Introduction
The World Health Organization estimates that 235
million individuals worldwide suffered from asthma in 2013
(1). In the United States, an
estimated 25.9 million individuals, including almost 7.1 million
children, suffered from asthma in 2013, and asthma was the third
most common cause of hospitalization for children under 15 years of
age (2). The symptoms of asthma
are difficult to control (3), and
its causes are diverse, including hereditary factors and external
factors, such as pet dander, dust mites, cockroaches, viral
infections, pollen, mold, fungi and tobacco smoke (1). The typical manifestations of asthma
vary from a cough to obstructive apnea, which may arise due to the
excessive production of mucus, goblet cell hyperplasia, epithelial
cell shedding, basement membrane thickening, as well as eosinophil
and lymphocyte infiltration (4,5).
Asthma is a chronic lung condition that involves an
imbalance between T helper (Th)1- and Th2-related factors (4,5),
including interleukin (IL)-12, interferon-γ (IFN-γ), IL-4, IL-5,
IL-13, IL-6 and tumor necrosis factor-α (TNF-α). IL-12 modulates
the induction of Th1-specific immune responses and the suppression
of Th2-specific immune responses (6,7).
IFN-γ is known to stimulate the Th1 transcription factor, T-bet,
resulting in a positive feedback loop between IFN-γ and Th1
(8,9). The cytokines, IL-4 and IL-13, play
important roles in asthma (10).
IL-4 regulates the immunoglobulin class switching from IgG to IgE,
attracts eosinophils into the interspace of pulmonary cells
(11), and is involved in the
induction of the Th2 transcription factor GATA-3 (12,13,15). IL-13 activates B cells and induces
asthma-related changes, such as the excessive production of mucus,
goblet cell hyperplasia, epithelial cell shedding, basement
membrane thickening, and eosinophil and lymphocyte infiltration
(14,16,17). Both IL-5 and IL-6 have been
demonstrated to regulate the development, activation, migration and
survival of eosinophils (6,18).
Furthermore, TNF-α is known to recruit granulocytes and to induce
the proliferation of fibroblasts (19).
The drugs currently used to treat asthma include
corticosteroids, bronchodilators, leukotriene modifiers,
theophylline and anti-IgE agents; however, their therapeutic
effects are not completely understood (20). Although inhaled corticosteroids
are the most widely used therapy for suppressing the symptoms of
asthma (21), they are associated
with many adverse effects, including growth inhibition in children
during the first year of treatment (22), cataracts and glaucoma,
hypertension, hyperlipidemia, peptic ulcers, myopathy and
immunosuppressive effects (23).
Thus, the unwanted side-effects of corticosteroids have increased
the need to develop anti-asthmatic drugs from natural products.
Erythronium japonicum (E. japonicum)
is an indigenous herb in Korea and East Asian countries (24). It is distributed throughout
Hokkaido in Japan, where the starch ('katakuriko' in Japanese) is
obtained from the bulb after a long, cold winter. Although a
limited number of studies have explored its potentially therapeutic
effects, E. japonicum extract was found in one study to
possess 2,2-diphenyl-1-picrylhydrazyl free radical scavenging
activity and to exert anti-proliferative effects in human
colorectal carcinoma cells (25).
In the present study, we evaluated the effects of E.
japonicum extract on ovalbumin (OVA)-induced asthma in
mice.
Materials and methods
Plant material and preparation of 80%
EtOH extract
During May 2013, E. japonicum was collected
from a site situated close to Wolchul mountain in the southern part
of Korea. A sample was deposited at the Jeonnam Biofood Technology
Center (identification number: JBF-FRI-S-2013-0099). E.
japonicum was dried in a dark, cool room. Dried E.
japonicum (2 kg) was chopped and then extracted twice with 80%
aqueous EtOH (30 liters) at room temperature for 24 h. The EtOH
extracts of E. japonicum were concentrated and evaporated
under a vacuum. E. japonicum extract was analyzed using
high-performance liquid chromatography (HPLC).
Establishment of a mouse model of
OVA-induced asthma
Female BALB/c mice (5 weeks old, n=80) were
purchased from Samtako Korea (Osan, Korea) and randomly divided
into the following 5 treatment groups: i) the control group (no
treatment, no OVA challenge); ii) the group administered sterilized
tap water and challenged with OVA; iii) the group administered 1
mg/kg/day dexamethasone followed by OVA challenge; iv) the group
administered 60 mg/kg/day E. japonicum extract followed by
OVA challenge; and v) the group administered 600 mg/kg/day E.
japonicum extract followed by OVA challenge. On days 1 and 8,
all mice were sensitized with an intraperitoneal injection of 20
µg OVA and 1 mg aluminum hydroxide hydrate (both from
Sigma-Aldrich, St. Louis, MO, USA) in 500 µl saline. On days
21 to 25, all mice except those used as controls were challenged
once daily with 5% OVA for 30 min using a nebulizer (3 ml/min,
NE-U17; Omron Co. Ltd., Kyoto, Japan). During the same 5-day
period, the mice in the treatment groups were also treated once
daily with oral doses of either sterilized tap water,
dexamethasone, or 60 or 600 mg/kg/day E. japonicum extract
(hereafter referred to as E. japonicum) 1 h prior to the OVA
challenge.
Ethics statement
E. japonicum was collected on private land
with permission granted by the owner. All experiments were approved
by the Institutional Animal Care and Use Committee at Dongshin
University (approval no. 2014-08-04).
Analysis of bronchoalveolar lavage fluid
(BALF)
One day after the final treatment, the mice were
anesthetized with intraperitoneal injections of 50 mg/kg Zoletin
(Virbac, Fort Worth, TX, USA), and thereafter the tracheas were
cannulated with disposable animal feeding needles. Lavages were
performed with three 0.4 ml aliquots of cold phosphate-buffered
saline (PBS). The BALF samples were collected and immediately
centrifuged at 3,000 rpm for 5 min (Sorvall Legend Micro 17R;
Thermo Fisher Scientific, Inc., Marietta, OH, USA). The cell
pellets were resuspended in PBS in order to determine the total and
differential white blood cell (WBC) counts. The numbers of total
and differential cells were counted using the Hemavet Multispecies
Hematology System (Drew Scientific, Inc., Waterbury, CT, USA)
(n=8/group). The levels of IgE in the BALF were measured using a
specific mouse IgE enzyme-linked immunosorbent assay kit (Abnova,
Atlanta, GA, USA), according to the manufacturer's instructions
(n=8/group).
Histopathological analysis
One day after the final treatment, the mice were
anesthetized with intraperitoneal injections of 50 mg/kg Zoletin
and lung tissue was obtained. The mice were then sacrificed by
exsanguination. The lung tissue was fixed in 10% (v/v) formaldehyde
solution, dehydrated in a graded ethanol series (99.9, 90, 80 and
70%), and embedded in paraffin. The paraffin-embedded lung tissue
was then sectioned (4-µm-thick sections) longitudinally and
stained with hematoxylin and eosin. The sections were also stained
with Periodic acid-Schiff for the semi-quantitative analysis of
glycoproteins.
Immunohistochemical analysis
The deparaffinized tissue sections were treated with
3% hydrogen peroxide in methanol for 10 min to remove the
endogenous peroxidase. Antigen retrieval was performed with sodium
citrate buffer (0.1 M) using the microwave method. The slides were
incubated with normal serum to block non-specific binding and then
incubated overnight at 4°C with the following primary antibodies
(diluted 1:100 to 1:200): rat anti-mouse CD4 monoclonal (14-9766;
eBioscience, San Diego, CA, USA); rat anti-mouse CD8 monoclonal
(sc-18913; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA);
rabbit anti-mouse CD19 polyclonal (250585; Abbiotec, San Diego, CA,
USA); rabbit anti-mouse Tbx21/T-bet polyclonal (bs-3599R; Bioss,
Woburn, MA, USA); goat anti-mouse GATA-3 (TA305795; OriGene,
Rockville, MD, USA); rat anti-mouse IFN-γ monoclonal (sc-74104),
goat anti-mouse IL-12p35 polyclonal (sc-9350) and rat anti-mouse
IL-12p40 monoclonal (sc-57258) (all from Santa Cruz Biotechnology,
Inc.); rabbit anti-mouse TNF-α polyclonal (3053R-100; BioVision,
Milpitas, CA, USA); rat anti-mouse IL-4 monoclonal (sc-73318) and
rabbit anti-mouse IL-5 polyclonal (sc-7887) (both from Santa Cruz
Biotechnology, Inc.); rabbit anti-mouse IL-6 polyclonal (PAB16165;
Abnova, Taipei City, Taiwan), and goat anti-mouse IL-13 polyclonal
(sc-1776; Santa Cruz Biotechnology, Inc.) antibodies. Subequently,
the slides were incubated for 2 h with biotinylated secondary
antibody (1:500; Dako, Carpinteria, CA, USA) and
horseradish-peroxidase conjugated streptavidin. Signals were
detected using 3,3-diaminobenzidine tetrahydrochloride substrate
chromogen solution, and the cells were counterstained with Mayer's
hematoxylin. To determine the number of positively stained cells,
we counted the cells in 5 randomly selected, non-overlapping fields
(magnification, ×200) from 3 separately immunostained lung sections
per animal (n=8/group).
HPLC analysis
HPLC analysis was performed using an HPLC system
(Agilent Technology, Santa Clara, CA, USA) according to the
following parameters: column, Zorbax extend-C18 (4.6 mm inner
diameter × 150 mm height, 5.0-µm particle diameter); mobile
phase, 0.2% acetic acid and methanol used as A and B, respectively,
with the following solvent gradient: 25% B, 0–5 min; 25–50% B, 5–10
min; and 50–25% B, 10–15 min; injection volume, 10 µl; flow
rate, 1 ml/min; UV detection, 330 nm. All solvents were of
extra-pure grade. Methanol and distilled water were purchased from
J.T. Baker (Phillipsburg, NJ, USA). Acetic acid, caffeic acid and
chlorogenic acid were purchased from Sigma-Aldrich.
Statistical analysis
Data are shown as the means ± standard deviation
(SD). Group differences were evaluated using one-way analysis of
variance followed by Dunnett's multiple comparison tests. A p-value
of <0.05 or <0.001 was considered to indicate a statistically
significant difference.
Results
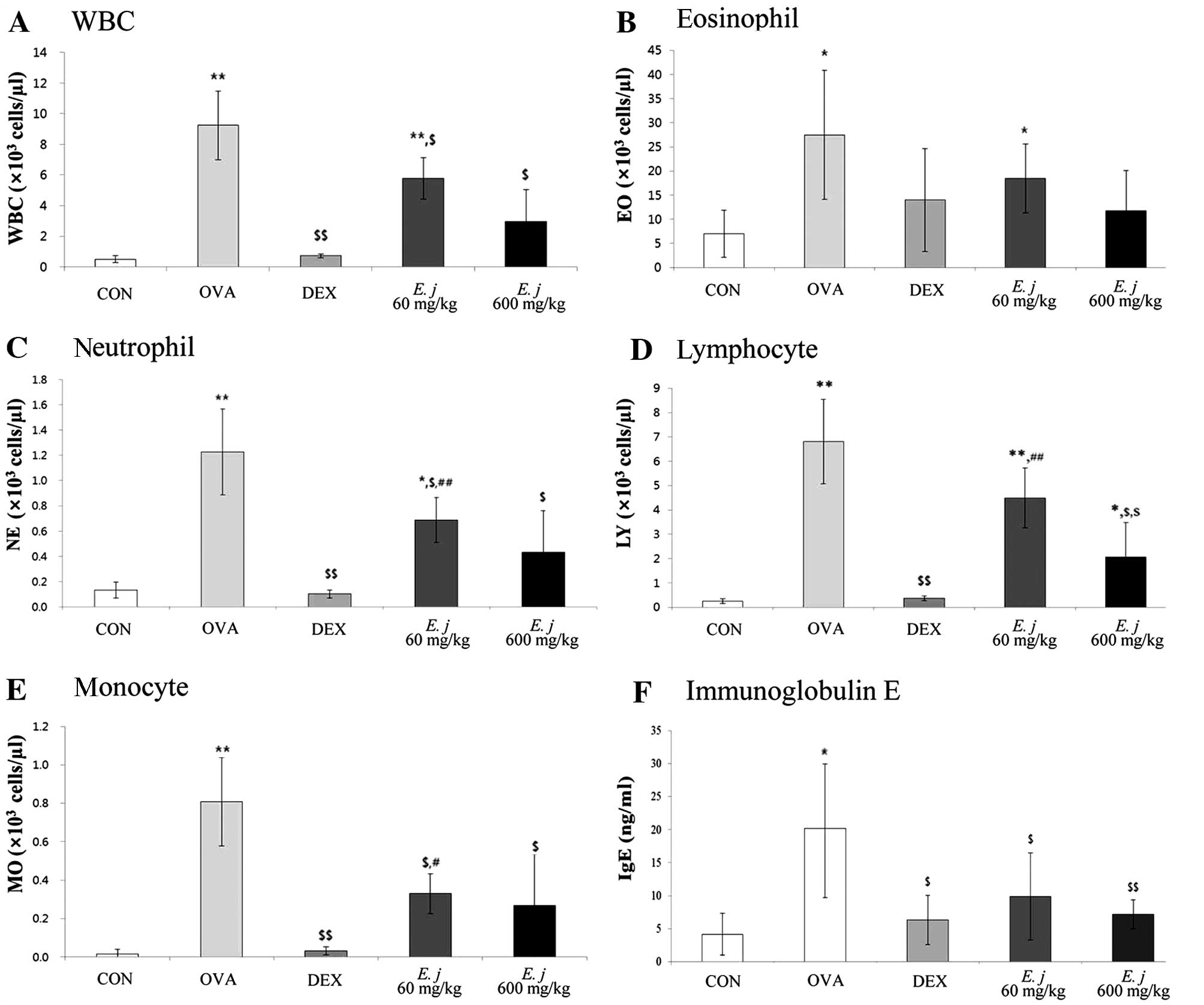
Effects of E. japonicum on the total and
differential WBC counts and on the IgE levels in the BALF
samples
Compared with the BALF obtained from the control
mice, the BALF collected from the OVA-challenged mice contained
significantly higher numbers of total WBCs and differential WBCs
(eosinophils, neutrophils, lymphocytes and monocytes), as well as
significantly higher levels of IgE (Fig. 1). Treatment with both
dexamethasone, a drug commonly used to suppress the symptoms of
asthma, and E. japonicum significantly decreased the number
of WBCs, as well as the IgE levels in the OVA-challenged mice.
Effects of E. japonicum on lung tissue
morphology
The asthma-related histopathological changes in the
mouse lung tissues were evaluated using hematoxylin and eosin
staining and Periodic acid-Schiff staining (Fig. 2). Compared with the lung tissue
isolated from the control mice, the lung tissue from the
OVA-challenged mice exhibited typical asthma-related changes,
including goblet cell and epithelial cell hyperplasia, eosinophil
infiltration around the bronchioles and vessels, as well as the
increased secretion of mucus in the bronchioles, and these changes
were attenuated by treatment with dexamethasone. Treatment with
E. japonicum also attenuated these histopathological changes
in a dose-dependent manner; 600 mg/kg/day of E. japonicum
appeared to be more effective at inhibiting the OVA-induced changes
in the mouse lung tissue compared with the dose of 60
mg/kg/day.
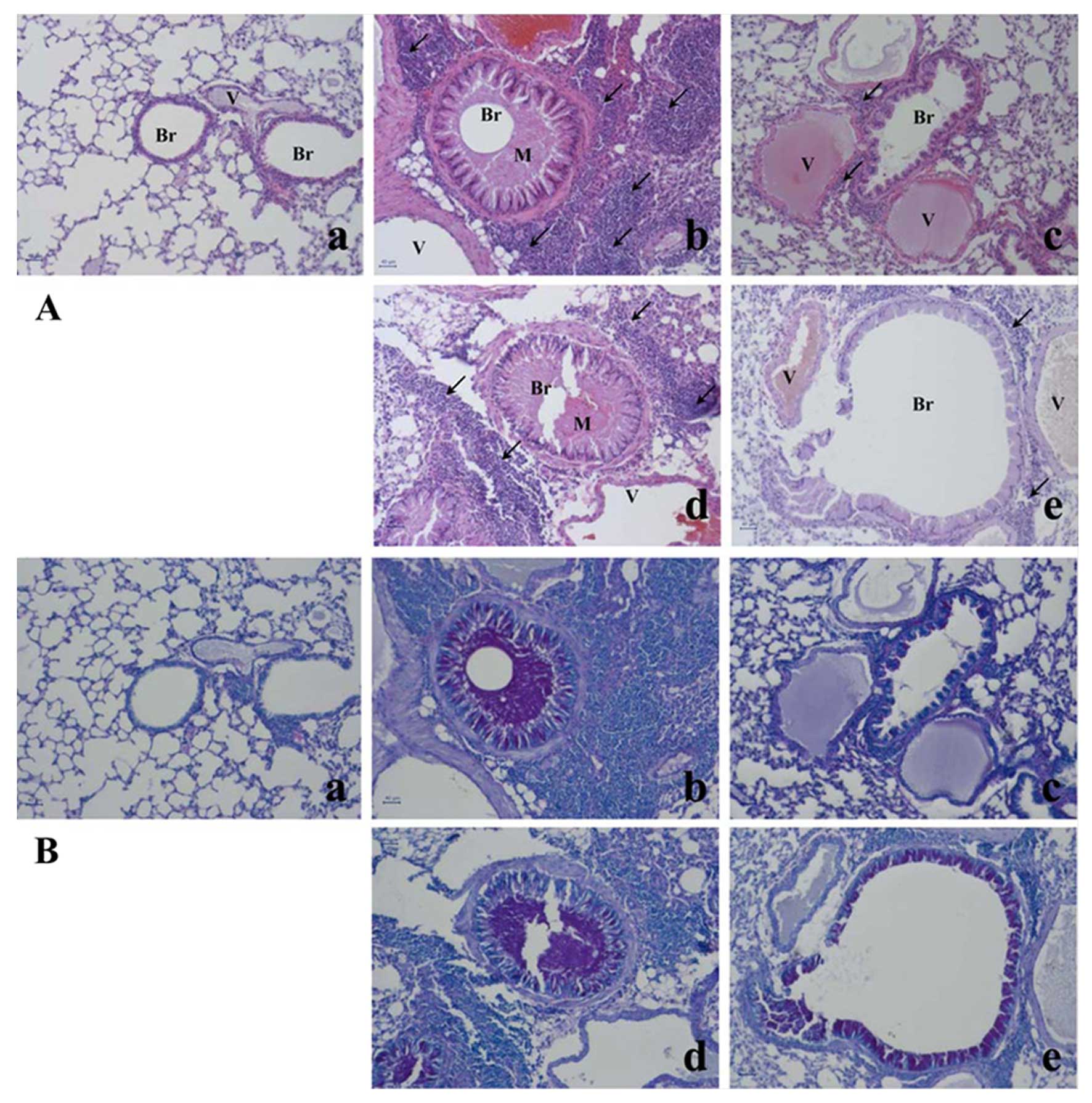 | Figure 2Effects of E. japonicum on
lung tissue morphology. (A) Representative images of hematoxylin
and eosin-stained lung tissue. (B) Representative images of
Periodic acid-Schiff-stained lung tissue. Scale bar, 50 µm;
arrows, eosinophil infiltration; Br, bronchiole; M, mucus
secretion; V, vessel. Treatment groups: panel a, control; panel b,
ovalbumin; panel c, dexamethasone; panel d, 60 mg/kg/day E.
japonicum; panel e, 600 mg/kg/day E. japonicum. |
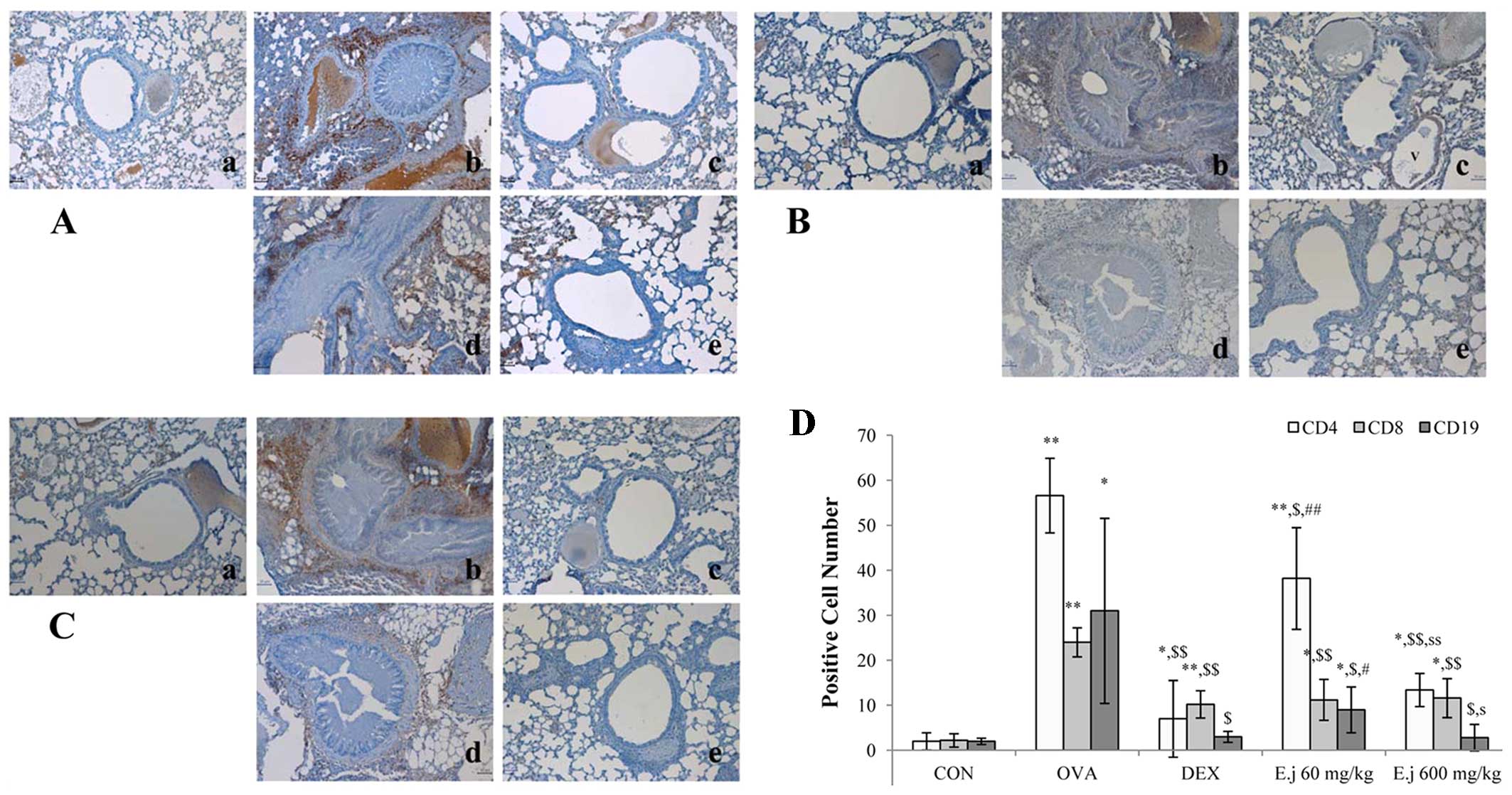
Effects of E. japonicum on the numbers of
Th cells, T cells, and B cells
The development of asthma involves an imbalance
between Th1 and Th2 cells (4,5).
In order to evaluate the effects of E. japonicum treatment
on Th cells, T cells and B cells, we quantified the numbers of
CD4+ Th cells, CD8+ cytotoxic T cells and
CD19+ B cells in the mouse lung tissue by performing
immunohistochemical analysis (Fig.
3). Compared with the control mice, the lung tissue isolated
from the OVA-challenged mice had significantly higher numbers of
all 3 types of cells, and these increased numbers were
significantly reduced by treatment with both dexamethasone and
E. japonicum. Furthermore, treatment with E.
japonicum exerted a reducing effect on the numbers of
CD4+ Th cells and CD19+ B cells in a
dose-dependent manner, with greater effects observed at the dose of
600 mg/kg/day compared with 60 mg/kg/day.
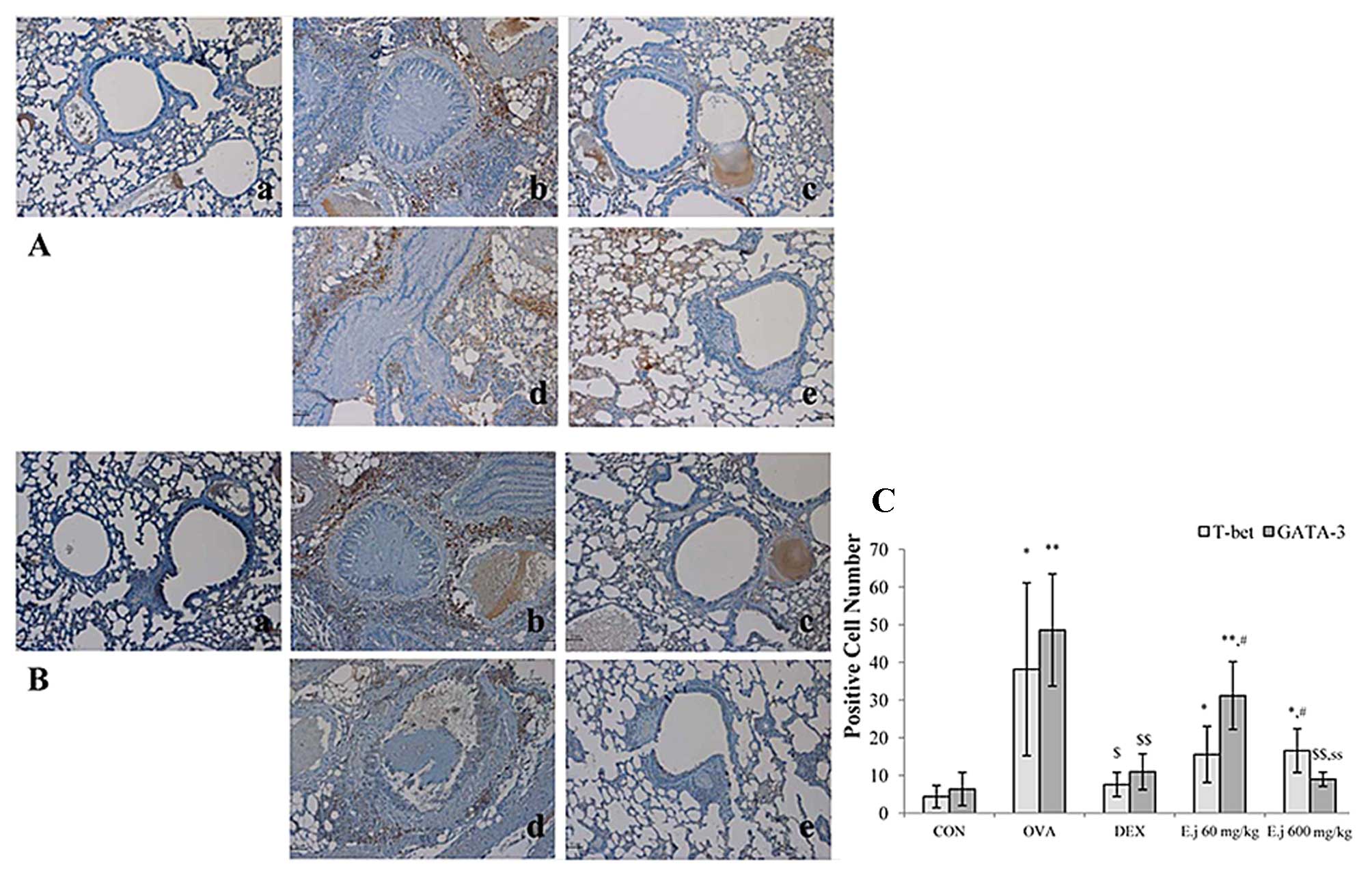
Effects of E. japonicum on the expression
of T-bet and GATA-3
To examine the effects of E. japonicum
treatment on Th cell transcription factors, we examined the
expression of T-bet (Th1 cell transcription factor) and GATA-3 (Th2
cell transcription factor) in the mouse lung tissue by perfoming
immunohistochemical analysis (Fig.
4). Compared with the control mice, the lung tissue isolated
from the OVA-challenged mice exhibited a significantly higher
expression of T-bet and GATA-3, and these effects were
significantly reduced by dexamethasone treatment. Treatment with
E. japonicum exerted a reducing effect on GATA-3 expression
in a dose-dependent manner, with the dose of 600 mg/kg/day
significantly reducing the OVA-induced expression of GATA-3 to a
greater extent compared with the dose of 60 mg/kg/day. However, the
expression of T-bet in the E. japonicum treatment groups
remained at higher levels compared to the control group, and there
was no significant difference between the 2 doses used.
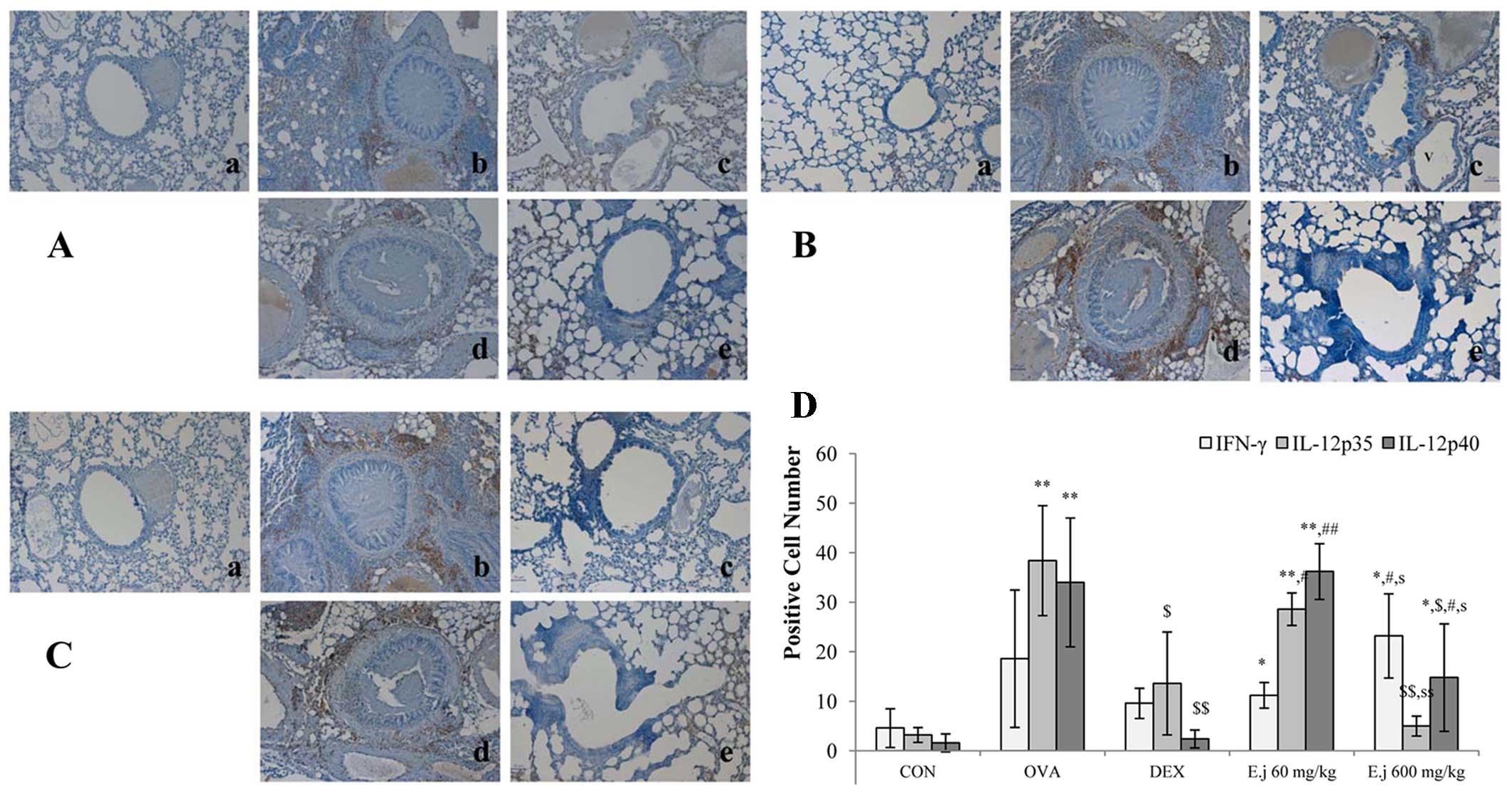
Effects of E. japonicum on the expression
of Th1-related cytokines
The imbalance between Th1 and Th2 cells which occurs
in asthma involves a decrease in Th1-related cytokine levels and an
increase in Th2-related cytokine levels (4,5).
In this study, to examine alterations in the levels of Th1-related
cytokines, we measured the expression levels of IFN-γ, IL-12p35 and
IL-12p40 in the mouse lung tissue (Fig. 5). Compared with the control mice,
the lung tissue isolated from the OVA-challenged mice exhibited a
significantly increased expression of IL-12p35 and IL-12p40, and
these effects were significantly suppressed by dexamethasone
treatment. E. japonicum treatment exerted a reducing
dose-dependent effect on the expression of IL-12p35 and IL-12p40,
with the dose of 600 mg/kg/day (but not the dose of 60 mg/kg/day)
significantly reducing the OVA-induced expression of IL-12p35 and
IL-12p40. E. japonicum treatment at the dose of 600
mg/kg/day increased the expression of IFN-γ.
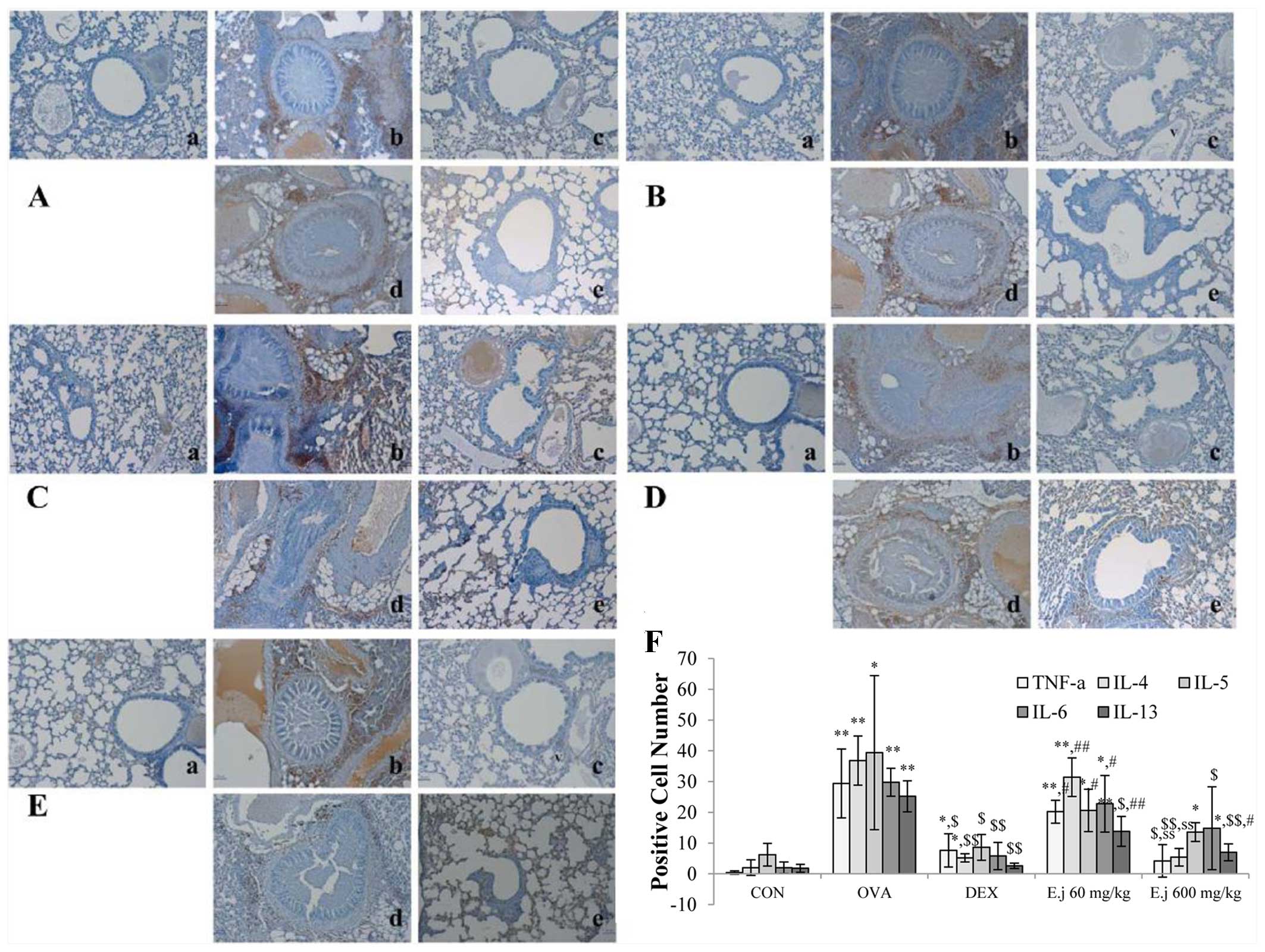
Effect of E. japonicum on the expression
of Th2-related cytokines
To examine alterations in the levels of Th2-related
cytokines, we measured the expression levels of TNF-α, IL-4, IL-5,
IL-6 and IL-13 in the mouse lung tissue (Fig. 5). Compared with the control mice,
the lung tissue isolated from the OVA-challenged mice exhibited a
significantly increased expression of all the measured cytokines,
and these changes were significantly reduced by dexamethasone
treatment (Fig. 6). E.
japonicum treatment decreased the OVA-induced expression of
most of these cytokines in a dose-dependent manner. Specifically,
E. japonicum treatment at the dose of 60 mg/kg/day
significantly reduced the expression of IL-13, whereas treatment
with E. japonicum at 600 mg/kg/day significantly reduced the
expression of TNF-α, IL-4, IL-6, IL-5 and IL-13.
Discussion
In the present study, we found that treatment with
E. japonicum suppressed the OVA-induced increase in the
number of WBCs, as well as the IgE level in BALF. Furthermore,
treatment with E. japonicum attenuated the asthma-related
morphological changes which had occurred in the mouse lung tissue
following exposure to OVA, including the increased secretion of
mucus, eosinophil infiltration around the bronchioles and vessels,
and goblet cell and epithelial cell hyperplasia. E.
japonicum treatment also suppressed the OVA-induced increase in
the number of Th cells (CD4+ cells), B cells
(CD19+ cells) and T cells (CD8+ cytotoxic
cells). Furthermore, we found that E. japonicum treatment
modulated the expression of not only Th2-related factors (GATA-3,
TNF-α, IL-4, IL-5, IL-6 and IL-13), but also that of Th1-related
factors (IFN-γ, IL-12p35 and IL-12p40).
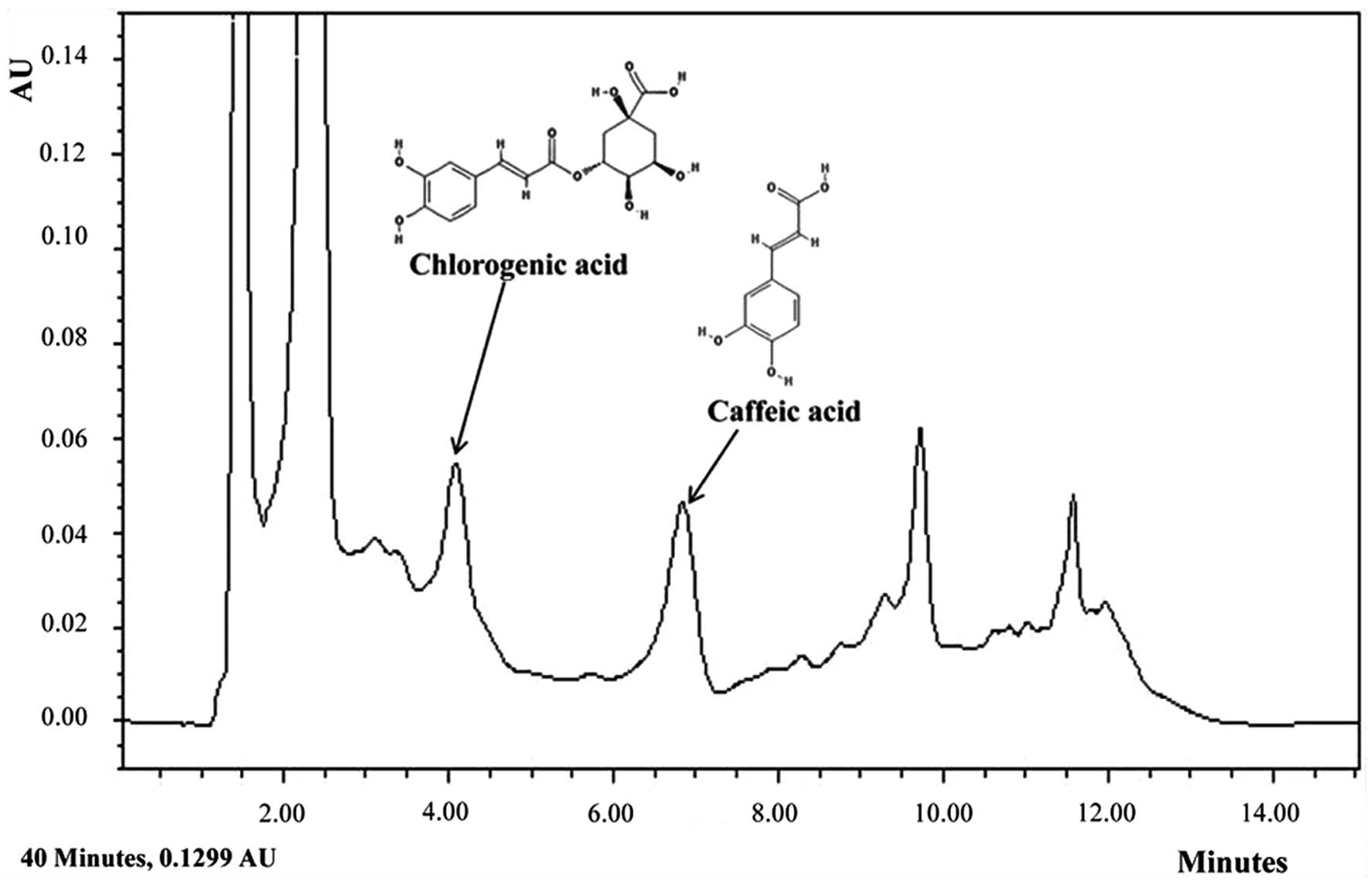
To identify the particular compounds in E.
japonicum that may exert anti-asthmatic effects, we conducted
HPLC analysis. Although not all the compounds in E.
japonicum were isolated, we found that chlorogenic acid and
caffeic acid were candidate compounds (Fig. 7). Chlorogenic acid is associated
with lignin biosynthesis (26)
and it has been demonstrated to exert several effects, including
protective effects against oxidative stress-induced hepatotoxicity
(27), anti-hypertensive effects
(28) and the suppressive effects
on several asthma-related factors in an animal model of
ovalbumin-induced asthma (29).
Furthermore, chlorogenic acid is an immunoregulatory molecule
(30).
Asthma involves an imbalance between Th1 and Th2
cell-related factors (13), which
results in airway hyperresponsiveness in asthma patients as well as
morphological changes in lung tissues such as mucus hypersecretion
in the bronchioles, goblet cell and epithelial cell hyperplasia,
and eosinophil infiltration near to the bronchioles and vessels
(4). Depending on its severity,
asthma may even be fatal (1).
Asthma is associated with changes in the expression of Th2- and
Th1-related cytokines (13),
including the increased expression of IL-4, IL-5, IL-6, IL-13 and
TNF-α and the decreased expression of IL-12 (31). In the present study, we found that
E. japonicum treatment reversed many of the OVA-induced
changes in Th1 and Th2 cell-related factors in a mouse model of
asthma, suggesting that E. japonicum may ameliorate the
symptoms of asthma by modulating the balance between Th1 and Th2
cells.
The most commonly prescribed drugs for asthma are
steroids; however, these have many adverse effects, such as growth
inhibition in children (22),
cataracts and glaucoma, hypertension, hyperlipidemia, peptic ulcers
and myopathy, as well as immunosuppressive effects (23). Moreover, steroids do not cure
asthma (20). Therefore, much
research has focused on identifying candidate pharmacological
agents, including natural products, which are more effective and
produce fewer unwanted side-effects than steroids (32,33). The present study suggests that
E. japonicum is a promising candidate for the treatment of
asthma, although additional studies are warranted in order to
improve our understanding of the potential effects of E.
japonicum, as well as the mechanisms repsonsible for these
effects.
References
|
1
|
World Health Organization: Asthma Fact
Sheet No307. November. 2013
|
|
2
|
United States Environmental Protection
Agency (EPA): Asthma Facts EPA-402-F-04-019. March. 2013
|
|
3
|
Slejko JF, Ghushchyan VH, Sucher B, Globe
DR, Lin SL, Globe G and Sullivan PW: Asthma control in the United
States, 2008–2010: indicators of poor asthma control. J Allergy
Clin Immunol. 133:1579–1587. 2014. View Article : Google Scholar
|
|
4
|
Kay AB: Allergy and allergic diseases.
First of two parts. N Engl J Med. 344:30–37. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
National Asthma Education and Prevention
Program: Expert Panel Report: Guidelines for the Diagnosis and
Management of Asthma Update on Selected Topics - 2002. J Allergy
Clin Immunol. 110(Suppl 5): S141–S219. 2002.
|
|
6
|
Mattes J, Yang M, Mahalingam S, Kuehr J,
Webb DC, Simson L, Hogan SP, Koskinen A, McKenzie AN, Dent LA, et
al: Intrinsic defect in T cell production of interleukin (IL)-13 in
the absence of both IL-5 and eotaxin precludes the development of
eosinophilia and airways hyperreactivity in experimental asthma. J
Exp Med. 195:1433–1444. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Manetti R, Parronchi P, Giudizi MG,
Piccinni MP, Maggi E, Trinchieri G and Romagnani S: Natural killer
cell stimulatory factor (interleukin 12 [IL-12]) induces T helper
type 1 (Th1)-specific immune responses and inhibits the development
of IL-4-producing Th cells. J Exp Med. 177:1199–1204. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Szabo SJ, Kim ST, Costa GL, Zhang X,
Fathman CG and Glimcher LH: A novel transcription factor, T-bet,
directs Th1 lineage commitment. Cell. 100:655–669. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhu J, Jankovic D, Oler AJ, Wei G, Sharma
S, Hu G, Guo L, Yagi R, Yamane H, Punkosdy G, et al: The
transcription factor T-bet is induced by multiple pathways and
prevents an endogenous Th2 cell program during Th1 cell responses.
Immunity. 37:660–673. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Brightling CE, Symon FA, Birring SS,
Bradding P, Pavord ID and Wardlaw AJ: TH2 cytokine expression in
bronchoalveolar lavage fluid T lymphocytes and bronchial submucosa
is a feature of asthma and eosinophilic bronchitis. J Allergy Clin
Immunol. 110:899–905. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mosmann TR and Coffman RL: TH1 and TH2
cells: different patterns of lymphokine secretion lead to different
functional properties. Annu Rev Immunol. 7:145–173. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yagi R, Zhu J and Paul WE: An updated view
on transcription factor GATA3-mediated regulation of Th1 and Th2
cell differentiation. Int Immunol. 23:415–420. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Barnes PJ: Immunology of asthma and
chronic obstructive pulmonary disease. Nat Rev Immunol. 8:183–192.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hershey GK: IL-13 receptors and signaling
pathways: an evolving web. J Allergy Clin Immunol. 111:677–691.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rankin JA, Picarella DE, Geba GP, Temann
UA, Prasad B, DiCosmo B, Tarallo A, Stripp B, Whitsett J and
Flavell RA: Phenotypic and physiologic characterization of
transgenic mice expressing interleukin 4 in the lung: lymphocytic
and eosinophilic inflammation without airway hyperreactivity. Proc
Natl Acad Sci USA. 93:7821–7825. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wills-Karp M, Luyimbazi J, Xu X, Schofield
B, Neben TY, Karp CL and Donaldson DD: Interleukin-13: central
mediator of allergic asthma. Science. 282:2258–2261. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhu Z, Homer RJ, Wang Z, Chen Q, Geba GP,
Wang J, Zhang Y and Elias JA: Pulmonary expression of
interleukin-13 causes inflammation, mucus hypersecretion,
subepithelial fibrosis, physiologic abnormalities, and eotaxin
production. J Clin Invest. 103:779–788. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rincon M and Irvin CG: Role of IL-6 in
asthma and other inflammatory pulmonary diseases. Int J Biol Sci.
8:1281–1290. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hamid Q, Shannon J and Martin J:
Physiologic basis of respiratory disease. Cytokines and Chemokines
in Asthma: An Overview. Tulic MK, Fiset PO, Muller Z and Hamid Q:
BC Decker Inc; Hamilton: pp. 453–467. 2005
|
|
20
|
Bosnjak B, Stelzmueller B, Erb KJ and
Epstein MM: Treatment of allergic asthma: modulation of Th2 cells
and their responses. Respir Res. 12:1142011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Barnes PJ: Current issues for establishing
inhaled corticosteroids as the antiinflammatory agents of choice in
asthma. J Allergy Clin Immunol. 101:S427–S433. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wise J: Corticosteroids for asthma may
suppress growth in children in first year of treatment, researchers
say. BMJ. 349:g46232014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ciriaco M, Ventrice P, Russo G,
Scicchitano M, Mazzitello G, Scicchitano F and Russo E:
Corticosteroid-related central nervous system side effects. J
Pharmacol Pharmacother. 4(Suppl 1): S94–S98. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kwon Soon-Kyung: Erythronium japonicum.
Yakup: http://www.yakup.com/pharmplus/plus_print.html?nid=3000131250.
Last updated March 19, 2014.
|
|
25
|
Heo BG, Park YS, Chon SU, Lee SY, Cho JY
and Gorinstein S: Antioxidant activity and cytotoxicity of methanol
extracts from aerial parts of Korean salad plants. Biofactors.
30:79–89. 2007. View Article : Google Scholar
|
|
26
|
Boerjan W, Ralph J and Baucher M: Lignin
biosynthesis. Annu Rev Plant Biol. 54:519–546. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Koriem KM and Soliman RE: Chlorogenic and
caftaric acids in liver toxicity and oxidative stress induced by
methamphetamine. J Toxicol. 2014:5834942014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Revuelta-Iniesta R and Al-Dujaili EA:
Consumption of green coffee reduces blood pressure and body
composition by influencing 11β-HSD1 enzyme activity in healthy
individuals: a pilot crossover study using green and black coffee.
BioMed Res Int. 2014:4827042014. View Article : Google Scholar
|
|
29
|
Kim HR, Lee DM, Lee SH, Seong AR, Gin DW,
Hwang JA and Park JH: Chlorogenic acid suppresses pulmonary
eosinophilia, IgE production, and Th2-type cytokine production in
an ovalbumin-induced allergic asthma: activation of STAT-6 and JNK
is inhibited by chlorogenic acid. Int Immunopharmacol.
10:1242–1248. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sy LB, Yang LK, Chiu CJ and Wu WM: The
immunoregulatory effects of caffeic acid phenethyl ester on the
cytokine secretion of peripheral blood mononuclear cells from
asthmatic children. Pediatr Neonatol. 52:327–331. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Larché M, Robinson DS and Kay AB: The role
of T lymphocytes in the pathogenesis of asthma. J Allergy Clin
Immunol. 111:450–463; quiz 464. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Seo JW1, Cho SC, Park SJ, Lee EJ, Lee JH,
Han SS, Pyo BS, Park DH and Kim BH: 1′-Acetoxychavicol acetate
isolated from Alpinia galanga ameliorates ovalbumin-induced asthma
in mice. PLosOne. 8:e564472013. View Article : Google Scholar
|
|
33
|
Bang MA, Seo JH, Seo JW, Jo GH, Jung SK,
Yu R, Park DH and Park SJ: Bacillus subtilis KCTC 11782BP-produced
alginate oligosaccharide effectively suppresses asthma via T-helper
cell type 2-related cytokines. PLoS One. 10:e01175242015.
View Article : Google Scholar : PubMed/NCBI
|