Introduction
Malignant gliomas are the most common and deadly
primary brain tumors of the central nervous system (1). Glioblastoma multiforme (GBM) is the
most malignant and common type of glioma, which arises from
astrocytes with poor differentiation and is associated with a
median survival of approximately 10–14 months (2,3).
Rapid proliferation and diffuse brain invasion are the
histopathological hallmarks of these tumors and are likely to
determine the unfavorable prognosis (4). Therefore, the recurrence of gliomas
remains inevitable. Novel therapeutic targets, based on the
identification of molecular events key to carcinogenesis and tumor
progression, are necessary in order to improve the overall outcome
of patients with GBM.
MicroRNAs (miRNAs or miRs) are a class of
non-coding, regulatory RNA molecules, 21–24 nucleotides (nt) in
length, that modulate the expression levels of specific proteins
based on sequence complementarity with their target mRNA molecules
(5). Accumulating evidence
indicates that miRNAs are abnormally expressed in various types of
cancer and act as both tumor suppressor genes and oncogenes by
negatively regulating their mRNA targets either by degradation or
translational repression (6–8).
Therefore, it has been proposed that miRNAs may serve as novel
targets for anticancer therapies.
It has been suggested that miR-130b is a novel
tumor-related miRNA and it has been found to be significantly
dysregulated in some types of cancer; this includes overexpression
in melanoma (9), gastric cancer
(10), bladder cancer (11), colorectal cancer (12), metastatic renal carcinoma
(13) and glioma (14), as well as downregulation in
papillary thyroid carcinomas (15), endometrial cancer (16), pituitary adenomas (17) and pancreatic cancer (18). Therefore, the functional
significance of miR-130b in cancer development and progression
appears to be specific to the type of cancer. Malzkorn et al
identified the increased expression of miR-130b in four patients
with WHO grade II primary gliomas that spontaneously progressed to
WHO grade IV secondary glioblastomas (14). However, the precise role that
miR-130b plays in the proliferation, differentiation and migration
of glioma cells remains unknown. In this study, we observed that
miR-130b expression was elevated in glioma tissues and cells.
Moreover, we examined how miR-130b affects the proliferation and
invasion of glioma cells as well as the mechanism responsible for
the miRNA-mediated direct suppression of the peroxisome
proliferator-activated receptor-γ (PPAR-γ) pathway in gliomas.
Materials and methods
Clinical samples and cell lines
Human glioma tumor tissue samples were obtained from
patients undergoing surgical resection at the Department of
Neurosurgery at Fuzhou General Hospital (Xiamen University Medical
College, Fuzhou, China) in accordance with procedures approved by
the Ethics Committee of our hospital and the study was performed
according to the Declaration of Helsinki. Written informed consent
was obtained from the family of each patient. Twelve glioma samples
were thoroughly reviewed by an experienced neuropathologist
according to the 2007 WHO classification, which classifies
astrocytomas as i) well-differentiated low-grade diffuse
astrocytoma (WHO grade II, 4 cases), ii) anaplastic astrocytoma
(WHO grade III, 4 cases) and iii) glioblastoma multiforme (GBM; WHO
grade IV, 4 cases). In addition, four non-neoplastic brain
specimens were obtained from patients with traumatic brain injury
at the site of decompression. All tissue samples were frozen in
liquid nitrogen immediately after resection and were stored at
−80°C. Normal human astrocytes (NHAs) were purchased from ScienCell
Research Laboratories (Carlsbad, CA, USA) in 2013. The glioma cell
lines U251, U87, SNB19 and LN229 that were used in this study were
obtained from the Institute of Biochemistry and Cell Biology
(Shanghai Institutes for Biological Sciences, Chinese Academy of
Science, Shanghai, China).
Cell culture and transfection
The cells were grown in Dulbecco's modified Eagle's
medium (DMEM; Invitrogen, Carlsbad, CA, USA) supplemented with 10%
fetal bovine serum (FBS; Invitrogen) and 1% penicillin/streptomycin
(Invitrogen/Thermo Fisher Scientific Inc., Waltham, MA, USA) in 5%
CO2 atmosphere at 37°C. The U251 cells
(1×105) were seeded into 6-well plates and transfected
with either the negative control (NC), miR-130b mimic, miR-130b
inhibitor or siPPAR-γ (5′-AAUAUGACCUGAAGCUCCAAGAAUAAG-3′) which
were purchased from GenePharma (Shanghai, China), using
Lipofectamine 2000 (Invitrogen) according to the manufacturer's
instructions. Following a 24 h transfection, the media were removed
and the cells were placed in complete medium and maintained at 37°C
in an atmosphere of 5% CO2.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the cultured cells and
fresh glioma tissues using TRIzol reagent (Invitrogen) and total
miRNAs were extracted using mirVana kits (Ambion, Austin, TX, USA)
according to the manufacturer's instructions. Gene-specific primers
were used to synthesize miR-130b cDNA from total RNA, according to
the miRNA-specific TaqMan miRNA assay kit (Applied Biosystems,
Foster City, CA, USA); U6 snRNA was used as an internal control.
The expression levels of miR-130b and PPAR-γ were examined by
performing qPCR with an SYBR-Green PCR Master Mix kit in
conjunction with an ABI PRISM 7300 system (both from Applied
Biosystems). The primer sequences were as follows: PPAR-γ forward,
5′-CATGCTTGTGAAGGATGCAAG-3′ and reverse,
5′-CCCATCATTAAGGAATTCATGTC-3′; GAPDH forward,
5′-TCGGAGTCAACGGATTTGG-3′ and reverse,
5′-CATGGGTGGAATCATATTGGA-3′.
Western blot analysis
Total cell lysates from different experiments were
obtained by lysing the cells in RIPA buffer. The protein
concentration was determined using a BCA Protein assay kit (Pierce
Biotechnology, Rockford, IL, USA). Forty micrograms of protein from
each sample were resolved by 10% SDS-PAGE gel and transferred to
PVDF membranes (Millipore, Billerica, MA, USA). The membranes were
further incubated with primary antibodies PPAR-γ (1:200 dilution),
E-cadherin (1:500 dilution) and β-catenin (1:200 dilution) (all
from Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) followed
by incubation with an HRP-conjugated secondary antibody (1:1,000
dilution; Zymed, San Diego, CA, USA). The membranes were stripped
and reprobed with a primary antibody against GAPDH. The signal
intensity was determined using gel analysis software, ImageJ. GAPDH
was used as an endogenous protein for normalization (1:1,000
dilution; Santa Cruz Biotechnology, Inc.).
Dual luciferase reporter assay
The 3′-untranslated region (3′-UTR) sequence of
PPAR-γ predicted to interact with miR-130b was identified using
TargetScan (http://www.targetscan.org) as well as
a mutated sequence with the predicted target sites. They were
synthesized and inserted into the XbaI and FseI sites
of a pGL3 control vector (Promega Corp., Madison, WI, USA). The
cells (8×103) were seeded into a 24-well plate. The
cells in each well were co-transfected with either the miR-130b
mimic, miR-130b inhibitor or NC. The transfections were performed
using the FuGENE HD transfection reagent (Promega Corp.). A
Renilla luciferase vector pRL-SV50 (Promega Corp.) was
co-transfected to normalize the differences in transfection
efficiency. Following a 24 h transfection, the cells were harvested
and assayed using the Dual-Luciferase Reporter assay system
(Promega Corp.) according to the manufacturer's instructions.
Transfection was repeated in triplicate in three independent
experiments.
Proliferation assays
The cells were plated at 5×103 cells/well
in 96-well plates and were grown for 24, 48 and 72 h after
transfection, according to the manufacturer's instructions. Cell
proliferation was documented every 24 h for four days using a cell
counting kit-8 (CCK8) assay (Dojindo, Tokyo, Japan). The absorbance
at a wavelength of 570 nm was detected using a microplate reader
(Thermo Fisher Scientific Inc.).
Clonogenicity assay
Transfected glioma U251 cells (1×103)
were seeded into 6-well plates and cultured in cell culture medium
for 2 weeks to allow colony formation. The culture medium was
changed every third day. The colonies were then fixed in 100%
methanol for 5 min and stained with 1.0% crystal violet solution
(BioLab, Inc., Shanghai, China) for 30 sec. The number of
macroscopically detectable colonies was registered.
Cell cycle assay
The cells were harvested by trypsinization 48 h
after transfection. They were washed three times with ice-cold
phosphate-buffered saline (PBS) and fixed with 70% ethanol
overnight at 4°C. The fixed cells were rehydrated in PBS and
subjected to propidium iodide/RNase staining followed by
fluorescence-activated cell sorting (FACS; Becton-Dickinson,
Mountain View, CA, USA). The percentage of cells in each phase of
the cell cycle was estimated using PV Elite software (Intergraph
Corp., Madison, AL, USA).
Transwell assay and scratch-wound
assay
Cell invasion and migration was examined using
Transwell and scratch-wound assays, respectively. For the Transwell
assay, the appropriate oligonucleotides were transfected into the
cells as described above. Following incubation for 48 h,
3×104 cells were transferred to the top of the
Matrigel-coated invasion chambers (BD Biosciences, San Jose, CA,
USA) in serum-free DMEM. DMEM containing 10% FBS was added to the
lower chamber. After 24 h, the non-invading cells were removed, and
the invading cells were fixed using 95% ethanol, stained with 0.1%
crystal violet and images were captured at ×100 magnification under
an inverted phase contrast microscope (Olympus CKX31/41; Olympus,
Tokyo, Japan). The experiments were repeated three times
independently. For the scratch-wound assay, the miR-130b mimic,
miR-130b inhibitor or NC, were transfected into the cells in 6-well
plates. The cell layers were then scratched using a 200 µl
sterile pipette tip to form wound gaps. The wound location in the
6-well plates was marked. Images of the cells were captured to
record the wound width at the marked wound locations at 0 and 24 h
in order to measure the migratory ability of the cells.
Statistical analysis
All data are presented as the means ± SD. The
experiments were repeated three times. All statistical analyses
were performed using a two-tailed Student's t-test in SPSS 12.0
software. A P-value of <0.05 was considered to indicate a
statistically significant difference.
Results
miR-130b is overexpressed in glioma
tissues and cell lines
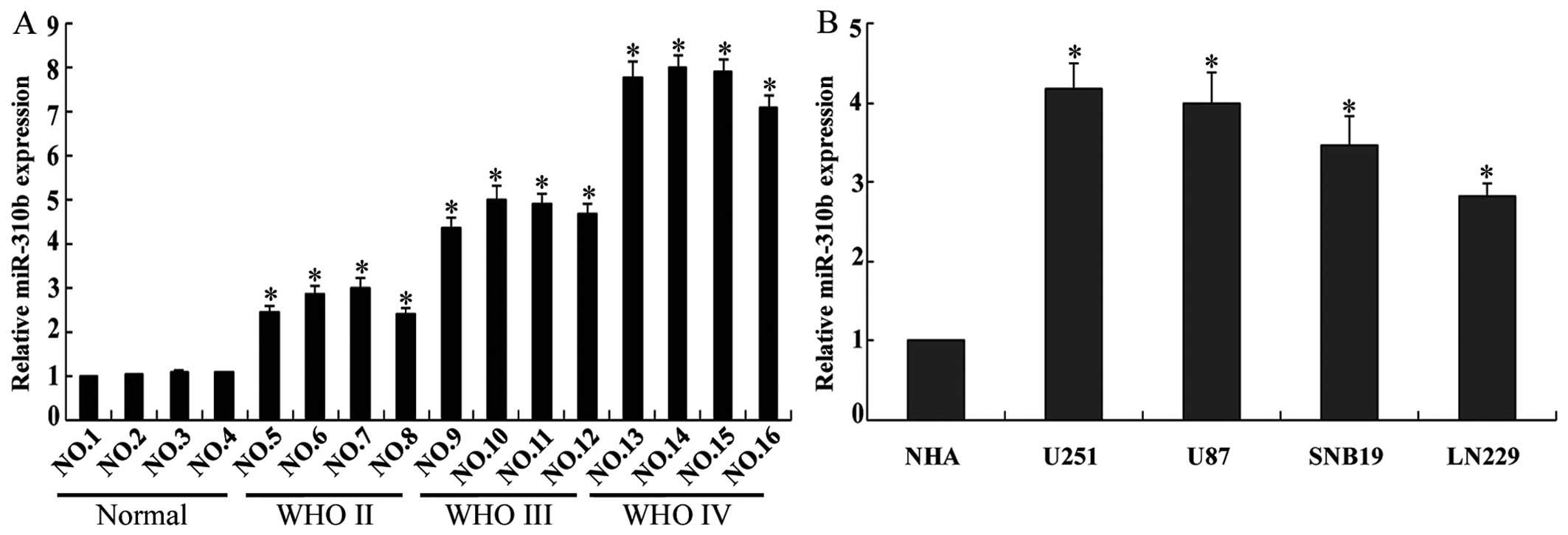
The expression of miR-310b in 12 glioma tissues and
4 non-neoplastic brain specimens was examined by RT-qPCR. The
expression of miR-310b was significantly higher in the glioma
tissues in comparison with that in the 4 non-neoplastic brain
specimens, particularly in the grade III/IV tissues (Fig. 1A). Compared with miR-310b
expression in the NHAs, it was higher in the glioma cell lines
U251, U87, SNB19 and LN229 (P<0.05) (Fig. 1B). Taken together, these findings
indicate that miR-310b is overexpressed in glioma tissues and
glioma cell lines.
miR-310b directly targets PPAR-γ in
glioma cells
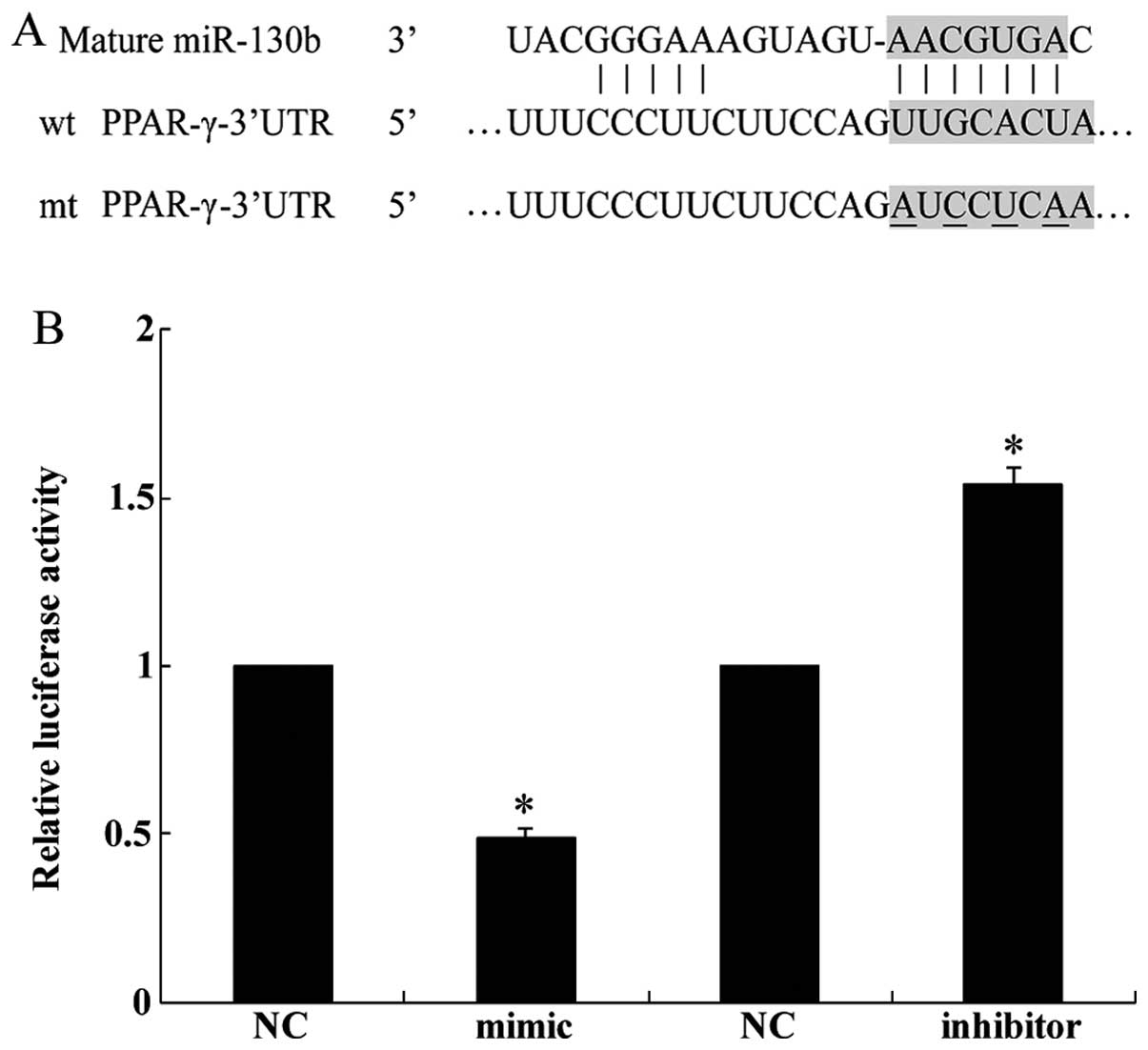
We identified the tumor suppressor gene PPAR-γ as a
potential target of miR-130b by using the publicly available
algorithm TargetScan (http://www.targetscan.org) (Fig. 2A). A dual luciferase reporter
assay was performed in order to verify that the 3′-UTR of PPAR-γ
mRNA is a direct target of miR-130b. The luciferase activity of the
U251 cells transfected with miR-130b mimic was reduced when
compared with the cells transfected with NC whereas miR-130b
inhibitor increased the luciferase activity of the U251 cells
(Fig. 2B). Collectively, this
experiment demonstrated that PPAR-γ is a direct target of
miR-310b.
miR-130b regulates the expression of
PPAR-γ, E-cadherin and β-catenin
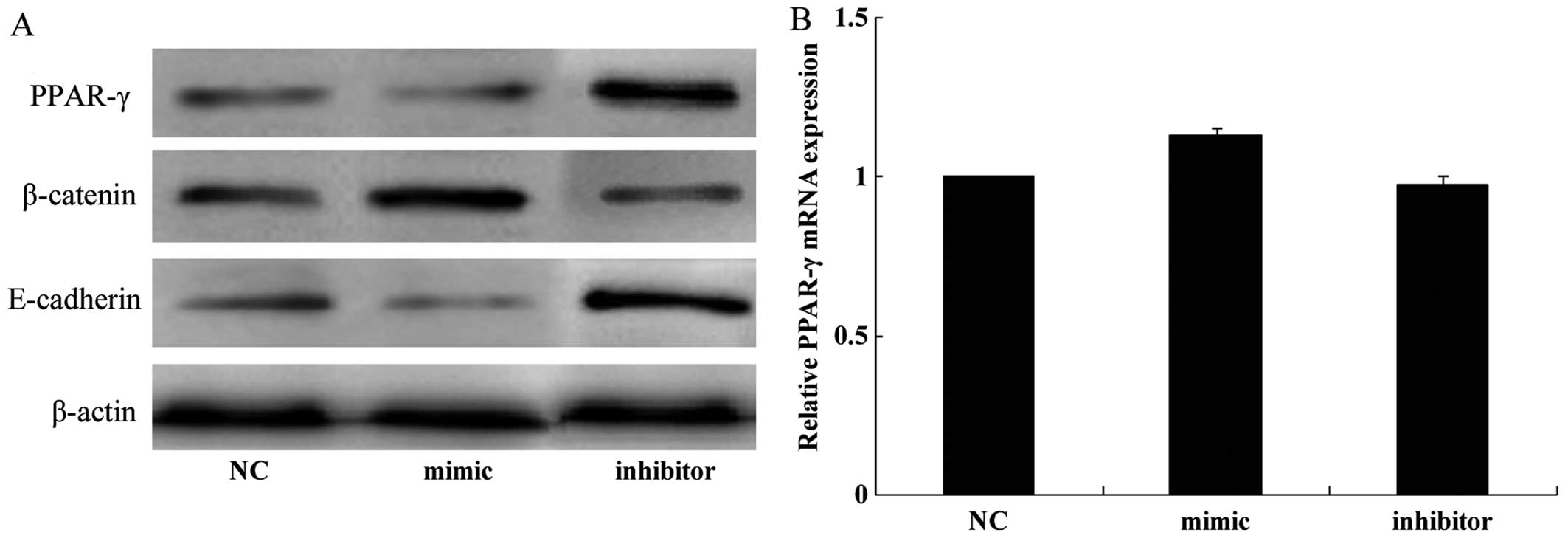
We performed western blot analysis and RT-qPCR in
order to elucidate the possible molecular mechanism through which
miR-130b exerts effects in glioma cells. The results of western
blot analysis revealed that the transfection of U251 cells with the
miR-130b mimic significantly reduced the expression of PPAR-γ and
E-cadherin whereas it increased the expression of β-catenin
(Fig. 3A). Conversely, the
protein expression levels of PPAR-γ and E-cadherin were increased
whereas those of β-catenin were decreased when endogenous miR-130b
was downregulated by the miR-130b inhibitor (Fig. 3A). However, the results of RT-qPCR
revealed that the mRNA expression of PPAR-γ was unaffected by the
miR-130b mimic and inhibitor (Fig.
3B). These results demonstrate that the overexpression of
miR-130b negatively regulates the protein levels but not the mRNA
levels of PPAR-γ, indicating that regulation occurs at the
translational level.
miR-130b regulates the proliferation of
U251 cells
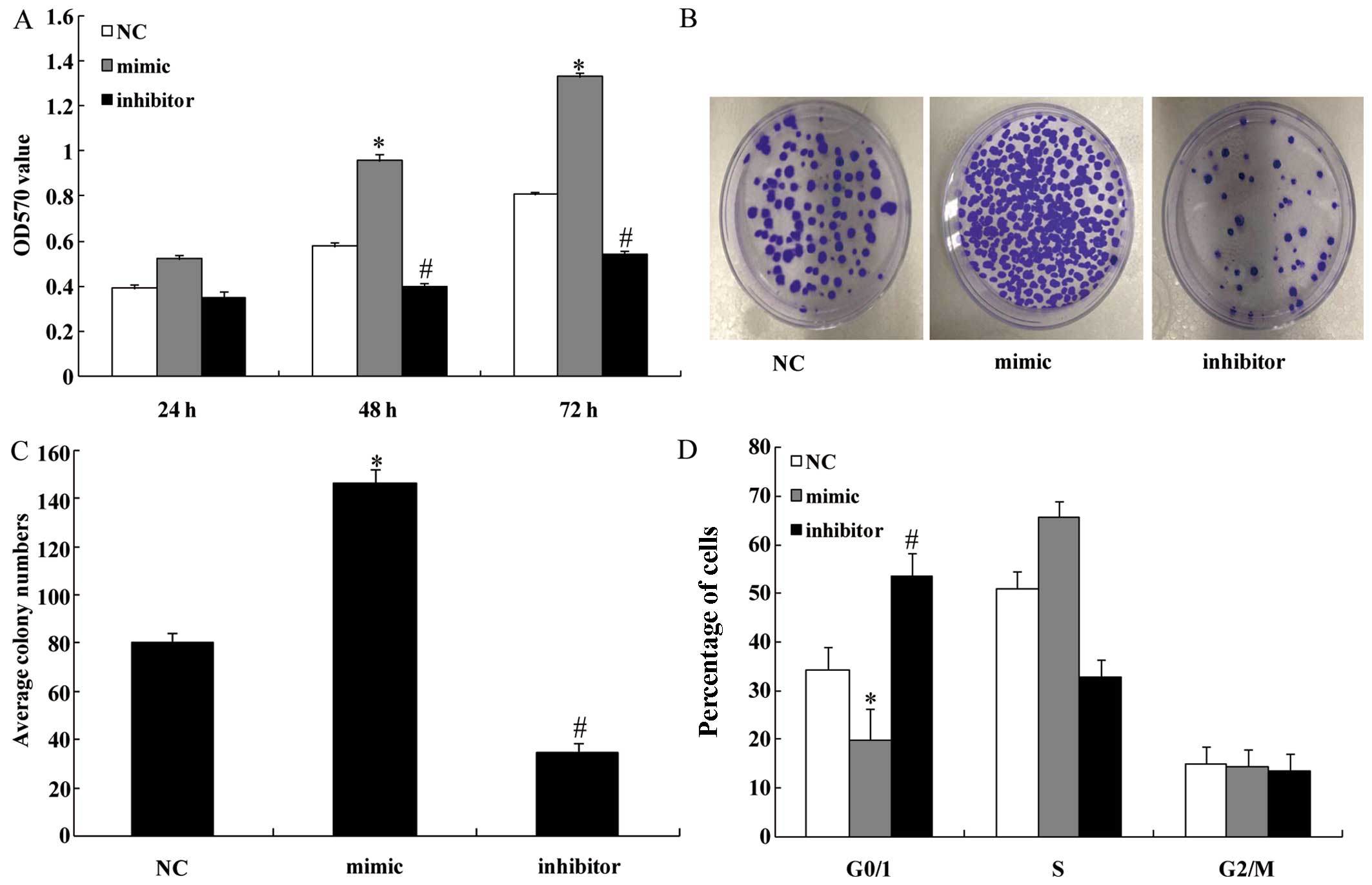
We performed the cell proliferation assay in order
to explore the effect of miR-130b upregulation and downregulation
on glioma cell proliferation in vitro. The cell
proliferation rate was clearly increased in the miR-130b
mimic-transfected group at 48 and 72 h compared with the NC groups
(P<0.05) (Fig. 4A). By
contrast, the cell proliferation rate was inhibited following
transfection with the miR-130b inhibitor (P<0.05) (Fig. 4A). In Fig. 4B and C, stable overexpression of
miR-130b markedly increased the number of surviving colonies,
whereas the miR-130b inhibitor reduced the number of surviving
colonies formed by U251 cells compared with the NC groups
(P<0.05). This finding indicates that miR-130b significantly
enhances the proliferation of glioma cells. As shown in Fig. 4D, FACS revealed that miR-130b
overexpression increased the percentage of cells in the S phase and
significantly decreased the percentage of cells in the G1/G0
(P<0.05). By contrast, the miR-130b inhibitor decreased the
percentage of cells in the S phase and significantly increased the
percentage of cells in the G1/G0 phase (P<0.05).
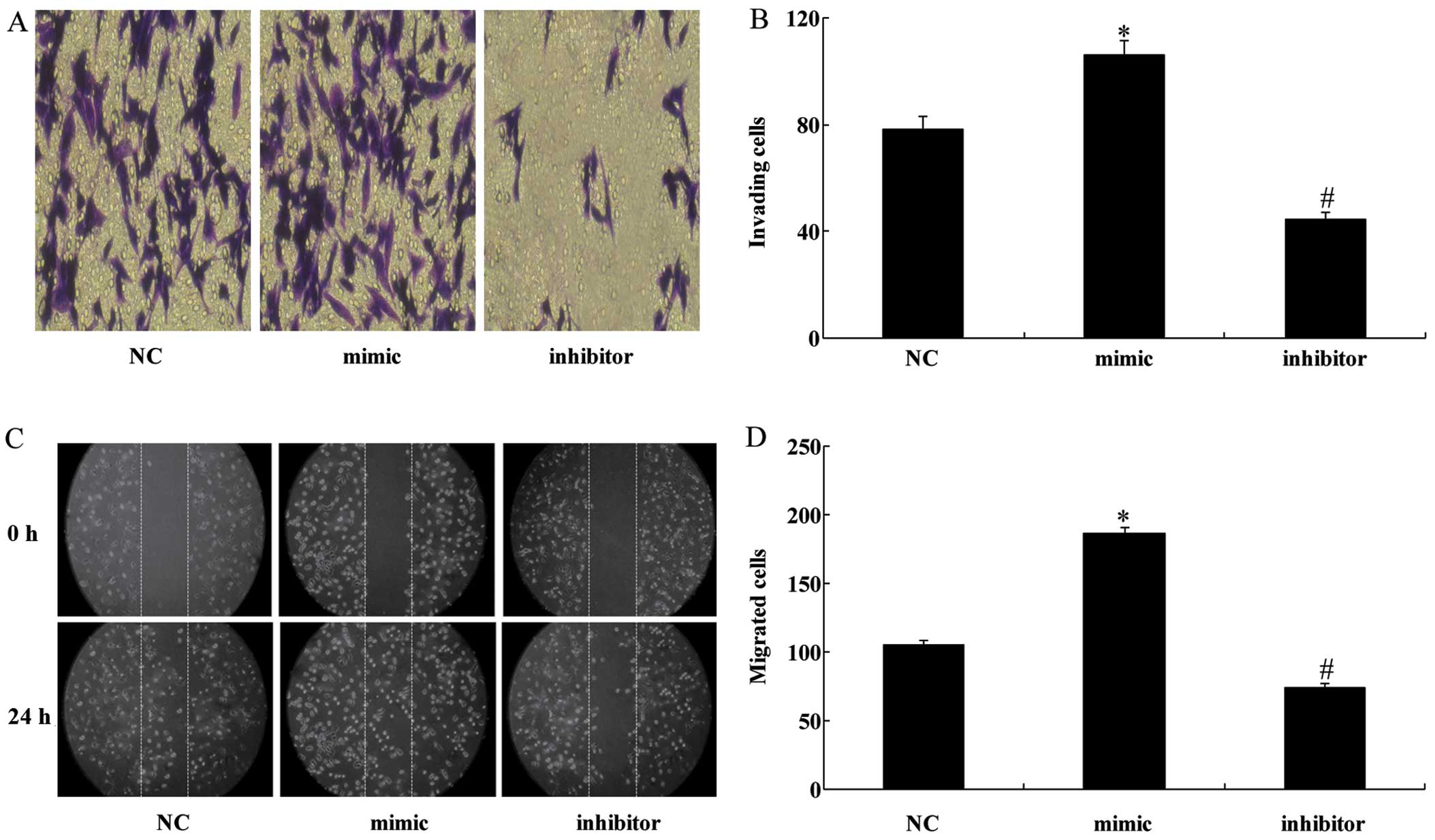
miR-130b regulates the migration and
invasion of U251 cells in vitro
As invasiveness is one of the pathophysiological
features of malignant human gliomas, the effects of miR-130b on the
invasiveness and migration of glioma cells were examined by
Transwell and scratch-wound assays, respectively. The former showed
that the overexpression of miR-130b significantly enhanced the
invasive ability of U251 cells whereas the downregulation of
miR-130b significantly decreased the number of cells capable of
invasion (Fig. 5A and B). The
scratch-wound assay also demonstrated that the miR-130b mimic
enhanced the invasiveness of U251 cells and the miR-130 inhibitor
inhibited the invasiveness of the cells (Fig. 5C and D).
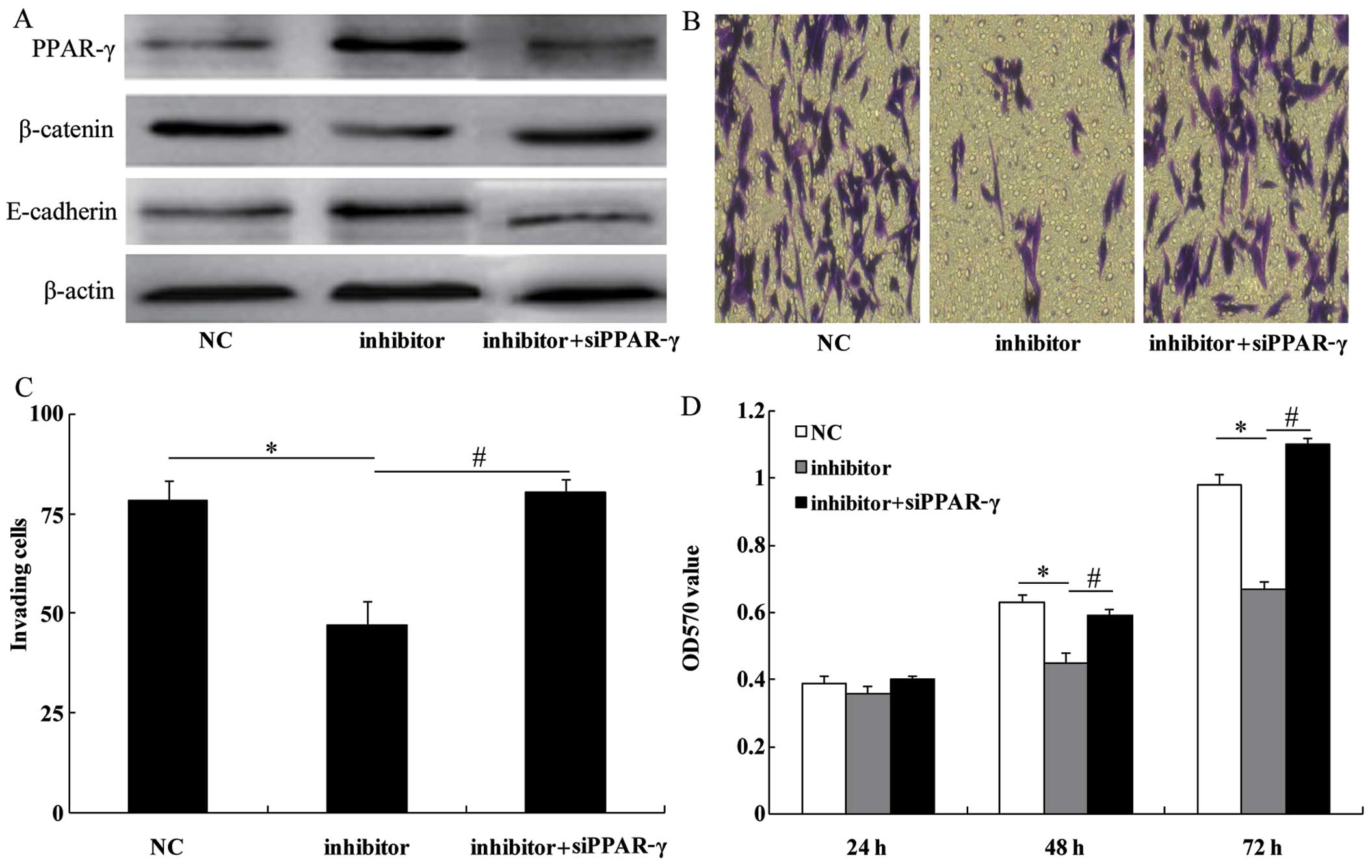
PPAR-γ siRNA imitates the role of
miR-130b in U251 glioma cells
To examine the role of PPAR-γ in the
miR-130b-dependent cell proliferation and invasion, the miR-130b
inhibitor was transfected into the U251 cells treated with PPAR-γ
siRNA. As expected, PPAR-γ siRNA co-transfected with the miR-130b
inhibitor decreased the expression levels of PPAR-γ (Fig. 6A). The effect of the miR-130b
inhibitor on cell invasion (Fig. 6B
and C) and proliferation (Fig.
6D) was reversed by the silencing of PPAR-γ expression. These
results indicate that PPAR-γ plays an essential role in
miR-130b-dependent cell proliferation and invasion.
Discussion
Gliomas are the most common primary tumors of the
central nervous system with glioblastomas as the most malignant
entity (1). Despite progress in
glioma therapy regimens, such as surgery, radiation, chemotherapy
or combined modalities, the prognosis for malignant glioma patients
remains dismal (19), and most
patients with glioma die within two years of diagnosis (20). Therefore, extensive study of the
biological characteristics of gliomas is necessary in order to
identify an effective treatment capable of suppressing the
invasiveness of glioma cells. Over the last few years, miRNAs have
emerged as a new class of gene regulators which are associated with
different malignancies. For example, miR-222 promoted the
proliferation of epithelial ovarian cancer cells by downregulating
p27Kip1 (21). miR-218
reversed the highly invasive nature of glioblastoma cells by
targeting the oncogenic transcription factor LEF1 (22).
miRNAs are emerging as a novel class of regulatory
molecules involved in numerous biological processes (23,24). They regulate gene expression at
the transcriptional or translational level by binding to the 3′-UTR
of mRNAs (25). miRNA
deregulation is a common feature of human malignancies as they
control the expression of oncogenes or tumor suppressors by acting
as onco-miRNAs or tumor suppressor miRNAs (24). Previous studies have demonstrated
the association between miR-130b with some types of solid tumors.
For example, Yu et al demonstrated that miR-130b is
significantly overexpressed in gastric cancer, which increased the
proliferation of esophageal squamous cell carcinoma cells and
enhanced their ability to migrate and invade through interactions
with the 3′-UTR of phosphatase and tensin homolog (PTEN) to
downregulate PTEN expression (26). On the contrary, it has been shown
that miR-130b is downregulated in ovarian cancer and papillary
thyroid carcinoma, and its expression inversely correlates with
tumor aggressiveness and multidrug resistance in these types of
cancer (15,27). In recent years, miR-130b has been
found to be overexpressed in colorectal cancer and hepatocellular
carcinoma (28). It has also been
observed that miR-130b represses the expression of PPAR-γ protein
by targeting the PPAR-γ 3′-UTR, leading to enhancement of the
oncogenic capacities of cancer cells (12,28). In the present study, we used
RT-qPCR to confirm that miR-130b is overexpressed in glioma tissues
and cells. Furthermore, upregulated miR-130b induced cell
proliferation, decreased the percentage of cells in the G0/G1
phase, and enhanced the invasiveness of U251 glioma cells whereas
these abilities were weakened when miR-130b was inhibited. PPAR-γ
was identified as a direct functional target of miR-130b using
bioinformatics analysis, and this finding was experimentally
confirmed by performing a dual luciferase reporter assay.
PPAR-γ is a ligand-activated transcription factor
that belongs to the superfamily of hormone receptors (29). It is abundantly expressed in many
cell types, where it regulates lipid metabolism, glucose
homeostasis, tumor progression and inflammation (30). Some studies have verified that
PPAR-γ is implicated in epithelial cell differentiation and
anti-proliferative responses, acting as a tumor suppressor
(31,32). It has also been demonstrated that
PPAR-γ is frequently downregulated in human glioma cell lines
including SWO-38 and U251 glioma cells (33), and PPAR-γ agonists induce growth
arrest and apoptosis in glioma cells, suggesting that they may be
suitable for use in the treatment of brain tumors (34). A previous study has proposed that
the PPAR-signaling pathway is connected to the β-catenin pathway;
β-catenin is a multifunctional, intracellular protein that binds to
either E-cadherin or APC proteins; β-catenin forms a complex with
the transmembrane receptor E-cadherin, becomes plasma
membrane-associated and mediates intercellular adhesion (35). Wan et al found that the
PPAR-γ agonist pioglitazone not only suppressed the proliferation
and migration of U251 glioma cells and induced cell apoptosis, but
also decreased the expression level of the β-catenin protein. The
knockdown of β-catenin expression mimicked the anti-neoplastic
potency of pioglitazone. These findings suggest that the
PPAR-γ/β-catenin signaling pathway plays a key role in the
development of glioma (36). The
present study demonstrated that miR-130b binds to the 3′-UTR of
PPAR-γ mRNA and downregulates the protein expression of PPAR-γ and
E-cadherin and increases the expression of β-catenin. Furthermore,
the silencing of PPAR-γ expression reversed the effect of the
miR-130b inhibitor on cell proliferation and invasion.
Taken together, our findings reveal that miR-130b
expression is markedly upregulated in glioma tissues and cells.
PPAR-γ is a direct functional target of miR-130b. miR-130b
repressed PPAR-γ expression which downregulated E-cadherin and
increased the expression of β-catenin, thereby promoting the
proliferation and invasion of glioma cells in vitro. These
results suggest that miR-130b plays a critical role in promoting
the development and progression of glioma, and the downregulation
of miR-130b may be a useful treatment strategy for the management
of glioma.
References
|
1
|
Van Meir EG, Hadjipanayis CG, Norden AD,
Shu HK, Wen PY and Olson JJ: Exciting new advances in
neuro-oncology: the avenue to a cure for malignant glioma. CA
Cancer J Clin. 60:166–193. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Louis DN, Ohgaki H, Wiestler OD, Cavenee
WK, Burger PC, Jouvet A, Scheithauer BW and Kleihues P: The 2007
WHO classification of tumours of the central nervous system. Acta
Neuropathol. 114:97–109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wiedemeyer R, Brennan C, Heffernan TP,
Xiao Y, Mahoney J, Protopopov A, Zheng H, Bignell G, Furnari F,
Cavenee WK, et al: Feedback circuit among INK4 tumor suppressors
constrains human glioblastoma development. Cancer Cell. 13:355–364.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Coras R, Hölsken A, Seufert S, Hauke J,
Eyüpoglu IY, Reichel M, Tränkle C, Siebzehnrübl FA, Buslei R,
Blümcke I and Hahnen E: The peroxisome proliferator-activated
receptor-γ agonist troglitazone inhibits transforming growth
factor-β-mediated glioma cell migration and brain invasion. Mol
Cancer Ther. 6:1745–1754. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wu Z, He B, He J and Mao X: Upregulation
of miR-153 promotes cell proliferation via downregulation of the
PTEN tumor suppressor gene in human prostate cancer. Prostate.
73:596–604. 2013. View Article : Google Scholar
|
|
6
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kent OA and Mendell JT: A small piece in
the cancer puzzle: microRNAs as tumor suppressors and oncogenes.
Oncogene. 25:6188–6196. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wu Z, Han Y, Li Y, Li X, Sun T, Chen G,
Huang Y, Zhou Y and Du Z: MiR-218-5p inhibits the stem cell
properties and invasive ability of the
A2B5+CD133− subgroup of human glioma stem
cells. 35:869–877. 2016.
|
|
9
|
Sand M, Skrygan M, Sand D, Georgas D,
Gambichler T, Hahn SA, Altmeyer P and Bechara FG: Comparative
microarray analysis of microRNA expression profiles in primary
cutaneous malignant melanoma, cutaneous malignant melanoma
metastases, and benign melanocytic nevi. Cell Tissue Res.
351:85–98. 2013. View Article : Google Scholar
|
|
10
|
Lai KW, Koh KX, Loh M, Tada K, Subramaniam
MM, Lim XY, Vaithilingam A, Salto-Tellez M, Iacopetta B, Ito Y, et
al: Singapore Gastric Cancer Consortium: MicroRNA-130b regulates
the tumour suppressor RUNX3 in gastric cancer. Eur J Cancer.
46:1456–1463. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Scheffer AR, Holdenrieder S, Kristiansen
G, von Ruecker A, Müller SC and Ellinger J: Circulating microRNAs
in serum:novel biomarkers for patients with bladder cancer? World J
Urol. 32:353–358. 2014. View Article : Google Scholar
|
|
12
|
Colangelo T, Fucci A, Votino C, Sabatino
L, Pancione M, Laudanna C, Binaschi M, Bigioni M, Maggi CA, Parente
D, et al: MicroRNA-130b promotes tumor development and is
associated with poor prognosis in colorectal cancer. Neoplasia.
15:1218–1231. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu X, Weng L, Li X, Guo C, Pal SK, Jin JM,
Li Y, Nelson RA, Mu B, Onami SH, et al: Identification of a
4-microRNA signature for clear cell renal cell carcinoma metastasis
and prognosis. PLoS One. 7:e356612012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Malzkorn B, Wolter M, Liesenberg F,
Grzendowski M, Stühler K, Meyer HE and Reifenberger G:
Identification and functional characterization of microRNAs
involved in the malignant progression of gliomas. Brain Pathol.
20:539–550. 2010. View Article : Google Scholar
|
|
15
|
Yip L, Kelly L, Shuai Y, Armstrong MJ,
Nikiforov YE, Carty SE and Nikiforova MN: MicroRNA signature
distinguishes the degree of aggressiveness of papillary thyroid
carcinoma. Ann Surg Oncol. 18:2035–2041. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dong P, Karaayvaz M, Jia N, Kaneuchi M,
Hamada J, Watari H, Sudo S, Ju J and Sakuragi N: Mutant p53
gain-of-function induces epithelial-mesenchymal transition through
modulation of the miR-130b-ZEB1 axis. Oncogene. 32:3286–3295. 2013.
View Article : Google Scholar :
|
|
17
|
Leone V, Langella C, D'Angelo D, Mussnich
P, Wierinckx A, Terracciano L, Raverot G, Lachuer J, Rotondi S,
Jaffrain-Rea ML, et al: Mir-23b and miR-130b expression is
downregulated in pituitary adenomas. Mol Cell Endocrinol. 390:1–7.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao G, Zhang JG, Shi Y, Qin Q, Liu Y,
Wang B, Tian K, Deng SC, Li X, Zhu S, et al: MiR-130b is a
prognostic marker and inhibits cell proliferation and invasion in
pancreatic cancer through targeting STAT3. PLoS One. 8:e738032013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Walbert T and Chasteen K: Palliative and
supportive care for glioma patients. Cancer Treat Res. 163:171–184.
2015. View Article : Google Scholar
|
|
20
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sun C, Li N, Zhou B, Yang Z, Ding D, Weng
D, Meng L, Wang S, Zhou J, Ma D and Chen G: miR-222 is upregulated
in epithelial ovarian cancer and promotes cell proliferation by
downregulating P27(kip1). Oncol Lett. 6:507–512. 2013.PubMed/NCBI
|
|
22
|
Liu Y, Yan W, Zhang W, Chen L, You G, Bao
Z, Wang Y, Wang H, Kang C and Jiang T: MiR-218 reverses high
invasiveness of glioblastoma cells by targeting the oncogenic
transcription factor LEF1. Oncol Rep. 28:1013–1021. 2012.PubMed/NCBI
|
|
23
|
Stefani G and Slack FJ: Small non-coding
RNAs in animal development. Nat Rev Mol Cell Biol. 9:219–230. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Garzon R, Calin GA and Croce CM: MicroRNAs
in cancer. Annu Rev Med. 60:167–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Esquela-Kerscher A and Slack FJ: Oncomirs
– microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yu T, Cao R, Li S, Fu M, Ren L, Chen W,
Zhu H, Zhan Q and Shi R: MiR-130b plays an oncogenic role by
repressing PTEN expression in esophageal squamous cell carcinoma
cells. BMC Cancer. 15:292015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang C, Cai J, Wang Q, Tang H, Cao J, Wu L
and Wang Z: Epigenetic silencing of miR-130b in ovarian cancer
promotes the development of multidrug resistance by targeting
colony-stimulating factor 1. Gynecol Oncol. 124:325–334. 2012.
View Article : Google Scholar
|
|
28
|
Tu K, Zheng X, Dou C, Li C, Yang W, Yao Y
and Liu Q: MicroRNA-130b promotes cell aggressiveness by inhibiting
peroxisome proliferator-activated receptor gamma in human
hepatocellular carcinoma. Int J Mol Sci. 15:20486–20499. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kliewer SA, Xu HE, Lambert MH and Willson
TM: Peroxisome proliferator-activated receptors: from genes to
physiology. Recent Prog Horm Res. 56:239–263. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pestereva E, Kanakasabai S and Bright JJ:
PPARγ agonists regulate the expression of stemness and
differentiation genes in brain tumour stem cells. Br J Cancer.
106:1702–1712. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Michalik L, Desvergne B and Wahli W:
Peroxisome-proliferator-activated receptors and cancers: complex
stories. Nat Rev Cancer. 4:61–70. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Drori S, Girnun GD, Tou L, Szwaya JD,
Mueller E, Xia K, Shivdasani RA and Spiegelman BM: Hic-5 regulates
an epithelial program mediated by PPARgamma. Genes Dev. 19:362–375.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang MH, Zhong XY, Lin CL, Xie YK, Jia JP,
Li SM and Mi C: Expression of peroxisome proliferator-activated
receptor gamma in glioma. Nan Fang Yi Ke Da Xue Xue Bao.
28:444–446. 2008.In Chinese. PubMed/NCBI
|
|
34
|
Papi A, Tatenhorst L, Terwel D, Hermes M,
Kummer MP, Orlandi M and Heneka MT: PPARgamma and RXRgamma ligands
act synergistically as potent antineoplastic agents in vitro and in
vivo glioma models. J Neurochem. 109:1779–1790. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jansson EA, Are A, Greicius G, Kuo IC,
Kelly D, Arulampalam V and Pettersson S: The Wnt/β-catenin
signaling pathway targets PPARgamma activity in colon cancer cells.
Proc Natl Acad Sci USA. 102:1460–1465. 2005. View Article : Google Scholar
|
|
36
|
Wan Z, Shi W, Shao B, Shi J, Shen A, Ma Y,
Chen J and Lan Q: Peroxisome proliferator-activated receptor γ
agonist pioglitazone inhibits β-catenin-mediated glioma cell growth
and invasion. Mol Cell Biochem. 349:1–10. 2011. View Article : Google Scholar : PubMed/NCBI
|