences (Shanghai, China) and cultured in
Dulbecco's modified Eagle's medium (DMEM) supplemented with 10%
fetal bovine serum (FBS) (both from Invitrogen, Carlsbad, CA, USA)
containing penicillin/streptomycin. The cells were grown in a
humidified 5% CO2 atmosphere at 37°C in an incubator.
The LoVo cell cultures were exposed to chemotherapy with
oxaliplatin as previously described (24), at the Cancer Research Institute of
Guangzhou Medical University (Guangzhou, China). The
oxaliplatin-resistant cell line was established from LoVo parental
cells by gradually treating the cells with increasing
concentrations of oxaliplatin for 3 months for the selection of
oxaliplatin-resistant cells. The cells were subcultured at 10%
confluency by splitting at a ratio of 1:200 and exposed to
increasing concentrations of oxaliplatin (0, 50, 100 and 200
µg/ml). The cultures that survived after 8 cycles of
subculturing and oxaliplatin treatment (maximum treatment, 200
µg/ml) were considered chemoresistant and used in the
subsequent experiments. It should be noted that the LoVo cells were
selected as the target cells for use in the experiments, as they
were more invasive than the other cells (data not shown). The
IC50 value was read from the drug concentration which
had the half maximal inhibitory effect on cell viability. In
addition, the accumulation of autophagosomes in the colon cancer
cells was measured using an electron microscope (Philips CM-120
transmission electron microscope; Philips, Amsterdam, The
Netherlands) as previously described (12). Autophagosomes appear in cells as
single membrane vesicles containing different cytoplasmatic
material.
A total of 30 human samples were used in this study,
including 20 human colon tumor tissue samples and 10 human normal
colon tissue (adjacent) samples. The samples were obtained from
patients undergoing surgery for colon cancer between 2013 and 2014
at the Department of Abdominal Surgery, The Affiliated Cancer
Hospital of Guangzhou Medical University (Guangzhou, China) and
stored in liquid nitrogen until analysis. The tumor and normal
samples were obtained following a protocol approved by the Ethics
Committee of the Department of Guangzhou Medical University, The
Affiliated Cancer Hospital (Guangzhou, China). All patients
provided written informed consent for the use of their samples for
experimental purposes.
Female BALB/c nude mice (n=20, 6 weeks old, weighing
25–30 g) were obtained from the Medical Experimental Animal Center
of Guangdong Province (Guangzhou, China) and housed under
pathogen-free conditions with free access to food and water. The
animal experimental procedures were reviewed and approved by the
Institutional Animal Care and Use Committee of Guangzhou Medical
University.
Determination of the interaction between
miR-409-3p and Beclin-1 3′-UTR using a Dual-Luciferase Reporter
assay
The wild-type (WT) 3′-UTR of human Beclin-1
(5′-AGGTTGAG AAAGGCGAGACA-3′) and the 3′-UTR sequences carrying
mutations (C5878T, A5999T or A6328G) were amplified and subcloned
into the pGL3 expression vector downstream of the promoter and
coding region of firefly luciferase (Promega Corp., Madison, WI,
USA); subcloning was performed using the XbaI and
NotI restriction sites. The resulting expression plasmids
were transfected into the chemoresistant LoVo cells in the presence
or absence of 20 nmol/ml miR-409-3p mimic (Gene Pharma, Shanghai,
China) using Lipofectamine transfection reagent (Invitrogen)
according to the manufacturer's instructions. The cells were
harvested 48 h after transfection and the luciferase activity was
measured using a Dual-Luciferase Reporter assay kit (Promega
Corp.). Luciferase activity was normalized to that measured in the
control cells transfected with the empty pGL3 expression vector in
the presence or absence of miR-409-3p mimic. In addition TargetScan
was used to confirm the target of miR-409-3p.
Cell viability and colony formation
assays
The chemoresistant LoVo cells were pre-treated with
miR-409-3p mimic or scrambled miRNA (5′-ATTAATCATAGAGGAAATCCACG-3′;
Shanghai Kangcheng Biological engineering Co. Ltd. Shanghai, China)
for 1 h and then exposed to oxaliplatin at 50 µg/ml for 24
h. Cell growth and viability were evaluated by
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay (Sangon Biotech, Shanghai, China). Briefly, the cells were
seeded in 96-well plates at a density of 5×103 cells/200
µl. Following transfection and chemotherapy, fresh medium
containing MTT solution [5 mg/ml diluted in phosphate-buffered
saline (PBS), 20 µl/well] was added followed by incubation
for an additional 4 h. The resulting formazan was resuspended in
dimethyl sulfoxide (200 µl/well). Finally, the absorbance
was determined at 490 nm using an ELISA reader (BioTek Instruments,
Inc., Winooski, VT, USA).
The colony-forming ability was assessed at 15 days
following transfection. The medium was discarded and the cell
colonies were stained with crystal violet (Sigma-Aldrich (Shanghai,
China) prior to counting using the Leica Zoom 2000 dissecting
microscope (Leica Microsystems Inc., Buffalo Grove, IL, USA).
Measurements were taken in quadruplicate from 3 independent
experiments.
miRNA or plasmid DNA transfection
miRNA mimics and control miRNAs were purchased from
Applied Biosystems Life Technologies (Foster City, CA, USA).
Lipofectamine 2000 (Invitrogen) was used for the transfection of
miRNA mimics or plasmid DNA according to the manufacturer's
instructions. At 48 h after transfection, the expression of
miR-409-3p was detected by reverse transcription
quantitative-polymerase chain reaction (RT-qPCR).
Cell viability assay
A total of 1×104 cells per well were
seeded in 48-well plates overnight. The medium was replaced with
fresh medium with or without oxaliplatin at the indicated
concentrations and incubated for 48 h. Cell viability was measured
by MTT assay. The absorbance was measured spectrophotometrically at
570 nm using the Universal Microplate Reader EL800 (BioTek
Instruments, Inc.).
RT-qPCR
Total RNA was isolated from the cells using TRIzol
reagent (Invitrogen) and small RNA was extracted using the
mirVana kit (Ambion, Austin, TX, USA) according to the
manufacturer's instructions. The corresponding cDNA was generated
using M-MLV reverse transcriptase (Clontech Laboratories, Palo
Alto, CA, USA) and the TaqMan microRNA reverse transcription kit
(Applied Biosystems, Foster City, CA, USA). The RT-qPCR mixtures,
which contained cDNA templates, primers and SYBR-Green qPCR Master
Mix, were subjected to RT-qPCR quantification, and Beclin-1 and
miR-409-3p expression levels were quantified using the
2−ΔΔCt method. The expression levels were expressed
relative to the levels of the following internal control genes:
glyceraldehyde- 3-phosphate dehydrogenase (GAPDH) for Beclin-1 or
U6 small nuclear (sn)RNA for miR-409-3p.
Western blot analysis
Total cell lysates were separated by 10% sodium
dodecyl sulfate-polyacrylamide gel electrophoresis and transferred
onto nitrocellulose membranes (Amersham; GE Healthcare, Little
Chalfont, UK). Non-fat dry milk (2.5%) was used to block the
membranes, which were then incubated with primary antibody against
LC3 (ab63817) and Beclin-1 (ab62557) (both from Abcam, Cambridge,
UK) or cleaved caspase-3 (PC679-50UG; Millipore Corp., Boston, MA,
USA) and anti-GAPDH antibody (bs-2188R; BIOSS, Beijing, China). The
membranes were then incubated with horseradish peroxide
(HRP)-conjugated antibody and antibody binding was visualized using
enhanced chemiluminescence (ECL; Amersham; GE Healthcare). Band
intensities were quantitated using Image Pro Plus 6.0 (Olympus,
Osaka, Japan) and normalized to the levels of GAPDH in the same
sample.
Tumorigenicity assay
Chemoresistant LoVo cells (2×106) diluted
in 200 µl PBS pre-transfected with lentiviral vector
expressing miR-409-3p or scrambled miRNA control were injected
subcutaneously into the right groin of the BALB/c nude mice. Tumor
length and width were measured daily and the volume was calculated
using the following formula: length × width2 × π/6. When
the tumor volume reached approximately 500 mm3, the
tumor was injected once daily with oxaliplatin (50 µg/ml, 1
ml) or PBS as a control and the volume was measured daily for at
least 36 more days. On day 41, the animals were euthanized by
cervical dislocation following exposure to CO2. The
tumors were removed and either frozen in liquid nitrogen or fixed
in 10% formalin for further analysis.
Statistical analysis
Data are expressed as the means ± standard deviation
(SD) of 3 independent experiments. Differences between or within
groups were assessed for significance using, respectively,
Student's two-tailed t-test or one-way ANOVA. The threshold for
significance was defined as P<0.05.
Results
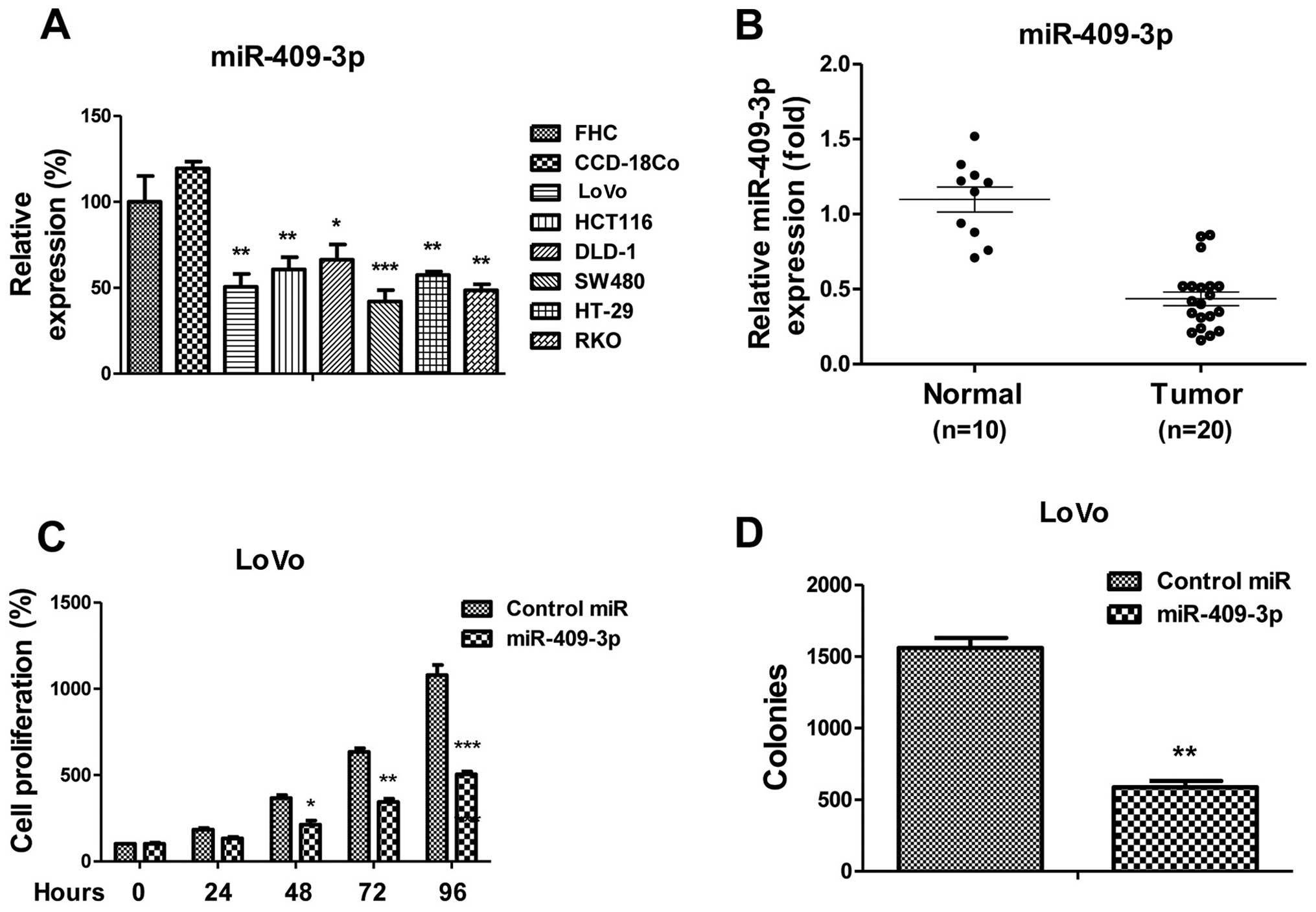
miR-409-3p is downregulated in human
colon tumors and suppresses colon cancer cell proliferation in
vitro
Previous studies have indicated that miR-409-3p acts
as a tumor suppressor in various types of cancer (21–23). To examine the role of miR-409-3p
in human colon cancer, we measured the expression of miR-409-3p in
colon cancer and normal colon cell lines. As expected, the
miR-409-3p levels were significantly downregulated in all the colon
cancer cell lines compared with the 2 normal cell lines, FHC and
CCD-18Co (Fig. 1A). To support
these findings, we compared the miR-409-3p levels between the human
colon cancer samples and the adjacent benign tissue samples.
Consistent with these results, miR-409-3p expression was
downregulated in the colon cancer samples, compared with the normal
colon tissue samples (Fig. 1B),
thus suggesting that miR-409-3p acts as a tumor suppressor in colon
cancer. We then transfected miR-409-3p mimics or control miRNA into
the LoVo colon cancer cells. Our results revealed that cell
proliferation was suppressed by the overexpression of miR-409-3p
(Fig. 1C). Cell proliferation at
48, 72 and 96 h was significantly suppressed following transfection
with miR-409-3p mimic. Moreover, we observed that miR-409-3p
overexpression in the LoVo cells significantly inhibited the
colony-forming ability of th cells (Fig. 1D). Taken together, these results
revealed a tumor suppressor role for of miR-409-3p in colon
cancer.
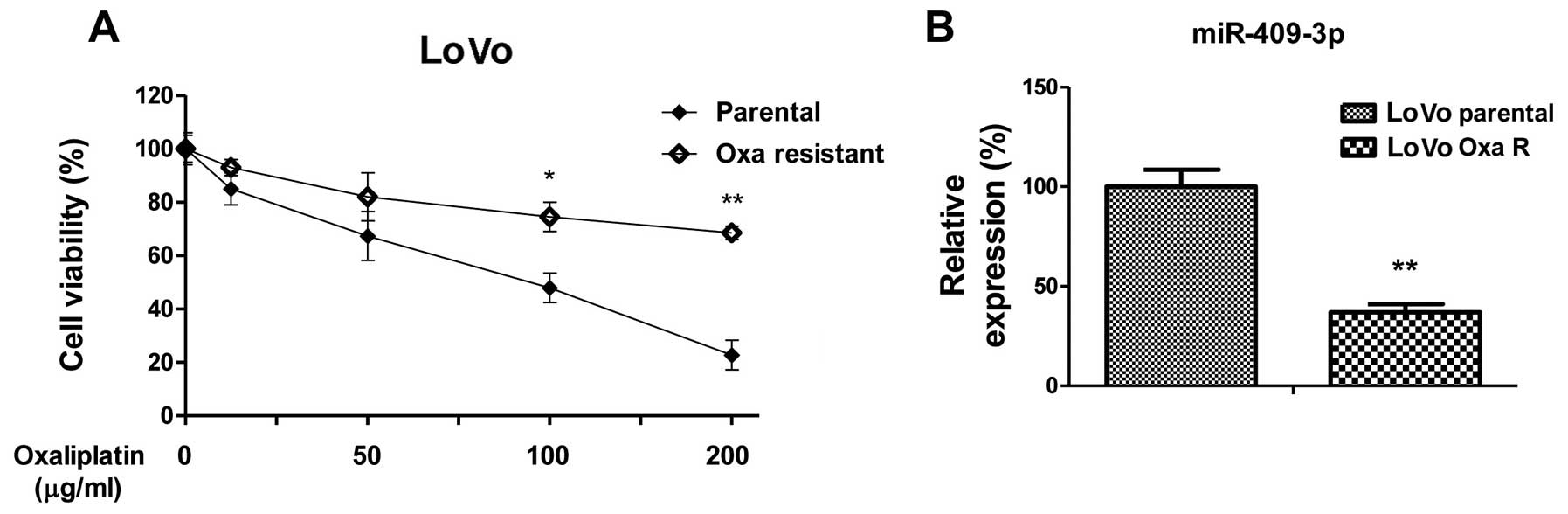
miR-409-3p epxression negatively
correlates with resistance to oxaliplatin in colon cancer
cells
The evasion of chemotherapeutic agents has been
recognized as one of the hallmarks of cancer (3). In this study, to investigate the
putative roles of miR-409-3p in enhancing the chemosensitivity of
colon cancer cells, we established the oxaliplatin-resistant cell
line from LoVo parental cells by gradually treating the parental
cells with increasing concentrations of oxaliplatin for 3 months to
select oxaliplatin-resistant cells. Fig. 2A illustrates that the
IC50 value of the oxaliplatin-resistant cells was
approximately 5-fold greater than the IC50 value of the
LoVo parental cells, indicating that the resistant cells may be
tolerant to higher concentrations of oxaliplatin. Of note,
miR-409-3p expression was significantly lower in the
oxaliplatin-resistant cultures than in the parental cells (Fig. 2B), indicating that miR-409-3p may
be a therapeutic target for the treatment of chemoresistant cancer
cells.
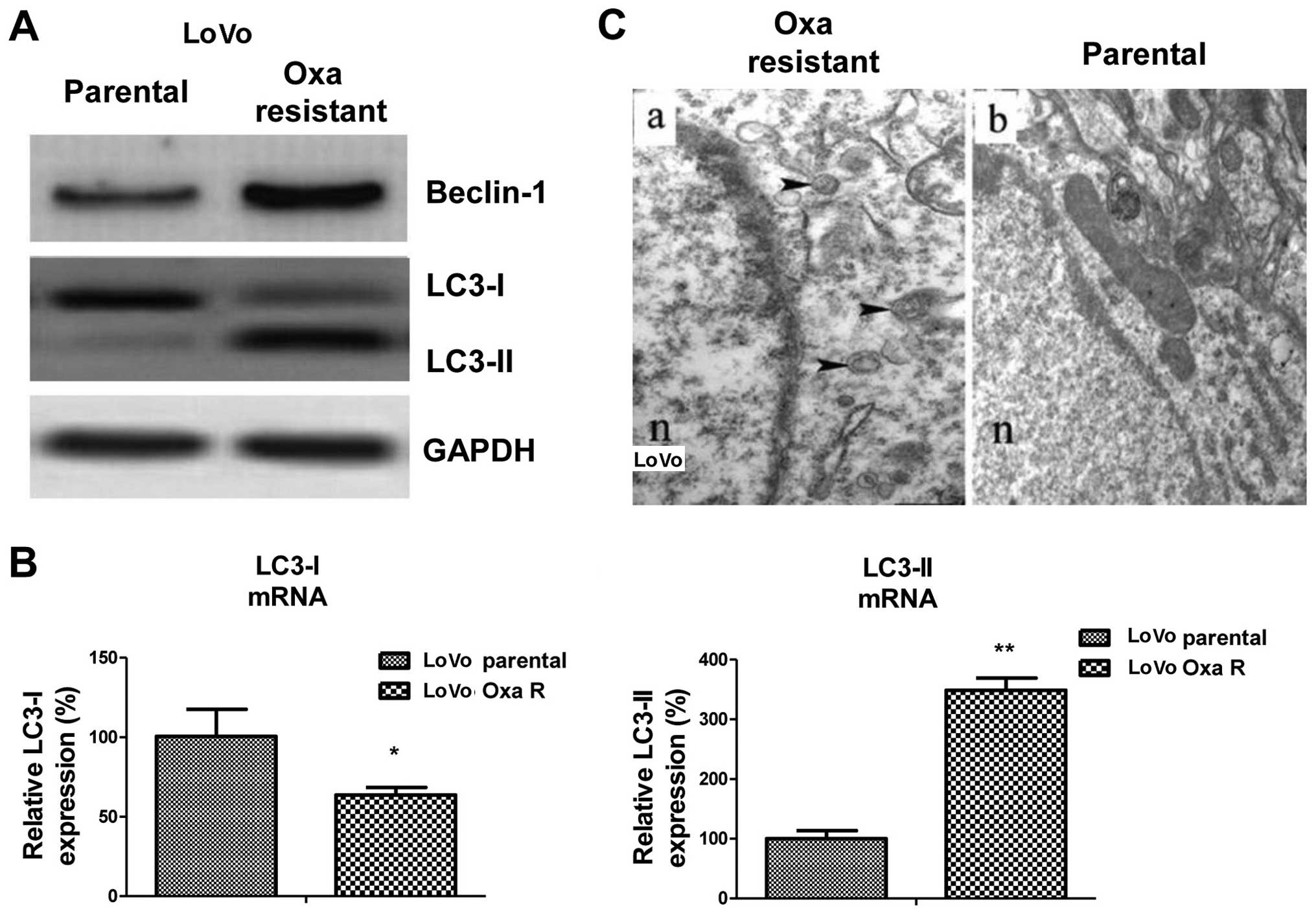
Oxaliplatin-resistant cancer cells
exhibit elevated autophagic activity
The above-mentioned findings demonstrated the
negative correlation between the expression of miR-409-3p and
sensitivity to oxaliplatin in colon cancer cells. To elucidate the
mechanisms responsible for the miR-409-3p-mediated
chemo-sensitivity, we compared the expression levels of Beclin-1
and its downstream factors, LC3-I and LC3-II between the parental
and oxaliplatin-resistant LoVo cells. As expected, western blot
analysis revealed that Beclin-1 and LC3-II protein expression was
present at much higher levels and that LC3-I protein expression was
downregulated in the chemoresistant cells compared with the
parental cells, indicating enhanced autophagic activity in the
oxaliplatin-resistant cells (Fig.
3A). Consistent with these findings, the mRNA expression levels
of LC3-I were downregulated and those of LC3-II were upregulated in
the chemoresistant cells (Fig.
3B). However, we did not observe a significant difference in
Beclin-1 mRNA expression levels between the resistant and sensitive
cells, indicating that the post-transcriptional regulation of
Beclin-1 may occur in oxaliplatin-resistant cells (data not shown).
To further confirm our findings, we measured autophagosome
formation in the chemoresistant (Fig.
3C, panel a) and parental (Fig.
3C, panel b) LoVo cancer cells. The LC3-II dots (representing
the autophagic activity; black arrowheads represent autophagosomes)
in the oxaliplatin-resistant cells were more evident than in the
sensitive cells (Fig. 3C). The
above-mentioned data (Figs. 2 and
3) indicate that miR-409-3p
expression is downregulated in chemotherapy-resistant colon cancer
cells and that autophagy is activated in these resistant cells,
thus suggesting a link between the downregulation of miR-409-3p
expression and the activation of autophagy.
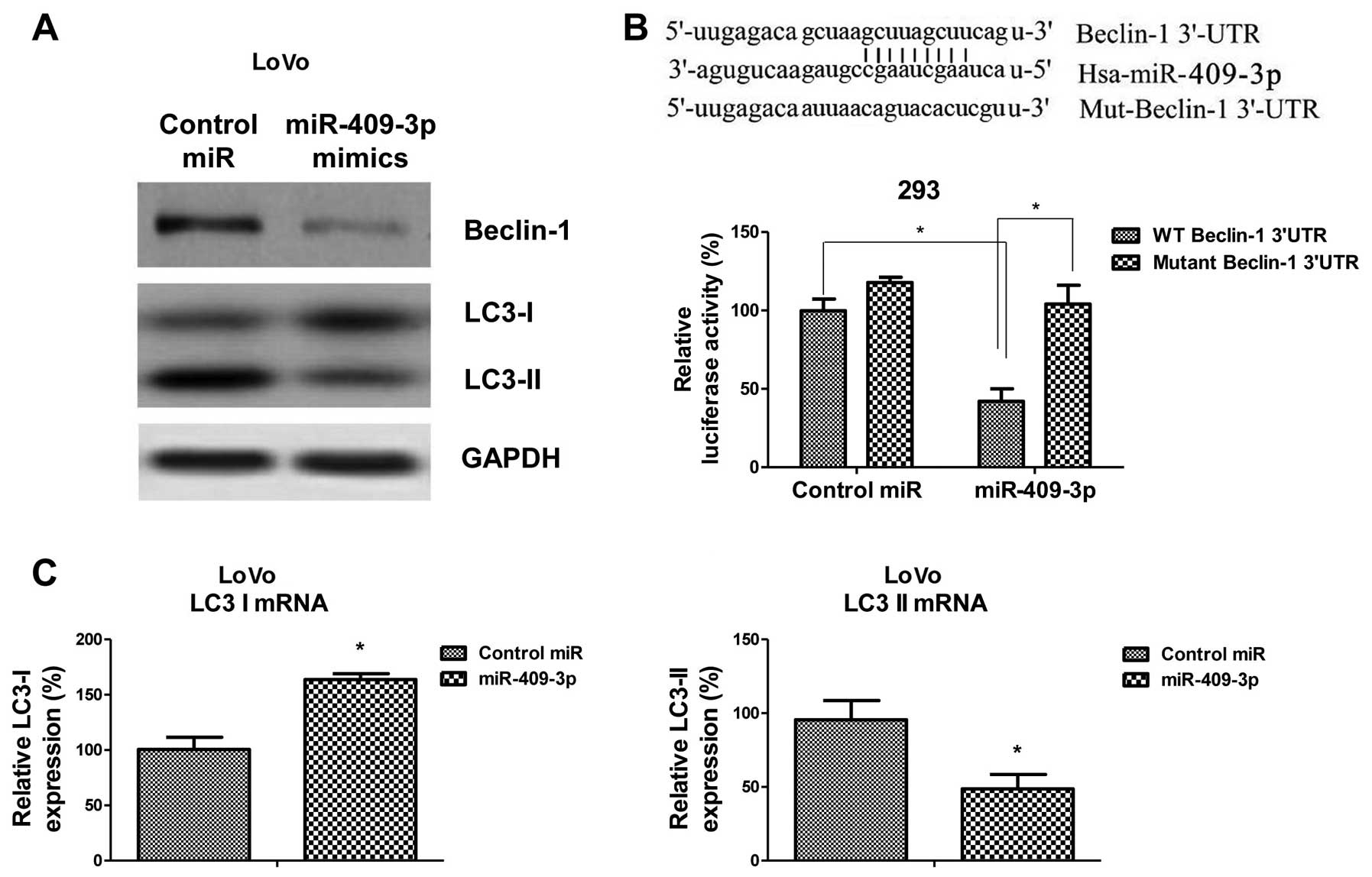
miR-409-3p directly targets Beclin-1 to
regulate autophagic activity
To obtain more direct evidence that miR-409-3p
regulates autophagy, we performed a screening of the possible
binding sites of miR-409-3p in the 3′-UTRs of gene transcripts
related to autophagy using TargetScan. This analysis highlighted
Beclin-1 as a putative target of miR-409-3p. To verify that
miR-409-3p targets Beclin-1, we measured the protein expression
levels of Beclin-1 and LC3-I/II in the LoVo cells in response to
miR-409-3p overexpression induced by transfection with miR-409-3p
mimics. In the cells transfected with miR-409-3p mimics, we
observed decreased Beclin-1 expression levels, as well as increased
LC3-I levels and decreased LC3-II levels (Fig. 4A). To verify that miR-409-3p binds
to the 3′-UTR of Beclin-1 directly, we transfected 293 cells with a
dual-luciferase reporter system in which the luciferase transcript
was fused to the 3′-UTR of Beclin-1. In this system, luciferase
activity is lower in the presence of elements that bind to the UTR.
The cells were transfected with either the reporter system alone or
with the reporter system together with the miR-409-3p mimic. Other
cells were co-transfected with a reporter system containing a
mutated version of the Beclin-1 3′-UTR lacking the predicted
miR-409-3p binding site. The relative luciferase activity was
significantly lower in the cultures co-transfected with miR-409-3p
mimic and pGL3 containing the WT 3′-UTR of Beclin-1 than in the
cells co-transfected with the mimic and pGL3 containing the mutant
3′-UTR (Fig. 4B). These results
confirmed that Beclin-1 is a target gene of miR-409-3p. Consistent
with the results from western blot analysis, the mRNA levels of
LC3-I and LC3-II were significantly altered by transfection with
miR-409-3p (Fig. 4C). Taken
together, the above-mentioned results demonstrate that miR-409-3p
suppresses autophagy through the direct inhibition of Beclin-1.
Exogenous overexpression of miR-409-3p
enhances the sensitivity of oxaliplatin-resistant and non-resistant
colon cancer cells through the inhibition of Beclin-1
To determine the effects of miR-409-3p on the drug
sensitivity of colon cancer cells, we pre-treated the LoVo cells
with miR-409-3p mimic or scrambled miRNA control and measured the
cell viabilitys in response to treatment with oxaliplatin.
Exogenous miR-409-3p overexpression led to significantly higher
apoptotic levels (Fig. 5A, shown
by a decrease in the number of surviving cells). These results
suggest that miR-409-3p enhances the sensitivity of LoVo colon
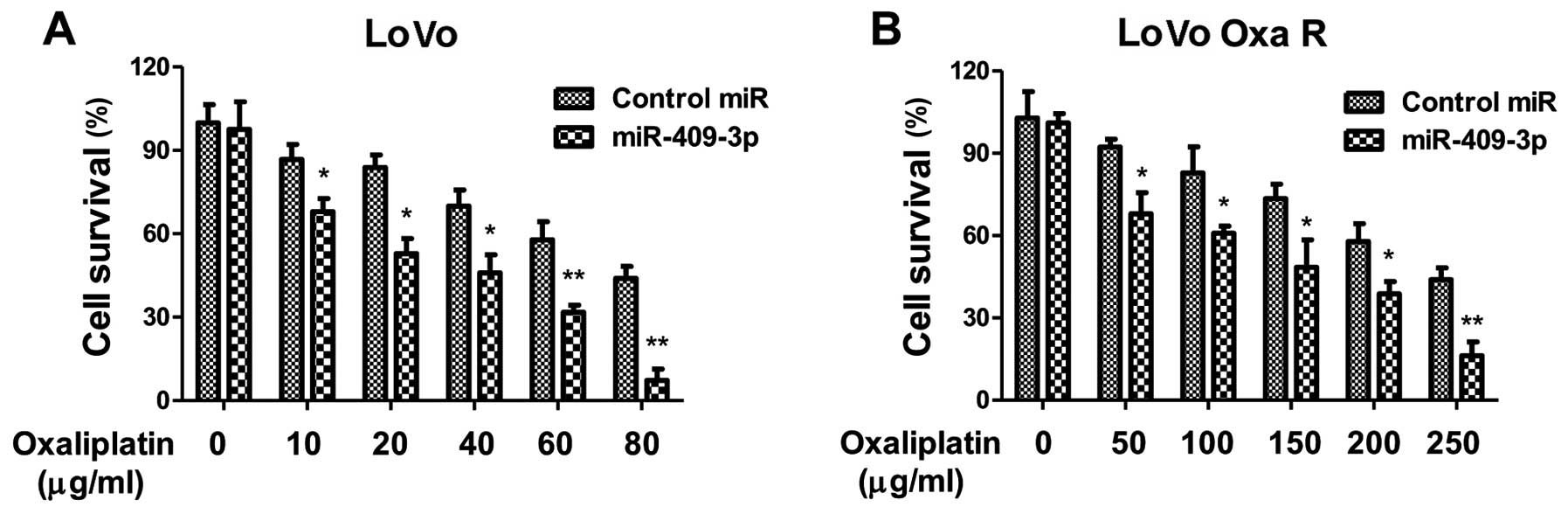
cance4r cells by triggering apoptosis. As expected, the
overexpression of miR-409-3p in LoVo oxaliplatin-resistant LoVo
cells sensitized the cells to oxaliplatin. The
oxaliplatin-resistant cells transfected with control miRNA showed
an IC50 value of 250 µg/ml, which was
significantly higher than that of the cells transfected with
miR-409-3p mimic (Fig. 5B),
indicating miR-409-3p may be a therapeutic agent which may be used
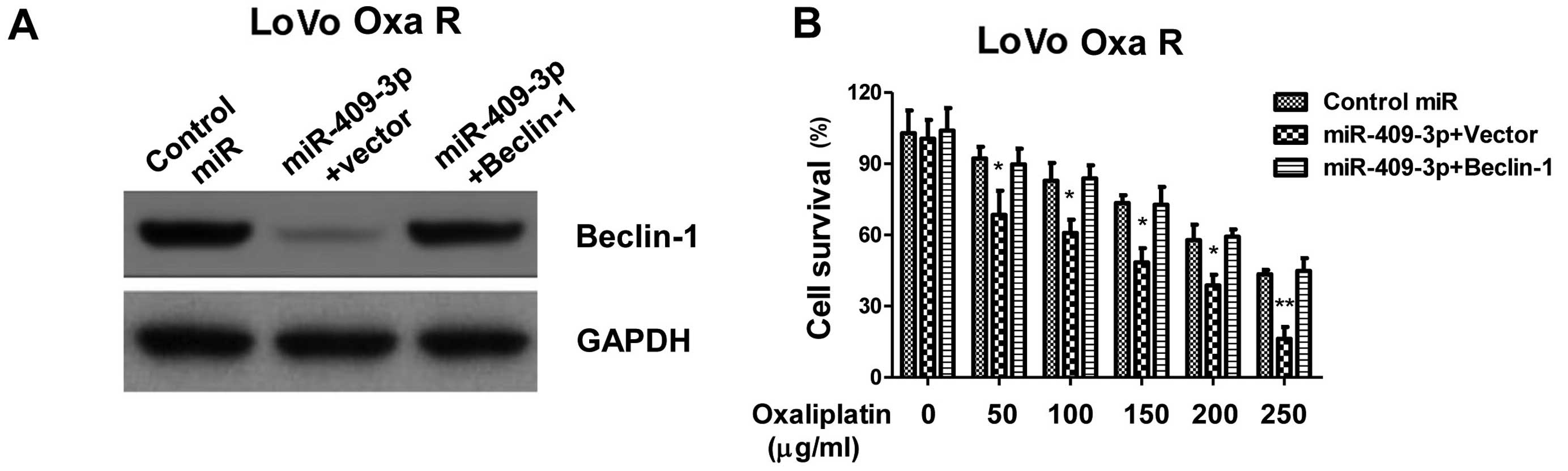
to overcome resistance to oxaliplatin. To obtain additional
evidence that miR-409-3p sensitizes chemoresistant LoVo cells to
chemotherapy by regulating Beclin-1 expression, we co-transfected
the cells with miR-409-3p mimic and an expression plasmid carrying
the Beclin-1 coding sequence upstream of a 3′-UTR mutated to
eliminate the miR-409-3p binding site. The overexpression of
Beclin-1 (Fig. 6A) almost
completely blocked the ability of miR-409-3p to inhibit the growth
of chemoresistant LoVo cells, as well as the ability of miR-409-3p
to trigger apoptosis (Fig. 6B).
These results support the hypothesis that miR-409-3p sensitizes
chemoresistant LoVo colon cancer cells to chemotherapy by targeting
Beclin-1-mediated autophagy.
Overexpression of miR-409-3p sensitizes
oxaliplatin-resistant colon cancer cells by suppressing
Beclin-1-mediated autophagy in vivo
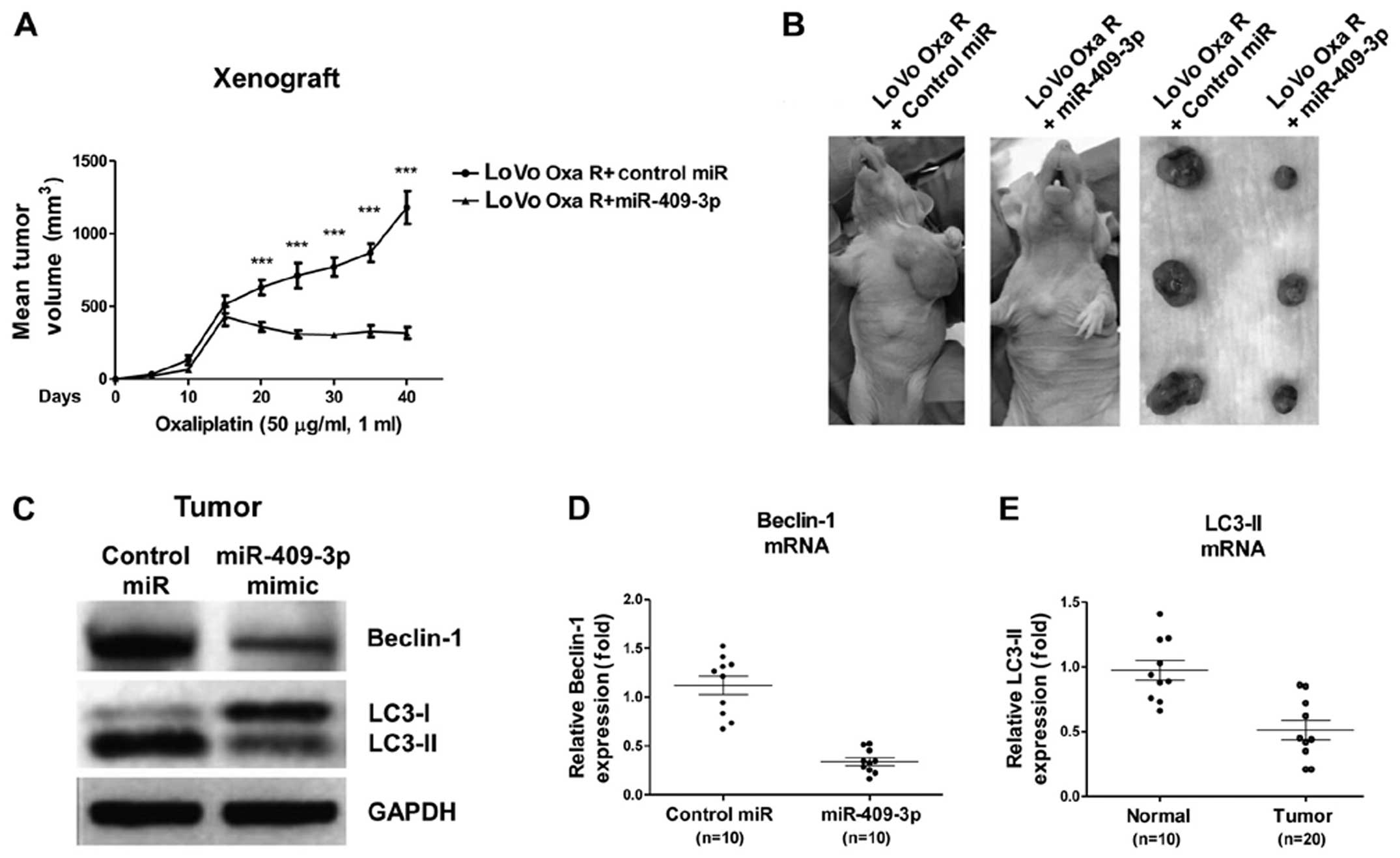
To verify our in vitro experiments using the
LoVo cell cultures, we determine whether miR-409-3p was capable of
strengthening the ability of chemotherapeutic drugs to suppress the
growth of tumor implanted tumors in nude mice. Twenty animals were
equally divided into 2 groups, and subcutaneously injected with
LoVo cells pre-transfected with lentiviral vector expressing
miR-409-3p or scrambled miRNA control. When the tumors attained a
volume of approximately 500 mm3 (around day 16), the
tumors were injected once daily with oxaliplatin (50 µg/ml,
1 ml). The tumors overexpressing miR-409-3p were significantly more
sensitive to chemotherapy than the tumors expressing the scrambled
control (Fig. 7A and B). In
addition, the analysis of the xenograft tumor samples at the end of
chemotherapy indicated that the tumors overexpressing miR-409-3p
expressed lower levels of Beclin-1 and exhibited less autophagic
activity (Fig. 7C–E). These
results are consistent with those from our in vitro
experiments, suggesting that miR-409-3p sensitizes LoVo cancer
cells to chemotherapy by blocking Beclin-1-mediated autophagy.
Discussion
In this study, using a combination of in
vitro experiments using chemoresistant LoVo human colon cancer
cells and in vivo experiments using a xenograft tumor model
in nude mice, we provide evidence that miR-409-3p inhibits
Beclin-1-mediated autophagy, thus rendering cancer cells sensitive
to oxaliplatin therapy. Our results are consistent with growing
evidence which suggests that autophagy plays a key role in the
chemoresistance of cancer cells (13,14), and our findings identify
miR-409-3p as having potential therapeutic potential. Our results
also support the hypothesis that miR-409-3p plays an important role
in the onset and/or progression of colon cancer.
The role of miR-409-3p in cancer is complex. In
prostate cancer, this miRNA appears to act as an oncogene,
inhibiting the expression of the tumor suppressor genes, Ras
suppressor 1 and stromal antigen 2 and thereby promoting
tumorigenesis (25). In laryngeal
cancer, miR-409-3p again appears to act as an oncogene,
specifically inhibiting Krüppel-like factor 17, E-cadherin and
phosphatidylinositol 3-kinase and thereby promoting the
epithelial-mesenchymal transition of tumor cells (26). Conversely, in gastric cancer,
miR-409-3p appears to act as a tumor suppressor (23). The levels of this miRNA have been
shown to be significantly lower in human colon cancer cells, than
in normal human colon cells (27). In fact, miR-409-3p has been
proposed to serve as a biomarker of gastric cancer (23). Moreover, miR-409-3p has been shown
to inhibit HT1080 cell proliferation, vascularization and
metastasis by targeting angiogenin (21). In the present study, we have
provide evidence that miR-409-3p suppresses colon cancer cell
proliferation, in part, by inhibiting autophagy mediated by
Beclin-1.
Studies on various types of cancer point to
autophagy as an important mechanism of cancer cell resistance to
treatment. Gamma radiation has been shown to induce autophagy that
renders glioma stem cells radioresistant (28,29). It has also been shown that
hypoxia-induced autophagy may contribute to the radioresistance of
breast cancer cells (30).
Hypoxia- inducible factor (HIF)1-α-mediated autophagy in cervical
cancer cells may contribute to paclitaxel chemoresistance (31), while autophagy in LoVo colon
cancer cells has been directly associated with resistance to
irinotecan (32). The inhibition
of autophagy in HCT 116 Bax-deficient colon cancer cells using the
drug 3-methyladenine has been shown to enhance purvalanol- induced
apoptotic cell death (33). The
overexpression of miR-502, which inhibits Rab1B-mediated autophagy,
has been demonstrated to inhibit the progression of HCT 116 and
SW480 colon cancer cells in vitro and the growth of a tumor
xenograft in vivo (34).
In this study, in continuation of these
above-mentioned findings, we demonstrated that in
oxaliplatin-resistant LoVo colon cancer cells, the miR-409-3p
levels were significantly downregulated and that autophagic
activity was increased. This suggests that miR-409-3p helps to
regulate autophagy. Consistent with this possibility, our
bioinformatics analysis predicted that the miRNA binds to the
3′-UTR of the key autophagy gene encoding Beclin-1, which we
confirmed in LoVo cells using a Dual-Luciferase Reporter assay. In
addition, the experiments in which LoVo cells were co-transfected
with the Beclin-1 expression plasmid and with a miR-409-3p mimic
revealed that the antagonistic effects of the two molecules were
lost when the putative miRNA binding sites in the 3′-UTR were
mutated. It appears that miR-409-3p does for oxaliplatin therapy
what miR-30a does for cis-dichloro-diamine platinum therapy:
both miRNAs inhibit Beclin-1-mediated autophagy, thereby
sensitizing cancer cells to chemotherapy (35). In lung cancer cells, the
downregulation of miR-17-5p expression has been shown to lead to
the upregulation of Beclin-1 expression and resistance to
paclitaxel (36).
Taken together, our findings and those of previous
studies highlight the possibility of targeting Beclin-1 as a
therapeutic strategy for cancer. In fact, the levels of this
protein have been shown to predict the overall survival of patients
with esophageal squamous cell carcinoma following radiation therapy
(37). However, it may not always
be desirable to reduce Beclin-1 expression and therefore autophagy.
While many studies point to autophagy as contributing to
chemoresistance, other studies have reported the opposite. Akt
inhibitors, for example, have been shown to promote
radiosensitivity by inducing autophagy (38). Inhibitors of the mammalian target
of rapamycin have been shown to radiosensitize non-small-cell lung
cancer cells by inhibiting the activity of phosphatase and tensin
homolog (PTEN) (39). Whether
these opposite findings regardin the association between autophagy
and resistance to treatment reflect differences between chemo- and
radiotherapy and/or between different types of cancers warrant
further investigation in future studies.
In conclusion, the present study provides evidence
that miR-409-3p targets and inhibits Beclin-1, which inhibits
chemotherapy-induced autophagy and thereby enhances the
chemosensitivity of colon cancer cells.
Abbreviations:
|
LC3
|
microtubule-associated protein light
chain 3
|
|
miRNA or miR
|
microRNA
|
|
MTT
|
3-(4,5-dimethylthiazol-2-yl)-2,5-
diphenyltetrazolium bromide
|
|
TUNEL
|
terminal deoxynucleotidyl transferase
dUTP nick-end labeling
|
|
UTR
|
untranslated region
|
Acknowledgments
The present study was supported by funding from the
Guangdong Province Natural Science Fund (no. S2013010016662), the
Health Bureau of Guangdong Province (nos. A2014224 and B2014196),
the Science and Technology Planning Project of Guangdong Province
(no. 2013B021800284) and the National Natural Science Foundation of
China (nos. 81201932 and 81372493).
References
|
1
|
Tárraga López PJ, Albero JS and
Rodríguez-Montes JA: Primary and secondary prevention of colorectal
cancer. Clin Med Insights Gastroenterol. 7:33–46. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bonetti A, Giuliani J and Muggia F:
Targeted agents and oxaliplatin-containing regimens for the
treatment of colon cancer. Anticancer Res. 34:423–434.
2014.PubMed/NCBI
|
|
3
|
Gordeev SS, Rasulov AO, Mamedli ZZ, Aliev
VA and Kulushev VM: Neoadjuvant chemotherapy for colorectal cancer
- time to learn a new treatment strategy? Vopr Onkol. 60:18–24.
2014.In Russian.
|
|
4
|
Walker AS, Zwintscher NP, Johnson EK,
Maykel JA, Stojadinovic A, Nissan A, Avital I, Brücher BL and
Steele SR: Future directions for monitoring treatment response in
colorectal cancer. J Cancer. 5:44–57. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen J, Wang W, Zhang Y, Chen Y and Hu T:
Predicting distant metastasis and chemoresistance using plasma
miRNAs. Med Oncol. 31:7992014. View Article : Google Scholar
|
|
6
|
Touil Y, Igoudjil W, Corvaisier M, Dessein
AF, Vandomme J, Monté D, Stechly L, Skrypek N, Langlois C, Grard G,
et al: Colon cancer cells escape 5FU chemotherapy-induced cell
death by entering stemness and quiescence associated with the
c-Yes/YAP axis. Clin Cancer Res. 20:837–846. 2014. View Article : Google Scholar
|
|
7
|
Herraez E, Gonzalez-Sanchez E, Vaquero J,
Romero MR, Serrano MA, Marin JJ and Briz O: Cisplatin-induced
chemoresistance in colon cancer cells involves FXR-dependent and
FXR-independent up-regulation of ABC proteins. Mol Pharm.
9:2565–2576. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhou Y, Sun K, Ma Y, Yang H, Zhang Y, Kong
X and Wei L: Autophagy inhibits chemotherapy-induced apoptosis
through downregulating Bad and Bim in hepatocellular carcinoma
cells. Sci Rep. 4:53822014.PubMed/NCBI
|
|
9
|
Helgason GV, Holyoake TL and Ryan KM: Role
of autophagy in cancer prevention, development and therapy. Essays
Biochem. 55:133–151. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Maiuri MC, Zalckvar E, Kimchi A and
Kroemer G: Self-eating and self-killing: crosstalk between
autophagy and apoptosis. Nat Rev Mol Cell Biol. 8:741–752. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mizushima N, Ohsumi Y and Yoshimori T:
Autophagosome formation in mammalian cells. Cell Struct Funct.
27:421–429. 2002. View Article : Google Scholar
|
|
12
|
Song J, Guo X, Xie X, Zhao X, Li D, Deng
W, Song Y, Shen F, Wu M and Wei L: Autophagy in hypoxia protects
cancer cells against apoptosis induced by nutrient deprivation
through a Beclin1-dependent way in hepatocellular carcinoma. J Cell
Biochem. 112:3406–3420. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Song J, Qu Z, Guo X, Zhao Q, Zhao X, Gao
L, Sun K, Shen F, Wu M and Wei L: Hypoxia-induced autophagy
contributes to the chemoresistance of hepatocellular carcinoma
cells. Autophagy. 5:1131–1144. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kang R, Wang ZH, Wang BQ, Zhang CM, Gao W,
Feng Y, Bai T, Zhang HL, Huang-Pu H and Wen SX: Inhibition of
autophagy- potentiated chemosensitivity to cisplatin in laryngeal
cancer Hep-2 cells. Am J Otolaryngol. 33:678–684. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mendell JT and Olson EN: MicroRNAs in
stress signaling and human disease. Cell. 148:1172–1187. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bartel DP: MicroRNAs: genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Oh JS, Kim JJ, Byun JY and Kim IA:
Lin28-let7 modulates radiosensitivity of human cancer cells with
activation of K-Ras. Int J Radiat Oncol Biol Phys. 76:5–8. 2010.
View Article : Google Scholar
|
|
18
|
Wang P, Chen L, Zhang J, Chen H, Fan J,
Wang K, Luo J, Chen Z, Meng Z and Liu L: Methylation-mediated
silencing of the miR-124 genes facilitates pancreatic cancer
progression and metastasis by targeting Rac1. Oncogene. 33:514–524.
2014. View Article : Google Scholar
|
|
19
|
Wang P, Zhuang L, Zhang J, Fan J, Luo J,
Chen H, Wang K, Liu L, Chen Z and Meng Z: The serum miR-21 level
serves as a predictor for the chemosensitivity of advanced
pancreatic cancer, and miR-21 expression confers chemoresistance by
targeting FasL. Mol Oncol. 7:334–345. 2013. View Article : Google Scholar
|
|
20
|
Winter J, Jung S, Keller S, Gregory RI and
Diederichs S: Many roads to maturity: microRNA biogenesis pathways
and their regulation. Nat Cell Biol. 11:228–234. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Weng C, Dong H, Chen G, Zhai Y, Bai R, Hu
H, Lu L and Xu Z: miR-409–3p inhibits HT1080 cell proliferation,
vascularization and metastasis by targeting angiogenin. Cancer
Lett. 323:171–179. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu X, Chen H, Lin Y, Hu Z, Mao Y, Wu J, Xu
X, Zhu Y, Li S, Zheng X and Xie L: MicroRNA-409–3p inhibits
migration and invasion of bladder cancer cells via targeting c-Met.
Mol Cells. 36:62–68. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zheng B, Liang L, Huang S, Zha R, Liu L,
Jia D, Tian Q, Wang Q, Wang C, Long Z, et al: MicroRNA-409
suppresses tumour cell invasion and metastasis by directly
targeting radixin in gastric cancers. Oncogene. 31:4509–4516. 2012.
View Article : Google Scholar
|
|
24
|
Zhang XL, Hu AB, Cui SZ and Wei HB:
Thermotherapy enhances oxaliplatin-induced cytotoxicity in human
colon carcinoma cells. World J Gastroenterol. 18:646–653. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Josson S, Gururajan M, Sung SY, Hu P, Shao
C, Zhau HE, Liu C, Lichterman J, Duan P, Li Q, et al: Stromal
fibroblast-derived miR-409 promotes epithelial-to-mesenchymal
transition and prostate tumorigenesis. Oncogene. 34:2690–2699.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tai J, Xiao X, Huang ZG, Yu ZK, Chen XH,
Zhou WG, Chen XJ, Rao YS, Fang JG and Ni X: MicroRNAs regulate
epithelial- mesenchymal transition of supraglottic laryngeal
cancer. Zhonghua Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 48:499–503.
2013.In Chinese. PubMed/NCBI
|
|
27
|
Liu M, Xu A, Yuan X, Zhang Q, Fang T, Wang
W and Li C: Downregulation of microRNA-409–3p promotes
aggressiveness and metastasis in colorectal cancer: an indication
for personal-ized medicine. J Transl Med. 13:1952015. View Article : Google Scholar
|
|
28
|
Lomonaco SL, Finniss S, Xiang C,
Decarvalho A, Umansky F, Kalkanis SN, Mikkelsen T and Brodie C: The
induction of autophagy by γ-radiation contributes to the
radioresistance of glioma stem cells. Int J Cancer. 125:717–722.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhuang W, Qin Z and Liang Z: The role of
autophagy in sensi-tizing malignant glioma cells to radiation
therapy. Acta Biochim Biophys Sin (Shanghai). 41:341–351. 2009.
View Article : Google Scholar
|
|
30
|
He WS, Dai XF, Jin M, Liu CW and Rent JH:
Hypoxia-induced autophagy confers resistance of breast cancer cells
to ionizing radiation. Oncol Res. 20:251–258. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Peng X, Gong F, Chen Y, Jiang Y, Liu J, Yu
M, Zhang S, Wang M, Xiao G and Liao H: Autophagy promotes
paclitaxel resistance of cervical cancer cells: involvement of
Warburg effect activated hypoxia-induced factor 1-α-mediated
signaling. Cell Death Dis. 5:e13672014. View Article : Google Scholar
|
|
32
|
Chen MC, Lee NH, Ho TJ, Hsu HH, Kuo CH,
Kuo WW, Lin YM, Tsai FJ, Tsai CH and Huang CY: Resistance to
irinotecan (CPT-11) activates epidermal growth factor
receptor/nuclear factor kappa B and increases cellular metastasis
and autophagy in LoVo colon cancer cells. Cancer Lett. 349:51–60.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Coker-Gurkan A, Arisan ED, Obakan P,
Guvenir E and Unsal NP: Inhibition of autophagy by 3-MA potentiates
purvalanol-induced apoptosis in Bax deficient HCT 116 colon cancer
cells. Exp Cell Res. 328:87–98. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhai H, Song B, Xu X, Zhu W and Ju J:
Inhibition of autophagy and tumor growth in colon cancer by
miR-502. Oncogene. 32:1570–1579. 2013. View Article : Google Scholar
|
|
35
|
Zou Z, Wu L, Ding H, Wang Y, Zhang Y, Chen
X, Chen X, Zhang CY, Zhang Q and Zen K: MicroRNA-30a sensitizes
tumor cells to cis-platinum via suppressing beclin 1-mediated
autophagy. J Biol Chem. 287:4148–4156. 2012. View Article : Google Scholar :
|
|
36
|
Chatterjee A, Chattopadhyay D and
Chakrabarti G: miR-17–5p downregulation contributes to paclitaxel
resistance of lung cancer cells through altering beclin1
expression. PLoS One. 9:e957162014. View Article : Google Scholar
|
|
37
|
Chen Y, Li X, Wu X, He C, Guo L, Zhang S,
Xiao Y, Guo W and Tan B: Autophagy-related proteins LC3 and
Beclin-1 impact the efficacy of chemoradiation on esophageal
squamous cell carcinoma. Pathol Res Pract. 209:562–567. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Fujiwara K, Iwado E, Mills GB, Sawaya R,
Kondo S and Kondo Y: Akt inhibitor shows anticancer and
radiosensitizing effects in malignant glioma cells by inducing
autophagy. Int J Oncol. 31:753–760. 2007.PubMed/NCBI
|
|
39
|
Kim EJ, Jeong JH, Bae S, Kang S, Kim CH
and Lim YB: mTOR inhibitors radiosensitize PTEN-deficient
non-small-cell lung cancer cells harboring an EGFR activating
mutation by inducing autophagy. J Cell Biochem. 114:1248–1256.
2013. View Article : Google Scholar : PubMed/NCBI
|