Introduction
Anterior segment dysgenesis (ASD), which is caused
by abnormal development of the anterior eye tissues, is a
collection of heterogeneous disorders which affect numerous tissues
including the iris, cornea and lens (1). It is closely linked with an
increased risk of inherited eye disorders such as congenital
corneal opacity, cataracts and glaucoma (2–4),
of which glaucoma is considered to be the second highest cause of
blindness worldwide. It has been estimated that the number of
patients with glaucoma will be 79.6 million in 2020, and this
number will increased by 31.5% compared to 2010 (5). Moreover, developmental glaucoma
accounts for approximately one-tenth of pediatric blindness
(6). Although several advanced
techniques, such as combined trabeculotomy-trabeculectomy, have
been utilized to manage developmental glaucoma, the long-term
outcomes are still not favorable (7). Thus, it is essential to find more
effective approaches, such as biological therapeutic methods, for
the treatment and prevention of glaucoma.
Of past research dedicated to uncovering the
pathogenesis of ASD, one study discovered that mutations in the
transcription factor (TF) genes such as paired box 6 (Pax6),
forkhead box C1 (Foxc1) and forkhead box E3 (Foxe3)
led to ASD (8). The TF gene
peroxidasin (Pxdn) is closely linked with the extracellular
matrix and is expressed in the corneal epithelium. Defective
Pxdn may impair the integrity of the basement membrane by
damaging the structure of its major constituent, collagen IV
(9). Previous studies have
verified that the Pxdn mutation is implicated in severe
forms of ASD, including developmental glaucoma and congenital
cataracts in human and mice (10,11). Moreover, the latter study further
investigated the gene alterations and physiological changes which
occurred after Pxdn mutation, by comparing the embryo
tissues from the KTA048 mutant line and those from the control
mouse model (11). However, the
authors emphasized different stages, and the regulatory mechanisms
of the Pxdn mutation in eye disorder-development,
particularly in relation to developmental glaucoma progression,
were not well elaborated.
Therefore, in the present study we used the
microarray dataset GSE49704, which was established by Yan et
al (11), to identify the
differentially expressed genes (DEGs) between Pxdn mutation
embryo and normal tissues. Additionally, functional enrichment
analysis and protein-protein interaction (PPI) network analysis
were also performed on these DEGs in order to explore their
biological processes (BPs) and the potential correlations. We used
the GeneCodis database to construct a transcriptional regulation
(TR) network involving TFs and miRNAs. By comparing the overlapped
genes in PPI and TR networks, crucial genes relating to the
Pxdn mutation were screened with a sub-network extracted
from the TR network. Using these bioinformatic methods, we aimed to
provide a comprehensive elucidation of the regulatory mechanisms in
Pxdn mutation-induced eye disorder development and provide
novel biomarkers which may be used for the prognosis and prevention
of these disorders.
Materials and methods
Microarray dataset
The microarray dataset with the accession number
GSE49704 was downloaded from the GEO database (Gene Expression
Omnibus; http://www.ncbi.nlm.nih.gov/geo). The mRNA expression
profile comprised 4 mouse samples from embryos with Pxdn
mutation and 4 samples of normal mouse tissues (control). The
platform used was the GPL6885 Illumina MouseRef-8 v2.0 expression
beadchip (Illumina, San Diego, CA, USA).
Data preprocessing and DEG
identification
The raw data were subjected to preprocessing, e.g.,
background correction, normalization and conversion to gene symbol
from the probe level, using the affy package of Bioconductor R, as
previously described (12). When
multiple probes corresponded to a single gene, the average level of
probes was calculated as the expression value of the specific
gene.
Subsequently, the limma (linear models for
microarray and RNA-seq data) package of Bioconductor R (http://www.bioconductor.org/packages/release/bioc/html/limma.html)
(13) was used, as previously
described, to select the DEGs between Pxdn mutation embryo
tissues and the control tissues, based on the empirical Bayes test.
The cut-off values for DEG identification were P<0.05 and |log2
(fold-change)| >2.
Functional enrichment analysis of the
DEGs
GO (gene ontology; http://www.geneontology.org/) enrichment analysis
(14) was carried out, as
previously described. Briefly, enrichment analysis was undertaken
for the screened DEGs in order to explore their potential roles in
eye disorder development resulting from Pxdn mutation, using
the DAVID (Database for Annotation, Visualization and Integration
Discovery; http://david.abcc.Ncifcrf.gov/) (15) online tool as previously described.
The over-represented terms with P-values <0.05 were considered
to indicate significantly enriched processes of the DEGs.
PPI network establishment
To further explore the potential interactions of
these DEGs on the protein level, the DEGs were mapped into STRING
(Search Tool for the Retrieval of Interacting Genes/Proteins;
http://string-db.org/), a database which provides
comprehensive data such as high throughput and genome-wide
association and the validated or predicted pairwise PPIs; we chose
the mouse as species (16). The
pairwise interactions with a combined score ≥0.4 were filtered out
to construct the PPI network, and were visualized using Cytoscape
software (http://cytoscape.org/), as previously
described (17). In the network,
a protein served as a 'node', and the 'degree' of a node, which was
calculated by topological structure analysis of the PPI network,
indicated the interaction numbers of that particular protein. The
nodes with high degrees were selected, and the encoded genes were
classified as group 1.
Constructing a TR network of the
DEGs
Considering that understanding the TR network
related to eye disorders induced by the Pxdn mutation may
contribute to elucidating the molecular mechanisms of the
pathogenesis of these disorders, it was deemed necessary to build a
TR network of the DEGs by comparing information from the GeneCodis
database (http://genecodis.cnb.csic.es), which was capable of
integrating the annotated files from various bioinformatic
analyses, and functional enrichment analyses using GO and the Kyoto
Encyclopedia of Genes and Genomes, protein domain analysis
(InterPro motifs), TF and miRNA regulation analysis. Finally, we
merged the data into a single annotation file by computing a
statistical rank score, as previously described (18). In the present study, we emphasized
the TR analysis of DEGs involving miRNAs and TFs, and the TR
network containing pairwise miRNA-DEG and TF-DEG interactions were
constructed and visualized by Cytoscape software, as previously
described (17).
In the present study, the node in the network was
referred to as a DEG, an miRNA or a TF. By applying the
aforementioned topological structure analysis to the PPI network,
the degrees of the nodes in this network were also calculated. The
crucial nodes with high degrees were screened and classified as
group 2.
Integrating the analysis of two
networks
By comparing the nodes in group 1 with those in
group 2, from the two networks, the nodes which overlapped to a
high degree were identified. Subsequently, the sub-network of the
overlapped nodes was extracted from the TR network.
Results
DEGs between Pxdn mutation embryo tissues
and the control tissues
According to the t-test method and the criteria of
P<0.05 and |log2 (fold-change)| >2, a total of 121 DEGs
between Pxdn mutation embryo tissues and the control tissues
in mice were identified, of which 75 were upregulated and 46 were
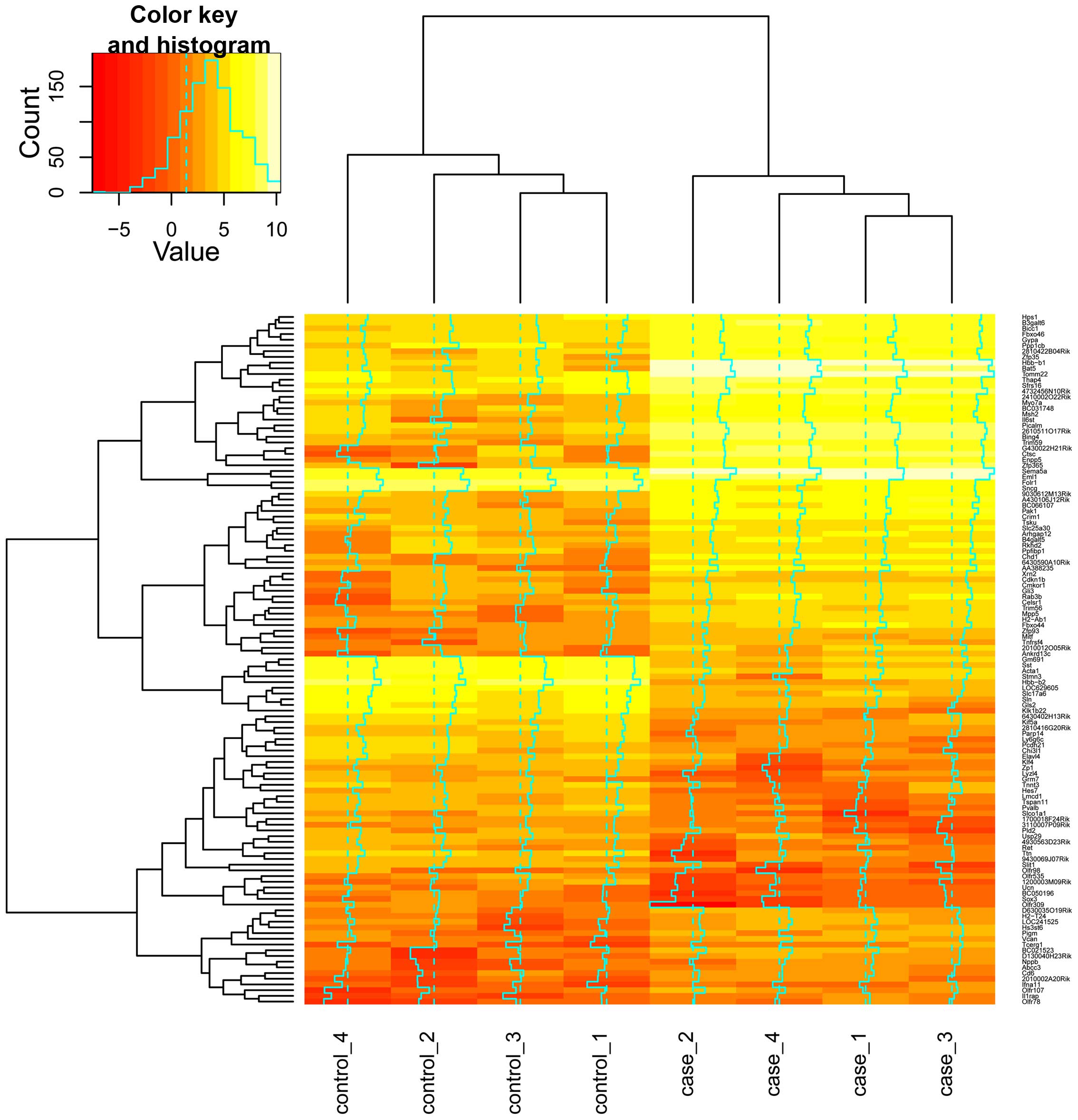
down-regulated. The heat map depicts the relative expression levels
of these DEGs (Fig. 1).
Functional enrichment using the GO
database
After taking into account the threshold for the
significant GO terms, the over-represented processes which the DEGs
may participate in were screened in the present study. As is
clearly demonstrated in Table I,
significant BP terms such as neuron development (GO: 0048666),
regulation of cellular localization (GO: 0060341), sensory organ
development (GO: 007423), polysaccharide metabolic process (GO:
0005976) as well as neuron differentiation (GO: 0030182) were
significantly enriched for the DEGs.
 | Table ISignificantly enriched biological
process terms for the identified differentially expressed genes
(ranked by P-values). |
Table I
Significantly enriched biological
process terms for the identified differentially expressed genes
(ranked by P-values).
| GO search term | P-value | Genes |
|---|
| GO: 0048666~neuron
development | 0.00416 | Sema5a,
Ret, Stmn3, Myo7a, Pak1, Gli3,
Slit1 |
| GO:
0060341~regulation of cellular localization | 0.009975 | Sncg,
Rab3b, Celsr1, Tnfrsf4, Gli3 |
| GO: 0007423~sensory
organ development | 0.010999 | Cdkn1b,
Myo7a, Mitf, Celsr1, Gli3,
Klf4 |
| GO:
0005976~polysaccharide metabolic process | 0.012321 | B3galt6,
Il6st, Chi3l1, Ppp1cb |
| GO: 0030182~neuron
differentiation | 0.017787 | Sema5a,
Ret, Stmn3, Myo7a, Pak1, Gli3,
Slit1 |
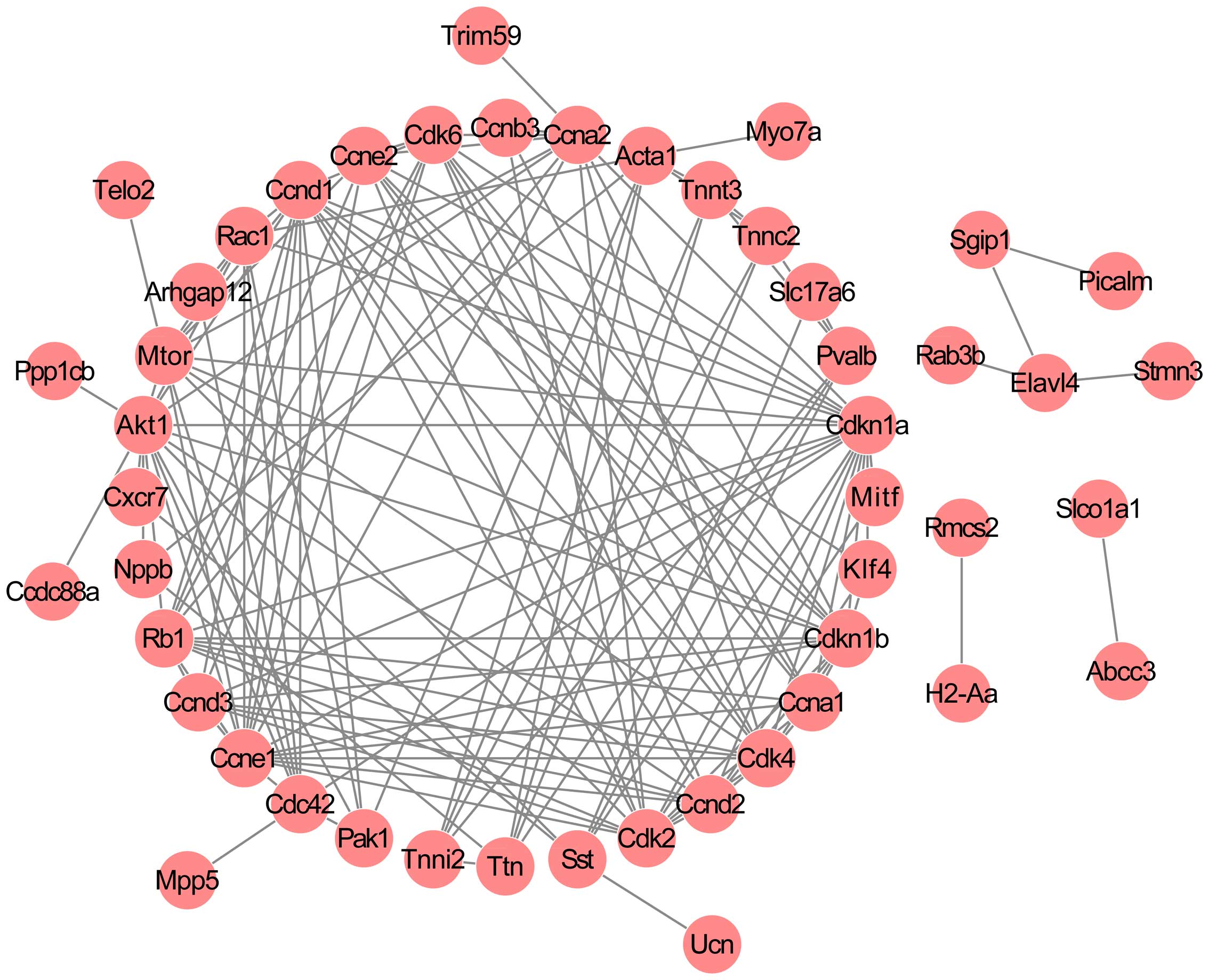
PPI network of the DEGs
By combining the information in the STRING database,
a PPI network was established, comprising 48 nodes and 148
interactions (Fig. 2). The nodes
with high degrees ≥5 were considered crucial to the network and as
a result, 25 critical nodes, including troponin C type 2 (fast)
(Tnnc2; degree = 5), troponin I type 2 (Tnni2; degree = 5),
troponin T type 3 (Tnnt3; degree = 5), P21 protein
(Cdc42/Rac)-activated kinase 1 (Pak1; degree = 5), titin (Ttn;
degree = 6), parvalbumin (Pvalb; degree = 6), somatostatin (Sst;
degree = 6), actin, alpha 1, skeletal muscle (Acta1; degree = 8),
ras-related C3 botulinum toxin substrate 1 (Rac1; degree = 9), cell
division cycle 42 (Cdc42; degree = 9), cyclin A1 (Ccna1; degree =
9), cyclin D3 (Ccnd3; degree = 9), cyclin E2 (Ccne2; degree = 10),
cyclin-dependent kinase 6 (Cdk6; degree = 11), cyclin D2 (Ccnd2;
degree = 11), mechanistic target of rapamycin (Mtor; degree = 12),
cyclin A2 (Ccna2; degree = 12), retinoblastoma 1 (Rb1; degree =
13), cyclin-dependent kinase 4 (Cdk4; degree = 13),
cyclin-dependent kinase 2 (Cdk2; degree = 14), cyclin D1 (Ccnd1;
degree = 15), cyclin-dependent kinase inhibitor 1B (Cdkn1b; degree
= 15), v-akt murine thymoma viral oncogene homolog 1 (Akt1; degree,
16), cyclin E1 (Ccne1; degree = 16), and cyclin-dependent kinase
inhibitor 1A (Cdkn1a; degree = 19) were classified as group 1.
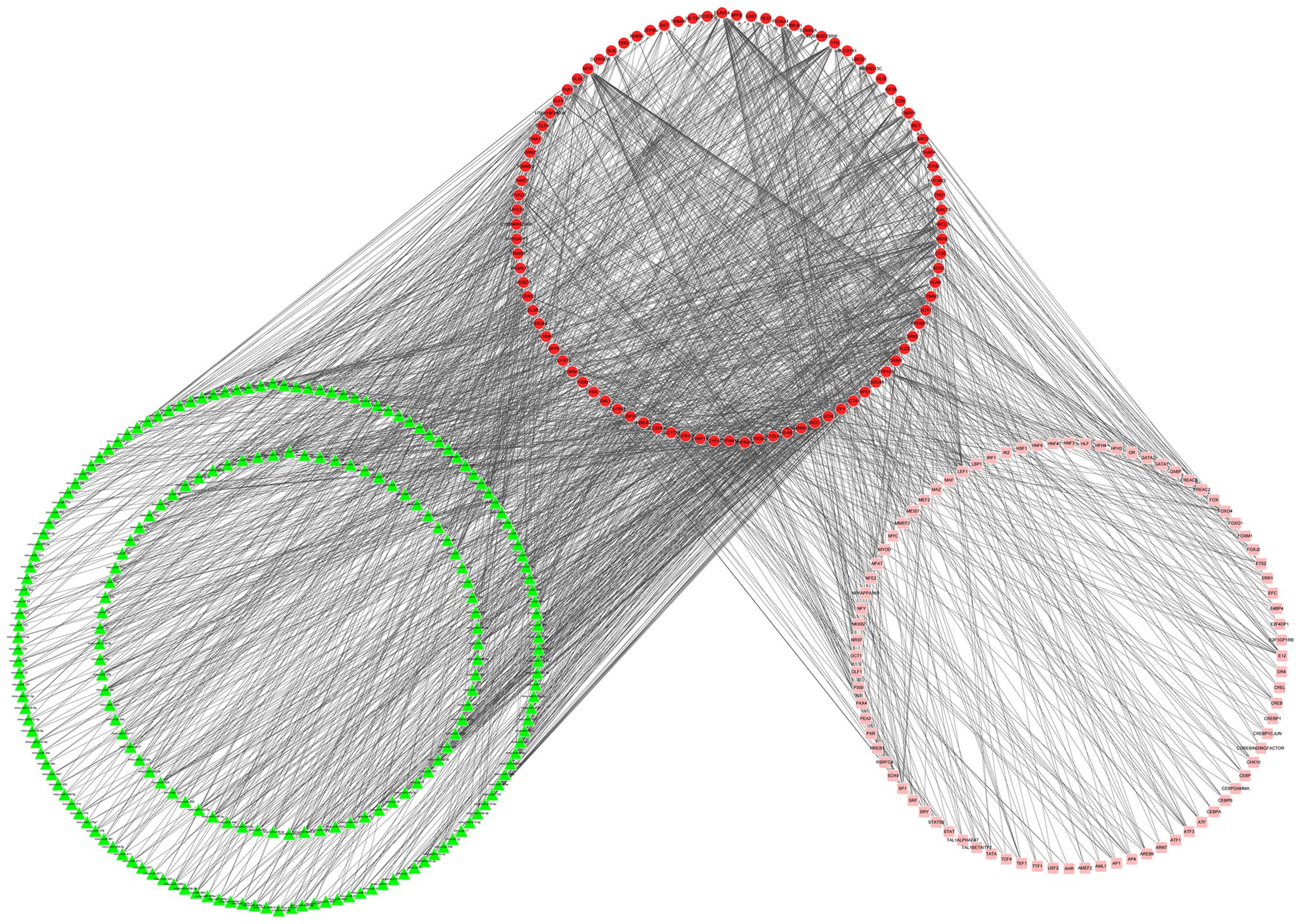
TR network of the DEGs
Using the GeneCodis database, a TR network
consisting of miRNAs and TFs of the DEGs was constructed,
encompassing 388 nodes (213 miRNAs, 91 DEGs and 84 TFs) and 1,523
pairwise interactions (Fig. 3).
In total, 120 nodes (66 DEGs, 38 miRNAs and 16 TFs) with high
degrees ≥7 were classified as group 2 nodes.
By comparing the genes in groups 1 and 2, seven
overlapped genes with high degrees were identified, namely
Cdkn1b, Acta1, Pak1, Pvalb, Sst,
Tnnt3 and Ttn. The degrees of the seven genes in the
two networks are indicated in Table
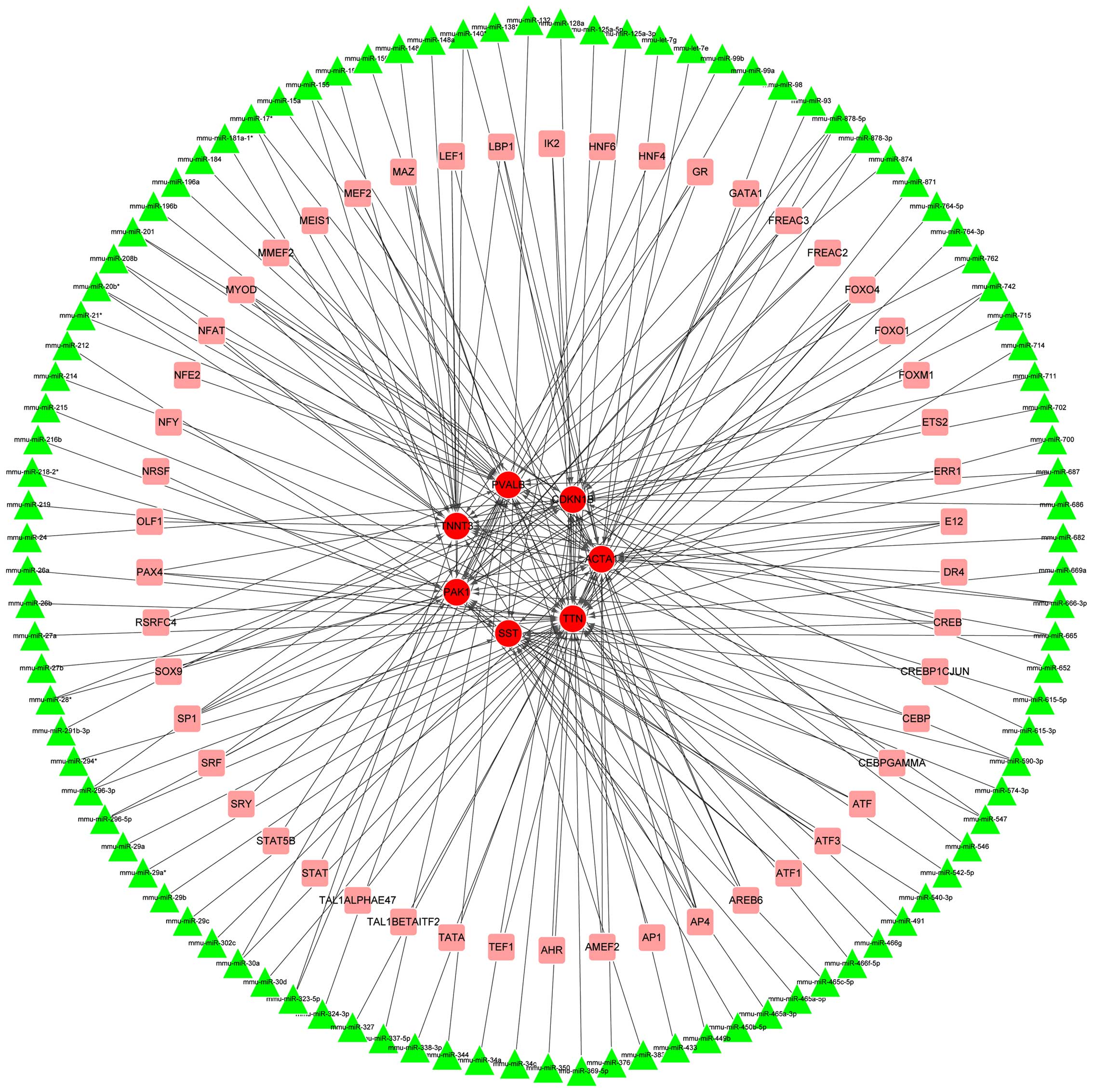
II. Subsequently, a sub-network involving these seven genes was
extracted from the TR network, comprising 156 nodes (7 DEGs, 99
miRNA and 50 TFs) and 205 interactions (Fig. 4). Notably, Cdkn1b was
predicted as the target of miRNAs such as mmu-miR-24,
mmu-miR-28*, mmu-miR-34a,
mmu-miR-369-5p and mmu-miR-449b and TFs including
forkhead box O4 (FOXO4) and activating enhancer binding protein 4
(AP4).
| Table IIDegrees of overlapped genes in both
networks. |
Table II
Degrees of overlapped genes in both
networks.
| Genes | Degree-TRN | Degree-PPI |
|---|
| Acta1 | 33 | 8 |
| Cdkn1b | 27 | 15 |
| Pak1 | 26 | 5 |
| Pvalb | 27 | 6 |
| Sst | 18 | 6 |
| Tnnt3 | 25 | 5 |
| Ttn | 49 | 6 |
Discussion
Pxdn plays an important role in basement
membrane synthesis, and its mutation is linked to ASD, which can
easily develop into developmental glaucoma and congenital cataracts
(19). In the present study, we
selected a total of 121 DEGs (75 upregulated and 46 downregulated)
between the Pxdn mutation embryo tissues and the control
tissues, which were enriched in BPs such as neuron development,
polysaccharide metabolic process and neuron differentiation. Seven
vital overlapped genes of the PPI and TR network were identified,
including Cdkn1b, Acta1 and Tnnt3, with the
sub-network extracted from the TR network. Of these, Cdkn1b
was predicted be the target of miRNAs such as mmu-miR-24,
and TFs including FOXO4 and AP4.
CDK2 and CDK4 are two cyclin-dependent kinases
(CDKs) that positively regulate cell cycle activities (20). In a previous study, it was noted
that the cyclin-dependent kinase inhibitor 1B (Cdkn1b, also
known as p27kip1) encoded protein binds to the
cyclin E-Cdk2 or cyclin D-Cdk4 complexes to prevent the activation
of the complex, and thus serves as an inhibitor of cell cycle
progression at the G1 stage. The overexpression of Cdkn1b in
Tenon's capsule fibroblasts after glaucoma filtration surgery
resulted in the downregulation of Cdk2 and Cdk4 and
the inhibition of fibroblast proliferation, which contributed to
decrease the severity of the surgical outcome (21). In addition, although Cdkn1b
was not the only regulator of Sertoli cell proliferation in adult
mouse testes, its downregulation was linked to the elevated
proliferative activity of Sertoli cells, suggesting that the
protein played a role as a suppressor in Sertoli cell proliferation
and regulation (22). These
previous studies verified that Cdkn1b played an important
role in inhibiting proliferation during cell cycle progression.
Notably, fibrovascular proliferation into the anterior segment was
considered a rare complication of acute angle-closure glaucoma
(23). Considering that the
Pxdn mutation may also influence the proliferation and
differentiation during eye development (11), and Cdkn1b was among the
downregulated DEGs in the Pxdn mutation line in our present
study, it may be inferred that Cdkn1b serves as an inhibitor
of proliferation during eye development and that the Pxdn
mutation-induced Cdkn1b decrease is related to eye disorders
such as glaucoma.
Extensive studies have been conducted which report
the transcriptional regulation of Cdkn1b by miRNAs and TFs.
miR-452 has been reported to dramatically accelerate
proliferation by targeting the 3′-untranslated region (UTR) of
Cdkn1b, which eventually contributed to tumorigenesis in
hepatocellular carcinoma (24).
miR-222 may also bond to the 3′ UTR of Cdkn1b, and it
has been suggested that it is an indicator of liver fibrosis
(25). Moreover, in human
prostate carcinoma cell lines, it was discovered that the
expression of miR-221 and miR-222 affected cell
proliferation by targeting Cdkn1b (26). As shown in Fig. 4, Cdkn1b was predicted to be
targeted by cohorts of miRNAs such as mmu-miR-24,
mmu-miR-28*, mmu-miR-34a,
mmu-miR-369-5p and mmu-miR-449b. It has previously
been noted that miR-24 inhibits cell cycle progression and
directly suppresses the transcription of v-myc avian
myelocytomatosis viral oncogene homolog (MYC), an important TF in
cell cycle progression which regulates the transcription of
Cdkn1b (27). Although no
further direct evidence exists to illustrate the regulatory
relationship between Cdkn1b and miR-24, this study
may provide a clue for the potential targeting relationship between
these molecules in eye disorders development. However, further
experimental validation is warranted.
With regard to the TFs that targeted Cdkn1b,
it has been reported that the binding of the TF PAX6 to multiple
regulators which are responsible for cell cycle progression and
proliferation, including Cdkn1b, may account for the
regulation of cell proliferation and determination in the cortex
(28). Moreover, it has been
noted that Cdkn1b is directly regulated by the TF E2F1 and
suppresses baculoviral IAP repeat containing 5 (Birc5)
expression via the inhibition of CDK activity, which results in
cell cycle arrest. The induction of Cdkn1b by E2F1 enabled
E2F1 to inhibit the expression of Birc5 in neuroblastomas
(29). In the present study,
Cdkn1b was linked to TFs including FOXO4 and AP4. FOXO4 is a
TF that is involved in the regulation of the insulin signaling
pathway (30). The peptidylprolyl
cis/trans isomerase, NIMA-interacting 1 (Pin1) encoded
protein acts as a catalyst in the regulation of
post-phosphorylation conformation of its substrates by binding to
the phosphorylated ser/thr-pro motifs (31). In relation to tumor development,
it has been indicated previously that Pin1-induced
deubiquitylation results in the inhibition of the nuclear
translocation of FOXO4 and the suppression of FOXO4 targeting
Cdkn1b (32). The TF AP4
has been noted to suppress the transcription of Cdkn1a,
another CDK inhibitor, by occupying four CAGCTG motifs in its
promoter. By binding to specific motifs of CACGTG in the first
intron of AP4, c-MYC directly regulates AP4
expression and thus contributes to maintaining the proliferative
stage in breast cancer cells (33). This evidence substantiates our
hypothesis that Cdkn1b is the target of FOXO4 and AP4 and
that transcriptional regulation by the TFs plays a significant role
in eye disorder development caused by the Pxdn mutation.
Of the remaining six vital genes identified in the
present study, Acta1, which encodes alpha actin family
protein, has previously been listed in the DEGs between retinal
ganglion cells (RGCs) and glial cells (34). Given that the loss of the RGCs
through apoptosis contributes to the progression of glaucoma
(35), we speculate that
Acta1 is also linked to eye disorder development,
particularly glaucoma progression. Tnnt3 has been identified
as a DEG between human trabecular meshwork in patients with primary
open-angle glaucoma and normal subjects (36), and in this study it was also
suggested that it is involved in eye disorder regulation. Although
there is not yet sufficient evidence to fully identify correlations
between these genes and eye disorder development, or the regulatory
relationships between these genes and TFs and miRNAs, all the
predictions described in the present study provide a novel insight
into the transcriptional regulators in eye disorders such as
glaucoma, which are induced by the Pxdn mutation, and this
link should be borne in mind for follow-up studies.
In conclusion, using the GSE49704 dataset, in the
Pxdn mutation embryo tissues of the mice, we identified
several crucial genes, including Cdkn1b, Acta1 and
Tnnt3, which were related to the progression of eye
disorders, such as glaucoma. Notably, it may be the case that
Cdkn1b exerts its roles by inhibiting proliferation and is
mediated by mmu-miR-24 and targeted by the TFs of FOXO4 and
AP4, during the regulation of eye disorder development. However,
further experimental validationis warranted.
Acknowledgments
The present study was supported by the Hubei
Provincial Natural Science Foundation of China (no.
2014CFB366).
References
|
1
|
Gould DB and John SW: Anterior segment
dysgenesis and the developmental glaucomas are complex traits. Hum
Mol Genet. 11:1185–1193. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nischal KK: Congenital corneal opacities -
a surgical approach to nomenclature and classification. Eye (Lond).
21:1326–1337. 2007. View Article : Google Scholar
|
|
3
|
Van Agtmael T, Schlötzer-Schrehardt U,
McKie L, Brownstein DG, Lee AW, Cross SH, Sado Y, Mullins JJ,
Pöschl E and Jackson IJ: Dominant mutations of Col4a1 result in
basement membrane defects which lead to anterior segment dysgenesis
and glomerulopathy. Hum Mol Genet. 14:3161–3168. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shigeyasu C, Yamada M, Mizuno Y, Yokoi T,
Nishina S and Azuma N: Clinical features of anterior segment
dysgenesis associated with congenital corneal opacities. Cornea.
31:293–298. 2012. View Article : Google Scholar
|
|
5
|
Quigley HA and Broman AT: The number of
people with glaucoma worldwide in 2010 and 2020. Br J Ophthalmol.
90:262–267. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yalvac IS, Satana B, Suveren A, Eksioglu U
and Duman S: Success of trabeculotomy in patients with congenital
glaucoma operated on within 3 months of birth. Eye (Lond).
21:459–464. 2007.
|
|
7
|
Mandal AK, Gothwal VK and Nutheti R:
Surgical outcome of primary developmental glaucoma: a single
surgeon's long-term experience from a tertiary eye care centre in
India. Eye (Lond). 21:764–774. 2007. View Article : Google Scholar
|
|
8
|
Reis LM and Semina EV: Genetics of
anterior segment dysgenesis disorders. Curr Opin Ophthalmol.
22:314–324. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Choi A, Lao R, Ling-Fung Tang P, Wan E,
Mayer W, Bardakjian T, Shaw GM, Kwok PY, Schneider A and Slavotinek
A: Novel mutations in PXDN cause microphthalmia and anterior
segment dysgenesis. Eur J Hum Genet. 23:337–341. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Khan K, Rudkin A, Parry DA, Burdon KP,
McKibbin M, Logan CV, Abdelhamed ZI, Muecke JS, Fernandez-Fuentes
N, Laurie KJ, et al: Homozygous mutations in PXDN cause congenital
cataract, corneal opacity, and developmental glaucoma. Am J Hum
Genet. 89:464–473. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yan X, Sabrautzki S, Horsch M, Fuchs H,
Gailus-Durner V, Beckers J, Hrabě de Angelis M and Graw J:
Peroxidasin is essential for eye development in the mouse. Hum Mol
Genet. 23:5597–5614. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gautier L, Cope L, Bolstad BM and Irizarry
RA: affy - analysis of Affymetrix GeneChip data at the probe level.
Bioinformatics. 20:307–315. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Smyth GK: Limma: linear models for
microarray data. Bioinformatics and computational biology solutions
using R and Bioconductor. Springer; New York, NY: pp. 397–420.
2005, View Article : Google Scholar
|
|
14
|
Blake JA, Dolan M, Drabkin H, Hill DP, Li
N, Sitnikov D, Bridges S, Burgess S, Buza T, McCarthy F, et al:
Gene Ontology Consortium: Gene Ontology annotations and resources.
Nucleic Acids Res. 41(D1): D530–D535. 2013. View Article : Google Scholar
|
|
15
|
Dennis G Jr, Sherman BT, Hosack DA, Yang
J, Gao W, Lane HC and Lempicki RA: DAVID: Database for annotation,
visualization, and integrated discovery. Genome Biol. 4:32003.
View Article : Google Scholar
|
|
16
|
Jensen LJ, Kuhn M, Stark M, Chaffron S,
Creevey C, Muller J, Doerks T, Julien P, Roth A, Simonovic M, et
al: STRING 8 - a global view on proteins and their functional
interactions in 630 organisms. Nucleic Acids Res. 37(Database):
D412–D416. 2009. View Article : Google Scholar
|
|
17
|
Kohl M, Wiese S and Warscheid B:
Cytoscape: software for visualization and analysis of biological
networks. Methods Mol Biol. 291–303. 2011. View Article : Google Scholar
|
|
18
|
Carmona-Saez P, Chagoyen M, Tirado F,
Carazo JM and Pascual-Montano A: GENECODIS: a web-based tool for
finding significant concurrent annotations in gene lists. Genome
Biol. 8:R32007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Péterfi Z and Geiszt M: Peroxidasins:
novel players in tissue genesis. Trends Biochem Sci. 39:305–307.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lapenna S and Giordano A: Cell cycle
kinases as therapeutic targets for cancer. Nat Rev Drug Discov.
7:547–566. 2009. View
Article : Google Scholar
|
|
21
|
Yang JG, Deng Y, Zhou LX, Li XY, Sun PR
and Sun NX: Overexpression of CDKN1B inhibits fibroblast
proliferation in a rabbit model of experimental glaucoma filtration
surgery. Invest Ophthalmol Vis Sci. 54:343–352. 2013. View Article : Google Scholar
|
|
22
|
Ahmed EA, Barten-van Rijbroek AD, Kal HB,
Sadri-Ardekani H, Mizrak SC, van Pelt AM and de Rooij DG:
Proliferative activity in vitro and DNA repair indicate that adult
mouse and human Sertoli cells are not terminally differentiated,
quiescent cells. Biol Reprod. 80:1084–1091. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ng J, Srinivasan S and Roberts F: Fibrous
proliferation into anterior segment after acute angle-closure
glaucoma. Cornea. 34:103–106. 2015. View Article : Google Scholar
|
|
24
|
Zheng Q, Sheng Q, Jiang C, Shu J, Chen J,
Nie Z, Lv Z and Zhang Y: MicroRNA-452 promotes tumorigenesis in
hepatocellular carcinoma by targeting cyclin-dependent kinase
inhibitor 1B. Mol Cell Biochem. 389:187–195. 2014.w. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ogawa T, Enomoto M, Fujii H, Sekiya Y,
Yoshizato K, Ikeda K and Kawada N: MicroRNA-221/222 upregulation
indicates the activation of stellate cells and the progression of
liver fibrosis. Gut. 61:1600–1609. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Galardi S, Mercatelli N, Giorda E,
Massalini S, Frajese GV, Ciafrè SA and Farace MG: miR-221 and
miR-222 expression affects the proliferation potential of human
prostate carcinoma cell lines by targeting p27Kip1. J Biol Chem.
282:23716–23724. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lal A, Navarro F, Maher CA, Maliszewski
LE, Yan N, O'Day E, Chowdhury D, Dykxhoorn DM, Tsai P, Hofmann O,
et al: miR-24 inhibits cell proliferation by targeting E2F2, MYC,
and other cell-cycle genes via binding to 'seedless̓ 3′UTR microRNA
recognition elements. Mol Cell. 35:610–625. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sansom SN, Griffiths DS, Faedo A, Kleinjan
D-J, Ruan Y, Smith J, van Heyningen V, Rubenstein JL and Livesey
FJ: The level of the transcription factor Pax6 is essential for
controlling the balance between neural stem cell self-renewal and
neurogenesis. PLoS Genet. 5:e10005112009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Eckerle I, Muth D, Batzler J, Henrich KO,
Lutz W, Fischer M, Witt O, Schwab M and Westermann F: Regulation of
BIRC5 and its isoform BIRC5-2B in neuroblastoma. Cancer Lett.
285:99–107. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jünger MA, Rintelen F, Stocker H,
Wasserman JD, Végh M, Radimerski T, Greenberg ME and Hafen E: The
Drosophila forkhead transcription factor FOXO mediates the
reduction in cell number associated with reduced insulin signaling.
J Biol. 2:202003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lu KP and Zhou XZ: The prolyl isomerase
PIN1: a pivotal new twist in phosphorylation signalling and
disease. Nat Rev Mol Cell Biol. 8:904–916. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lu Z and Hunter T: Prolyl isomerase Pin1
in cancer. Cell Res. 24:1033–1049. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jung P, Menssen A, Mayr D and Hermeking H:
AP4 encodes a c-MYC-inducible repressor of p21. Proc Natl Acad Sci
USA. 105:15046–15051. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tezel G and Yang X: Comparative gene array
analysis of TNF-α-induced MAPK and NF-kappaB signaling pathways
between retinal ganglion cells and glial cells. Exp Eye Res.
81:207–217. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kamphuis W, Dijk F, Kraan W and Bergen AA:
Transfer of lens-specific transcripts to retinal RNA samples may
underlie observed changes in crystallin-gene transcript levels
after ischemia. Mol Vis. 13:220–228. 2007.PubMed/NCBI
|
|
36
|
Liu Y, Allingham RR, Qin X, Layfield D,
Dellinger AE, Gibson J, Wheeler J, Ashley-Koch AE, Stamer WD and
Hauser MA: Gene expression profile in human trabecular meshwork
from patients with primary open-angle glaucoma. Invest Ophthalmol
Vis Sci. 54:6382–6389. 2013. View Article : Google Scholar : PubMed/NCBI
|