Introduction
Sensorineural hearing loss is a severe hearing
impairment that affects millions of individuals worldwide.
Moreover, neuronal regeneration is uncommon in the mature inner ear
(1,2), in contrast to neurogenesis in the
adult brain, such as in the subventricular zone (SVZ) of the
lateral ventricles and the dentate gyrus of the hippocampus
(3). Inner ear neural stem cells
(NSCs) may exist in the spiral ganglion. Since NSCs can proliferate
and differentiate into neurons (4–6),
they may be used to regenerate damaged neurons in the inner ear.
However, the intrinsic 'self-repair' capacity of inner ear NSCs is
inactive in vivo for unclear reasons (unsuitable
microenvironment, lack of guidance, etc.). Therefore, successfully
harnessing the differentiation potential of inner ear NSCs may
restore hearing. While the molecular mechanisms underlying the
neuronal differentiation of inner ear NSCs are not yet fully
understood, microRNAs (miRNAs or miRs) may play an important role
in this process.
miRNAs are small non-coding RNAs that affect mRNA
stability or inhibit translation by binding complementary sequences
in the 3′-translated regions (3′-UTRs) of target mRNAs (7–10).
Thus, miRNAs regulate multiple biological functions, including cell
proliferation, differentiation and apoptosis (11,12). miRNA expression studies in mammals
using microarrays and reverse transcription-quantitative PCR
(RT-qPCR) have demonstrated that miRNAs are expressed in the
developing nervous system and in mature neurons (13–18). miRNAs have crucial functions in
neuronal development and plasticity (19,20). The miR-17 family has been shown to
play an integral role in the regulation of neuronal differentiation
(21). miR-124 expression has
been shown to be upregulated during neuronal differentiation
(22,23). Several lines of evidence have
indicated that miR-124 regulates neuronal differentiation by
inhibiting small C-terminal domain phosphatase (SCP1), a component
of the RE1-silencing transcription factor (REST)/neuron-restrictive
silencer factor (NRSF) transcriptional repression complex, and by
targeting polypyrimidine tract binding protein 1 (PTBP1), a global
repressor of brain-specific alternative splicing (24,25). Moreover, global miRNA expression
profiling by microarray analysis, RT-qPCR, and/or in situ
hybridization have revealed several miRNAs with distinct
spatio-temporal expression patterns in the embryonic and post-natal
mouse inner ear (25–28). While miRNAs regulate the
development, morphogenesis and function of the inner ear (29–33), it is not yet known whether miRNAs
also regulate the neuronal differentiation of inner ear NSCs.
In this study, 6 miRNAs (miR-124, miR-132, miR-134,
miR-20a, miR-17-5p and miR-30a-5p) were selected based on their
involvement in neuronal differentiation/neurogenesis as discussed
in the 'Introduction' and in the 'Discussion'; their expression
patterns during the neuronal differentiation of inner ear NSCs were
examined by RT-qPCR. Our data demonstrate that miR-124 is important
for the differentiation of inner ear NSCs into neurons. Our results
revealed that miR-124 expression is upregulated during neuronal
differentiation. Moreover, miR-124 increased the percentage of
cells expressing neuron-specific class III β-tubulin in culture,
and increased the neurite length in mouse inner ear NSCs. These
changes were accompanied by changes in the expression of
tropomyosin receptor kinase B (TrkB) and cell division control
protein 42 homolog (Cdc42). TrkB is a receptor of brain-derived
neurotrophic factor (BDNF), and participates in the regulation of
neurogenesis, neurite outgrowth and in the survival of spiral
ganglion neurons. The small GTP-ase, Cdc42, which regulates both
microtubules and actin filaments in a number of cells, regulates
the neurite extension of spiral ganglion neurons. Our data
demonstrated that miR-124 promotes the neuronal differentiation of
and neurite outgrowth in inner ear NSCs, and regulates the
expression of TrkB and Cdc42 in inner ear NSCs.
Materials and methods
Animals
Post-natal day 1 (P1) C57BL/6 mice (n=135;
Laboratory Animal Center of Sun Yat-sen University, Guangzhou,
China) were used for the experiments. The animals were sacrificed
according to policies set forth by the Animal Care and Use
Committee of Sun Yat-sen University based on the National
Institutes of Health guidelines for animal care. All animal
experiments were also approved the Animal Care and Use Committee of
Sun Yat-sen University.
Inner ear NSC cultures, neuronal
differentiation and immunostaining
The isolation and culture of NSCs from the spiral
ganglia of newborn C57BL/6 mice were performed as previously
described (4). Spiral ganglia,
isolated from 5 mice, were digested in 200 µl of 0.125%
trypsin in phosphate-buffered saline (PBS; Gibco/Thermo Fisher
Scientific, Waltham, MA, USA) at 37°C for 15 min. The cells were
carefully triturated with plastic 200 µl pipette tips,
centrifuged and suspended in 2 ml Dulbecco's modified Eagle's
medium (DMEM)/high glucose and F12 medium (mixed 1:1; Gibco/Thermo
Fisher Scientific) supplemented with N2 and B27 supplements
(Invitrogen/Thermo Fisher Scientific), epidermal growth factor
(EGF; 20 ng/ml), basic fibroblast growth factor (bFGF; 20 ng/ml),
insulin-like growth factor 1 (IGF-1; 20 ng/ml) (PeproTech, Rocky
Hill, NJ, USA). The cell suspension was passed through a 70-mm cell
strainer (BD Falcon, Franklin Lakes, NJ, USA) to remove cell
clumps. For neuronal differentiation, mouse inner ear NSCs at 5
days were harvested and cultured in laminin and
poly-L-lysine-coated dishes in DMEM-F12 (1:1) medium supplemented
with 20 ng/ml of neurotrophin-3 (NT-3), 20 ng/ml of BDNF) (both
from PeproTech), 10 ng/ml of leukemia inhibitory factor (LIF)
(R&D Systems, Inc., Minneapolis, MN, USA), 2 mM of L-glutamine
(Gibco/Thermo Fisher Scientific) and 3 mM of KCl (Sigma, St. Louis,
MO, USA) for 7 days. Half of the differentiation medium was
replaced every other day. The NSCs were allowed to differentiate
for up to 14 days (day 14). Cell proliferative ability was assessed
by BrdU incorporation assay as previously described (34). Undifferentiated and differentiated
inner ear NSCs were fixed with 4% paraformaldehyde and
immunostained with antibodies specific for BrdU (C2181; 1:10,000;
Sigma), Tuj1 (class III β-tubulin; MO15013; 1:250; Neuromics,
Edina, MN, USA), p27Kip1 (554069; 1:150; BD Biosciences,
San Jose, CA, USA), ATP-binding cassette sub-family G member 2
(Abcg2; sc-25822; 1:150; Santa Cruz Biotechnology, Inc., Dallas,
TX, USA), Sox2 (SAB2104660;1:250; Sigma), Nestin (MO15012; 1:200;
Neuromics), TrkB (ab18987; 1:200; Abcam, Cambridge, UK) and Cdc42
(sc-8401; 1:150 Santa Cruz Biotechnology, Inc.). Nuclei were
counterstained with Fluoroshield Mounting Medium with
4′,6-diamidino-2-phenylindole (DAPI; ab104139; Abcam).
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
The cells were collected in cold D-Hank's solution
(Gibco/Thermo Fisher Scientific), and RNA was extracted using
TRIzol reagent (Invitrogen/Thermo Fisher Scientific) and
column-purified with an RNeasy kit [Qiagen (Suzhou) Translational
Medicine Co., Ltd., Suzhou Industrial Park, China]. Reverse
transcription was performed using M-MLV reverse transcriptase
(Roche Diagnostics, Mannheim, Germany) according to the
instructions provided by the manufacturer. The sequences of the
primers were as follows: Nestin forward,
5′-GCCGAGCTGGAGCGCGAGTTAGAG-3′ and reverse,
5′-GCAAGGGGGAAGAGAAGGATGTCG-3′; Sox2 forward,
5′-ACCCGGGCCTCAACGCTCACG-3′ and reverse,
5′-TCCCCTTCTCCAGTTCGCAGTCC-3′; p27Kip1 forward,
5′-CTGGAGCGGATGGACGCCAGAC-3′ and reverse,
5′-CGTCTGCTCCACAGTGCCAGC-3′; Abcg2 forward,
5′-GCTGTGGAGCTGTTCGTAGTGG-3′ and reverse,
5′-GCTAAAGTACTGAAGCCAGGAC-3′; Nanog forward,
5′-CCCTTCCCTCGCCATCACA-3′ and reverse,
5′-ACCGCTTGCACTTCATCCTTTG-3′; Tuj1 forward,
5′-TAGACCCCAGCGGCAACTAT-3′ and reverse,
5′-GTTCCAGGTTCCAAGTCCACC-3′; and glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) forward, 5′-AACGGGAAGCCCATCACC-3′ and
reverse, 5′-CAGCCTTGGCAGCACCAG-3′. Primers were synthesized by
Invitrogen. Transcripts of GAPDH were amplified and used as an
internal reference. Quantitative PCR (qPCR) was performed using 2
µl of SYBR-Green/ROX qPCR Master Mix (Suzhou GenePharma Co.,
Ltd., Suzhou, China). All procedures were subjected to a TaqMan
miRNA assay (Applied Biosystem 7500) according to the
manufacturer's instructoins. U6 was employed as an endogenous
control (for miRNAs) to normalize data using the 2−ΔΔCt
method. Probe and primers specific for miR-124, miR-132, miR-134,
miR-20a, miR-17-5p, miR-30a-5p, or for U6 RNA internal control were
purchased from LC Sciences (Hangzhou, China). PCR and qPCR analyses
of at least 3 independent cultures were performed.
Western blot analysis
For protein extraction, the cells were washed twice
and resuspended in lysis buffer containing 50 mM of Tris-HCl, pH
7.4, 150 mM of NaCl, 1% NP-40, 10% glycerol, 1 mM of sodium
orthovanadate, 1 mM of PMSF and a protease inhibitor cocktail
(Shanghai Roche Pharmaceutical Co. Ltd., Shanghai, China).
Subsequently, 20 µg of cell lysates were separated by 8–15%
of resolving sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE), transferred onto polyvinylidene
fluoride membranes, and immunoblotted with antibodies according to
the manufacturer's instructions. The antibodies used in western
blot analysis included β-actin (sc-47778; 1:3,000; Santa Cruz
Biotechnology, Inc.), Tuj1 (MO15013; 1:1,000, Neuromics), TrkB
(ab18987; 1:1,000; Abcam) and Cdc42 (sc-8401; 1:1,000; Santa Cruz
Biotechnology, Inc.).
Electroporation
RNA oligoribonucleotides for miR-124 mimics (which
mimic endogenous mature miR-124 molecules), non-specific miRNA
(negative control) and miR-124 inhibitor [small interfering RNA
(siRNA) targeting miR-124 (miR-124 siRNA)] were all purchased from
Genepharma (Shanghai, China). For electroporation, mouse inner ear
NSCs were dissociated into a single-cell suspension with 0.25%
trypsin (Gibco/Thermo Fisher Scientific) at 37°C for 3 min.
Subsequently, 1×106 cells were electroporated with 5 g
of miR-124 mimics, non-specific miRNA or miR-124 siRNA using an
Amaxa Mouse NSC Nucleofector kit (Lonza Group Ltd., Basel,
Switzerland) following the manufacturer's instructions. RNA
oligoribonucleotides specific for enhanced green fluorescent
protein (eGFP) were used to monitor electroporation efficiency.
Electroporated cells were plated on cover glasses coated with
poly-L-lysine and laminin (R&D Systems, Inc.), and incubated in
differentiation medium at 37°C. The transfected inner ear NSCs were
then subjected to neuronal differentiation for up to 3 days.
Measurement of neuron percentage and
neurite length
Transfected inner ear NSCs were fixed and
immunostained at 3 days post-differentiation. The neuron percentage
was determined as the number of cells stained positive for the
neuron-specific marker, Tuj1, divided by the number of cells
stained with Hoechst 33342 (the total number of differentiated
inner ear NSCs) from 15 randomly selected fields at ×20
magnification. All neurons within the image frame were counted.
Cells located in clumps were excluded. Images were acquired using
an Olympus microscope [Olympus IX71 Inverted Microscope and Olympus
FV1000 confocal microscope; Olympus Inc. (China), Guangzhou, China]
and quantitative analysis was carried out using ImageJ software
(each image of ×20 magnification generally captures at least 200
differentiated inner ear NSCs).
As previously described (35), we measured the entire length of
the longest neurite extending from 95 differentiated neurons
(identified by the presence of Tuj1), which was selected randomly
(using a 40× objective lens). Neurites that were not entirely in
the frame were excluded. Neurite lengths were determined using
ImageJ software. Processes that could not be distinguished from
others were eliminated.
Statistical analysis
One-way ANOVA and Chi-square tests were used to
analyze the data, and the differences between variables were
considered to be statistically significance when the value was
P<0.05. All data are expressed as the means ± SEM, unless
otherwise stated.
Results
Culture and differentiation of inner ear
NSCs
The cells were isolated from the mouse spiral
ganglion and were cultured for 5 days. The cells formed floating
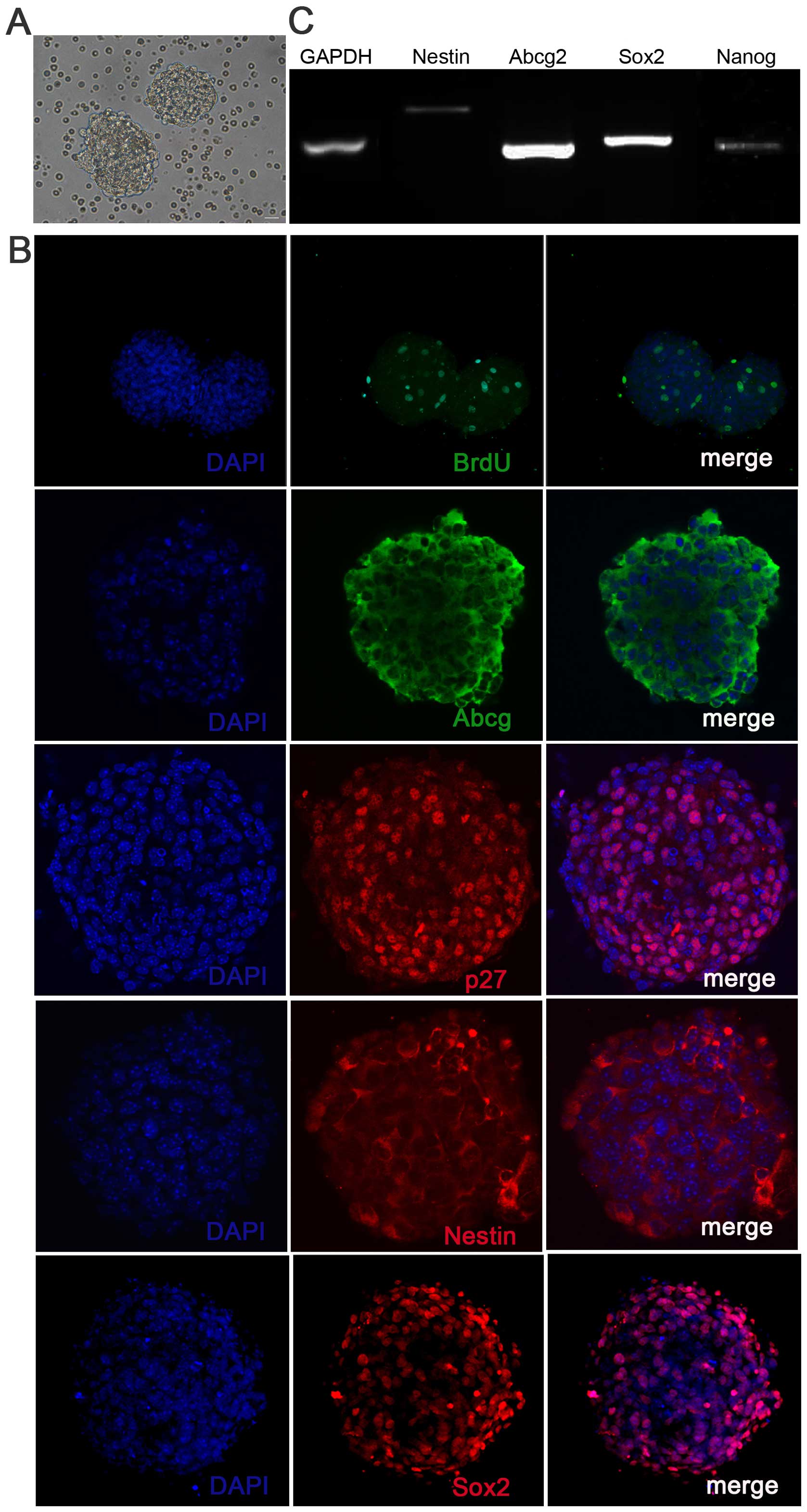
solid neurospheres (Fig. 1A),
which were positive for BrdU, Sox2 and other stem cell markers,
such as Nestin, p27Kip1 and Abcg2 (Fig. 1B), confirming the self-renewal and
proliferative capacity of these cells. To further characterize the
stemness properties of the inner ear neurospheres, we confirmed the
expression of markers of progenitor cells (Nestin,
p27Kip1, Abcg2 and Nanog) by RT-qPCR (Fig. 1C).
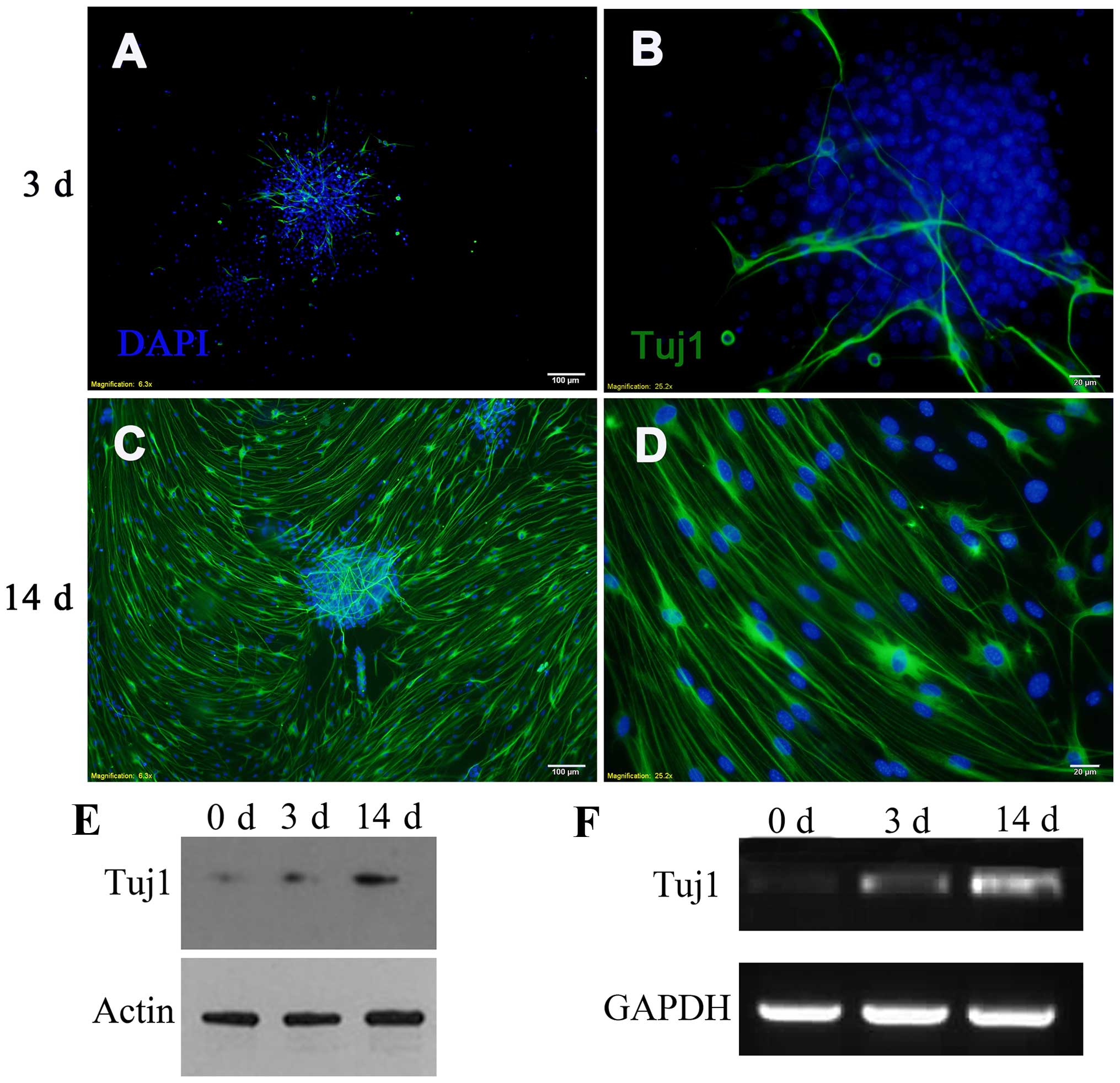
To initiate the differentiation of mouse inner ear
NSCs into neurons, we plated the neurospheres onto a coated
substratum in differentiation medium. Four hours after adherence,
differentiated neurons that expressed neuron-specific class III
β-tubulin (detected by Tuj1 antibody) began to sprout neurites from
the attached neurospheres. Immunofluorescence staining demonstrated
that the number of Tuj1-positive cells increased on day 3, and
reached maximal levels on day 14 (Fig. 2A–D). To confirm the
differentiation of NSCs, the expression of neuron-specific class
III β-tubulin was examined by western blot analysis and by RT-qPCR
(Fig. 2E–F). Proliferating
spheres expressed low levels of neuron-specific class III
β-tubulin, which is a marker of mature neurons. This phenomenon may
be related to the differentiation of individual peripheral NSCs.
During differentiation, the levels of neuron-specific class III
β-tubulin increased on day 3 and reached the highest levels on day
14, which was consistent with the data obtained by
immunofluorescence.
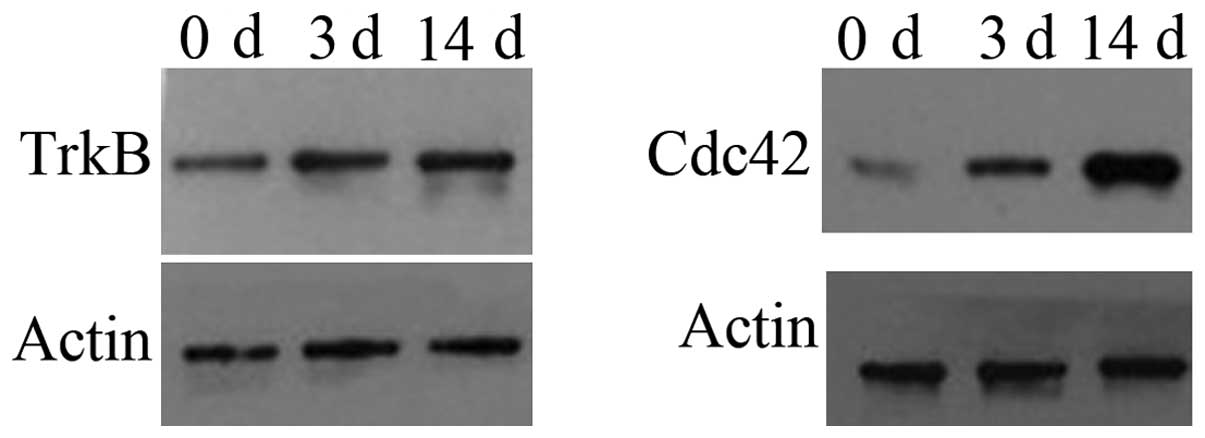
TrkB and Cdc42 are upregulated during the
neuronal differentiation of inner ear NSCs
Although Cdc42 and TrkB have been shown to affect
neuronal differentiation and polarity (36–40), the role of TrkB and Cdc42 in the
neuronal differentiation of mouse NSCs remains unknown. In this
study, we cultured inner ear NSCs in differentiation medium for 14
days; our results revealed that TrkB and Cdc42 expression were low
in the undifferentiated inner ear NSCs, and their expression
gradually increased during differentiation (Fig. 3C–D). Since TrkB and Cdc42 modulate
neuronal differentiation and neurite extension, TrkB and Cdc42 may
be involved in the differentiation of inner ear NSCs into
neurons.
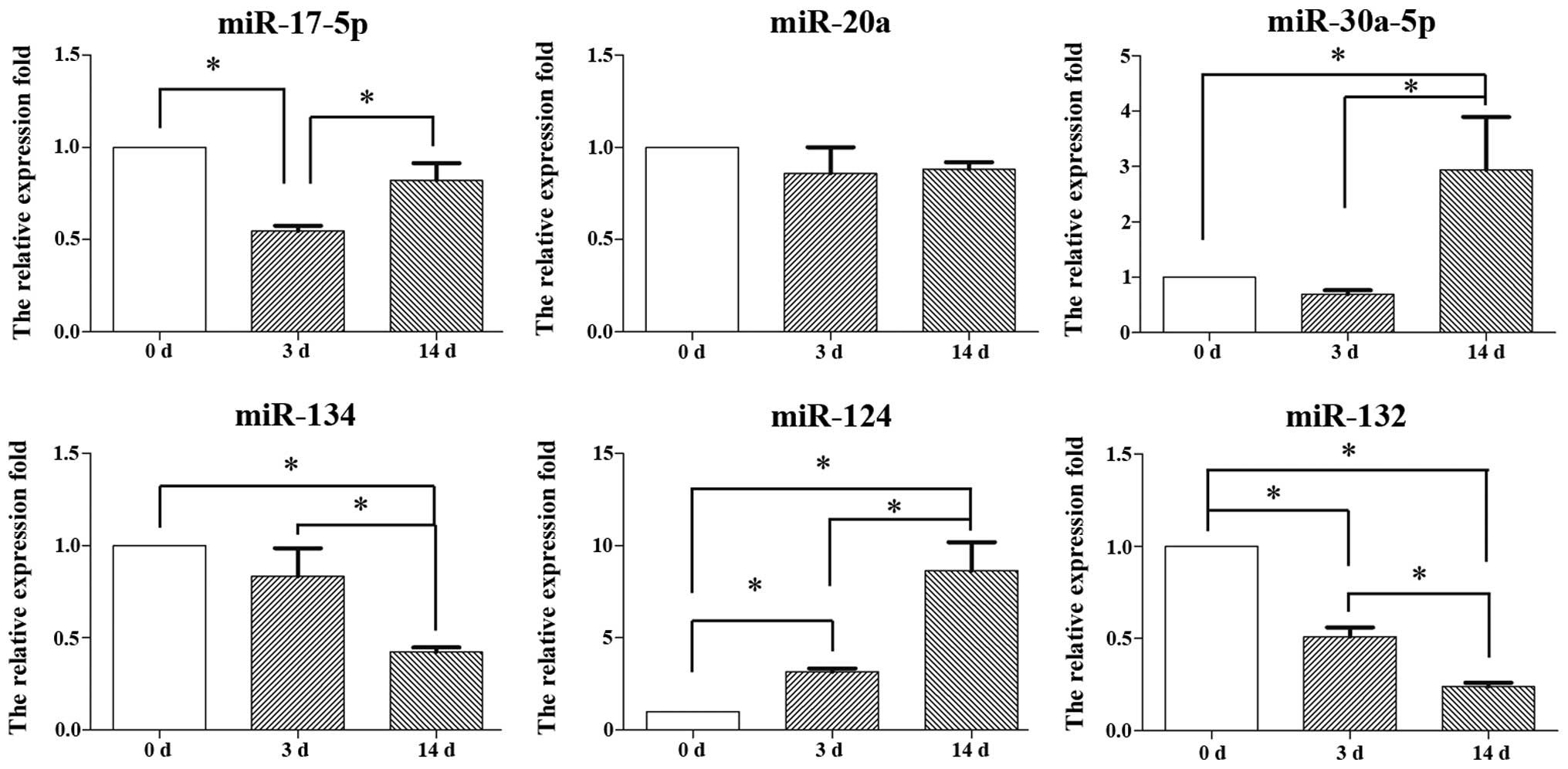
Expression of miRNAs during the
differentiation of inner ear NSCs
To determine the function of miRNAs in the neuronal
differentiation of inner ear NSCs, we compared the expression
levels of 6 miRNAs between undifferentiated and differentiated
inner ear NSCs. A total of 6 miRNAs were selected based on their
established role in neuronal differentiation.
We set the expression level in undifferentiated
inner ear NSCs as the baseline, and compared it to the expression
in differentiated inner ear NSCs on days 3 and 14 using the
2−ΔΔCt method. The levels of miR-17-5p decreased by day
3 (p<0.05) and increased by day 14 (p<0.05); however, the
overall expression level of miR-17-5p did not show any significant
change over the 14-day period (Fig.
4). Similarly, no significant change in miR-20a expression
levels were observed during neuronal differentiation (p>0.05).
By contrast, the miR-30a-5p expression levels increased and the
miR-134 levels significantly decreased during the differentiation
of NSCs on day 14, suggesting that miR-30a-5p and miR-134 play a
role in neuronal differentiation. The expression levels of 2
miRNAs, miR-124 and miR-132 were altered on days 3 and 14 of
differentiation. The levels of miR-124 increased 6-fold, while the
levels of miR-132 decreased 4-fold during the entire 14-day period.
In this study, we focused on the role of miR-124 in the
differentiation of inner ear NSCs.
miR-124 promotes the neuronal
differentiation of and neurite outgrowth in inner ear NSCs
Inner ear NSCs were transfected with RNA
oligoribonucleotides for miR-124 mimics, non-specific miRNA and
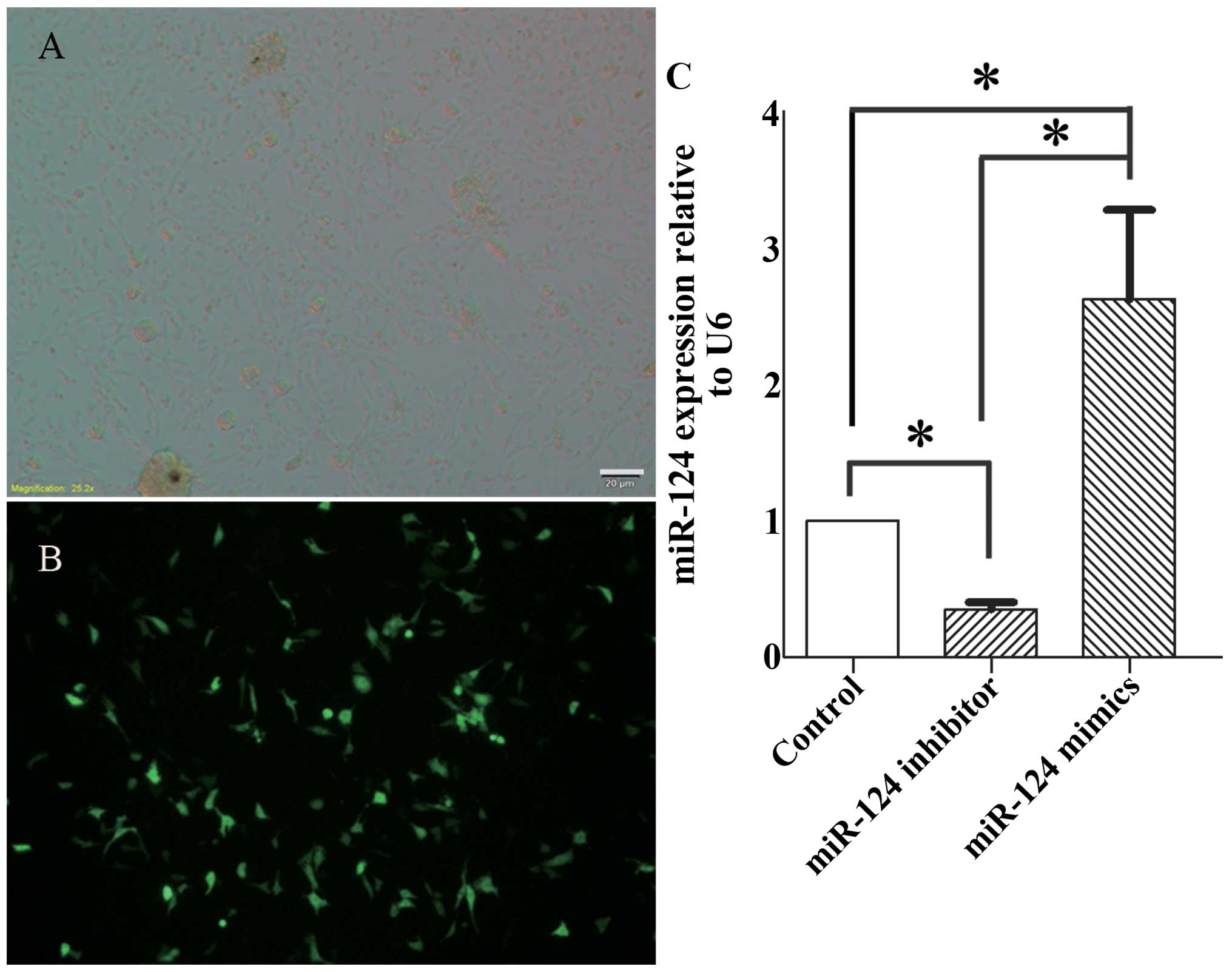
miR-124 inhibitor. The transfection efficiency was 50–60%, as
determined by eGFP (Fig. 5A and
B). In addition, we measured the miR-124 expression levels in
differentiated inner ear NSCs by RT-qPCR. Three days following
electroporation with miR-124 inhibitor, the miR-124 levels were
decreased by 2.13-fold. By contrast, the electroporation of miR-124
mimics led to a 3.02-fold increase in miR-124 expression (Fig. 5C).
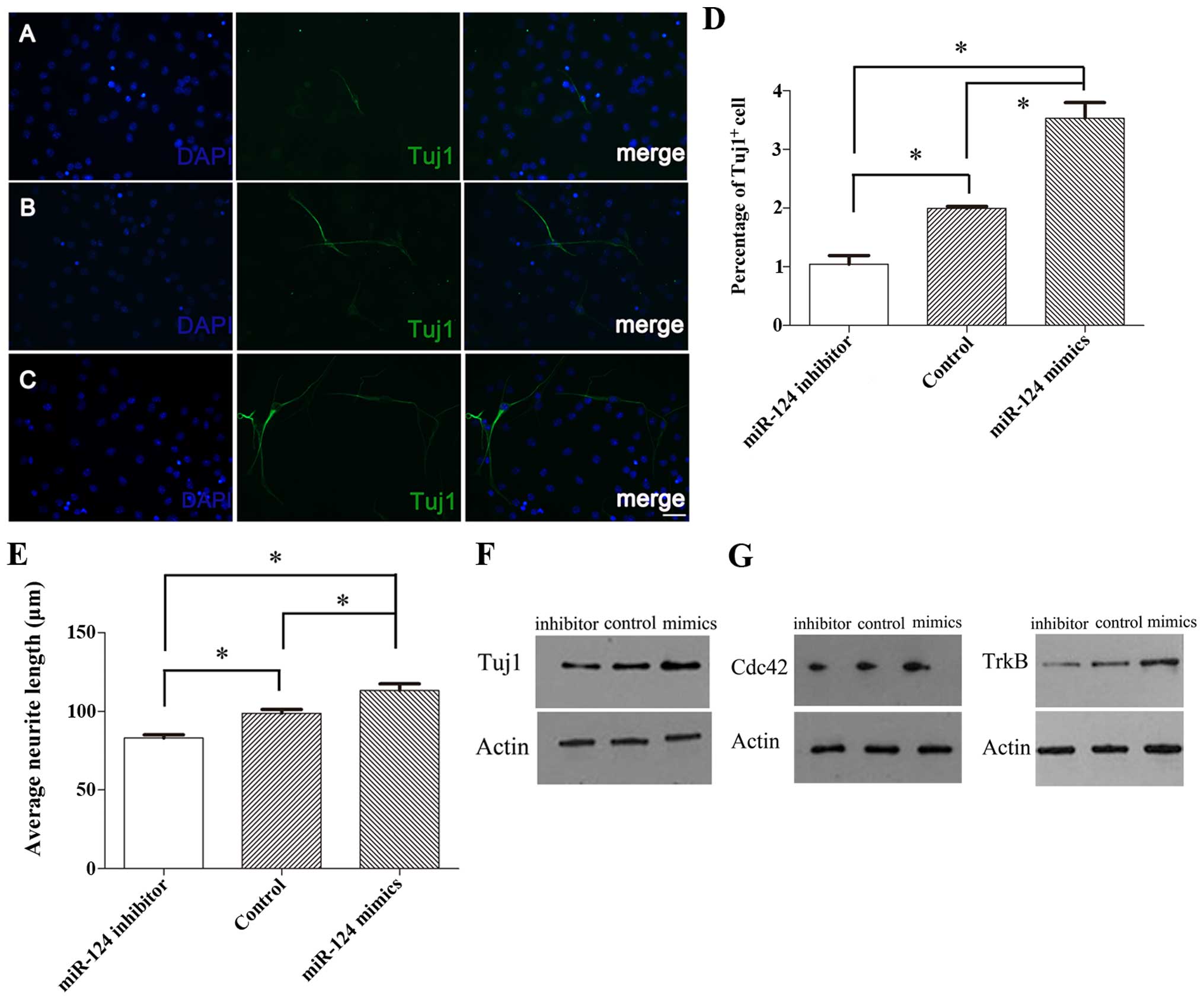
Next, we determined the effect of miR-124 on the
percentage of neuron and neurite outgrowth. We found that the
overexpression of miR-124 in differentiated inner ear NSCs
increased neuronal differentiation and neurite outgrowth, whereas
the knockdown of miR-124 had an opposite effect (Fig. 6A–C). The results revealed that at
3 days of differentiation, 3.53±0.46% of the cells transfected with
miR-124 mimics were Tuj1-positive cells, while the percentage of
Tuj1-positive cells in the controls was 1.99±0.06% (p<0.05;
Fig. 6D). Moreover, the average
neurite length of the Tuj1-positive cells transfected with the
miR-124 mimics was 113.22±7.31 µm, which was significantly
higher than that of the control cells (98.83±4.18 µm;
p<0.05) (Fig. 6E). By
contrast, 1.05±0.25% of the cells transfected with miR-124
inhibitor were Tuj1-positive, while the percentage of Tuj1-positive
cells in the controls was 1.99±0.06% (p<0.05; Fig. 6D). Additionally, the average
neurite length of the Tuj1-positive cells transfected with miR-124
inhibitor was reduced to 83.12±3.40 µm, which was
significantly lower than that of the controls (98.83±4.18
µm; p<0.05) (Fig.
6E).
We confirmed that inner ear NSCs transfected with
miR-124 mimics had higher levels of neuron-specific class III
β-tubulin compared to the control cells, while inner ear NSCs
transfected with miR-124 inhibitor had a reduced expression of
neuron-specific class III β-tubulin (Fig. 6F). These findings are in agreement
with the immunofluorescence observations and confirmed the role of
miR-124 in neuronal differentiation.
miR-124 affects the expression of TrkB
and Cdc42 in the neuronal differentiation of mouse inner ear
NSCs
TrkB and Cdc42 play a role in neuronal
differentiation and neurite elongation. We demonstrated that Cdc42
and TrkB were upregulated during neuronal differentiation. Our
findings revealed that miR-124 promoted the differentiation of
inner ear NSCs, which led us to investigate whether miR-124
modulates TrkB and Cdc42 expression in inner ear NSCs. We evaluated
the TrkB and Cdc42 expression levels in inner NSCs transfected with
miR-124 by western blot analysis (Fig. 6G). Compared to the control cells,
miR-124 overexpression increased Cdc42 and TrkB expression during
neuronal differentiation. Conversely, miR-124 knockdown caused a
marked decrease in the TrkB and Cdc42 protein levels, suggesting
that Cdc42 and TrkB act as downstream effectors of miR-124 during
inner ear NSC differentiation.
Discussion
NSCs are neural progenitor cells that have the
ability to self-renew, proliferate and differentiate into lineages,
such as neurons, oligodendroglia and astroglia (41). Thus, NSCs, which have regenerative
potential may serve as an important source for replacing damaged
neurons in hearing-impaired individuals. It is therefore important
to understand the differentiation pathway of inner ear NSCs into
neurons. While miRNAs are generally important in neuronal
development and differentiation (19,20), the profiling of miRNAs in the
inner ear has been a major research goal (27,28,42,43). Conditional Dicer knockout mice
have major defects in the inner ear, including hair cell
malformations, abnormal innervations, disrupted neurosensory
development and hearing loss (29,33). A role of miR-96 in progressive
hearing loss in mice and humans has been reported. Furthermore,
miR-181a influences proliferation within the basilar papilla
(44), while miR-200b can cause
severe malformations of the auditory and vestibular labyrinth by
affecting the Zeb1 pathway (45).
miR-183 may affect zebrafish inner ear organization (31), as well as mouse inner ear function
(46,47). However, the role of miRNAs in the
neuronal differentiation of mouse inner ear NSCs warrants further
investigation.
In this study, we examined the expression of 6
miRNAs in the differentiation of mouse inner ear NSCs into neurons.
In this study, the levels of miR-30a-5p, an established inhibitor
of BDNF expression (48),
increased 14 days after neuronal differentiation, suggesting that
miR-30a-5p plays a role in the late stages of neuronal
differentiation or neurite plasticity. In contrast to previously
published results (48–53), miR-134 and miR-132 were
downregulated during the neuronal differentiation of inner ear
NSCs. Although miR-20a has been shown to be downregulated in the
neuronal lineage differentiation of unrestricted somatic stem cells
(USSCs) from cord blood and NSCs (54,55), no changes in miR-20a were observed
during differentiation of NSCs in our study. In addition, in our
study, miR-17-5p expression increased during the first 3 days of
differentiation and then increased on day 14; however, in a
previous study, its expression decreased continued to decrease in
differentiated SH-SY5Y cells (21). The differential expression of
specific miRNAs may be due to specific neuronal types and time
points being investigated.
In this study, we focused on the role of miR-124,
which has been shown to enhance the development of mature neurons
(56,57) and to play a role in the neuronal
differentiation of several cell lines in vivo and in
vitro (16,22,23,25,58). Moreover, miR-124 acts as a key
mediator in regulating the differentiation of human embryonic and
mesenchymal stem cells (59,60). We demonstrated that miR-124 was
detectable at low levels in the undifferentiated inner ear NSCs;
however, its expression gradually increased and peaked on day 14 of
differentiation. The overexpression of miR-124 increased the
percentages of neurons and neurite length, whereas the knockdown of
miR-124 had an opposite effect, demonstrating that miR-124 is
important for the in vitro neuronal differentiation of inner
ear NSCs.
Cdc42 is a member of the Rho GTPases and regulates
both microtubules and actin filaments in a variety of cells
(61–63), and plays a role in the
establishment of neuronal polarity (36,37,39,64). In addition, TrkB, the cognate
receptor of BDNF, participates in multiple aspects of neuronal
functions (65), modulating
neurite outgrowth, structural plasticity and survival (36,38–40). Cdc42 functions downstream of BDNF
stimulation (36,40,66,67). Furthermore, BDNF activates Cdc42
via the Cdk5-mediated phosphorylation of TrkB, which in turn
influences dendritic growth (36). TrkB and Cdc42 play a role in
neural development and plasticity. The pregulation of TrkB and
Cdc42 expression during inner ear NSC differentiation suggests that
TrkB acts upstream of Cdc42 in the regulation of the neuronal
differentiation of and neurite outgrowth in mouse inner ear
NSCs.
Recently, several studies have examined the function
of Cdc42 in dendrite and axon development, as well as its
regulation by miR-124 (58,68). It has been shown that miR-124
upregulates the expression of Cdc42 (24). Moreover, an increased TrkB level
of has been shown to be accompanied by the upregulation of miR-124
in exercise-dependent neuronal differentiation in the hippocampus
of adult male rats (69).
In this study, we demonstrated that miR-124 promoted
neuronal differentiation and neurite outgrowth, and we investigated
the effects of miR-124 on TrkB and Cdc42 expression during the
neuronal differentiation of and neurite outgrowth in inner ear
NSCs. We demonstrated that TrkB and Cdc42 expression coincided with
miR-124 during inner ear NSC differentiation. These results suggest
that TrkB and Cdc42 may play an important role in miR-124-regulated
neuronal differentiation. In support of this hypothesis, we found
that miR-124 regulates their expression, raising the possibility
that the effects of miR-124 on inner ear NSC differentiation and
neurite outgrowth are mediated by TrkB and Cdc42.
Although TrkB promotes neuronal survival and
plasticity (36,38–40), Cdc42 may have opposite effects on
neurite outgrowth. Several studies have revealed that Cdc42
enhances dendritic growth or arborization (36,37,39), while other studies have
demonstrated negative effects (58,68,70). These studies are consistent with
our hypothesis, wherein Cdc42 promotes neurite outgrowth. In the
inner ear, TrkB is enriched in cochlear neurons and plays a role in
neuronal survival and structural plasticity (38,71,72). In contrast to our findings and the
results reported by Brors et al (73), Rac/Cdc42 inhibition has been shown
to enhance neurite formation and decrease neurite length in
cochlear spiral ganglion explants in neonatal rat pups (74).
In conclusion, our results suggest that miR-124
promotes the neuronal differentiation of and neurite outgrowth in
inner ear NSCs by regulating TrkB and Cdc42. Future studies are
required to determine the mechanisms through which miR-124 controls
TrkB and Cdc42 expression to regulate neuronal differentiation in
the mouse inner ear.
Acknowledgments
The study was supported by grants from the National
Natural Science fund of China (no. 81200748).
References
|
1
|
Spoendlin H: Retrograde degeneration of
the cochlear nerve. Acta Otolaryngol. 79:266–275. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hawkins JE Jr: Comparative otopathology:
aging, noise, and ototoxic drugs. Adv Otorhinolaryngol. 20:125–141.
1973.PubMed/NCBI
|
|
3
|
Zhao C, Deng W and Gage FH: Mechanisms and
functional implications of adult neurogenesis. Cell. 132:645–660.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Oshima K, Grimm CM, Corrales CE, Senn P,
Martinez Monedero R, Géléoc GS, Edge A, Holt JR and Heller S:
Differential distribution of stem cells in the auditory and
vestibular organs of the inner ear. J Assoc Res Otolaryngol.
8:18–31. 2007. View Article : Google Scholar
|
|
5
|
Oshima K, Senn P and Heller S: Isolation
of sphere-forming stem cells from the mouse inner ear. Methods Mol
Biol. 493:141–162. 2009. View Article : Google Scholar
|
|
6
|
Rask-Andersen H, Boström M, Gerdin B,
Kinnefors A, Nyberg G, Engstrand T, Miller JM and Lindholm D:
Regeneration of human auditory nerve. In vitro/in video
demonstration of neural progenitor cells in adult human and guinea
pig spiral ganglion. Hear Res. 203:180–191. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gu S, Jin L, Zhang F, Sarnow P and Kay MA:
Biological basis for restriction of microRNA targets to the 3′
untranslated region in mammalian mRNAs. Nat Struct Mol Biol.
16:144–150. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lai EC: MicroRNAs are complementary to 3′
UTR sequence motifs that mediate negative post-transcriptional
regulation. Nat Genet. 30:363–364. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Valencia-Sanchez MA, Liu J, Hannon GJ and
Parker R: Control of translation and mRNA degradation by miRNAs and
siRNAs. Genes Dev. 20:515–524. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vaucheret H: Post-transcriptional small
RNA pathways in plants: Mechanisms and regulations. Genes Dev.
20:759–771. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Darnell DK, Kaur S, Stanislaw S, Konieczka
JH, Yatskievych TA and Antin PB: MicroRNA expression during chick
embryo development. Dev Dyn. 235:3156–3165. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Deo M, Yu JY, Chung KH, Tippens M and
Turner DL: Detection of mammalian microRNA expression by in situ
hybridization with RNA oligonucleotides. Dev Dyn. 235:2538–2548.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kapsimali M, Kloosterman WP, de Bruijn E,
Rosa F, Plasterk RH and Wilson SW: MicroRNAs show a wide diversity
of expression profiles in the developing and mature central nervous
system. Genome Biol. 8:R1732007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Krichevsky AM, Sonntag KC, Isacson O and
Kosik KS: Specific microRNAs modulate embryonic stem cell-derived
neurogenesis. Stem Cells. 24:857–864. 2006. View Article : Google Scholar
|
|
17
|
Mansfield JH, Harfe BD, Nissen R, Obenauer
J, Srineel J, Chaudhuri A, Farzan-Kashani R, Zuker M, Pasquinelli
AE, Ruvkun G, et al: MicroRNA-responsive 'sensor' transgenes
uncover Hox-like and other developmentally regulated patterns of
vertebrate microRNA expression. Nat Genet. 36:1079–1083. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nelson PT, Baldwin DA, Kloosterman WP,
Kauppinen S, Plasterk RH and Mourelatos Z: RAKE and LNA-ISH reveal
microRNA expression and localization in archival human brain. RNA.
12:187–191. 2006. View Article : Google Scholar :
|
|
19
|
Fineberg SK, Kosik KS and Davidson BL:
MicroRNAs potentiate neural development. Neuron. 64:303–309. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gao FB: Context-dependent functions of
specific microRNAs in neuronal development. Neural Dev. 5:252010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Beveridge NJ, Tooney PA, Carroll AP, Tran
N and Cairns MJ: Down-regulation of miR-17 family expression in
response to retinoic acid induced neuronal differentiation. Cell
Signal. 21:1837–1845. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hohjoh H and Fukushima T: Marked change in
microRNA expression during neuronal differentiation of human
teratocarcinoma NTera2D1 and mouse embryonal carcinoma P19 cells.
Biochem Biophys Res Commun. 362:360–367. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Le MT, Xie H, Zhou B, Chia PH, Rizk P, Um
M, Udolph G, Yang H, Lim B and Lodish HF: MicroRNA-125b promotes
neuronal differentiation in human cells by repressing multiple
targets. Mol Cell Biol. 29:5290–5305. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Makeyev EV, Zhang J, Carrasco MA and
Maniatis T: The MicroRNA miR-124 promotes neuronal differentiation
by triggering brain-specific alternative pre-mRNA splicing. Mol
Cell. 27:435–448. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Visvanathan J, Lee S, Lee B, Lee JW and
Lee SK: The microRNA miR-124 antagonizes the anti-neural REST/SCP1
pathway during embryonic CNS development. Genes Dev. 21:744–749.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sacheli R, Nguyen L, Borgs L, Vandenbosch
R, Bodson M, Lefebvre P and Malgrange B: Expression patterns of
miR-96, miR-182 and miR-183 in the development inner ear. Gene Expr
Patterns. 9:364–370. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang XR, Zhang XM, Zhen J, Zhang PX, Xu G
and Jiang H: MicroRNA expression in the embryonic mouse inner ear.
Neuroreport. 21:611–617. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Weston MD, Pierce ML, Rocha-Sanchez S,
Beisel KW and Soukup GA: MicroRNA gene expression in the mouse
inner ear. Brain Res. 1111:95–104. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Friedman LM, Dror AA, Mor E, Tenne T,
Toren G, Satoh T, Biesemeier DJ, Shomron N, Fekete DM, Hornstein E
and Avraham KB: MicroRNAs are essential for development and
function of inner ear hair cells in vertebrates. Proc Natl Acad Sci
USA. 106:7915–7920. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lewis MA, Quint E, Glazier AM, Fuchs H, De
Angelis MH, Langford C, van Dongen S, Abreu-Goodger C, Piipari M,
Redshaw N, et al: An ENU-induced mutation of miR-96 associated with
progressive hearing loss in mice. Nat Genet. 41:614–618. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li H, Kloosterman W and Fekete DM:
MicroRNA-183 family members regulate sensorineural fates in the
inner ear. J Neurosci. 30:3254–3263. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mencía A, Modamio-Høybjør S, Redshaw N,
Morín M, Mayo-Merino F, Olavarrieta L, Aguirre LA, del Castillo I,
Steel KP, Dalmay T, et al: Mutations in the seed region of human
miR-96 are responsible for nonsyndromic progressive hearing loss.
Nat Genet. 41:609–613. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Soukup GA, Fritzsch B, Pierce ML, Weston
MD, Jahan I, McManus MT and Harfe BD: Residual microRNA expression
dictates the extent of inner ear development in conditional Dicer
knockout mice. Dev Biol. 328:328–341. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Martinez-Monedero R, Yi E, Oshima K,
Glowatzki E and Edge AS: Differentiation of inner ear stem cells to
functional sensory neurons. Dev Neurobiol. 68:669–684. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Roehm PC, Xu N, Woodson EA, Green SH and
Hansen MR: Membrane depolarization inhibits spiral ganglion neurite
growth via activation of multiple types of voltage sensitive
calcium channels and calpain. Mol Cell Neurosci. 37:376–387. 2008.
View Article : Google Scholar
|
|
36
|
Cheung ZH, Chin WH, Chen Y, Ng YP and Ip
NY: Cdk5 is involved in BDNF-stimulated dendritic growth in
hippocampal neurons. PLoS Biol. 5:e632007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Endo M, Antonyak MA and Cerione RA:
Cdc42-mTOR signaling pathway controls Hes5 and Pax6 expression in
retinoic acid-dependent neural differentiation. J Biol Chem.
284:5107–5118. 2009. View Article : Google Scholar
|
|
38
|
Sciarretta C, Fritzsch B, Beisel K,
Rocha-Sanchez SM, Buniello A, Horn JM and Minichiello L:
PLCγ-activated signalling is essential for TrkB mediated sensory
neuron structural plasticity. BMC Dev Biol. 10:1032010. View Article : Google Scholar
|
|
39
|
Sosa L, Dupraz S, Laurino L, Bollati F,
Bisbal M, Cáceres A, Pfenninger KH and Quiroga S: IGF-1 receptor is
essential for the establishment of hippocampal neuronal polarity.
Nat Neurosci. 9:993–995. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lai KO, Wong AS, Cheung MC, Xu P, Liang Z,
Lok KC, Xie H, Palko ME, Yung WH, Tessarollo L, et al: TrkB
phosphorylation by Cdk5 is required for activity-dependent
structural plasticity and spatial memory. Nat Neurosci.
15:1506–1515. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gage FH: Mammalian neural stem cells.
Science. 287:1433–1438. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Elkan-Miller T, Ulitsky I, Hertzano R,
Rudnicki A, Dror AA, Lenz DR, Elkon R, Irmler M, Beckers J, Shamir
R and Avraham KB: Integration of transcriptomics, proteomics, and
microRNA analyses reveals novel microRNA regulation of targets in
the mammalian inner ear. PLoS One. 6:e181952011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Patel M, Cai Q, Ding D, Salvi R, Hu Z and
Hu BH: The miR-183/Taok1 target pair is implicated in cochlear
responses to acoustic trauma. PLoS One. 8:e584712013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Frucht CS, Santos-Sacchi J and Navaratnam
DS: MicroRNA181a plays a key role in hair cell regeneration in the
avian auditory epithelium. Neurosci Lett. 493:44–48. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hertzano R, Elkon R, Kurima K, Morrisson
A, Chan SL, Sallin M, Biedlingmaier A, Darling DS, Griffith AJ,
Eisenman DJ and Strome SE: Cell type-specific transcriptome
analysis reveals a major role for Zeb1 and miR-200b in mouse inner
ear morphogenesis. PLoS Genet. 7:e10023092011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Gu C, Li X, Tan Q, Wang Z, Chen L and Liu
Y: MiR-183 family regulates chloride intracellular channel 5
expression in inner ear hair cells. Toxicol In Vitro. 27:486–491.
2013. View Article : Google Scholar
|
|
47
|
Wang XR, Zhang XM, Du J and Jiang H:
MicroRNA-182 regulates otocyst-derived cell differentiation and
targets T-box1 gene. Hear Res. 286:55–63. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Mellios N, Huang HS, Grigorenko A, Rogaev
E and Akbarian S: A set of differentially expressed miRNAs,
including miR-30a-5p, act as post-transcriptional inhibitors of
BDNF in prefrontal cortex. Hum Mol Genet. 17:3030–3042. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Gaughwin P, Ciesla M, Yang H, Lim B and
Brundin P: Stage-specific modulation of cortical neuronal
development by Mmu-miR-134. Cereb Cortex. 21:1857–1869. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Mellios N, Sugihara H, Castro J, Banerjee
A, Le C, Kumar A, Crawford B, Strathmann J, Tropea D, Levine SS, et
al: miR-132, an experience-dependent microRNA, is essential for
visual cortex plasticity. Nat Neurosci. 14:1240–1242. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Clovis YM, Enard W, Marinaro F, Huttner WB
and De Pietri Tonelli D: Convergent repression of Foxp2 3′UTR by
miR-9 and miR-132 in embryonic mouse neocortex: Implications for
radial migration of neurons. Development. 139:3332–3342. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Lin LF, Chiu SP, Wu MJ, Chen PY and Yen
JH: Luteolin induces microRNA-132 expression and modulates neurite
outgrowth in PC12 cells. PLoS One. 7:e433042012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Pathania M, Torres-Reveron J, Yan L,
Kimura T, Lin TV, Gordon V, Teng ZQ, Zhao X, Fulga TA, Van Vactor D
and Bordey A: miR-132 enhances dendritic morphogenesis, spine
density, synaptic integration, and survival of newborn olfactory
bulb neurons. PLoS One. 7:e381742012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Aranha MM, Santos DM, Xavier JM, Low WC,
Steer CJ, Solá S and Rodrigues CM: Apoptosis-associated microRNAs
are modulated in mouse, rat and human neural differentiation. BMC
Genomics. 11:5142010. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Trompeter HI, Abbad H, Iwaniuk KM, Hafner
M, Renwick N, Tuschl T, Schira J, Müller HW and Wernet P: MicroRNAs
MiR-17, MiR-20a, and MiR-106b act in concert to modulate E2F
activity on cell cycle arrest during neuronal lineage
differentiation of USSC. PLoS One. 6:e161382011. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Miska EA, Alvarez-Saavedra E, Townsend M,
Yoshii A, Sestan N, Rakic P, Constantine-Paton M and Horvitz HR:
Microarray analysis of microRNA expression in the developing
mammalian brain. Genome Biol. 5:R682004. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Smirnova L, Gräfe A, Seiler A, Schumacher
S, Nitsch R and Wulczyn FG: Regulation of miRNA expression during
neural cell specification. Eur J Neurosci. 21:1469–1477. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Yu JY, Chung KH, Deo M, Thompson RC and
Turner DL: MicroRNA miR-124 regulates neurite outgrowth during
neuronal differentiation. Exp Cell Res. 314:2618–2633. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lee MR, Kim JS and Kim KS: miR-124a is
important for migratory cell fate transition during gastrulation of
human embryonic stem cells. Stem Cells. 28:1550–1559. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Maisel M, Habisch HJ, Royer L, Herr A,
Milosevic J, Hermann A, Liebau S, Brenner R, Schwarz J, Schroeder M
and Storch A: Genome-wide expression profiling and functional
network analysis upon neuroectodermal conversion of human
mesenchymal stem cells suggest HIF-1 and miR-124a as important
regulators. Exp Cell Res. 316:2760–2778. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Hall A: Rho GTPases and the actin
cytoskeleton. Science. 279:509–514. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Jaffe AB and Hall A: Rho GTPases:
Biochemistry and biology. Annu Rev Cell Dev Biol. 21:247–269. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Li E, Stupack D, Bokoch GM and Nemerow GR:
Adenovirus endocytosis requires actin cytoskeleton reorganization
mediated by Rho family GTPases. J Virol. 72:8806–8812.
1998.PubMed/NCBI
|
|
64
|
Schwamborn JC and Püschel AW: The
sequential activity of the GTPases Rap1B and Cdc42 determines
neuronal polarity. Nat Neurosci. 7:923–929. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Huang EJ and Reichardt LF: Trk receptors:
Roles in neuronal signal transduction. Annu Rev Biochem.
72:609–642. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Chen TJ, Gehler S, Shaw AE, Bamburg JR and
Letourneau PC: Cdc42 participates in the regulation of ADF/cofilin
and retinal growth cone filopodia by brain derived neurotrophic
factor. J Neurobiol. 66:103–114. 2006. View Article : Google Scholar
|
|
67
|
Shen W, Wu B, Zhang Z, Dou Y, Rao ZR, Chen
YR and Duan S: Activity-induced rapid synaptic maturation mediated
by presynaptic cdc42 signaling. Neuron. 50:401–414. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Franke K, Otto W, Johannes S, Baumgart J,
Nitsch R and Schumacher S: miR-124-regulated RhoG reduces neuronal
process complexity via ELMO/Dock180/Rac1 and Cdc42 signalling. EMBO
J. 31:2908–2921. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Mojtahedi S, Kordi MR, Hosseini SE, Omran
SF and Soleimani M: Effect of treadmill running on the expression
of genes that are involved in neuronal differentiation in the
hippo-campus of adult male rats. Cell Biol Int. 37:276–283. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Hing H, Xiao J, Harden N, Lim L and
Zipursky SL: Pak functions downstream of Dock to regulate
photoreceptor axon guidance in Drosophila. Cell. 97:853–863. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Schimmang T, Minichiello L, Vazquez E, San
Jose I, Giraldez F, Klein R and Represa J: Developing inner ear
sensory neurons require TrkB and TrkC receptors for innervation of
their peripheral targets. Development. 121:3381–3391.
1995.PubMed/NCBI
|
|
72
|
Schimmang T, Tan J, Müller M, Zimmermann
U, Rohbock K, Kôpschall I, Limberger A, Minichiello L and Knipper
M: Lack of Bdnf and TrkB signalling in the postnatal cochlea leads
to a spatial reshaping of innervation along the tonotopic axis and
hearing loss. Development. 130:4741–4750. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Brors D, Aletsee C, Dazert S, Huverstuhl
J, Ryan AF and Bodmer D: Clostridium difficile toxin B, an
inhibitor of the small GTPases Rho, Rac and Cdc42, influences
spiral ganglion neurite outgrowth. Acta Otolaryngol. 123:20–25.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Mullen LM, Pak KK, Chavez E, Kondo K,
Brand Y and Ryan AF: Ras/p38 and PI3K/Akt but not Mek/Erk signaling
mediate BDNF-induced neurite formation on neonatal cochlear spiral
ganglion explants. Brain Res. 1430:25–34. 2012. View Article : Google Scholar
|