Introduction
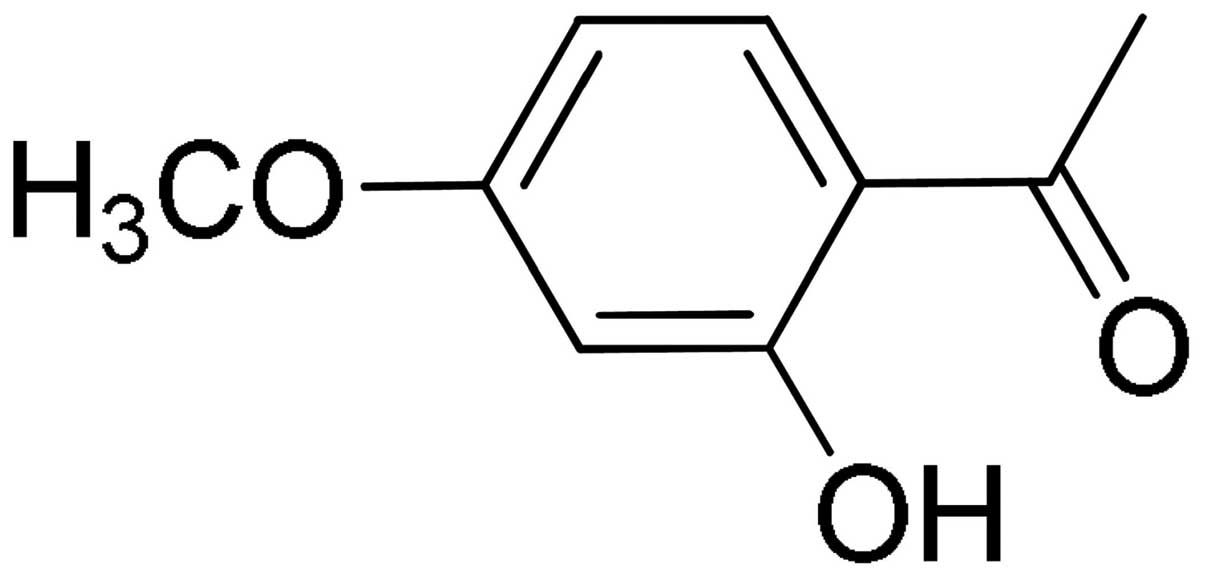
Paeonol (2′-hydroxy-4′-methoxyacetophenone)
(chemical structure shown in Fig.
1) is a main constituent of the root of Paeonia
Suffruticosa Andrews, which has long been used as an
anti-inflammatory agent in China (1,2).
We previously found that paeonol was effective in the prevention
and treatment of atherosclerosis (AS) at the initiation and
developmental stages through the inhibition of the inflammatory
response in animal models. Paeonol inhibited vascular cell adhesion
molecule-1 (VCAM-1) expression to suppress monocyte adhesion to
tumor necrosis factor-α (TNF-α)-stimulated vascular endothelial
cells (VECs) (3). However, the
underlying mechanisms are not yet well known. It has been
demonstrated that paeonol can suppress serum and aorta lipid
peroxidation in models of hyperlipidemia, and can decrease serum
oxidative modification induced by oxidized low-density lipoprotein
(ox-LDL) in healthy humans (4–6).
Paeonol has also been shown to inhibit the expression of
interleukin (IL)-1β and TNF-α to inhibit the over-proliferation of
vascular smooth muscle cells (VSMCs), and decrease local
inflammation in AS (7).
VCAM-1 belongs to the immunoglobulin superfamily and
is only expressed in VECs (8),
and it can be stimulated by cytokines, thus playing an important
role in the pathogenesis of AS (9,10).
MicroRNAs (miRNAs or miRs) are small non-coding RNAs that regulate
the post-transcription of target genes (11,12), and have been shown to be involved
in the pathogenesis of AS and to regulate the biological functions
of VECs (13). miRNA-126
(miR-126) is located in an intron of the epidermal growth factor
like domain-7, and is selectively expressed in VECs (14). The knockdown of miR-126 has been
shown to induce the proliferation and migration of VECs, which
results in endangium damage and angiogenesis-dependent disorders
(15). Notably, VCAM-1 is a
specific target gene of miR-126, and the inhibition of miR-126
expression leads to a higher expression of VCAM-1 in
TNF-α-stimulated VECs (11).
Moreover, phosphatidylinositol 3-kinase (PI3K) is one of the most
relevant players in VEC biological processes, and activates protein
kinase B (Akt) (16). The
transcription factor, nuclear factor-κB (NF-κB), participates in
cellular processes, such as cell adhesion (17).
However, it has been reported that the expression of
miRNAs can be regulated by natural compound (18). In this study, we examined the
effect of paeonol on miR-126 expression, and its ability to inhibit
monocyte adhesion to ox-LDL-injured VECs. Thus far, the underlying
mechanisms responsible for the regulatory effects of paeonol on
miR-126 expression have not yet been fully elucidated. Ox-LDL was
used for the stimulation of VECs, as its formation in the
subendothelial space of vascular wall is a key stage in the
pathogenesis of AS (19). Our
findings indicate that miR-126 may be an important target in
treatment of AS with the use of paeonol.
Materials and methods
Animals
At 6–8 weeks after birth, a total of 20 male
Sprague-Dawley rats (weighing 160–180 g) were obtained from the
Experimental Animal Center of Anhui Medical University (Hefei,
China). All rats were maintained under specific-free conditions in
the animal facility at the Anhui University of Chinese Medicine.
They were housed in a polypropylene cage (25±5°C, 40–50% relative
humidity) under controlled lighting (12 h light/dark cycle), and
allowed free access to food and water. All animal experiments were
carried out in accordance with the Guide for the Care and Use of
Laboratory Animals prepared by the National Academy of Sciences and
were approved by the Ethics Committee of Anhui University of
Chinese Medicine (Hefei, China).
Chemicals
Paeonol (purity, 99%) was obtained from Baicao
Plants Co., Ltd. (Xuancheng, China). Dulbecco's modified Eagle's
medium (DMEM) was purchased from Gibco Life Technologies Co. (Grand
Island, NY, USA). ox-LDL (YB-002-1) was purchased from Yiyuan
Biotechnologies Co., Ltd. (Guangzhou, China). HiPerFect
transfection reagent, miScript miR-126 mimic and inhibitor, the
miRNeasy mini kit (Cat. no. 217004) and the miScript PCR Starter
kit (Cat. no. 218193) were purchased from Qiagen Co., Ltd.
(Dusseldorf, Germany). The Dual-Luciferase Reporter kit was
purchased from Promega Biotech Co., Ltd. (Madison, WI, USA), and
the constructs of wild- and mutant-type 3′UTR of VCAM-1 mRNA were
obtained from Huaan Pingkang Biotechnology Co., Ltd. (Shen zhen,
China). The PI3K inhibitor, LY294002, and ammonium
pyrrolidinedithiocarbamate (PDTC), a selective NF-κB inhibitor,
were purchased from Beyotime Biotech, Inc. (Shanghai, China).
Antibody to p-PI3K (Cat. no. 4228L) was purchased from Cell
Signaling Technology, Inc. (Danvers, MA, USA). Antibodies to NF-κB
p65 (Cat. no. BS9879M), inhibitory factor κB-α (IκBα; Cat. no.
BS3601), p-IκBα (Cat. no. BS4736) and VCAM-1 (Cat. no. BS6005) were
purchased from Bioworld Technology Co., Ltd. (Louis Park, MN, USA).
Antibodies to PI3K (Cat. no. sc-48639), Akt (Cat. no. sc-24500),
p-Akt (Cat. no. sc-7985-R) were purchased from Santa Cruz
Biotechnology, Inc. (Santa Cruz, CA, USA). All other reagents were
purchased from reliable companies.
Isolation of rat VECs
The VECs were isolated from the rat thoracic aorta
as follows: the rats were sacrificed by euthanasia. The thoracic
aortas were removed from the rats using surgical scissors, and the
clean blood vessels were obtained from stripped connective and
fatty tissues under sterile conditions, reversed and stringed at
both ends tightly. The vascular intima was washed with DMEM
supplemented with 10% fetal bovine serum (FBS), and incubated to
digestion at 37°C, and 5% CO2 for 1 h in 0.2%
collagenase I and rinsed using medium. After cutting off, the
remaining aorta was cut into sections, and the tissue fragments
were incubated under the same conditions. Once the cells had formed
a monolayer, the explants were removed, and the cells were
subcultured. The cells were identified by their endothelial
cell-specific, cobblestone-like morphology, and the cells were
determined by the immunocytochemical staining for Factor VIII
(Boster, Wuhan, China). The cells were collected by centrifugation
and resuspended in DMEM containing 10% FBS for the investigation.
In all experiments, the VECs were used at passages 3–5.
3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide
(MTT) assay
The VECs were seeded in 96-well plates at a density
of 1×104 cells/well in 200 µl DMEM over night to
allow attachment. The cells were then treated with fresh medium
containing various concentrations of 5, 10, 20, 40 and 80 mg/l of
ox-LDL and cultured for 12, 24 and 48 h, or pre-treated with
various concentrations of 15, 30, 60, 120, 240 and 480 µM of
paeonol and then cultured for 12, 24 and 48 h as above. MTT
solution was added to each well followed by incubation for 4 h. The
growth medium was then removed and replaced with dimethylsulfoxide
(DMSO), followed by incubation for 10 min. A microplate reader
(SpectraMax Mze; Molecular Devices, Sunnyvale, CA, USA) was used to
measure the absorbance (A) at 490 nm. Cell viability was
calculated as follows: relative viability (%) =
A490(test) −
A490(normal)/A490(control) −
A490(normal) ×100.
Transient transfection
The VECs were seeded in 6-well plates at a density
of 1×105 cells/well in the presence of 2 ml DMEM
containing 10% FBS, and incubated under normal conditions
(typically 37°C and 5% CO2). HiPerFect transfection
reagent (12 µl) was added to the culture medium without
serum to a volume of 300 µl. Subsequently, 0.6 µl of
the miR-126 mimic (20 µM stock solution) or 6 µl of
the miR-126 inhibitor (20 µM stock solution) were added to
these mixtures, and mixed by vortexing. The complexes were added to
the VEC medium and gently swirled. The VECs were then incubated
with the transfection complexes for 24 h, and these media were
changed as required.
Quantitative polymerase chain reaction
(qPCR)
Total RNA was extracted from the VECs using the
Qiazol lysis reagent (Qiagen, Dusseldorf, Germany). cDNA prepared
in a transcript reaction using miScript PCR Starter kit (Qiagen)
served as the template for qPCR analysis. The reverse transcription
conditions were as follows: 37°C for 60 min, and 95°C for 5 min.
Subsequently, the cDNA was used for the quantification of miR-126
and VCAM-1 mRNA expression. The amplification conditions were 95°C
for 15 min at the initial activation, followed by 40 cycles of 94°C
for 15 sec, 55°C for 30 sec, and 70°C for 30 sec. β-actin primers
were used as an internal control in equal amounts to those of the
mRNA used. Gene expression was analyzed using QuantiTect SYBR-Green
PCR master mix miRNA or mRNA assays, according to the
manufacturer's instructions (Applied Biosystems, Foster City, CA,
USA). Dissociation curve analysis of each sample yields only one
peak, resulting from the specific amplification product
(primer-dimer not co-amplified). These values were normalized to
the reference gene β-actin as follows: fold =2−ΔΔCt,
where ΔCt represents the differences in cycle threshold numbers
between each of the target gene and β-actin, and ΔΔCt represents
the relative change between compared groups.
Western blot analysis
The VECs were lysed in ice-cold RIPA buffer (1%
NP-40, 0.1% sodium dodecyl sulfate, 0.5% sodium deoxycholate and 50
mM Tris, pH 7.4) containing protease inhibitor
phenylmethanesulfonyl fluoride. The total protein was quantified by
BCA assay, and the loaded concentration was 300–500 µg/ml.
The protein was separated by 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis. The protein was
transferred onto a polyacrylamide difluoride membrane, which was
blocked in phosphate-buffered saline-Tween-20 containing 5% non-fat
dry milk, and incubated with primary antibodies to VCAM-1, p-PI3K,
PI3K, p-Akt, Akt, p65, p-IκB, IκB or β-actin. Immunoreactive
protein was detected with secondary antibodies (Cat. nos. ZB2301
and ZB2305; Zhongshan, Beijing, China) conjugated to horseradish
peroxidase which enhanced chemiluminescence. To investigate the
relation between PI3K/Akt and NF-κB pathways in the effect of
paeonol, the VECs were pretreated with LY294002 (an inhibitor of
PI3K, 50 µM) and PDTC (an inhibitor of NF-κB, 50 µM)
for 1 h, and then treated with ox-LDL (20 mg/l) in the presence of
paeonol (120 µM). The protein expression levels were
detected.
Generation of luciferase reporter
construct
miR-126 consensus response element, UGAGUGCCAUGCU,
VCAM-1 wild-type 3′UTR response element, ACUGGACGGUACGG, and VCAM-1
mutant-type 3′UTR response element, and ACU GUUGCCAUGCG, were
inserted into the multiple cloning site of the miR-126 expression
vector, p-LUC-Report, downstream of cDNA for luciferase. Each
vector, along with miR-126 mimic, was transfected into the VECs
using HiPerFect reagent. The VECs were cultured for 24 h and
assayed using the Dual-Luciferase Reporter assay system. Relative
luciferase activity was calculated by normalizing the firefly
luminescence as to Renilla luminescence.
Isolation of rat monocytes
The rats were anaesthetized with chloral hydrate.
The abdominal aorta blood of the rats was obtained using the
heparin anticoagulant tube from the rats and centrifuged at 160 × g
for 10 min. The monocyte fraction was prepared by the
Ficoll-Hypaque gradient technique. The cells were then suspended
with lymphocyte separation solution (9% sodium diatrizoate, 5.7%
Ficoll PM100), and centrifuged at 670 × g for 15 min. Monocytes
were washed with phosphate buffer solution and DMEM, and
centrifuged at 240 × g for 6 min. The proportion of monocytes
>90% was confirmed by flow cytometry assay using anti-CD14 (Cat.
no. MAB1219) and anti-CD56 (Cat. no. MAB2120Z) antibodies
(Bioworld, Nanjing, China). Cell viability >95% was examined
using trypan blue staining assay. For the following experiments,
the monocyte concentration was adjusted to 1×106
cells/ml.
Cell adhesion analysis
The VECs were seeded in 96-well plates at a density
of 1×104 cells/well, and transfected with miR-126 mimic
or inhibitor. The VECs were pre-treated with paeonol for 24 h, and
incubated with fresh medium containing ox-LDL (20 mg/l) for 24 h.
The monocyte mixture was washed and co-cultured with the VECs for
60 min at 37°C and 5% CO2. Subsequently, 200 µl
of PBS were added to the dishes and gently flicked on tissue paper.
The unbound monocytes were removed. The adherent cells were fixed
in 100 µl methanol for 10 min, washed and incubated with 100
µl Rose-Bengal (0.25%, pH 7.3; Aladdin, Shanghai, China) for
10 min, as previously described (20). These cells were washed 3 times
with PBS and incubated with 50% ethanol for 1 h. The absorbance
(A) value was measured using a microplate reader at 490
nm.
Statistical analysis
All data are expressed as the means ± SD, and
analyzed using the SPSS version 17.0 software package. The
Student's t-test and ANOVA were used for the analysis of
experimental data. A value of P<0.05 was considered to indicate
a statistically significant difference.
Results
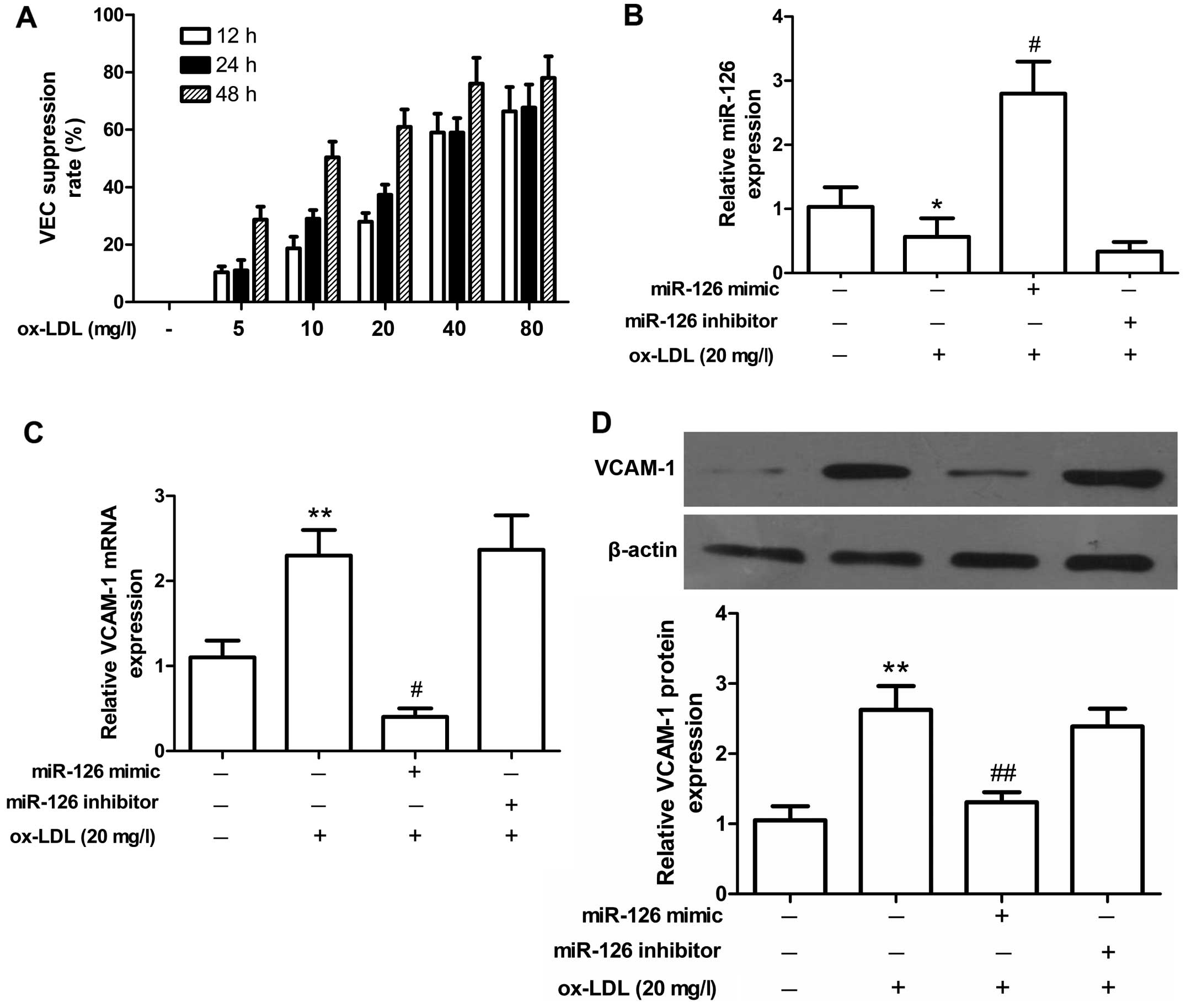
Effect of ox-LDL on miR-126 expression in
VECs
To verify whether miR-126 is specifically expressed
in inflammatory VECs, ox-LDL was used to stimulate the rat VECs
in vitro. We found that stimulation with ox-LDL (20, 40 and
80 mg/l) suppressed VEC viability in a concentration- and
time-dependent manner (as shown by the increased VEC suppression
rate; Fig. 2A). The suppression
rate of cell viability was close to 50% in the cells stimulated
with 20 mg/l ox-LDL for 24 h. These conditions were used as the
optimal stimulation concentration and time in subsequent
experiments. Of note, miR-126 had a lower expression in the
ox-LDL-stimulated VECs. Transfection with the miR-126 mimic
significantly promoted miR-126 expression, and transfection with
the miR-126 inhibitor decreased miR-126 expression (Fig. 2B). To investigate the function of
miR-126, the expression of VCAM-1, which is a target gene of
miR-126, was analyzed. miR-126 negatively regulated VCAM-1
expression at the mRNA and protein level (Fig. 2C and D). The expression of VCAM-1
increased with ox-LDL stimulation, and decreased by transfection
with the miR-126 mimic. Transfection with the miR-126 inhibitor led
to an increase in VCAM-1 expression. Taken together, our results
suggested that miR-126 had a lower expression in the
ox-LDL-stimulated VECs, and that miR-126 negatively regulated
VCAM-1 expression.
Identification of VCAM-1 serving as a
target gene of miR-126
To determine the predicted target sites for miR-126
in the 3′UTR of VCAM-1 mRNA which is responsible for the
down-regulation of VCAM-1 expression, dual-luciferase reporter
assay with a vector containing the VCAM-1 wild-type or mutant-type
3′UTR downstream of the luciferase reporter gene was performed. The
wild-type or mutant-type 3′UTR mRNA of VCAM-1 and miR-126 mimic
were transfected into the VECs using HiPerFect reagent. Following
24 h of culture, firefly luciferase activity was detected in the
cells transfected with the reporter plasmids and normalized to the
transfection efficiency with Renilla luciferase activity,
which was derived from a co-transfection with an expression
plasmid. Transfection with the wild-type plasmids and the presence
of miR-126 target sites led to a marked reduction in luciferase
activity. By contrast, the incorporation of mutant-type 3′UTR mRNA
plasmids did not affect luciferase efficiency, although the
luciferase activity was increased compared to the group transfected
with the wild-type plasmid (Fig.
3). It was clearly demonstrated that VCAM-1 served as a target
gene of miR-126 in the VECs.
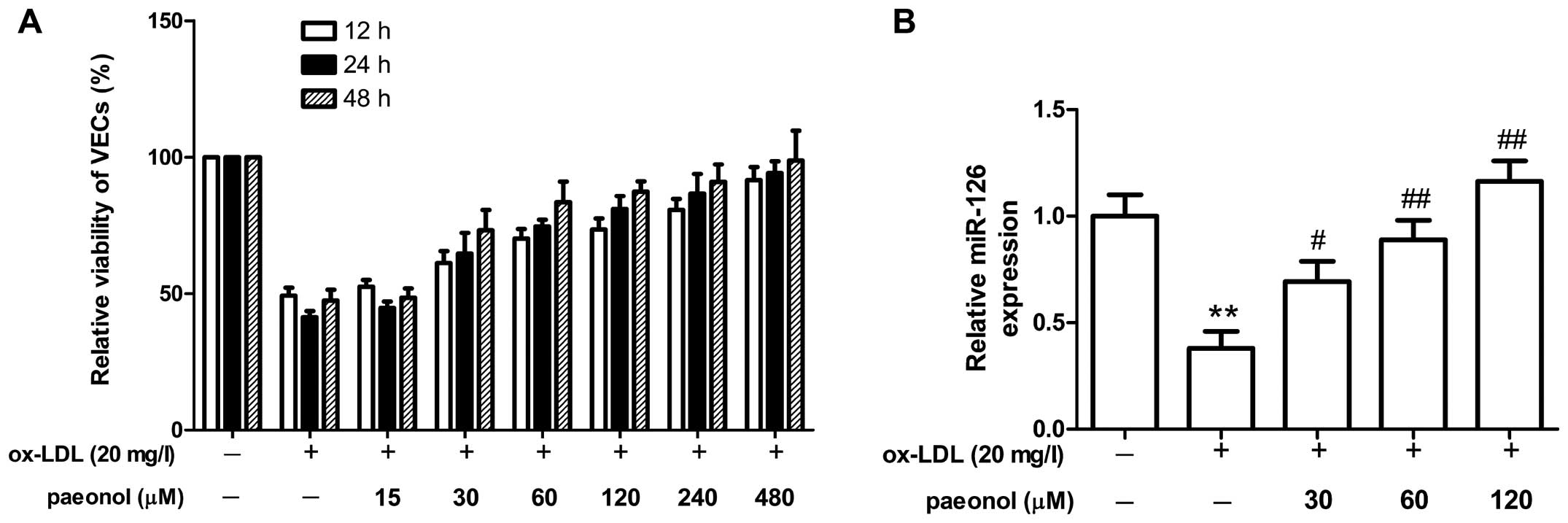
Effect of paeonol on miR-126 expression
in ox-LDL-injured VECs
To further determine the mechanisms through which
paeonol can effectively prevent and treat AS, we examined the
effect of paeonol on miR-126 expression in the ox-LDL-injured VECs.
The VECs were first treated with paeonol, and then stimulated with
ox-LDL (20 mg/l). We found that treatment with paeonol (30, 60,
120, 240 and 480 µM) protected the VECs from ox-LDL-induced
injury, as shown by the increase in cell viability (Fig. 4A). However, paeonol had a slightly
toxic effect as a concentration >480 µM and cultured
times longer than 48 h (data not shown). Notably, paeonol
significantly promoted miR-126 expression in a
concentration-dependent manner (Fig.
4B). Our resutls thus suggested that paeonol promoted miR-126
expression to inhibit VEC injury.
Paeonol exerts inhibitory effects on
VCAM-1 expression by promoting miR-126 expression
In order to demonstrate the effect of paeonol on the
function of miR-126, we analyzed the expression of its target gene,
VCAM-1. Firstly, the mimic/inhibitor of miR-126 was transfected
into rat VECs; after 24 h, the VECs were cultured with medium
containing paeonol (120 µM). We found that the miR-126
expression level was markedly upregulated by paeonol, increased by
the miR-126 mimic, and decreased by the miR-126 inhibitor (Fig. 5A). Paeonol markedly suppressed the
mRNA expression of VCAM-1, and the overexpression of miR-126
induced by transfection with the miR-126 mimic markedly inhibited
the mRNA expression of VCAM-1; this effect of paeonol was
attenuated by transfection with the miR-126 inhibitor (Fig. 5B). We then sought to determine the
regulatory effect of miR-126 mediated by paeonol on VCAM-1
expression at the protein level. The upregulation of miR-126
expression by transfection with miR-126 mimic markedly enhanced the
inhibitory effect of paeonol on VCAM-1 protein expression. By
contrast, the knockdown of miR-126 markedly diminished the
inhibitory effects of paeonol on VCAM-1 expression (Fig. 5C). These results indicated that
paeonol inhibited VCAM-1 expression by promoting miR-126
expression.
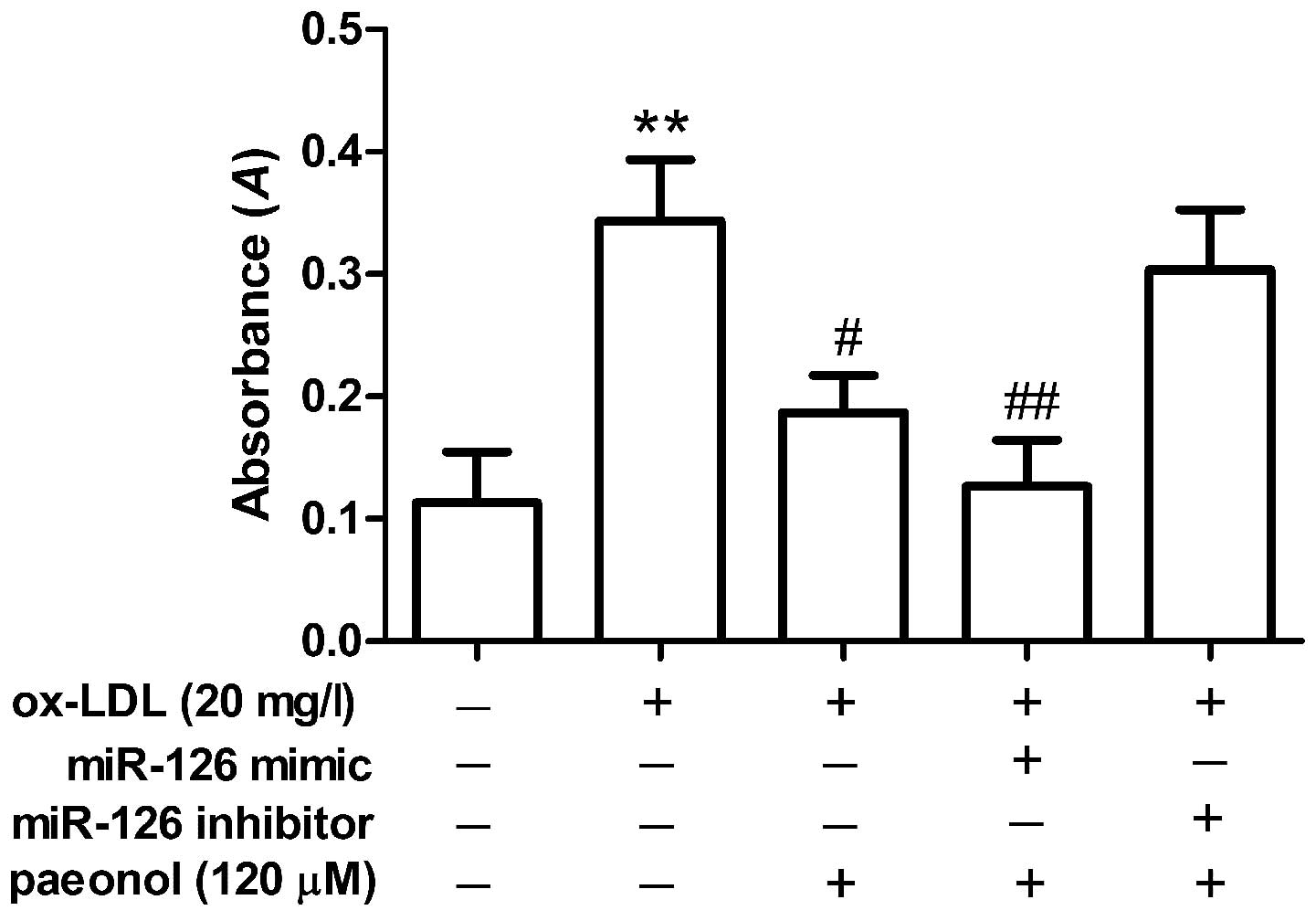
Effects of paeonol on monocyte adhesion
to ox-LDL-injured VECs are mediated via miR-126 expression
To investigate whether paeonol inhibits monocyte
adhesion to ox-LDL-injured VECs by regulating miR-126 expression,
the mimic/inhibitor of miR-126 was transfected into the VECs; after
24 h, the VECs were stimulated with ox-LDL in the presence or
absence of paeonol. ox-LDL induced monocyte adhesion to VECs. It
was shown that paeonol suppressed monocyte adhesion to VECs induced
by ox-LDL, and the effect of paeonol was enhanced by the miR-126
mimic, but was reduced by the miR-126 inhibitor (Fig. 6). These results suggested that
paeonol suppressed monocyte adhesion to ox-LDL-injured VECs via
miR-126 expression.
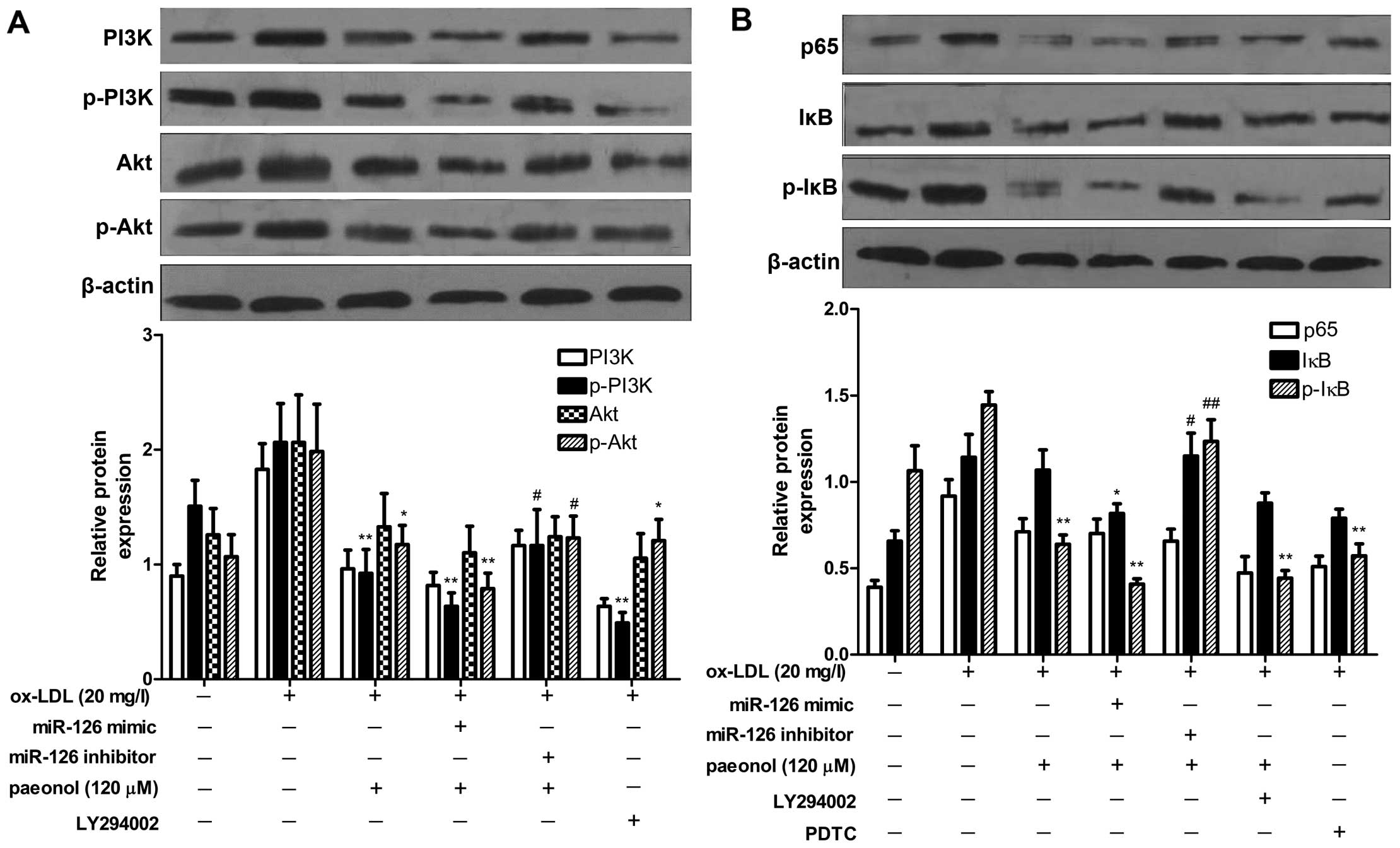
Effects of paeonol on the PI3K/Akt/NF-κB
pathway are mediated via miR-126 expression
To explore whether the promoting effects of paeonol
on miR-126 expression are closely related to the PI3K/Akt/NF-κB
pathway, miR-126 mimic/inhibitor was transfected into the VECs.
After 24 h, the VECs were treated with LY294002 (an inhibitor of
PI3K) and PDTC (an inhibitor of NF-κB) for 1 h, and then stimulated
with ox-LDL (20 mg/l) in the presence of paeonol (120 µM).
Paeonol significantly suppressed the phosphorylation of PI3K and
Akt. The effect of paeonol was enhanced by transfection with the
miR-126 mimic, but was diminished by transfection with the miR-126
inhibitor (Fig. 7A). Moreover, it
was found that paeonol significantly suppressed the phosphorylation
of IκB and the activation of the NF-κB pathway, and its effect was
enhanced by the miR-126 mimic, but diminished by the miR-126
inhibitor. In addition, the NF-κB pathway was indicated as
downstream of the PI3K/Akt pathway by the inhibitor of the PI3K
pathway (Fig. 7B). Taken
together, our results revealed that paeonol blocked the activation
of the PI3K/Akt/NF-κB signaling pathway via the upregulation of
miR-126 expression.
Discussion
In this study, ox-LDL markedly induced damage to
VECs, and miR-126 had a lower expression in ox-LDL-injured VECs
in vitro. It has been reported that miR-126 is a strongly
expressed miRNA specific to endothelial cells, and also affects the
physiological and pathological processes of vascular diseases, such
as AS (11,21). AS is a complex chronic
inflammatory and metabolic disease (22). Injured VECs may be detrimental to
the structure of plaque, and based on the fact that endothelial
destruction precedes atherogenesis, the injury of VECs may also
predispose to arterial thrombosis (23). VCAM-1 plays an important role in
the slow rolling of monocytes along the surface of the endothelial
layer and in the pathological process of AS (9,24).
miRNAs play an important role in both inflammatory and
cardiovascular diseases (25). In
this study, miR-126 downregulated VCAM-1 expression and protected
the rat VECs from ox-LDL-induced injury.
miRNAs can bind and interfere with the mRNAs of
target genes specifically, and negatively regulate their
expression. Accordingly, in this study, miR-126 suppressed VCAM-1
expression by combination of the miR-126-binding sites, and the
upregulation of miR-126 inhibited VCAM-1 gene expression. Exogenous
miR-126 suppresses luciferase activity when the miR-126-binding
site is fused to the luciferase 3′UTR mRNA of VCAM-1 (11). These findings raise the
possibility that medical strategies aimed at causing controlled
alterations to the levels of gene-targeting miRNAs, such as
miR-126, may be effective therapies for inflammatory diseases.
miRNAs are good candidates, particularly for the therapeutic
intervention, as they have an impact on the expression of target
genes, rather than simply turning genes on or off, thereby
regulating cellular inflammation through external regulation.
Moreover, we found that paeonol promoted miR-126
expression to suppress VCAM-1 expression in ox-LDL-injured VECs and
monocyte adhesion to the VECs. miR-126 expression can be modulated
by natural compound, and participates in the vascular inflammatory
response (26). The deletion of
miR-126 causes the loss of vascular integrity and leads to defects
in the proliferation and migration of endothelial cells (27). VCAM-1 is expressed in VECs and
plays an important role in monocyte or lymphocyte rolling and
adhesion in inflammation in early-onset AS (28,29). Thus, the inhibition of VCAM-1
expression suggests anti-inflammatory properties for endothelial
cells. As reported in a previous study, paeonol suppressed VCAM-1
expression in TNF-α-stimulated VECs (3). VCAM-1 expression can be regulated at
the post-transcriptional level (30). miR-126 inhibits leukocyte
adherence through the decrease of VCAM-1 expression (11). In is suggested that miR-126
expression induced by paeonol can downregulate the expression of
adhesion molecule and may exert additional inhibitory effects on
vascular inflammation.
However, it has been demonstrated that the
transcription factor, NF-κB, is sensitve to oxidants such as
ox-LDL, and is also associated with the cellular function of
adhesion (31,32). NF-κB activation is required for
the induction of adhesion molecules, such as VCAM-1, and is
responsible for monocyte adhesion and vascular inflammation
(33). On the other hand, the
PI3K/Akt signaling pathway is known as a critical regulator of
NF-κB activation in response to extracellular stress (34). In this study, to further clarify
the detailed mechanisms responsible for the inhibitory effects of
paeonol on cell adhesion, the effects of paeonol on the
PI3K/Akt/NF-κB pathway were analyzed. Our results revealed that
paeonol blocked the activation of the PI3K/Akt/NF-κB signaling
pathway by promoting miR-126 expression. The overexpression of
miR-126 has been reported to inhibit the expression of p-Akt and to
regulate PI3K signaling by targeting the PI3K regulatory subunit β
(p85β) (35,36) a target gene of miR-126 identified
previously in endothelial cells (37), indicating that its function is
related to the PI3K/Akt pathway. Similarly, the decrease in p-Akt
protein levels follwoing the suppression of p85β protein expression
by miR-126 has also reported (38). In this study, it was proven that
NF-κB is downstream of the PI3K/Akt pathway as regards the effect
of paeonol. It played an important role in the inhibitory effect of
paeonol on monocyte adhesion to ox-LDL-injured VECs via the
induction of miR-126 expression.
In conclusion, in the present study, we demonstrated
that paeonol promoted miR-126 expression to inhibit monocyte
adhesion to ox-LDL-injured VECs and block the activation of the
PI3K/Akt/NF-κB signaling pathway. We provide evidence that miR-126
may serve as an important therapeutic target in the treatment of AS
with the use of paeonol.
Acknowledgments
This study was supported by research grants from the
National Natural Science Foundation of China (grant. nos. 81473386
and 81274134).
Abbreviations:
|
Akt
|
protein kinase B
|
|
ox-LDL
|
oxidized low-density lipoprotein
|
|
FBS
|
fetal bovine serum
|
|
AS
|
atherosclerosis
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
DMSO
|
dimethyl-sulfoxide
|
|
IκBα
|
inhibitory factor κB-α
|
|
miRNAs or miRs
|
micro-RNAs
|
|
IL
|
interleukin
|
|
MTT
|
3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium
bromide
|
|
NF-κB
|
nuclear factor-κB
|
|
PDTC
|
pyrrolidinedithiocarbamate
|
|
PI3K
|
phosphatidylinositol 3-kinase
|
|
TNF-α
|
tumor necrosis factor-α
|
|
VSMCs
|
vascular smooth muscle cells
|
|
VCAM-1
|
vascular cell adhesion molecule-1
|
|
VECs
|
vascular endothelial cells
|
References
|
1
|
Chou TC: Anti-inflammatory and analgesic
effects of paeonol in carrageenan-evoked thermal hyperalgesia. Br J
Pharmacol. 139:1146–1152. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nizamutdinova IT, Jin YC, Kim JS, Yean MH,
Kang SS, Kim YS, Lee JH, Seo HG, Kim HJ and Chang KC: Paeonol and
paeoni-florin, the main active principles of Paeonia albiflora,
protect the heart from myocardial ischemia/reperfusion injury in
rats. Planta Med. 74:14–18. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pan LL and Dai M: Paeonol from Paeonia
suffruticosa prevents TNF-alpha-induced monocytic cell adhesion to
rat aortic endothelial cells by suppression of VCAM-1 expression.
Phytomedicine. 16:1027–1032. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hsieh CL, Cheng CY, Tsai TH, Lin IH, Liu
CH, Chiang SY, Lin JG, Lao CJ and Tang NY: Paeonol reduced cerebral
infarction involving the superoxide anion and microglia activation
in ischemia-reperfusion injured rats. J Ethnopharmacol.
106:208–215. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Su SY, Cheng CY, Tsai TH, Hsiang CY, Ho TY
and Hsieh CL: Paeonol attenuates H2O2-induced
NF-κB-associated amyloid precursor protein expression. Am J Chin
Med. 38:1171–1192. 2010. View Article : Google Scholar
|
|
6
|
Vitaglione P, Morisco F, Caporaso N and
Fogliano V: Dietary antioxidant compounds and liver health. Crit
Rev Food Sci Nutr. 44:575–586. 2004. View Article : Google Scholar
|
|
7
|
Li H, Dai M and Jia W: Paeonol attenuates
high-fat-diet-induced atherosclerosis in rabbits by
anti-inflammatory activity. Planta Med. 75:7–11. 2009. View Article : Google Scholar
|
|
8
|
Li W, Ishihara K, Yokota T, Nakagawa T,
Koyama N, Jin J, Mizuno-Horikawa Y, Wang X, Miyoshi E, Taniguchi N
and Kondo A: Reduced alpha4beta1 integrin/VCAM-1 interactions lead
to impaired pre-B cell repopulation in alpha 1,6-fucosyltransferase
deficient mice. Glycobiology. 18:114–124. 2008. View Article : Google Scholar
|
|
9
|
Hwa JS, Mun L, Kim HJ, Seo HG, Lee JH,
Kwak JH, Lee DU and Chang KC: Genipin selectively inhibits
TNF-α-activated VCAM-1 but not ICAM-1 expression by upregulation of
PPAR-γ in endothelial cells. Korean J Physiol Pharmacol.
15:157–162. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tsoyi K, Kim WS, Kim YM, Kim HJ, Seo HG,
Lee JH, Yun-Choi HS and Chang KC: Upregulation of PTEN by CKD712, a
synthetic tetrahydroisoquinoline alkaloid, selectively inhibits
lipopolysaccharide-induced VCAM-1 but not ICAM-1 expression in
human endothelial cells. Atherosclerosis. 207:412–419. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Harris TA, Yamakuchi M, Ferlito M, Mendell
JT and Lowenstein CJ: MicroRNA-126 regulates endothelial expression
of vascular cell adhesion molecule 1. Proc Natl Acad Sci USA.
105:1516–1521. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sturgeon CM, Chicha L, Ditadi A, Zhou Q,
McGrath KE, Palis J, Hammond SM, Wang S, Olson EN and Keller G:
Primitive erythropoiesis is regulated by miR-126 via
nonhematopoietic VCAM-1+ cells. Dev Cell. 23:45–57.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Harris TA, Yamakuchi M, Kondo M, Oettgen P
and Lowenstein CJ: Ets-1 and Ets-2 regulate the expression of
microRNA-126 in endothelial cells. Arterioscler Thromb Vasc Biol.
30:1990–1997. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nikolic I, Plate KH and Schmidt MH: EGFL7
meets miRNA-126: An angiogenesis alliance. J Angiogenes Res.
2:92010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bai Y, Bai X, Wang Z, Zhang X, Ruan C and
Miao J: MicroRNA-126 inhibits ischemia-induced retinal
neovascularization via regulating angiogenic growth factors. Exp
Mol Pathol. 91:471–477. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sui L, Wang J and Li BM: Role of the
phosphoinositide 3-kinase-Akt-mammalian target of the rapamycin
signaling pathway in long-term potentiation and trace fear
conditioning memory in rat medial prefrontal cortex. Learn Mem.
15:762–776. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tsubosaka Y, Murata T, Yamada K, Uemura D,
Hori M and Ozaki H: Halichlorine reduces monocyte adhesion to
endothelium through the suppression of nuclear factor-kappaB
activation. J Pharmacol Sci. 113:208–213. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ghosh S, Banerjee S and Sil PC: The
beneficial role of curcumin on inflammation, diabetes and
neurodegenerative disease: A recent update. Food Chem Toxicol.
83:111–124. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Salvayre R, Auge N, Benoist H and
Negre-Salvayre A: Oxidized low-density lipoprotein-induced
apoptosis. Biochim Biophys Acta. 1585:213–221. 2002. View Article : Google Scholar
|
|
20
|
Basoni C, Nobles M, Grimshaw A, Desgranges
C, Davies D, Perretti M, Kramer IM and Genot E: Inhibitory control
of TGF-beta1 on the activation of Rap1, CD11b, and transendothelial
migration of leukocytes. FASEB J. 19:822–824. 2005.PubMed/NCBI
|
|
21
|
Asgeirsdóttir SA, van Solingen C, Kurniati
NF, Zwiers PJ, Heeringa P, van Meurs M, Satchell SC, Saleem MA,
Mathieson PW, Banas B, et al: MicroRNA-126 contributes to renal
microvascular heterogeneity of VCAM-1 protein expression in acute
inflammation. Am J Physiol Renal Physiol. 302:F1630–F1639. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rioufol G, Finet G, Ginon I, André-Fouët
X, Rossi R, Vialle E, Desjoyaux E, Convert G, Huret JF and Tabib A:
Multiple atherosclerotic plaque rupture in acute coronary syndrome:
A three-vessel intravascular ultrasound study. Circulation.
106:804–808. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Peng N, Meng N, Wang S, Zhao F, Zhao J, Su
L, Zhang S, Zhang Y, Zhao B and Miao J: An activator of mTOR
inhibits oxLDL-induced autophagy and apoptosis in vascular
endothelial cells and restricts atherosclerosis in apolipoprotein
E−/− mice. Sci Rep. 4:55192014.
|
|
24
|
Fuentes E, Palomo I and Alarcón M:
Platelet miRNAs and cardiovascular diseases. Life Sci. 133:29–44.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Condorelli G, Latronico MV and Cavarretta
E: microRNAs in cardiovascular diseases: Current knowledge and the
road ahead. J Am Coll Cardiol. 63:2177–2187. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Angel-Morales G, Noratto G and
Mertens-Talcott SU: Standardized curcuminoid extract (Curcuma longa
L.) decreases gene expression related to inflammation and interacts
with associated microRNAs in human umbilical vein endothelial cells
(HUVEC). Food Funct. 3:1286–1293. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang S, Aurora AB, Johnson BA, Qi X,
McAnally J, Hill JA, Richardson JA, Bassel-Duby R and Olson EN: The
endothelial-specific microRNA miR-126 governs vascular integrity
and angiogenesis. Dev Cell. 15:261–271. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cybulsky MI, Iiyama K, Li H, Zhu S, Chen
M, Iiyama M, Davis V, Gutierrez-Ramos JC, Connelly PW and Milstone
DS: A major role for VCAM-1, but not ICAM-1, in early
atherosclerosis. J Clin Invest. 107:1255–1262. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Libby P: Inflammation in atherosclerosis.
Nature. 420:868–874. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Croft D, McIntyre P, Wibulswas A and
Kramer I: Sustained elevated levels of VCAM-1 in cultured
fibroblast-like synoviocytes can be achieved by TNF-alpha in
combination with either IL-4 or IL-13 through increased mRNA
stability. Am J Pathol. 154:1149–1158. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hayden MS and Ghosh S: Signaling to
NF-kappaB. Genes Dev. 18:2195–2224. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Albelda SM, Smith CW and Ward PA: Adhesion
molecules and inflammatory injury. FASEB J. 8:504–512.
1994.PubMed/NCBI
|
|
33
|
Lockyer JM, Colladay JS, Alperin-Lea WL,
Hammond T and Buda AJ: Inhibition of nuclear factor-kappaB-mediated
adhesion molecule expression in human endothelial cells. Circ Res.
82:314–320. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yang M, Huang J, Pan HZ and Jin J:
Triptolide overcomes dexamethasone resistance and enhanced
PS-341-induced apoptosis via PI3k/Akt/NF-κB pathways in human
multiple myeloma cells. Int J Mol Med. 22:489–496. 2008.PubMed/NCBI
|
|
35
|
Wang XC, Du LQ, Tian LL, Wu HL, Jiang XY,
Zhang H, Li DG, Wang YY, Wu HY, She Y, et al: Expression and
function of miRNA in postoperative radiotherapy sensitive and
resistant patients of non-small cell lung cancer. Lung Cancer.
72:92–99. 2011. View Article : Google Scholar
|
|
36
|
Guo C, Sah JF, Beard L, Willson JK,
Markowitz SD and Guda K: The noncoding RNA, miR-126, suppresses the
growth of neoplastic cells by targeting phosphatidylinositol
3-kinase signaling and is frequently lost in colon cancers. Genes
Chromosomes Cancer. 47:939–946. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fish JE, Santoro MM, Morton SU, Yu S, Yeh
RF, Wythe JD, Ivey KN, Bruneau BG, Stainier DY and Srivastava D:
miR-126 regulates angiogenic signaling and vascular integrity. Dev
Cell. 15:272–284. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sessa R, Seano G, di Blasio L, Gagliardi
PA, Isella C, Medico E, Cotelli F, Bussolino F and Primo L: The
miR-126 regulates angiopoietin-1 signaling and vessel maturation by
targeting p85β. Biochim Biophys Acta. 1823:1925–1935. 2012.
View Article : Google Scholar : PubMed/NCBI
|