Introduction
Oral squamous cell carcinoma (OSCC) accounts for
approximately 90% of all malignant oral tumors (1). OSCC often causes issues with
chewing, swallowing and speaking and affects the physical
appearance of patients. Although surgery, chemotherapy and
radiotherapy cure the majority of cases of early stage OSCC, the
outcome of most patients with advanced stage OSCC has not improved
over the last three decades (2).
The development of OSCC is a multi-step process involving the
progressive acquisition and study of genetic and epigenetic
alterations, such as deletions, point mutations and DNA methylation
(3). Studying the key molecules
that play important roles in the progression of OSCC will yield
novel insights and assist in the search for new therapeutic
targets.
MicroRNAs (miRNAs or miRs) are small non-coding RNAs
that regulate mRNA transcription and translation in various
cellular activities such as proliferation and differentiation
(4,5). Previous research has demonstrated
that aberrant miRNA expression is related to the development of
several types of human cancer (6). MiR-155 maps within and is processed
from an exon of a non-coding RNA transcribed from the B-cell
Integration Cluster (BIC) located on chromosome 21 (7). Previous studies have reported that
miR-155 plays an important role in many solid tumors, including
laryngeal cell carcinoma, lung cancer, gliomas, colon cancer,
hepatocellular carcinoma, pancreatic and breast cancer (8–15).
In relation to laryngeal squamous cell carcinoma, it was noted that
miR-155 increased cancer cell proliferation, migration and invasion
through suppressor of cytokine signaling 1 (SOCS1) and signal
transducer and activator of transcription 3 (STAT3) (9). In relation to colon cancer, Zhang
et al (15) reported that
the upregulation of miR-155 promotes tumor cell migration and
invasion via claudin-1. Recently, a study revealed that miR-155
overexpression in OSCC promoted cell proliferation through cell
division cycle 73 (CDC73) (16).
However, it is not yet known whether miR-155 affects other aspects
of the biological behavior of OSCC cells through other downstream
target genes.
B-cell CLL/lymphoma 6 (BCL6) is a transcriptional
repressor that has been proven to be a direct target gene of
miR-155 in macrophages (17).
BCL6 regulates a series of molecules that participate in regulating
the cell cycle and inflammation, including cyclin D2 and various
chemokines (18). Cyclin D2 plays
an important role in the cell cycle, and the ectopic expression of
cyclin D2 is closely related to invasion in OSCC cells (19). Moreover, BCL6 directly regulates
cyclin D2 by binding with its promoter (20). Previous studies have revealed that
the BCL6/cyclin D2 axis is involved in many disorders and that
pro-heparin binding (HB)-epidermal growth factor (EGF) and forkhead
box O3A (FOXO3A) regulate this axis (20,21). However, to the best of our
knowledge, no previous study has discussed the pathological role of
miR-155 in the BCL6/cyclin D2 axis in OSCC.
In the present study, we found that miR-155 was
upregulated in the OSCC CAL27 cell line and in OSCC specimens. The
overexpressed miR-155 enhanced the proliferative, migratory and
invasive ability of CAL27 cells by decreasing BCL6 expression and
increasing cyclin D2 levels. These results all suggest that miR-155
plays a tumor-promoting role during OSCC development, partly
through the BCL6/cyclin D2 axis.
Materials and methods
Cell culture
The human OSCC cell line CAL27 was purchased from
the American Type Culture Collection (ATCC; Manassas, VA, USA), and
cultured in Dulbecco's modified Eagle's medium high-glucose medium
(Life Technologies, Grand Island, NY, USA) supplemented with
heat-inactivated 10% fetal bovine serum (HyClone Laboratories,
Logan, UT, USA). The normal oral keratinocyte NOK-SI cell line was
kindly provided by the National Institute of Health (Bethesda, MD,
USA) and was grown in keratinocyte serum-free medium containing
human recombinant epidermal growth factor and bovine pituitary
extract (Life Technologies). Cells were cultured at 37°C in a
humidified atmosphere of 5% CO2 and 95% air.
Human tissue specimens
Primary OSCC tissues and matching adjacent non-tumor
tissues were obtained from 12 patients who underwent OSCC resection
without receiving preoperative chemotherapy or radiation at the
Guanghua School of Stomatology, Hospital of Stomatology, Sun
Yat-sen University (Guangzhou, China) between October 2011 and
November 2012 (Table I). We
obtained informed consent from all the patients. All tissue
specimens were immediately frozen in liquid nitrogen and then
stored at −80°C until the RNA was extracted. The samples were
stained with hematoxylin and eosin and examined histopathologically
(Table I). The Ethics Committee
of Sun Yat-sen University approved the present study.
 | Table IClinical data of patients with
OSCC. |
Table I
Clinical data of patients with
OSCC.
| Case no. | Gender | Age (years) | Tumor location | Pathological
diagnosis |
|---|
| Patient 1 | Female | 44 | Tongue | Well
differentiated |
| Patient 2 | Female | 68 | Gingiva | Well
differentiated |
| Patient 3 | Female | 32 | Tongue | Well
differentiated |
| Patient 4 | Female | 33 | Tongue | Well
differentiated |
| Patient 5 | Male | 49 | Floor of the
mouth | Poorly
differentiated |
| Patient 6 | Male | 80 | Tongue | Poorly
differentiated |
| Patient 7 | Female | 68 | Tongue | Well
differentiated |
| Patient 8 | Male | 62 | Oropharynx | Moderately
differentiated |
| Patient 9 | Male | 46 | Tongue | Moderately
differentiated |
| Patient 10 | Male | 57 | Tongue | Poorly
differentiated |
| Patient 11 | Female | 52 | Tongue | Moderately
differentiated |
| Patient 12 | Male | 63 | Tongue | Poorly
differentiated |
Reverse transcription quantitative PCR
(RT-qPCR)
In order to quantify miR-155, BCL6 and cyclin
D2 (CCND2) mRNA expression using RT-qPCR, total RNA was
extracted from cultured cells and clinical specimens using TRIzol
(Life Technologies). miR-155 and its control U6 were stem-loop
reverse transcribed using specific Bulge-Loop miRNA qRT-PCR Primer
sets (Guangzhou RiboBio Co., Ltd., Guangzhou, China) and also a
PrimeScript RT reagent kit (Takara, Shiga, Japan) in accordance
with the manufacturers' instructions. BCL6, cyclin D2 and
the control β-actin were reverse transcribed using a
Transcriptor First Strand cDNA Synthesis kit (Roche, Mannheim,
Germany). Subsequently, qPCR of miR-155 and U6 was performed using
a LightCycler 480 Real-Time PCR system (Roche) and SYBR Premix Ex
Taq II™ (Takara). The amplification profile involved initial
denaturation at 95°C for 30 sec, followed by 40 cycles of
denaturation at 95°C for 5 sec, annealing at 60°C for 20 sec and a
final extension at 65°C for 15 sec. qPCR of BCL6, cyclin D2
and β-actin was performed using the LightCycler 480
Real-Time PCR system and LightCycler 480 SYBR-Green I Master Mix
(Roche) with specific primers (Life Technologies) (Table II). The amplification profile
involved initial denaturation at 95°C for 5 min, followed by 40
cycles of denaturation at 95°C for 10 sec, annealing at 60°C for 20
sec and a final extension at 72°C for 20 sec. miR-155, BCL6
and cyclin D2 expression relative to U6 or β-actin
was determined as the respective comparative threshold cycle values
(2−ΔΔCt).
| Table IIPrimers used for RT-qPCR. |
Table II
Primers used for RT-qPCR.
| Gene | Primer | 5′→3′ |
|---|
| BCL6 | Forward |
ACACATCTCGGCTCAATTTGC |
| Reverse |
AGTGTCCACAACATGCTCCAT |
| Cyclin
D2 | Forward |
TTTGCCATGTACCCACCGTC |
| Reverse |
AGGGCATCACAAGTGAGCG |
| β-actin | Forward |
CTCTGGCCGTACCACTGGC |
| Reverse |
GTGAAGCTGTAGCCGCGC |
miRNA transfection
Homo sapiens (hsa)-miR-155-5p mimic (miR-155
mimic), miRNA mimic negative control (mimic-NC) (both from Life
Technologies), hsa-miR-155-5p antagomir (miR-155 antagomir), and
miRNA antagomir negative control (antagomir-NC) (both from
Guangzhou RiboBio) were used for transfection. Cells were incubated
in plates 24 h before transfection. When cells reached 30–40%
confluence, 5 nM miR-155 mimic, 5 nM mimic-NC, 100 nM miR-155
antagomir, or 100 nM antagomir-NC mixed with Lipofectamine RNAiMAX
transfection reagent according to the manufacturer's instructions
(Life Technologies) was added. Six hours after transfection, the
culture medium was replaced with fresh medium. We also constructed
a mock group, in which we added Lipofectamine RNAiMAX and culture
medium to cells but no synthetic miRNA sequence.
Cell proliferation assay
CAL27 cells were seeded in 96-well plates at 3,500
cells/well and cultured at 37°C. Twenty-four hours after seeding,
the cells were transfected with the synthetic miRNA sequences.
Every 24 h after transfection, 10 μl Cell Counting Kit-8
solution (Dojindo Laboratories, Kumamoto, Japan) was added to each
well, which contained 100 μl culture medium, and the plates
were further incubated for 2 h at 37°C. Absorbance was measured
using an automated microplate reader (Thermo Fisher Scientific,
Waltham, MA, USA) at 450 nm.
Migration and invasion assay
The cell migration and invasion assays were
performed in cell culture inserts packaged in companion plates
(Becton-Dickinson, Franklin Lakes, NJ, USA) with 8-μm
polycarbonate nuclepore filters. Transfected CAL27 cells were
plated at 7×104 cells/insert suspended in 100 μl
serum-free culture medium. For the invasion assay, each insert was
coated with Matrigel matrix (BD) before the cells were seeded.
After 24 h incubation, cells on the apical side of the inserts were
removed with cotton swabs, and the inserts were then fixed with 4%
paraformaldehyde for 30 min and stained with 0.4% crystal violet
for 15 min. The number of migrated cells in 10 random visual fields
was directly counted under a light microscope (Zeiss, Jena,
Germany). The cells were also counted indirectly by eluting the
membrane-binding crystal violet with 500 μl 10% glacial
acetic acid, and then measuring absorbance at 590 nm using an
automated microplate reader (Thermo Fisher Scientific).
Western blot analysis
To isolate the proteins, cells were washed twice
with cold phosphate-buffered saline and then lysed with RIPA and
PSFM (Beyotime Institute of Biotechnology, Nanjing, China). Protein
concentrations were determined using a BCA protein assay kit
(Nanjing KeyGen Biotech, Co., Ltd., Nanjing, China). The proteins
were separated using 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis, and were transferred onto polyvinylidene fluoride
membranes (Millipore, Billerica, MA, USA). After blocking in 5%
milk, the membranes were incubated with rabbit monoclonal anti-BCL6
antibody (D65C10, diluted 1:1,000), rabbit monoclonal anti-cyclin
D2 antibody (D52F9, diluted 1:1,000) (both from Cell Signaling
Technology, Boston, MA, USA), mouse monoclonal anti-GAPDH antibody
(ab125247, diluted 1:2,000), and rabbit polyclonal anti-H2A.X
antibody (ab10475, diluted 1:500) (both from Abcam, Cambridge, MA,
USA) at 4°C overnight. The membranes were then incubated with the
secondary antibodies goat polyclonal anti-rabbit IgG (ab6721,
diluted 1:5,000) and goat polyclonal anti-mouse IgG (ab85760,
diluted 1:2,000) (both from Abcam) for 2 h. The bands were then
visualized using an enhanced chemiluminescence detection system
(Alpha Innotech Cοrp., San Leandro, CA, USA) and quantified using
ImageJ software.
Statistical analysis
All statistical analyses were carried out using SPSS
version 16.0 (SPSS, Inc., Chicago, IL, USA). Data are expressed as
the means ± standard deviation. Statistical analysis was performed
using Student's t-test, or one-way ANOVA or the Kruskal-Wallis
test. A P-value <0.05 was considered to indicate a statistically
significant difference.
Results
miR-155 is highly expressed in the OSCC
cell line and OSCC tissue samples
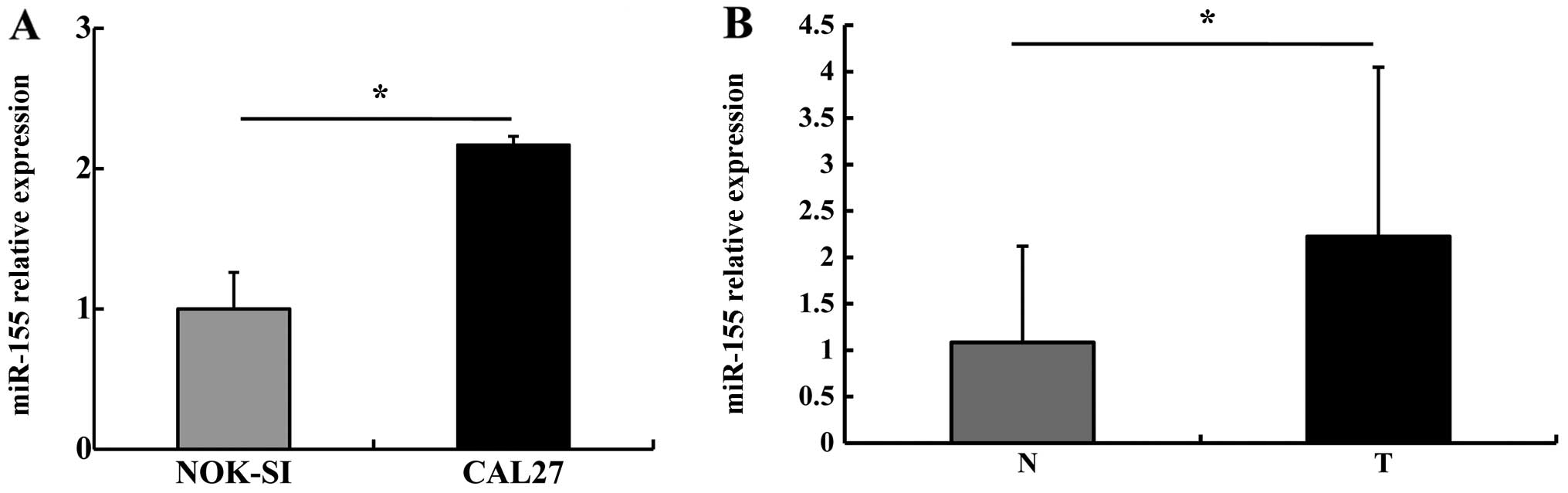
We first detected miR-155 expression in NOK-SI and
CAL27 cells using stem-loop RT-qPCR. miR-155 expression in the
CAL27 cells was 2.17±0.26 times higher than that in the NOK-SI
cells (P<0.05) (Fig. 1A). We
then detected miR-155 levels in the OSCC specimens: miR-155
expression in the OSCC tumor tissues was higher than that in the
paired adjacent non-tumor tissues (P<0.05) (Fig. 1B). These results suggest that
miR-155 is typically upregulated in OSCC.
miR-155 promotes CAL27 cell
proliferation
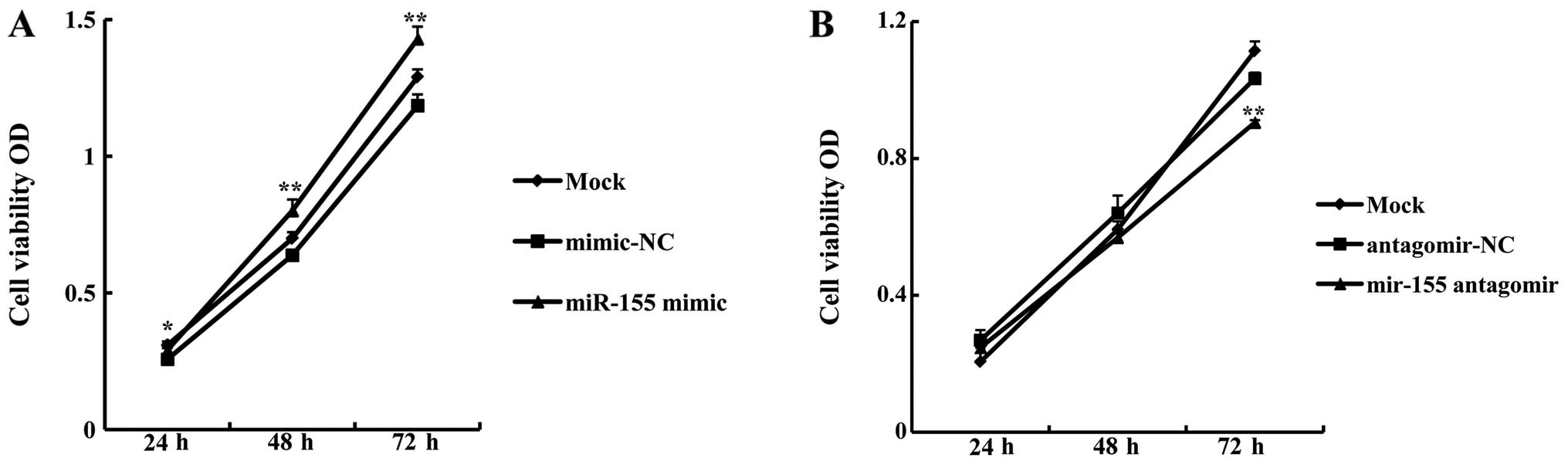
In order to examine whether miR-155 affected OSCC
cell growth, a proliferation assay was carried out using CAL27
cells following transfection with miR-155 mimic or antagomir. The
growth rate of CAL27 cells transfected with 5 nM miR-155 mimic was
significantly higher than that of cells transfected with mimic-NC
(miR-155 mimic OD/mimic-NC OD) at 24 h (113.14±2.75%, P<0.05),
48 h (125.39±6.49%, P<0.01), and 72 h (120.6±0.41%, P<0.01)
(Fig. 2A). By contrast, the
growth rate of cells transfected with 100 nM miR-155 antagomir was
significantly lower compared with antagomir-NC at 72 h
(87.61±1.67%, P<0.01) (Fig.
2B). These data indicate that miR-155 promotes OSCC cell
proliferation.
miR-155 increases CAL27 cell migration
and invasion
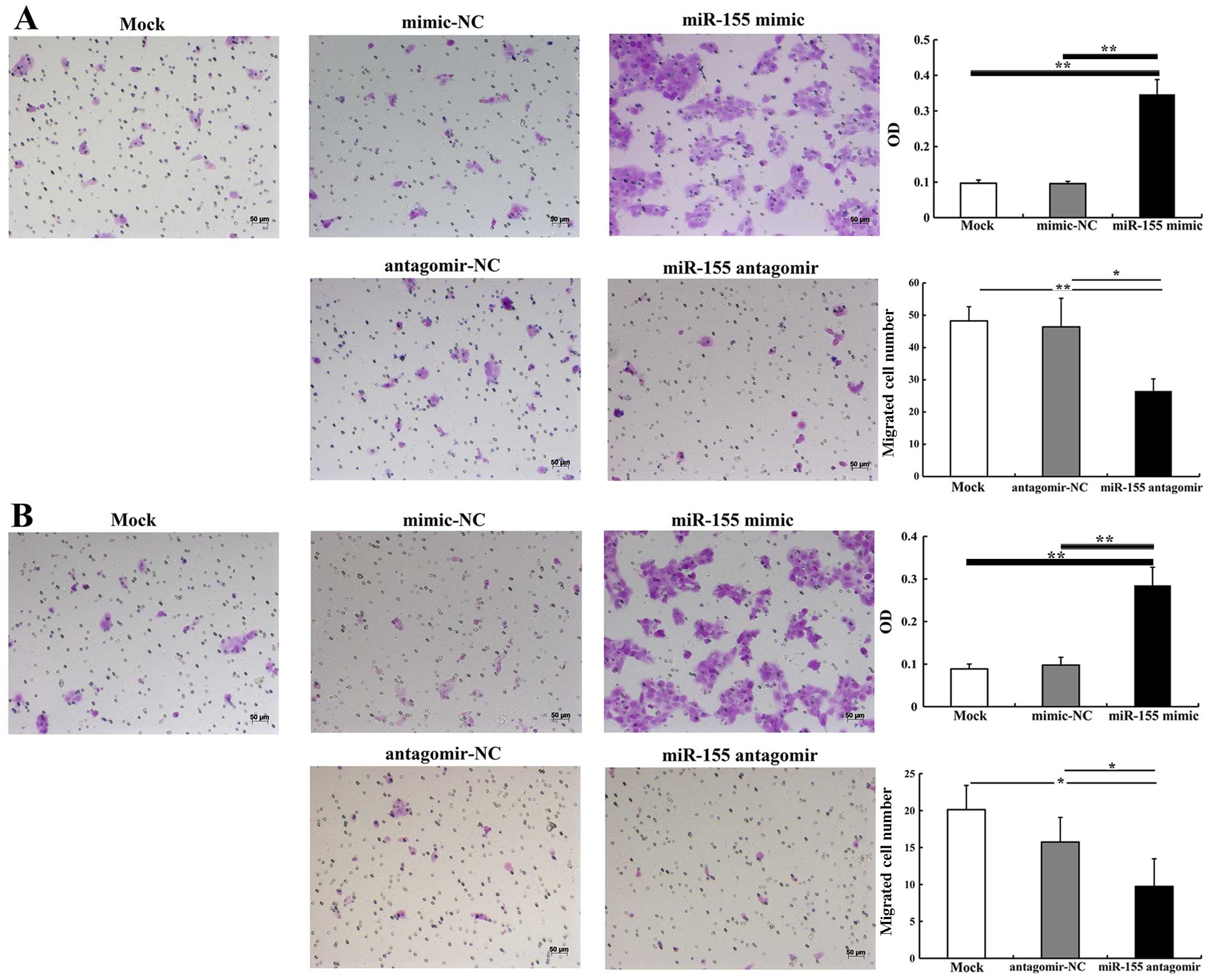
The effect of miR-155 on CAL27 cell migration and
invasion was detected using cell migration and invasion assays. At
24 h after transfection, the number of migrated cells in the group
transfected with 5 nM miR-155 mimic was significantly higher than
in the group treated with mimic-NC (P<0.01) (Fig. 3A). However, there were
significantly fewer migrated cells in the group transfected with
100 nM miR-155 antagomir compared with the group treated with
antagomir-NC (P<0.05) (Fig.
3B). The invasion assay demonstrated that 5 nM miR-155 mimic
significantly increased the number of invasive cells when compared
with mimic-NC (P<0.01). There were significantly fewer invasive
cells in the group treated with 100 nM miR-155 antagomir than in
the group treated with antagomir-NC (P<0.05) (Fig. 3B). Thus, we suggest that miR-155
effectively enhances the migratory and invasive abilities of OSCC
cells.
miR-155 regulates BCL6 and cyclin D2
expression in OSCC cells
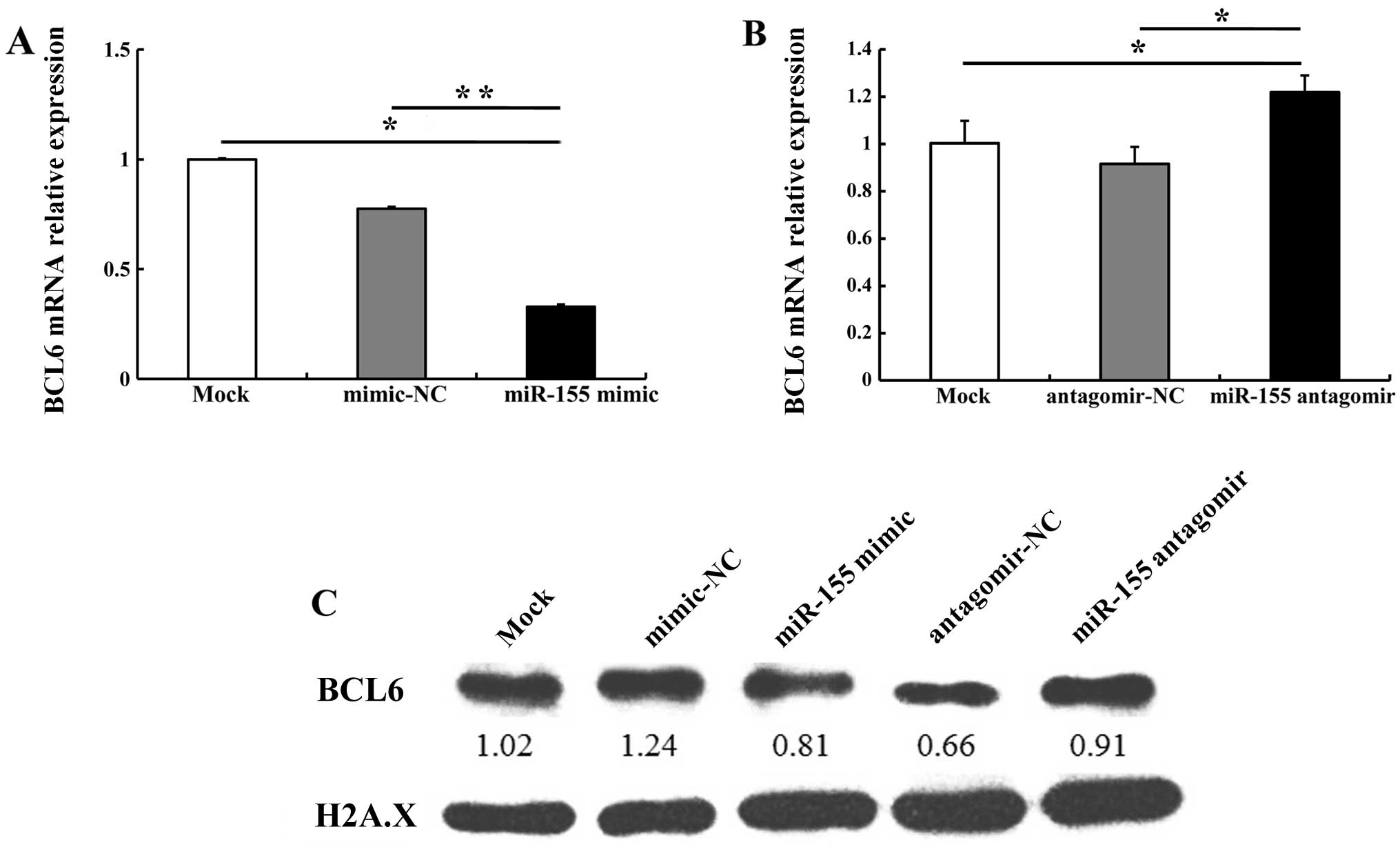
It has previously been reported that miR-155
directly regulates BCL6 expression (17). In the present study,
bioinformatics analyses using miRanda (http://www.microrna.org/microrna/home.do) and
miRTarBase (http://mirtarbase.mbc.nctu.edu.tw/) revealed that the
3′ untranslated region (3′UTR) of BCL6 mRNA contained a
highly conserved miR-155 binding site (data not shown). Therefore,
in order to validate whether miR-155 regulates BCL6 and its
downstream gene cyclin D2 in OSCC cells, we compared the alteration
in BCL6 and cyclin D2 expression in CAL27 cells transfected with
miR-155 mimic, mimic-NC, miR-155 antagomir or antagomir-NC. miR-155
mimic (5 nM) downregulated BCL6 mRNA and protein levels; 100 nM
miR-155 antagomir upregulated BCL6 mRNA and protein levels as
compared with antagomir-NC (Fig.
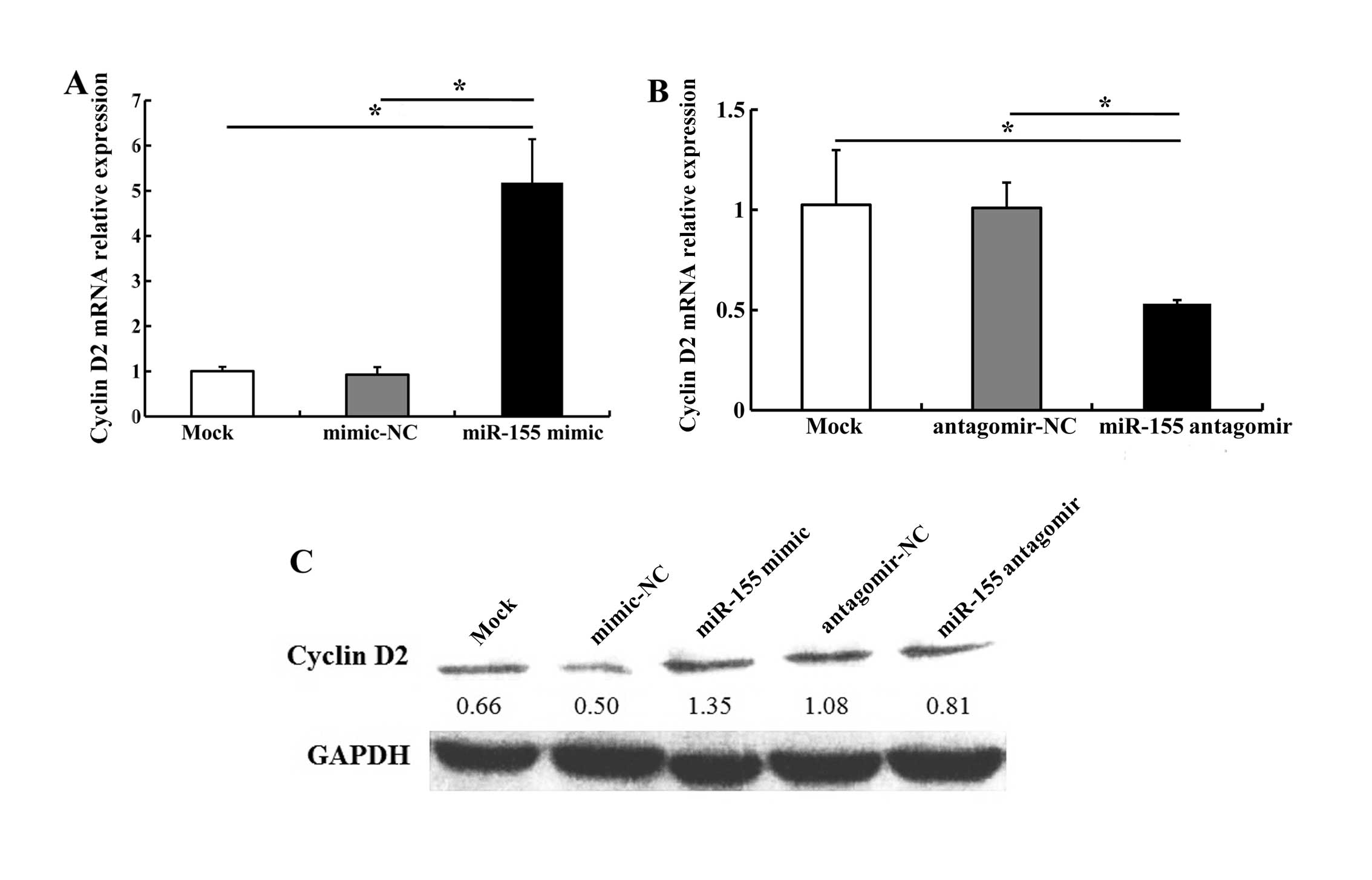
4). We noted that transfection with 5 nM miR-155 mimic
increased cyclin D2 mRNA and protein levels; 100 nM miR-155
antagomir markedly decreased cyclin D2 expression as compared with
antagomir-NC (Fig. 5). These
results indicate that miR-155 regulates the expression of BCL6 and
cyclin D2.
Discussion
It has been shown in previous studies that miR-155
is an important regulatory gene which is involved in the
carcinogenesis of various epithelial tumors, and is also involved
in tumor cell proliferation, invasion and other biological
behaviors. miR-155 is always upregulated in laryngeal squamous cell
carcinoma, cervical and colon cancer (9,10,23). To the best of our knowledge, there
are but few studies on the role of miR-155 in OSCC (16,24,25). In the present study, we detected
miR-155 expression in the OSCC cell line CAL27 and in NOK-SI, the
control cell line. We found that miR-155 expression was higher in
CAL27 cells than in NOK-SI cells. We also compared miR-155
expression in paired tumor and non-tumor tissues from patients with
OSCC, and found that miR-155 was upregulated in the tumor tissues.
These results are consistent with the findings of previous studies
(16,24,25). Our results and those of others
indicate that miR-155 participates in the development of OSCC, and
thus our understanding of the pathogenesis of OSCC has been
improved.
Cell proliferation is required for tumor growth and
acts as the foundation for cell invasion and migration. miR-155
upregulates cell proliferation in several types of cancer,
including laryngeal squamous cell carcinoma, breast and pancreatic
cancer (9,12,13). To date, relatively few studies on
the effect of miR-155 on OSCC cell proliferation have been
undertaken. Recently, Rather et al (16) reported that antagomir-155
decreased the proliferation of the OSCC cell line KB through CDC73.
In the present study, to explore whether miR-155 promotes
proliferation in OSCC, we transfected CAL27 cells with miR-155
mimic and antagomir. miR-155 mimic was used to imitate miR-155
function, and miR-155 antagomir was used to abolish the effect of
miR-155. We demonstrated that miR-155 mimic increased CAL27 cell
proliferation, whereas miR-155 antagomir reduced it, indicating
that miR-155 plays a proliferation-promoting role in OSCC.
Cell motility is critical for the spread of tumor
cells to the lymph nodes and blood vessels. miR-155 functions as a
migration- and invasion-promoting gene in many cancers: e.g., it
promotes migration and invasion in laryngeal squamous cell
carcinoma by targeting the SOCS1-STAT3 pathway (9). In breast cancer, miR-155 mediates
the loss of CCAAT/enhancer binding protein β (C/EBPβ) and shifts
the response to transforming growth factor-β (TGF-β) from growth
inhibition to invasion and metastasis (26). However, to the best of our
knowledge, no study on the effect of miR-155 on OSCC cell migratory
and invasive ability had previously been undertaken. In the present
study, by examining the gain or loss of function of miR-155, we
discovered that miR-155 enhances OSCC cell migration and invasion.
These findings demonstrate that miR-155 plays an important role in
the progression of OSCC by enhancing tumor cell migratory and
invasive abilities.
Typically, miR-155 exerts its regulatory functions
by binding to the 3′UTR of the target gene. miR-155 targets
FOXO3, SOCS1, claudin-1 and other genes in tumors
(9,14,15). Moreover, it has been reported that
the transcriptional repressor BCL6, which inhibits cell growth and
induces apoptosis, is a target gene of miR-155, and it has been
confirmed using a luciferase assay that miR-155 binds to the
conserved binding site in the 3′UTR of BLC6 mRNA and
regulates the expression of BCL6 (17,27). In the present study, we found that
miR-155 mimic significantly decreased BCL6, while miR-155 antagomir
significantly increased it. Thus, we deduced that miR-155 regulated
BCL6. Considering that the aforementioned previous studies have
verified that miR-155 binds to the conserved binding site in the
3′UTR of BCL6 through luciferase assays, we did not
undertake the same assay in our study.
BCL6 plays a role in cell growth and other
biological behaviors by regulating downstream genes such as
CCND2, and various chemokines (18). Previous research has reported that
cyclin D2 plays an important role in G1-to-S phase shifting in the
cell cycle (28). Moreover,
cyclin D2 overexpression results in excessive proliferation in many
cell types and enhances OSCC cell invasive ability in vitro
and in vivo (19). It has
also been reported that BCL6 directly regulates the expression of
cyclin D2 by binding with the conserved site in its promoter, as
evidenced by luciferase reporter assay, and BCL6 protein bound
specifically to cyclin D2, as determined using electrophoretic
mobility shift DNA binding assay (20). Several studies have demonstrated
that BCL6/cyclin D2 are involved in many disorders. The BCL6/cyclin
D2 axis affects the cell cycle and proliferation by regulating
pro-HB-EGF and FOXO (21,22,29). In the present study, cyclin D2
levels were increased, and BCL6 was repressed, by miR-155 mimic,
and these levels were decreased when miR-155 antagomir decreased
BCL6 repression. These data indicate that ectopic expression of
miR-155 downregulates BCL6 expression and increases cyclin D2
expression, and subsequently facilitates cell proliferation,
migration and invasion. Moreover, as miR-155 binds to the 3′UTR of
BCL6 mRNA to regulate BCL6 expression, and as CCND2
is a target gene of BCL6, we infer that miR-155 promotes OSCC
carcinogenesis and development partly through the BCL6/cyclin D2
axis. In future research, we will explore the critical roles of
BCL6/cyclin D2 in miR-155-induced proliferation, migration and
invasion in OSCC cells.
In conclusion, miR-155 overexpression plays a
promoting role in the proliferative, migratory and invasive
behavior of OSCC cells. Its effects on OSCC are possibly associated
with its regulation of the BCL6/cyclin D2 axis. Collectively, our
results are of great significance for understanding OSCC
development. However, further research is required to fully
elucidate the regulatory mechanisms of BCL6/cyclin D2 mediating the
miR-155-induced promoting effects in OSCC.
Acknowledgments
This study was supported by grants from the National
Natural Science Foundation of China (nos. 81272948 and 81000446),
the Specialized Research Fund for the Doctoral Program of Higher
Education of China (201001711201050), the Provincial Natural
Scientific Foundation of Guangdong (10451008901005050), the
Guangdong Department of Science and Technology Translational
Medicine Center Grant (2011A080300002) and the Priming Scientific
Research Foundation for Junior Teachers of Medicine in Sun Yat-sen
University (10ykpy17).
References
|
1
|
Choi S and Myers JN: Molecular
pathogenesis of oral squamous cell carcinoma: implications for
therapy. J Dent Res. 87:14–32. 2008. View Article : Google Scholar
|
|
2
|
Gibson MK and Forastiere AA:
Multidisciplinary approaches in the management of advanced head and
neck tumors: state of the art. Curr Opin Oncol. 16:220–224. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sasahira T, Kirita T and Kuniyasu H:
Update of molecular pathobiology in oral cancer: a review. Int J
Clin Oncol. 19:431–436. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bushati N and Cohen SM: microRNA
functions. Annu Rev Cell Dev Biol. 23:175–205. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kluiver J, van den Berg A, de Jong D,
Blokzijl T, Harms G, Bouwman E, Jacobs S, Poppema S and Kroesen BJ:
Regulation of pri-microRNA BIC transcription and processing in
Burkitt lymphoma. Oncogene. 26:3769–3776. 2007. View Article : Google Scholar
|
|
8
|
Heegaard NH, Schetter AJ, Welsh JA, Yoneda
M, Bowman ED and Harris CC: Circulating micro-RNA expression
profiles in early stage nonsmall cell lung cancer. Int J Cancer.
130:1378–1386. 2012. View Article : Google Scholar :
|
|
9
|
Zhao XD, Zhang W, Liang HJ and Ji WY:
Overexpression of miR-155 promotes proliferation and invasion of
human laryngeal squamous cell carcinoma via targeting SOCS1 and
STAT3. PLoS One. 8:e563952013. View Article : Google Scholar
|
|
10
|
Bakirtzi K, Hatziapostolou M,
Karagiannides I, Polytarchou C, Jaeger S, Iliopoulos D and
Pothoulakis C: Neurotensin signaling activates microRNAs-21 and
-155 and Akt, promotes tumor growth in mice, and is increased in
human colon tumors. Gastroenterology. 141:1749–61.e1. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yan XL, Jia YL, Chen L, Zeng Q, Zhou JN,
Fu CJ, Chen HX, Yuan HF, Li ZW, Shi L, et al: Hepatocellular
carcinoma-associated mesenchymal stem cells promote hepatocarcinoma
progression: role of the S100A4-miR155-SOCS1-MMP9 axis. Hepatology.
57:2274–2286. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gironella M, Seux M, Xie MJ, Cano C,
Tomasini R, Gommeaux J, Garcia S, Nowak J, Yeung ML, Jeang KT, et
al: Tumor protein 53-induced nuclear protein 1 expression is
repressed by miR-155, and its restoration inhibits pancreatic tumor
development. Proc Natl Acad Sci USA. 104:16170–16175. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang CM, Zhao J and Deng HY: MiR-155
promotes proliferation of human breast cancer MCF-7 cells through
targeting tumor protein 53-induced nuclear protein 1. J Biomed Sci.
20:79–88. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ling N, Gu J, Lei Z, Li M, Zhao J, Zhang
HT and Li X: microRNA-155 regulates cell proliferation and invasion
by targeting FOXO3a in glioma. Oncol Rep. 30:2111–2118.
2013.PubMed/NCBI
|
|
15
|
Zhang GJ, Xiao HX, Tian HP, Liu ZL, Xia SS
and Zhou T: Upregulation of microRNA-155 promotes the migration and
invasion of colorectal cancer cells through the regulation of
claudin-1 expression. Int J Mol Med. 31:1375–1380. 2013.PubMed/NCBI
|
|
16
|
Rather MI, Nagashri MN, Swamy SS, Gopinath
KS and Kumar A: Oncogenic microRNA-155 down-regulates tumor
suppressor CDC73 and promotes oral squamous cell carcinoma cell
proliferation: implications for cancer therapeutics. J Biol Chem.
288:608–618. 2013. View Article : Google Scholar :
|
|
17
|
Nazari-Jahantigh M, Wei Y, Noels H, Akhtar
S, Zhou Z, Koenen RR, Heyll K, Gremse F, Kiessling F, Grommes J, et
al: MicroRNA-155 promotes atherosclerosis by repressing Bcl6 in
macrophages. J Clin Invest. 122:4190–4202. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dent AL, Vasanwala FH and Toney LM:
Regulation of gene expression by the proto-oncogene BCL-6. Crit Rev
Oncol Hematol. 41:1–9. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu SC, Bassi DE, Zhang SY, Holoran D,
Conti CJ and Klein-Szanto AJ: Overexpression of cyclin D2 is
associated with increased in vivo invasiveness of human squamous
carcinoma cells. Mol Carcinog. 34:131–139. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fernández de Mattos S, Essafi A, Soeiro I,
Pietersen AM, Birkenkamp KU, Edwards CS, Martino A, Nelson BH,
Francis JM, Jones MC, et al: FoxO3a and BCR-ABL regulate cyclin D2
transcription through a STAT5/BCL6-dependent mechanism. Mol Cell
Biol. 24:10058–10071. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hirata Y, Ogasawara N, Sasaki M, Mizushima
T, Shimura T, Mizoshita T, Mori Y, Kubota E, Wada T, Tanida S, et
al: BCL6 degradation caused by the interaction with the C-terminus
of pro-HB-EGF induces cyclin D2 expression in gastric cancers. Br J
Cancer. 100:1320–1329. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Glauser DA and Schlegel W: The
FoxO/Bcl-6/cyclin D2 pathway mediates metabolic and growth factor
stimulation of proliferation in Min6 pancreatic beta-cells. J
Recept Signal Transduct Res. 29:293–298. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lao G, Liu P, Wu Q, Zhang W, Liu Y, Yang L
and Ma C: Mir-155 promotes cervical cancer cell proliferation
through suppression of its target gene LKB1. Tumour Biol.
35:11933–11938. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shi LJ, Zhang CY, Zhou ZT, Ma JY, Liu Y,
Bao ZX and Jiang WW: MicroRNA-155 in oral squamous cell carcinoma:
Overexpression, localization, and prognostic potential. Head Neck.
37:970–976. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gombos K, Horváth R, Szele E, Juhász K,
Gocze K, Somlai K, Pajkos G, Ember I and Olasz L: miRNA expression
profiles of oral squamous cell carcinomas. Anticancer Res.
33:1511–1517. 2013.PubMed/NCBI
|
|
26
|
Johansson J, Berg T, Kurzejamska E, Pang
MF, Tabor V, Jansson M, Roswall P, Pietras K, Sund M, Religa P and
Fuxe J: MiR-155-mediated loss of C/EBPβ shifts the TGF-β response
from growth inhibition to epithelial-mesenchymal transition,
invasion and metastasis in breast cancer. Oncogene. 32:5614–5624.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sandhu SK, Volinia S, Costinean S, Galasso
M, Neinast R, Santhanam R, Parthun MR, Perrotti D, Marcucci G,
Garzon R and Croce CM: miR-155 targets histone deacetylase 4
(HDAC4) and impairs transcriptional activity of B-cell lymphoma 6
(BCL6) in the Eμ-miR-155 transgenic mouse model. Proc Natl Acad Sci
USA. 109:20047–20052. 2012. View Article : Google Scholar
|
|
28
|
Ponzio G, Loubat A, Rochet N, Turchi L,
Rezzonico R, Farahi Far D, Dulic V and Rossi B: Early G1 growth
arrest of hybridoma B cells by DMSO involves cyclin D2 inhibition
and p21(CIP1) induction. Oncogene. 17:1159–1166. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shaffer AL, Yu X, He Y, Boldrick J, Chan
EP and Staudt LM: BCL-6 represses genes that function in lymphocyte
differentiation, inflammation, and cell cycle control. Immunity.
13:199–212. 2000. View Article : Google Scholar : PubMed/NCBI
|