Introduction
The majority of wounds, such as a surgical incision
or superficial injury will heal in an orderly manner within an
acceptable period of time. In a chronic wound, there is often an
underlying pathological process, either causing a prolonged injury
to tissues or greatly impairing normal healing (1). Wounds that fail to heal within 3
months are considered chronic and 3.55/1,000 people in the UK and
3% of those over 60 years of age suffer from chronic wounds
(2). The prevalence is likely to
rise with an ageing population. Chronic wounds have a major
financial impact. The cost to the NHS of frequent nurse and doctor
appointments, dressings as well as outpatient and inpatient
treatment complications is significant. The cost of chronic wound
management in the UK has been estimated to be between £2.3 and £3.1
billion annually (3) or 2–3% of
the NHS budget (4).
The process of repairing tissue after an injury is
complex, involving overlapping steps. The phases are tightly
regulated by a plethora of cytokines and growth factors controlling
multiple cell types to proliferate, differentiate and migrate.
Originally, 4 distinct phases were described. Namely: haemostasis,
inflammation, proliferation and maturation or remodelling (5). In an acute wound, the phases
progress in an orderly and timely fashion, with the prompt
achievement of healing. Chronic wounds can become arrested in one
of the above phases of healing or the phases progress more slowly
than normal. In a chronic wound, the complex cascade of cytokines
and cellular activity is impaired leading to prolonged
inflammation, defective wound matrix and a failure to
re-epithelialize. Several cytokines have been implicated as having
a role in the process of wound healing. As inflammation is a key
stage in wound healing, it is perhaps not surprising that many of
these cytokines are known to be regulators of inflammation.
One such pro-inflammatory cytokine is interleukin
(IL)-15. IL-15 was identified and characterised as a 14–18 kDa
protein using the simian kidney epithelial cell line CV-1/EBNA
(6). The data collected from this
study allowed for the isolation of human IL-15 by specifically
probing the human stromal cell line IMTLH cDNA library (6). The human IL-15 structure was defined
as a four α-helical bundle cytokine and is located on the human
chromosome 4q31 (6). IL-15 can
exert its biological effects using two separate signalling
pathways, the first of which involves conventional binding and
signalling via the IL-15 receptor α (IL-15Rα), β and γ chains.
IL-15 and its family member IL-2 are separate entities with
different sequence homologies, but there is data to prove
significant cross-over between IL-15 and the β and γ chains of the
IL-2 receptor (6,7). This then results in the upregulation
of natural killer (NK) cell and T-cell activity (6,8,9).
Conversely, IL-15 in mast cells binds to its cognate IL-15RX
receptor and does not require IL-2 receptor input (9,10).
However, the mRNA expression of IL-15 in activated T cells is not a
significant alternative to the large pools of IL-2 present
(6). Previous findings confirm
that IL-15Rα and its juxtracrine-signalling ligand IL-15 are
present in human adult low calcium temperature (HaCaT)
keratinocytes (11). Previous
studies have explored IL-15 in wound biology, regeneration and
inflammation and IL-15 has been demonstrated to promote the
proliferation of the human keratinocyte HaCaT cell line, through
ERK- and AKT-dependent pathways and to exert anti-apoptotic effects
on epidermal keratinocytes (12,13). Other evidence has suggested that
IL-15 promotes wound healing responses in the liver and may enhance
regeneration following liver damage (14). A study by Kagimoto et al
using transgenic IL-15 mouse models, has suggested a regulatory
role for IL-15 in wound healing and mucosal infection, where
transgenic IL-15 mice displayed accelerated wound healing but
enhanced susceptibility to genital HSV-2 infection (15). IL-15 has also been implicated in a
range of inflammatory diseases [reviewed in (16)].
As the clinical field of chronic wounds is such a
significant burden, new diagnostic markers and potential
therapeutic targets are needed. This study aims to assess whether
IL-15 has a role in the healing of chronic wounds and to assess its
effects in vitro on keratinocyte growth and migration.
Materials and methods
Cells and materials
The HaCaT cell line was purchased from the German
Cancer Research Institute (Heidelberg, Germany). Cells were
cultured in Dulbecco's modified Eagle's medium (DMEM)/Ham's F12
with L-glutamine medium (Sigma-Aldrich, Gillingham, UK)
supplemented with antibiotics (final concentration streptomycin 0.1
mg/ml, penicillin 100 U/ml, amphotericin 0.25 µg/ml) and 10%
foetal calf serum (Sigma-Aldrich). Cells were routinely cultured in
25 or 75 cm2 tissue culture flasks (Greiner Bio-One
Ltd., Stonehouse, UK) and incubated at 37°C, 5% CO2 and
95% humidity.
Recombinant IL-15 (rhIL-15) was purchased from
Insight Biotechnology (Wembley, UK). ERK inhibitor II, AKT
inhibitor and PLCγ inhibitor (U73122) were purchased from
Calbiochem (Merck Chemicals Ltd., Nottingham, UK). FAK inhibitor
was purchased from Tocris (Bristol, UK).
Clinical samples
Biopsies were taken from chronic venous leg ulcers
in a cohort of 71 patients. The inclusion criteria were as follows:
no evidence of other diagnosis, no evidence of infection and ulcer
present for at least 3 months. The samples were collected at the
wound healing clinic, University Hospital of Wales (UHW; Cardiff,
UK), after obtaining ethics approval from the South East Wales
Research Ethics Committee and informed written consent from the
subjects. The patients were treated for 12 weeks with standard
treatment. Specifically, this consisted of regular wound care with
appropriate dressings and compression. Compression was produced
with 3 layers of Tubigrip™ bandages (Mölnlycke Health Care,
Dunstable, UK) at an appropriate size to deliver 40 mmHg of
pressure. At the end of the 12 week treatment period, the wounds
were assessed. The biopsies from those that had decreased in size
or healed were termed 'healing'. The biopsies from those that had
increased in size or had shown no change were termed 'non-healing'.
The cohort has been described previously (17).
RNA extraction, quantification and
reverse transcription
RNA was extracted using TRI reagent (Sigma-Aldrich)
in accordance with the manufacturer's instructions. In brief,
medium was aspirated from a flask containing a confluent monolayer
of cells and the cells were lysed in TRI reagent. The cell lysate
was placed in a 1.8 ml Eppendorf tube and incubated at 4°C for 5
min. Chloroform (0.2 ml) was subsequently added and the solution
thoroughly shaken for 15 sec before standing for 15 min at room
temperature. Samples were then centrifuged at 12,000 × g for 15 min
at 4°C and the upper aqueous phase was removed with a pipette and
added to an equal volume of isopropanol. This sample was incubated
at 4°C for 10 min and then centrifuged at 12,000 × g for 10 min to
precipitate RNA. The RNA pellet was washed twice in 75% ethanol in
DEPC water before being dried in a drying oven (Techne Hybridiser
HB-1D; Wolf Laboratories, York, UK) for 10 min at 55°C. The RNA
pellet was dissolved in 50–100 µl of DEPC water prior to
quantification using an Implen nanophotometer (Implen Gmbh, Munich,
Germany) and standardisation.
Following quantification, standardised amounts of
RNA were reverse transcribed to generate cDNA using a reverse
transcription kit (iScript cDNA synthesis kit; Bio-Rad
Laboratories, Hemel Hempstead, UK). Subsequently, cDNA was diluted
appropriately and stored at −20°C until required.
Polymerase chain reaction (PCR)
RT-PCR was performed using the GoTaq Green master
mix (Promega, Madison, WI, USA). In each reaction, GoTaq Green
master mix, forward primer (10 pmol), reverse primer (10 pmol) and
nuclease-free water were added to cDNA in a 200 µl PCR tube.
The primers used in this study are detailed in Table I. Primer3 software was used to
identify primer binding sites and predicted product size. The
specific cycling conditions were 94°C for 5 min followed by 32–34
cycles of 94°C for 30 sec, 55°C for 30 sec and 72°C for 40 sec.
Following this, a final extension was undertaken at 72°C for 10
min. Following PCR, samples were size separated using gel
electrophoresis, stained in SYBR Safe (Life Technologies, Paisley,
UK) and visualised under blue light using a Syngene U:Genius 3
system (Geneflow, Lichfield, UK).
 | Table IPrimers used in the study. |
Table I
Primers used in the study.
| Primer | Sense | Antisense |
|---|
| GAPDH |
GGCTGCTTTTAACTCTGGTA |
GACTGTGGTCATGAGTCCTT |
| IL-15 |
AACAGTCATTTTCTAACTGAAGC |
ACTGAACCTGACCGTACATTCAAATCACTTATTACATTCACC |
| IL-15Rα |
TCTCAGACAACAGCCAAGA |
ACTGAACCTGACCGTACAGACAGTGGACGTGGAGATAG |
| GAPDH (qPCR) |
AAGGTCATCCATGACAACTT |
ACTGAACCTGACCGTACAGCCATCCACAGTCTTCTG |
Quantitative PCR (qPCR)
The Ampliflor UniPrimer Universal system (Intergen
Co., Purchase, NY, USA) was used to quantify relative transcript
copy numbers in each cDNA sample as previously described (18). Sample cDNA was combined with 2X iQ
supermix (Bio-Rad Laboratories), forward primer (10 pmol), reverse
primer containing the z sequence (1 pmol) and a UniPrimer probe (10
pmol). The specific primers are outlined in Table I, and the cycling conditions were
initial denaturing at 94°C for 5 min followed by 60 cycles of 94°C
for 10 sec, 55°C for 15 sec and 72°C for 20 sec. The incorporation
and unfolding of the UniPrimer probe, due to its similarity to the
z sequence, separates a fluorophore from a quencher moiety and
allows detection of the fluorescent signal which is indicative of
DNA amplification. Quantification of this signal against a standard
set of samples allows the quantification of relative transcript
copy numbers. Sample quantity was further normalised against the
GAPDH housekeeping gene levels present in the samples.
In vitro growth assay
Cells were seeded into duplicate 96-well plates at a
density of 3,000 cells/well. Subsequently, the same volume of
either double concentrate rhIL-15 or normal medium was added to the
wells to give final concentrations of 1, 10, 40 or 100 ng/ml as
required. Each 96-well plate was incubated at 37°C with 5%
CO2, either overnight or for 3 days. At the end of the
incubation period, the medium was aspirated and the cells in each
well were fixed in 4% formaldehyde (v/v) (Sigma-Aldrich) for 10
min. The formaldehyde was then removed and each well was stained
with crystal violet (0.5% w/v) (Sigma-Aldrich) for 10 min. The
crystal violet was then washed off with water and re-suspended in
10% acetic acid (Sigma-Aldrich) prior to reading absorbance on a
plate reading spectrophotometer (ELx800; Bio-Tek, Swindon, UK).
Percentage growth after a 3-day incubation period was calculated
against the overnight plates.
Scratch assay
Cells were grown to confluence in a 24-well plate.
Following this, the confluent monolayer was scratched using a 200
µl pipette tip to generate an artificial wound.
Subsequently, the media was replaced with either normal media or
media containing 100 ng/ml rhIL-15 and the wound tracked over a 120
min time period under an inverted microscope (Leica DMIL LED) on a
heated plate at 37°C (both from Leica Microsystems GmbH, Wetzlar,
Germany). Images were captured every 30 min using a digital camera
(Leica DFC3000G; Leica Microsystems GmbH) and the distance between
the wound fronts were calculated using ImageJ software. The
distance migrated was calculated in comparison to the 0 h
wound.
Electric cell-substrate impedance sensing
(ECIS)
ECIS is a novel, real time method used to measure
the rate of cells repopulating an area from which they have been
eliminated using electrical wounding (19). This method can be used to screen
large sample numbers and has been previously described (17). In brief, the ECIS Zθ system with
96WE1 array plates were used (Applied Biophysics Inc., Troy, NY,
USA). Plates were stabilised prior to the addition of a
standardised number of HaCaT cells and either rhIL-15 treatment or
control media and incubated at 37°C. Resistance of the cells was
monitored by the system and once confluence was achieved, the
monolayer was wounded electrically (3,000 µA for 30
sec/well). Following wounding, the change in resistance across the
array was monitored and recorded as cells migrated back into the
wounded area.
Immunohistochemical (IHC) staining
A representative subset of tissues from chronic
wounds that contained both healing (n=12) and non-healing (n=11)
wounds was immunhistochemically analysed using a standard
peroxidase technique. Briefly, 7 µm frozen sections were
firstly fixed in dried acetone (Thermo Fisher Scientific,
Loughborough, UK) for 15 min, air dried for a further 15 min, prior
to rehydration in PBS. This was then followed by a permeabilisation
step where the sections were washed with 0.1% saponin/PBS
(Sigma-Aldrich) for 30 min. All subsequent washes contained 0.1%
saponin/PBS since this reaction is reversible. The sections were
then blocked for 1 h with a solution that contained 0.1% BSA/0.1%
saponin/10% horse serum in PBS. All incubations were performed in a
humidified box in order to prevent the sections from drying. Excess
blocking solution was then removed and the sections incubated with
the relevant primary antibody for 1 h. Two primary antibodies were
used: anti-IL-15 (sc-8437;) and anti-IL-15Rα (sc-9172) (both from
Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) at a
concentration of 1:100 (final concentration 2 µg/ml). The
sections were then washed with 0.1% saponin/PBS, prior to
incubation for 30 min with biotinylated secondary antibody (Vector
Laboratories, Nottingham, UK). Following washing, and a further 30
min incubation with the avidinbiotin complex (ABC) reagent, the
final reaction product was developed with 3′3-diaminobenzidine
solution (5 mg/ml). The sections were then washed in tap water,
counterstained with Gill's hematoxylin (Vector Laboratories),
dehydrated, cleared in xylene (Thermo Fisher Scientific) and
mounted in DPX (Merck Chemicals Ltd.). Negative controls were
performed by omitting the primary antibody and replacing it with
PBS.
Statistical analysis
The SigmaPlot 11 statistical package was used for
statistical analysis. Two-sample, two-tailed t-tests, Mann-Whitney
U test and one or two way ANOVA analysis were undertaken depending
on data normality. Experimental repeats were undertaken for all
assays and p<0.05 was considered to indicate a statistically
significant difference.
Results
Expression of IL-15 in clinical
samples
The expression profile of IL-15 was explored in
clinical samples from chronic wounds using both qPCR transcript
analysis and IHC staining. Quantitative transcript analysis of the
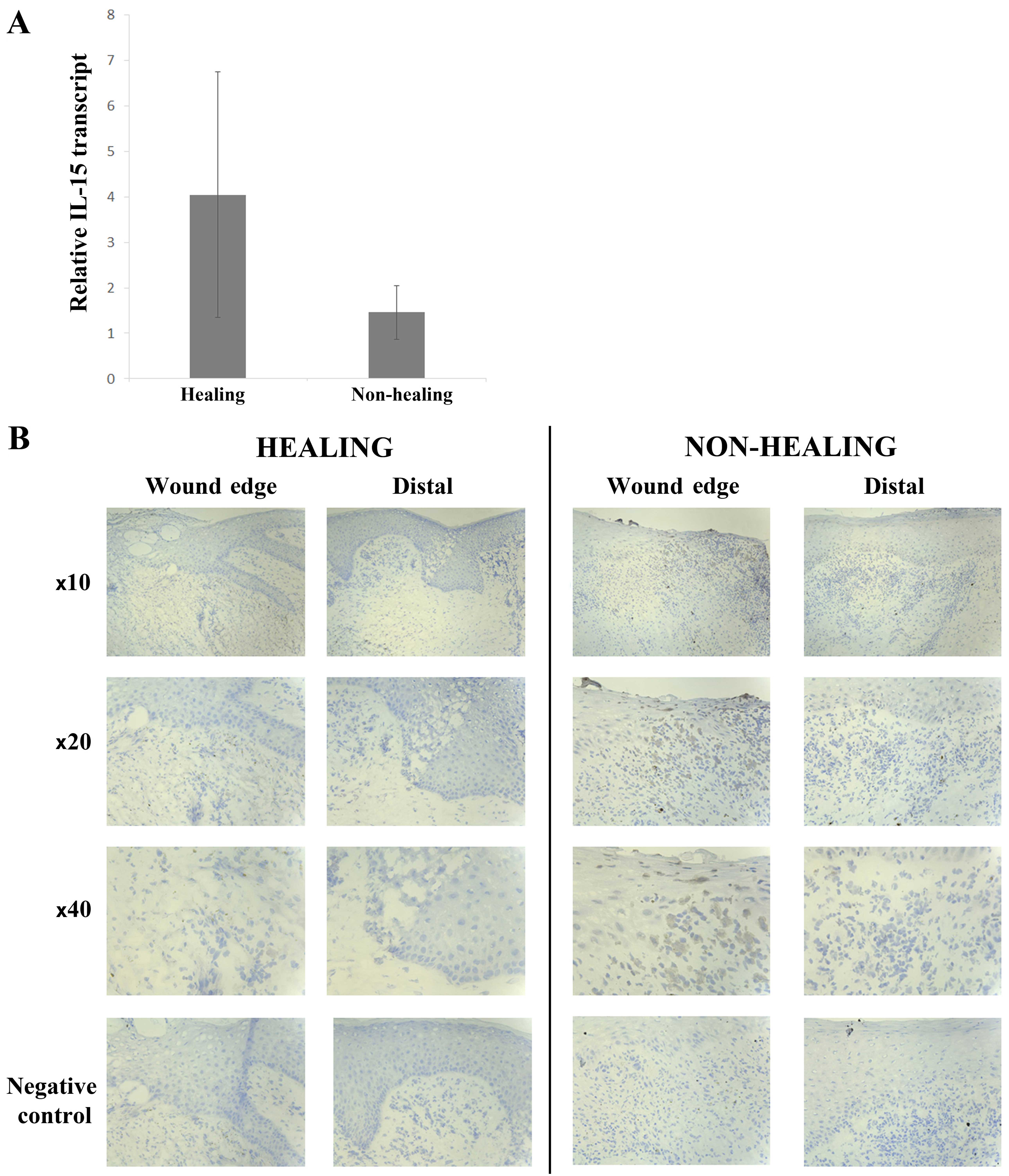
clinical samples showed a greater expression of IL-15 mRNA in
healing wounds (4.058±2.658) compared with non-healing wounds
(1.458±0.587) (Fig. 1A), though
this was not found to be statistically significant (p=0.17).
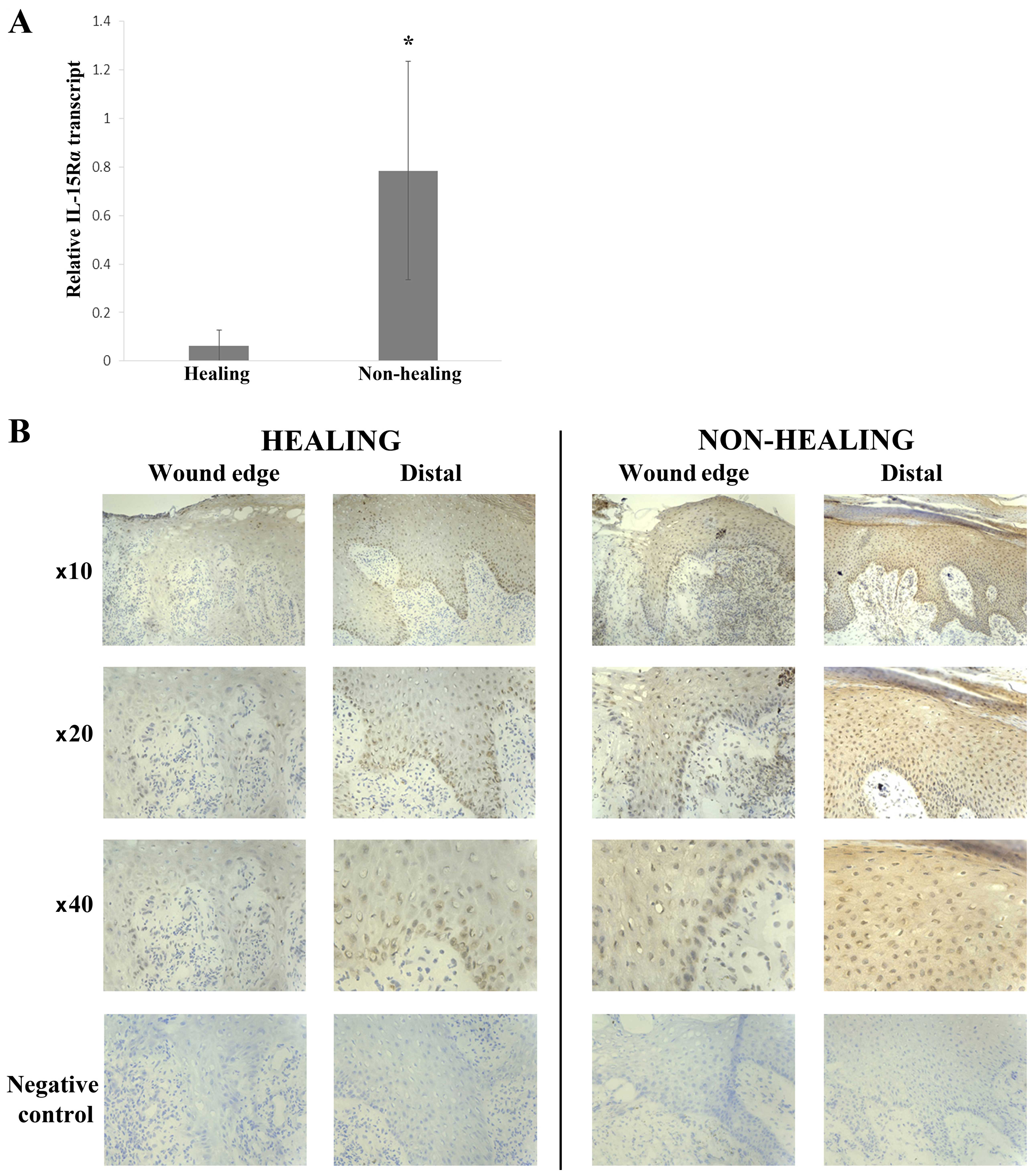
Conversely, there was a significantly greater expression of IL-15Rα
in non-healing wounds (0.785±0.45) compared with healing wounds
(0.0638±0.0638) (p=0.031) (Fig.
2A).
IHC staining for IL-15 was generally negative in
keratinocytes of all chronic wounds, both healing and non-healing
(Fig. 1B). Some cells of the
inflammatory infiltrate directly below the leading migratory wound
edge showed positive cytoplasmic staining for IL-15. These cells
had the appearance of macrophages. This was found mostly in
non-healing wounds (8/11 or 73%) compared with healing wounds (2/12
or 17%). IL-15Rα expression was seen in the majority of chronic
wounds (14/23) (Fig. 2B) with a
slightly higher number of non-healing wounds expressing IL-15Rα
compared with healing wounds [8/11 (73%) and 6/12 (50%),
respectively]. Generally, IL-15Rα expression in the healing wounds
was nuclear in the basal and lower layers of the epidermis,
increasing with intensity towards the distal/normal tissue (moving
away from the wound edge). This staining pattern was also seen in
the non-healing chronic wounds accompanied by cytoplasmic
expression in the mature keratinocytes distal to the wound
edge.
Impact of IL-15 on HaCaT
proliferation
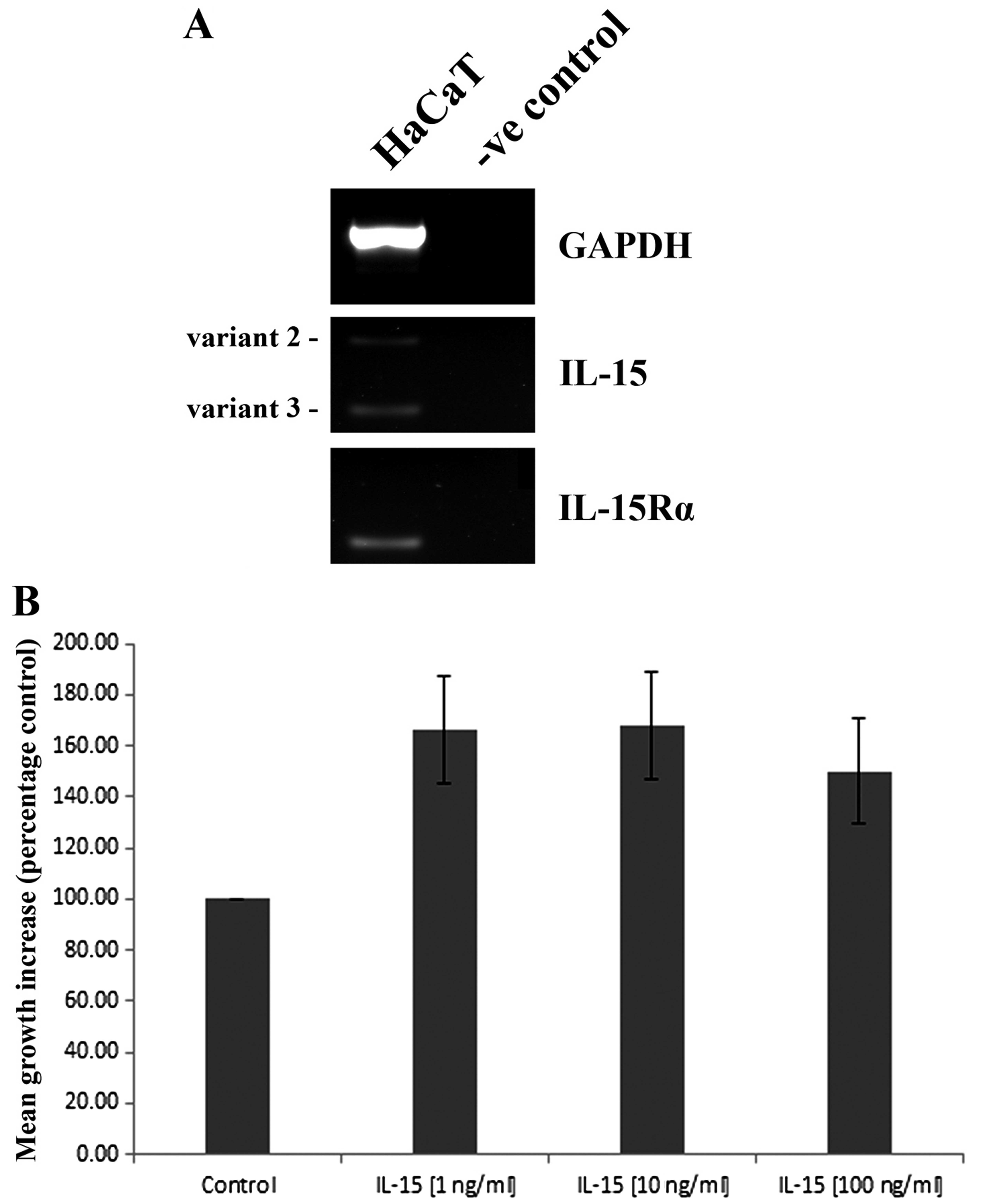
RT-PCR analysis indicated that HaCaT cells express
both IL-15 and IL-15Rα transcripts (Fig. 3A). Expression of the receptor
appears to be stronger than IL-15, of which two transcript variants
were detected. A recombinant form of IL-15 (rhIL15) was
subsequently used to explore the effects of this molecule on HaCaT
cell function. The treatment of HaCaT cells with either 1, 10 or
100 ng/ml appeared to enhance HaCaT cell growth over a 3 day
incubation, though ANOVA analysis was just outside of statistical
significance (p=0.07) (Fig.
3B).
IL-15 impacts on HaCaT cell
migration
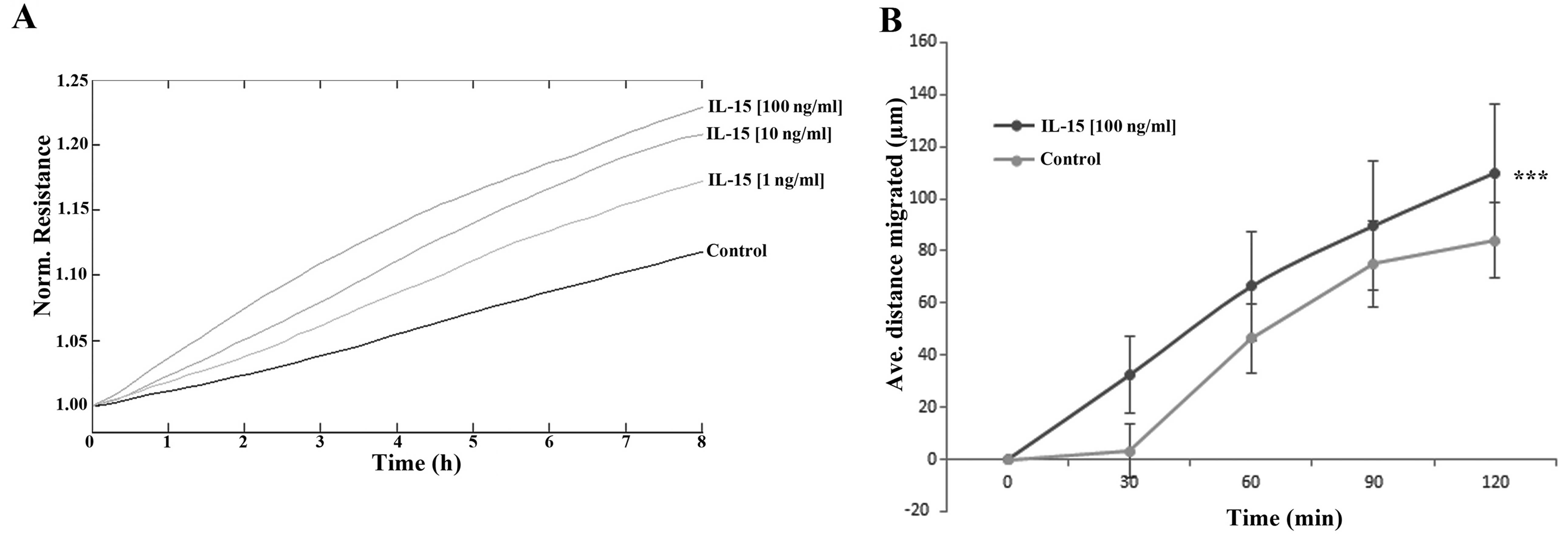
The impact of rhIL-15 was explored using both ECIS-
(Fig. 4A) and conventional
scratch- (Fig. 4B) based assays.
ECIS-based analysis indicated a dose-dependent enhancement of HaCaT
cell migration following treatment with rhIL-15, with the greatest
effects observed following treatment with 100 ng/ml rhIL-15. The
effect of this concentration was similarly confirmed using a
scratch assay methodology. Similar to the findings of the ECIS
assay, 100 ng/ml rhIL-15 enhanced HaCaT cell migration in
comparison with the untreated control cells and this effect was
found to be statistically significant (p<0.001).
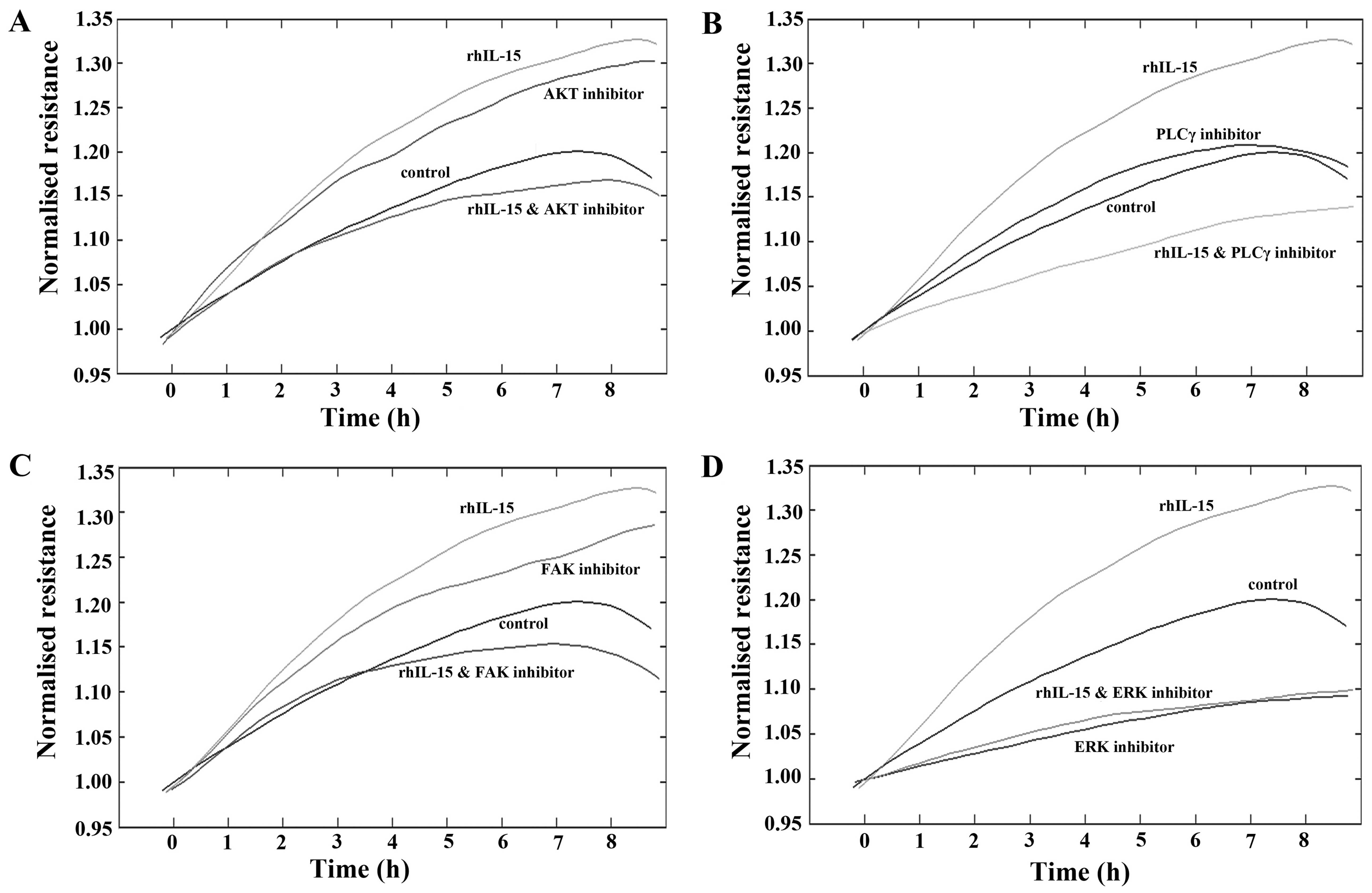
To further explore the potential mechanisms
responsible for the pro-migratory effects of rhIL-15 on HaCaT
cells, ECIS assays were undertaken using a number of small molecule
inhibitors of key migratory pathways (Fig. 5). Similar to previous findings,
treatment with rhIL-15 (40 ng/ml) continued to display a
pro-migratory response in comparison with the untreated cells.
The treatment of HaCaT cells with AKT inhibitor
appeared to enhance cell migration. Notably, the pro-migratory
effects observed due to both rhIL-15 or AKT inhibitor alone were
negated when these compounds were combined (Fig. 5A). The addition of a PLCγ
inhibitor did not substantially impact on the migration rates of
HaCaT cells in comparison with the untreated controls. However, in
combination with rhIL-15, PLCγ inhibition negated the pro-migratory
effect of rhIL-15 alone, and also substantially inhibited HaCaT
cell migration rates in comparison with the untreated control cells
(Fig. 5B). Similar trends to the
AKT inhibitor were observed with the addition of FAK inhibitor,
which appeared to exert pro-migratory effects independently but
when added in combination with rhIL-15, negated the pro-migratory
effect of both compounds (Fig.
5C). Finally, the addition of ERK inhibitor substantially
reduced HaCaT cell migration in comparison with the untreated
control cells and when added in combination with rhIL-15,
completely abolished the rhIL-15 pro-migratory effect,
demonstrating migratory rates similar to those seen with ERK
inhibition alone.
Discussion
IL-15 is a cytokine involved in a wide range of
biological processes and disease states. Given its involvement in
inflammatory diseases, keratinocyte function and potential to
affect wound healing, the present study aimed to explore the role
of this interleukin in chronic wounds.
Quantitative transcript analysis of the clinical
samples from the chronic wound cohort indicated a greater
expression of IL-15 mRNA in chronic wounds that go on to heal when
compared with those that do not heal, though this difference was
not found to be statistically significant. To the best of our
knowledge, this observation has not been previously documented as
most studies on IL-15 to date have focused on its role in
tumourigenesis, particularly in blood-borne tumours including
multiple myeloma (20), cutaneous
T-cell lymphoma (21) and T-cell
leukaemia (22). IHC staining
indicated little IL-15 protein expression in the keratinocytes of
healing or non-healing wounds, with little difference in expression
levels observed between the healing and non-healing chronic wound
samples. This discrepancy may be attributed to previous
observations that IL-15 transcripts are abundantly expressed
whereas IL-15 protein is tightly controlled with expression
generally limited to monocytes, macrophages and dendritic cells
(6,23). This is consistent with the IHC
observation that IL-15 protein was expressed in cells in the
inflammatory infiltrate directly below the migratory wound edge.
The cells in this infiltrate have been reported as being
predominantly leukocytes and macrophages (24). However, this was mostly found in
non-healing wounds compared with healing wounds, conflicting with
the qPCR results. Several translational and intracellular protein
trafficking mechanisms serve to limit the production of IL-15
protein from the far larger stores of transcript (25). This could explain the apparent
discrepancy between the mRNA and protein expression of IL-15.
Somewhat in contrast to this, the IL-15Rα receptor was found to be
upregulated in non-healing chronic wounds compared with healing
chronic wounds at both transcript and protein levels.
Following on from the clinical data, we explored the
impact of rhIL-15 on HaCaT keratinocytes and its impact on cell
growth and migration. The PCR results confirmed that IL-15 mRNA and
that of IL-15Rα are expressed in HaCaT keratinocytes. This is
consistent with previous studies of IL-15 expression in
keratinocytes (11,26). In vitro treatment of HaCaT
cells with rhIL-15 did not result in statistically significant
changes to 3 day growth rates, though all treatment concentrations
displayed a trend of enhanced growth. Previous studies have
demonstrated a pro-proliferative role for IL-15 in HaCaT cells
(12) and this interleukin is
associated with the enhancement of cell proliferation and the
inhibition of apoptosis in numerous cell lines (11,27–29). One key effect outlined in this
study was the impact of rhIL-15 on HaCaT cell migration. Using both
ECIS- and scratch assay-based methods, the addition of rhIL-15
resulted in an increased rate of HaCaT cell migration. Our data
indicates that IL-15 is capable of enhancing the migratory capacity
of HaCaT keratinocytes. Similar trends have been seen in other cell
types and IL-15 has recently been reported to act in a
pro-migratory manner on the 5637 bladder carcinoma cell line in a
wound healing-based assay (30).
Subsequently, we explored the ability of a number of small molecule
inhibitors of key pathways to alter the effect of rhIL-15 on HaCaT
cell migration. A combination of rhIL-15 with either AKT, FAK, PLCγ
or ERK small molecule pathway inhibitors was capable of negating
the pro-migratory effect of rhIL-15, reducing migrational rates to
control or below control levels. Notably, the combination of
rhIL-15 with either AKT, FAK or PLCγ inhibitor all appeared to
bring about migrational rates below that of the individual
inhibitor alone, suggesting potential links between these pathways
and the pro-migratory effect of IL-15. IL-15 has previously been
shown to activate/signal via AKT and ERK1/2 pathways (12,30) and the subsequent inhibition of
these pathways could negate the proliferative role of IL-15 in
HaCaT keratinocytes (12) and the
pro-migratory role of IL-15 in 5637 bladder cancer cells (30). The present data implies a possible
association between these tested pathways and HaCaT cell migration,
though further research is warranted in order to fully explore this
association.
The present study suggests some complex roles for
IL-15 in chronic wound healing. Enhanced IL-15Rα levels are
observed in non-healing chronic wounds in comparison with healing
chronic wounds, though treatment of keratinocytes with rhIL-15
enhanced pro-healing traits such as growth, albeit
non-significantly, and migration. Further studies are necessary to
establish expression profiles in larger wound healing cohorts
containing normal skin as well as acute and chronic wound
subtypes.
Acknowledgments
The authors wish to thank the A4B Scheme of the
Welsh Government Ser Cymru, NRN Life Sciences Research Network
Wales and Cancer Research Wales for supporting this study.
References
|
1
|
Wright K: Acute and chronic wounds:
current managment concepts. Clin Nurse Spec. 21:172–173. 2007.
View Article : Google Scholar
|
|
2
|
Vowden K, Vowden P and Posnett J: The
resource costs of wound care in Bradford and Airedale primary care
trust in the UK. J Wound Care. 18:93–94. 96–98, 100 passim. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Posnett J and Franks PJ: The burden of
chronic wounds in the UK. Nurs Times. 104:44–45. 2008.PubMed/NCBI
|
|
4
|
Drew P, Posnett J and Rusling L; Wound
Care Audit T; Wound Care Audit Team: The cost of wound care for a
local population in England. Int Wound J. 4:149–155. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schilling JA: Wound healing. Surg Clin
North Am. 56:859–874. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Grabstein KH, Eisenman J, Shanebeck K,
Rauch C, Srinivasan S, Fung V, Beers C, Richardson J, Schoenborn
MA, Ahdieh M, et al: Cloning of a T cell growth factor that
interacts with the beta chain of the interleukin-2 receptor.
Science. 264:965–968. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Giri JG, Anderson DM, Kumaki S, Park LS,
Grabstein KH and Cosman D: IL-15, a novel T cell growth factor that
shares activities and receptor components with IL-2. J Leukoc Biol.
57:763–766. 1995.PubMed/NCBI
|
|
8
|
Carson WE, Giri JG, Lindemann MJ, Linett
ML, Ahdieh M, Paxton R, Anderson D, Eisenmann J, Grabstein K and
Caligiuri MA: Interleukin (IL) 15 is a novel cytokine that
activates human natural killer cells via components of the IL-2
receptor. J Exp Med. 180:1395–1403. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Waldmann TA and Tagaya Y: The multifaceted
regulation of interleukin-15 expression and the role of this
cytokine in NK cell differentiation and host response to
intracellular pathogens. Annu Rev Immunol. 17:19–49. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tagaya Y, Burton JD, Miyamoto Y and
Waldmann TA: Identification of a novel receptor/signal transduction
pathway for IL-15/T in mast cells. EMBO J. 15:4928–4939.
1996.PubMed/NCBI
|
|
11
|
Rückert R, Asadullah K, Seifert M,
Budagian VM, Arnold R, Trombotto C, Paus R and Bulfone-Paus S:
Inhibition of keratinocyte apoptosis by IL-15: a new parameter in
the pathogenesis of psoriasis? J Immunol. 165:2240–2250. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yano S, Komine M, Fujimoto M, Okochi H and
Tamaki K: Interleukin 15 induces the signals of epidermal
proliferation through ERK and PI 3-kinase in a human epidermal
keratinocyte cell line, HaCaT. Biochem Biophys Res Commun.
301:841–847. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lindner G, Rückert R, Bulfone-Paus S and
Paus R: Inhibition of chemotherapy-induced keratinocyte apoptosis
in vivo by an interleukin-15-IgG fusion protein. J Invest Dermatol.
110:457–458. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Suzuki A, McCall S, Choi SS, Sicklick JK,
Huang J, Qi Y, Zdanowicz M, Camp T, Li YX and Diehl AM:
Interleukin-15 increases hepatic regenerative activity. J Hepatol.
45:410–418. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kagimoto Y, Yamada H, Ishikawa T, Maeda N,
Goshima F, Nishiyama Y, Furue M and Yoshikai Y: A regulatory role
of interleukin 15 in wound healing and mucosal infection in mice. J
Leukoc Biol. 83:165–172. 2008. View Article : Google Scholar
|
|
16
|
McInnes IB and Gracie JA: Interleukin-15:
A new cytokine target for the treatment of inflammatory diseases.
Curr Opin Pharmacol. 4:392–397. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bosanquet DC, Harding KG, Ruge F, Sanders
AJ and Jiang WG: Expression of IL-24 and IL-24 receptors in human
wound tissues and the biological implications of IL-24 on
keratinocytes. Wound Repair Regen. 20:896–903. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jiang WG, Martin TA, Lewis-Russell JM,
Douglas-Jones A, Ye L and Mansel RE: Eplin-alpha expression in
human breast cancer, the impact on cellular migration and clinical
outcome. Mol Cancer. 7:712008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Keese CR, Wegener J, Walker SR and Giaever
I: Electrical wound-healing assay for cells in vitro. Proc Natl
Acad Sci USA. 101:1554–1559. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tinhofer I, Marschitz I, Henn T, Egle A
and Greil R: Expression of functional interleukin-15 receptor and
autocrine production of interleukin-15 as mechanisms of tumor
propagation in multiple myeloma. Blood. 95:610–618. 2000.PubMed/NCBI
|
|
21
|
Leroy S, Dubois S, Tenaud I, Chebassier N,
Godard A, Jacques Y and Dréno B: Interleukin-15 expression in
cutaneous T-cell lymphoma (mycosis fungoides and Sézary syndrome).
Br J Dermatol. 144:1016–1023. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kukita T, Arima N, Matsushita K, Arimura
K, Ohtsubo H, Sakaki Y, Fujiwara H, Ozaki A, Matsumoto T and Tei C:
Autocrine and/or paracrine growth of adult T-cell leukaemia tumour
cells by interleukin 15. Br J Haematol. 119:467–474. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bamford RN, Battiata AP and Waldmann TA:
IL-15: the role of translational regulation in their expression. J
Leukoc Biol. 59:476–480. 1996.PubMed/NCBI
|
|
24
|
Moore K, Ruge F and Harding KG: T
lymphocytes and the lack of activated macrophages in wound margin
biopsies from chronic leg ulcers. Br J Dermatol. 137:188–194. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kurys G, Tagaya Y, Bamford R, Hanover JA
and Waldmann TA: The long signal peptide isoform and its
alternative processing direct the intracellular trafficking of
interleukin-15. J Biol Chem. 275:30653–30659. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Blauvelt A, Asada H, Klaus-Kovtun V,
Altman DJ, Lucey DR and Katz SI: Interleukin-15 mRNA is expressed
by human keratinocytes Langerhans cells, and blood-derived
dendritic cells and is downregulated by ultraviolet B radiation. J
Invest Dermatol. 106:1047–1052. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kuniyasu H, Ohmori H, Sasaki T, Sasahira
T, Yoshida K, Kitadai Y and Fidler IJ: Production of interleukin 15
by human colon cancer cells is associated with induction of mucosal
hyperplasia, angiogenesis, and metastasis. Clin Cancer Res.
9:4802–4810. 2003.PubMed/NCBI
|
|
28
|
Shinozaki M, Hirahashi J, Lebedeva T, Liew
FY, Salant DJ, Maron R and Kelley VR: IL-15, a survival factor for
kidney epithelial cells, counteracts apoptosis and inflammation
during nephritis. J Clin Invest. 109:951–960. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hjorth-Hansen H, Waage A and Börset M:
Interleukin-15 blocks apoptosis and induces proliferation of the
human myeloma cell line OH-2 and freshly isolated myeloma cells. Br
J Haematol. 106:28–34. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Park SL, Kim WJ and Moon SK: p21WAF1
mediates the IL-15-induced migration and invasion of human bladder
cancer 5637 cells via the ERK1/2/NF-κB/MMP-9 pathway. Int
Immunopharmacol. 22:59–65. 2014. View Article : Google Scholar : PubMed/NCBI
|