Introduction
As the economy rapidly grows in the developing
world, the incidence of depression is poised to increase. WHO
statistics have indicated that depression will be the leading cause
of disease burden worldwide by the year 2030 (1). Even with the armamentarium of
pharmacotherapy and psychotherapy, caring for patients with
depression remains a challenge due to the lack of universal
responses to anti-depressive therapies and frequent recurrences of
depressive episodes (2,3).
The currently used anti-depressants mostly target
brain neurotransmitters, including 5-hydroxytryptamine (5-HT),
norepinephrine [NE also known as noradrenaline (NA)] and dopamine
(DA). Following the consumption of antidepressants, it usually
takes several weeks for patients to exhibit any improvement
(4). In addition, in a great
number of patients, antidepressants are unable to achieve favorable
effects. Thus, fully elucidating the mechanisms responsible for the
pathogenesis of depression is of great significance and may lead to
the development of novel therapeutic methods.
Depression is thought to develop as a result of
genetic and environmental changes, and biological factors (5). Although the specific molecular and
cellular mechanisms are unclear as regards these factors, an
association between chronic stress and glucocorticoid levels has
attracted attention (6). Clinical
studies have indicated that patients with psychiatric disorders,
such as major depression, develop symptoms of chronic stress and
mount attenuated immune responses (7). Kiank et al (8) found that glucocorticoid levels
increase significantly and induce a negative immune response in
mice and humans.
It has been reported that chronic stress induces
structural alterations in splenic lymphoid tissue, which are
relevant to the changes in corticosterone levels in rats (9). However, there is no direct evidence
of alterations in splenic function and gene expression associated
with depression. Thus, in this study, we aimed to examine the
histopathological characteristics of spleen tissue, functional
changes and differentially expressed genes in mice with
depressive-like behavior. We also examined the levels of certain
related proteins in the main pathways assessed in the brain and the
spleen separately.
Materials and methods
Animals
Adult Balb/c mice (6–7 weeks old, weighing 18–22 g)
were purchased from the Experimental Animal Centre of Hualan
Bioengineering Co., Xinxiang, China (quality certificate no.
0005496). The mice were bred under aseptic conditions, and allowed
to acclimatize for 1 week in an environment with a controlled
temperature (25–27°C), constant humidity (50–60%) and 12 h
light/dark cycle, and were given free access to food and water. The
animal protocols for this study were approved by the Xinxiang
Medical University Institutional Animal Care and Use Committee
according to the Guidelines on the Humane Treatment of Laboratory
Animals published by the Ministry of Science and Technology of the
People's Republic of China in 2006.
Animal grouping and establishment of
mouse model of depression
Female mice were used for all the experiments
according to the literature (10,11). A total of 72 mice were randomly
divided into 2 groups: 22 mice served as the normal control group,
and another 50 mice were used to establish the model of depression
by an intraperitoneal injection of corticosterone [Sigma-Aldrich,
Munich, Germany; purity >98.5% determined by high performance
liquid chromatography (HPLC) assay (data not shown)] suspension at
20 mg/kg as previously described by Ago et al (12). The mice in the normal control
group were administered normal saline at the same volume. All mice
were injected once a day for 21 days. The success rate of the
depression model was approximately 74%. After excluding all the
demised animals (1 mouse died in the contorl group due to unknown
reasons) and those that failed to exhibit depressionlike behaviors,
21 mice remained in the normal control group, and 37 mice in the
depression model group. A total of 9 animals in each group were
used for the microarray analysis of spleen and the measurement of
neurotransmitters in the brain, and 4 animals in each group were
used for the histopathological examination. The remaining animals
were divided into 3 groups as follows: the normal control group
(n=8), the depression model group (n=12) and the fluoxetine group
(n=12), of which 8 mice in each group were used for western blot
analysis. The mice in the fluoxetine group were administered
fluoxetine by gavage for 15 days, and the mice of normal control
group and depression model group were given 0.9% saline solution at
the same time.
Open field test
The mice were individually housed in a cage
measuring 55×55×40 cm3 without a cover. The bottom of
the cage was divided into 11×11 cm2 squares and marked
with white lines. The entire interior walls and the bottom floor
were painted with black color. A canon XF300 video camera (Canon
China Co., Beijing, China) was installed on the ceiling at the
center of the room. The movements of the mice were recorded using
the video camera. The locomotive activities based on the distance
of movement and the numbers of standing-up were measured on the
21st day after the beginning of the experiment. Specifically, the
mouse was placed at the bottom center of the cage and allowed to
run freely for the first 5 min in order to allow the animal to
adapt to the new environment. Subsequently, the activities of the
mouse in the next 5 min were recorded using the video camera. These
measurements were performed at around 10 a.m. after the final
administration of normal saline (normal control group) or
corticosterone (depression model group). The activities of the
mouse were analyzed using points allocated to all the movements.
Walking across one small square or 10 cm in distance was allocated
1 point, as previously described (13,14). The lifting of both front limbs or
touching the walls of the cage was also allocated 1 point. Mobile
scores were calculated based on all the points recorded.
Sucrose preference test (SPT)
The sucrose preference test was performed as
previously described by Bhatt et al with slight
modifications (15). The mice
were trained before the experiment. Each mouse was housed in a
single cage with two bottles of sterile sucrose water, each bottle
containing 100 ml solution. Within the next 24 h, one bottle of
sterile sucrose water was replaced with a bottle of sterile
distilled water, and the position of the left bottle and right
bottle was switched. Sucrose and water consumption within the last
1 h of a 24-h period was measured, which was regarded as the
baseline values. The sucrose preference rate was calculated using
the following formula: sucrose consumption/sucrose consumption +
water consumption. On the 21st day of the injection, sucrose and
water consumption were measured within 1 h, and the sucrose
preference rate was then calculated.
Histopathological examination
The brains and spleen tissue of the mice from both
groups were examined using H&E staining as previously described
(16). Four animals from each
group were sacrificed by decapitation, and the brains and spleens
were removed. All specimens were fixed for 24 h by 4%
paraformaldehyde phosphate buffer solution (pH 7.4). After the
specimens were regularly dehydrated and embedded in paraffin, a 5
mm portion of the forebrain and the spleen tissue was cut into
coronal sections. The paraffin-embedded sections
(5-µm-thick) were placed on glass slides and stained with
hematoxylin for 5–10 min and eosin for 2–3 min after routine
dewaxing. The image-analysis software of HMIAS-2000 system (Tongji
Medical College, Huazhong University of Science and Technology,
Wuhan, China) was used for the histological examination of the
specimens.
Measurement of neurotransmitters
The NA, adrenaline (Ad), DA, tryptophan (Try) and
5-HT levels in the mouse brains were determined using the HPLC
fluorescence method (Varian Prostar 210; Varian, Palo Alto, CA,
USA) as previously described (17). Nine mice from each group (control
group and model group) were sacrificed by decapitation, and the
brains were removed. The prefrontal cortex was weighed and
dissected over ice. The brain area sample was placed in a
homogenizer, and 0.1 M perchlorate solution that contains 0.004 M
sodium metabisulfite was added. The brain tissue to the perchlorate
solution ratio was 1:3. The homogenate of each sample were
centrifuged for 30 min (8,183 x g) at −4°C, the supernatants were
filtered by 0.45 µm hydrophilic membranes and transferred to
a centrifuge tube and stored at 4°C. The reagents of NA, Ad, DA,
Try and 5-HT (Sigma-Aldrich Co.) were used for the standard
solution. All the other chemicals used in this study were of HPLC
grade. Measurements were made by injecting 20 µl of each
sample into the Varian Prostar 210 series HPLC system with a
fluorescence detector (excitation wavelength, 280 nm; emission
wavelength, 360 nm), which employed a Varian C18 column (4. ×250
mm, 5 µm; velocity, 1 ml/min; column temperature, 20°C). The
range of the concentration of NA, Ad, Try or 5-HT in the standard
liquid was 0.0625–1.0 µg/ml and that of DA was 0.125–2.0
µg/ml. The mobile phase was composed of 30% methanol
(Concord Technology Co., Tianjin, China), 4% ultrapure water and
66% phosphate buffer, which consisted of 0.02 M
KaH2PO4 and 0.005 M heptyl sulfonic acid
sodium (both from Daily Chemical Co., Shanghai, China) (pH .3). The
flow rate was maintained at 1.0 ml/min.
Evaluation of splenic function
Retro-orbital blood collection was conducted before
sacrificing the animals by decapitation. A drop of blood was placed
on the slide and spread into a thin layer. Cellular morphology was
observed by Wright staining, as previously described (18). The red blood cells, Howell-Jolly
bodies (HJB) and various types of white blood cells (WBC) were
observed using a light microscope (Olympus 009083; Olympus, Tokyo,
Japan) and the numbers of the cells were counted with ×100 oil
lens. Five fields were randomly selected from the upper left, lower
left, upper right, lower right and the middle areas of the blood
smear and photographed using a HMIAS-2000 high-resolution color
medical image analysis system (Qingping Imaging Co., Tongji Medical
College, Shanghai, China). HJB and hemosiderosis were observed and
analyzed, as previously described (19).
Splenic gene determination and
analysis
The spleens of 9 mice from each group were removed
immediately after sacrifice. After the spleen tissue was processed
by nitrogen and TRIzol reagent (Life Technologies, Carlsbad, CA,
USA), the samples were stored in a −80°C freezer until analysis.
Following the manufacturer's instructions, total RNA from the
spleen of each mouse was extracted using TRIzol reagent. The equal
amounts of RNA from 3 mice in the same group were mixed to yield a
testing sample. Totally, 3 mixed testing samples from 9 mice were
used for the examination of gene chip. Total RNA was amplified and
labeled using the Low Input Quick Amp Labeling kit, One-Color
(Agilent Technologies, Santa Clara, CA, USA). The labeled cRNA were
purified using the RNeasy mini kit (Qiagen GmBH, Hilden,
Germany).
The samples were hybridized to the Agilent Whole
Mouse Genome Oligo Microarray (4×44K, including 41,174 genes and
transcript) by Gene Expression Hybridization kit (both from Agilent
Technologies), stained, washed and scanned according to the
standard Agilent oligonucleotide microarray hybridization protocol.
The computer data files to be used in data analysis were generated
with the GeneSpring software 11.0 (Agilent Technologies) using the
statistical algorithm provided. All chip samples were scanned and
data were read using the same instrument and following the same
protocol. The above process was performed by Shanghai Biotechnology
Corp. (Shanghai, China). Data quality assessment was then performed
following the guidelines outlined in the Agilent Data Analysis
Fundamentals Manual. All quality control results met the criteria
recommended by Agilent Technologies.
Western blot analysis
The spleens and prefrontal cortices of 8 mice from
each group were removed after sacrifice by cervical dislocation,
and all the tissues were kept at −40°C. The protein concentration
of each sample was determined using a bicinchoninic acid assay kit
using BSA as standard (Pierce, Rockford, IL, USA). Equal amounts of
proteins from each sample (40 µg) were separated by dodecyl
sulfate, sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE) and transferred onto polyvinylidene fluoride (PVDF)
membranes (Merck Millipore, Billerica, MA, USA). The membranes were
blocked with 5% non-fat milk in 0.01 M phosphate-buffered saline
(PBS) at room temperature for 1 h and then incubated overnight at
4°C with the primary antibodies (Abcam, Cambridge, MA, USA),
including anti-D site albumin promoter binding protein (DBP;
1:1,000), anti-γ-aminobutyric acid B receptor, 2 (GABBR2; 1:1,000)
and anti-substance P (SP; 1:200) antibodies. The secondary antibody
(IRDye 800-conjugated goat anti-rabbit secondary antibodies) was
added at 1:2,000 for 1 h at room temperature. Mouse anti-β-actin
antibody (1:2,000 dilution; Santa Cruz Biotechnology, Inc., Santa
Cruz, CA, USA) was used as internal reference to confirm that the
same amount of each type of proteins was loaded. The bands were
photographed by a fully automatic chemo-luminescence image analysis
system (Tanon-450; Tanon Science and Technology Co., Ltd..
Shanghai, China). The gray-scale of each band was detected using
ImageJ image analysis software. The quantities of each western blot
product were normalized by dividing the average gray level of the
signal by that of the corresponding β-actin, and observed as the
semi-quantitative value of the target fragments.
Statistical analysis
The data analysis of the behavioral tests (3
variables), monoamine depletion (5 variables) and blood tests (6
variables) in the mice were performed using an independent-samples
t-test. One-way analysis of variance (ANOVA) with post hoc tests,
followed by the least significant difference (LSD) test was
performed to compare the protein levels of DBP, GABBR2 and SP. The
results of quantitative analysis were summarized as the means ± SEM
and analyzed using SPSS 17.0 statistical software. A value of
P<0.05 was set to define statistical significance. All P-values
are two-tailed. Gene chip data were analyzed using the SAS system
(SBC analysis system).
Results
Changes in animal behavior
The open field test was used to explore the
locomotor activity of the mice, as previously described (20). As a test for the degree of
anhedonia, sucrose preference rate was determined based on the
sucrose consumption. The results revealed that there was no
distinctive behavioral difference between the 2 groups before the
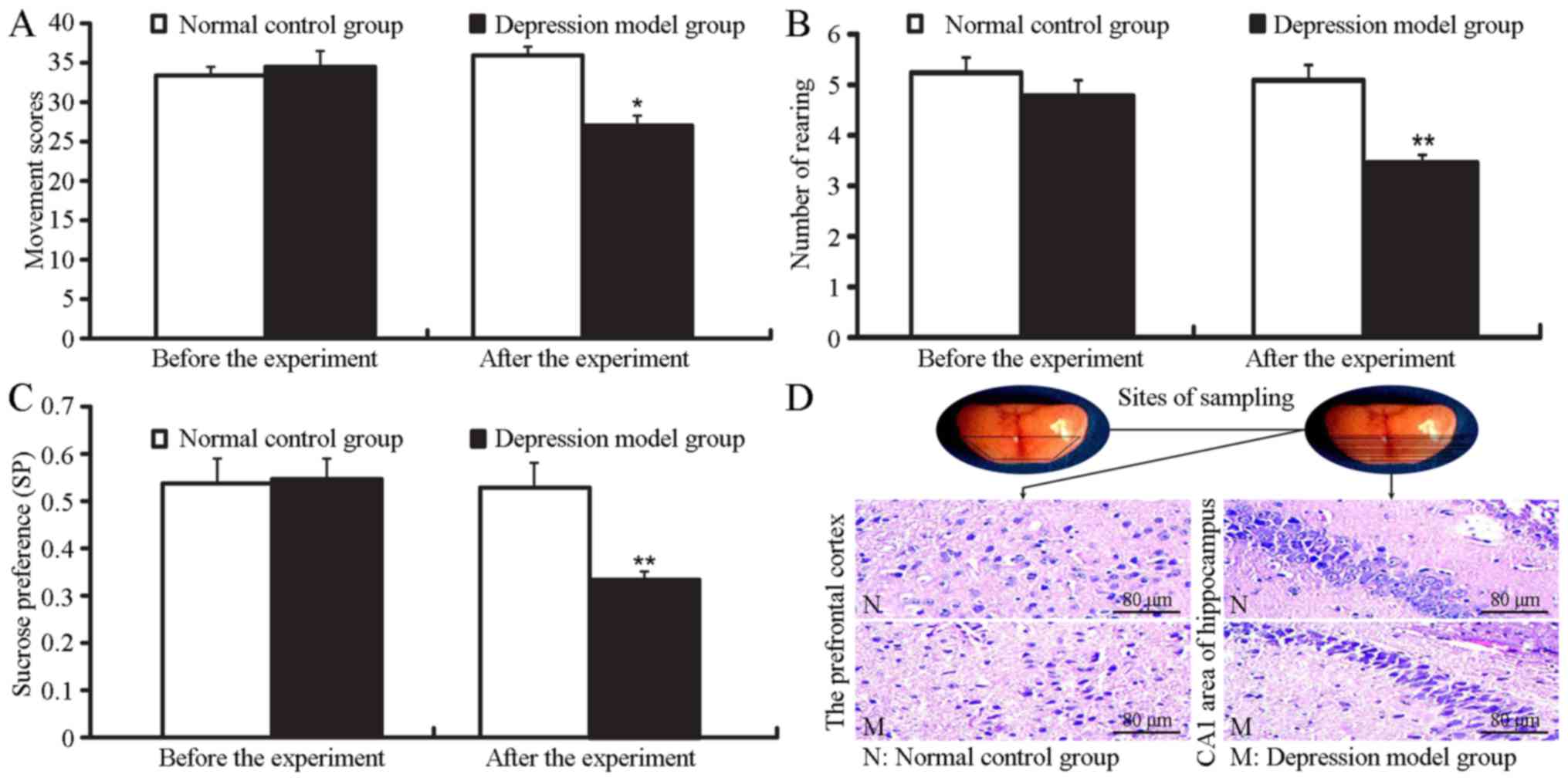
experiment (P>0.05; Fig. 1A).
The mobile scores, the number of rearings (standing on hind limbs)
and the sucrose intake of the mice in the depression model group
were significantly decreased compared with the normal control group
(mobile scores, t=2.342, P=0.030; number of rearing, t=4.814,
P=0.000; sucrose intake, t=3.829, P=0.004) (Fig. 1A–C). The indicators of depression
were strongly associated with the length of corticosterone
injections. The model of depression was successfully established on
the 21st day.
Histopathological changes
The histopathology of the cerebral cortex and
hippocampus was evaluated with H&E staining (Fig. 1D). In the normal control group,
the morphology of the neurons and glial cells in the prefrontal
cortex and hippocampal CA1 region was normal. The cellular
structures were compact with regular arrangement and a clear
hierarchy. The nucleoli were distinctly visible without any
anomalies. In the mice in the depression model group, the brain
cells exhibited an altered morphology and the cells presented a
spindle-like, or irregular shape. The cellular borders appeared
fuzzy, and the brain structures were disordered. A large number of
neurons had lost structural integrity with prominent karyopyknosis
and the disintegration of nucleoli in the prefrontal cortex.
Although the cellular arrangement of the hippocampus CA1 region
remained largely normal, the morphology of the nerve cells had
altered significantly, and some nerve cells were absent in the mice
in the depression model group.
Measurement of monoamine
Brain neurotransmitters assessed by HPLC
fluorescence method were used to further confirm the reliability of
the depression model from the biochemical point of view. The HPLC
chromatogram of a cocktail of DA, NA, Ad, Try and 5-HT standard is
presented in Fig. 2A. The peak of
each neurotransmitter in the mixed standard solution was well
separated and no other significant peaks were present. A series of
standard solutions was prepared with a concentration gradient and
injected into HPLC. The value of peak areas and concentrations with
reference to the each neurotransmitter standard was used to carry
out linear regression analysis and create standard curves. As a
result, the regression equation and correlation co-efficient of
each neurotransmitter is shown in Table I. The results indicated that the
peak area had a significant linear correlation with the content
level of NE, Ad, DA, Try or 5-HT. The contents of each
neurotransmitter in the brains of the mice were then respectively
calculated using the regression equations. The chromatogram of a
sample from one of the experimental groups is shown in Fig. 2B and C. The peak shape of each
neurotransmitter was well delineated, and there was no overlap,
double peak and bad tail phenomenon. There were two unknown,
well-separated peaks between the peak of Ad and DA, which were
worthy of note. All the neurotransmitters in the prefrontal cortex
of the mice in the depression model group yielded a lower value and
peak size than those of the control group (levels of
neurotransmitters: NA, t=2.606, P=0.040; Ad, t=2.930, P=0.026; DA,
t=4.931, P=0.003; Try, t=6.530, P=0.001; 5-HT, t=2.819, P=0.030;
and peak size: NA, t=2.672, P=0.037; Ad, t=3.544, P=0.012; DA,
t=7.442, P=0.000; Try, t=3.608, P=0.011; 5-HT, t=2.612, P=0.040)
(Fig. 2D and E).
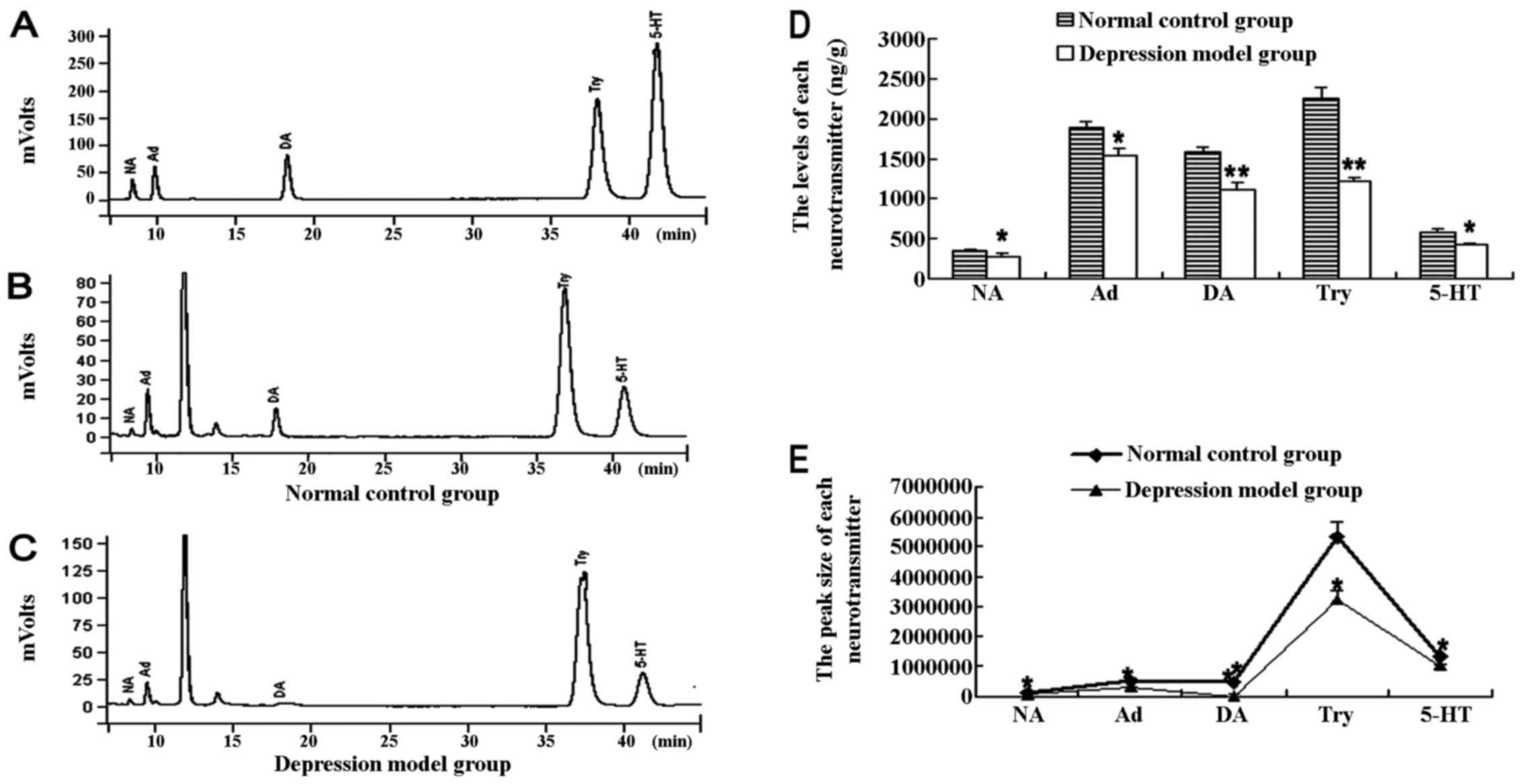 | Figure 2The content and the peak area of
neurotransmitters in the prefrontal cortex (n=6). (A) High
performance liquid chromatography (HPLC) chromatogram of the
standard substance in noradrenaline (NA) (1 µg/ml),
adrenaline (Ad) (1 µg/ml), dopamine (DA) (2 µg/ml),
tryptophan (Try) (1 µg/ml) and 5-hydroxytryptamine (5-HT) (1
µg/ml). The retention time of NA, Ad, DA, Try and 5-HT is
~7, 9, 18, 37 and 41 min, respectively. (B and C) HPLC
chromatograms of neurotransmitters in the brain samples. The
retention time of NA, Ad, DA, Try and 5-HT in the sample is the
same as each standard substance. (D and E) Contents (ng/g) and peak
area of each neurotransmitter in the sample. Summary data are
expressed as the means ± SEM. As indicated, the content and the
peak size of each neurotransmitter in the prefrontal cortex of
depressed mice were lower than those of control group (P<0.05 or
P<0.01); *P<0.05 or **P<0.01
compared with normal control group. |
 | Table IThe linear regression equation,
linear range and the correlation co-efficient of 5
neurotransmitters were measured using the HPLC fluorescence
method. |
Table I
The linear regression equation,
linear range and the correlation co-efficient of 5
neurotransmitters were measured using the HPLC fluorescence
method.
|
Neurotransmitters | Linear
equation | Linear range
(µg/ml) | Correlation
co-efficient (r) |
|---|
| NA |
y=5.29·105x+6.19·103 | 0.0625–1.0 | 0.999815 |
| Ad |
y=7.34·105x+8.66·103 | 0.0625–1.0 | 0.998305 |
| DA |
y=9.32·105x+1.61·104 | 0.125–2.0 | 0.999789 |
| Try |
y=8.12·106x−3.00·105 | 0.0625–1.0 | 0.996419 |
| 5-HT |
y=1.35·107x−1.15·106 | 0.0625–1.0 | 0.995741 |
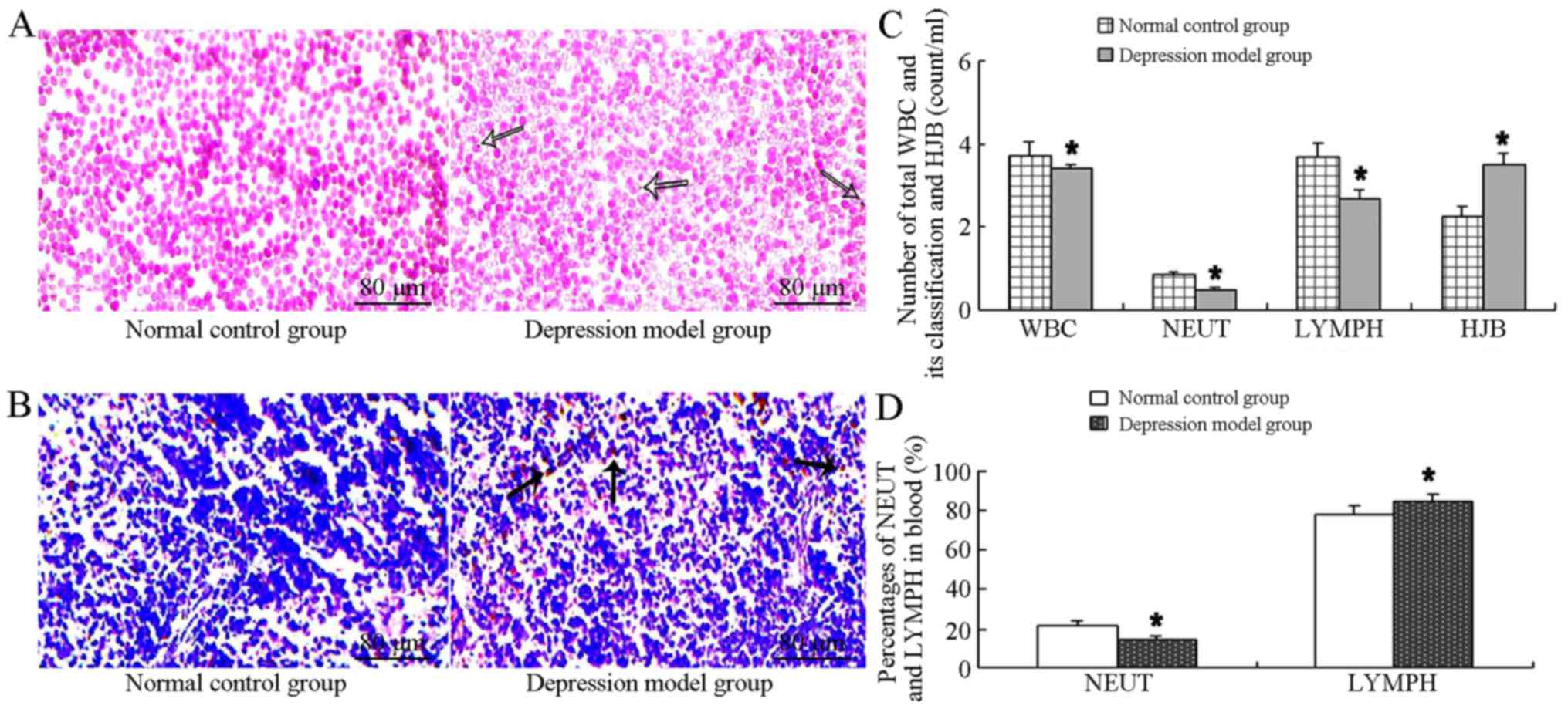
Changes in splenic function and
pathology
The changes in splenic function in the mice in the
depression model group was one of our main areas of focus. The
numbers of HJB and WBC in the blood correlate with splenic function
(21). HJB, WBC, neutrophils and
lymphocytes were counted using a light microscope following Wright
staining. In the control group, cellular morphology was normal
without obvious HJB. In the mice in the depression model group,
cellular size and staining were mildly abnormal and HJB were clear
and obvious (where the virtual arrow is pointing) (Fig. 3A). The histopathology was observed
using H&E staining of the splenic sections in both groups
(Fig. 3B). In the control group,
the white pulp was stained dark-blue, and was round or
ellipse-shaped. Cellular structures were dense with numerous
lymphocytes and lymphoid nodules. In the mice in the depression
model group, the cellular structures of white pulp became sparse,
lymphocytes and lymphoid nodules were diminished, and hemosiderin
was clearly evident in the splenic tissue. As shown in Fig. 3C and D, the counts of HJB and the
lymphocyte ratio in the depression model group increased greatly
compared with the normal control group (counts of HJB: t=−3.273,
P=0.017; lymphocyte ratio: t=2.755, P=0.033). However, the number
of neutrophils, WBC and lymphocytes and the ratio of neutrophils
was significantly decreased compared with the normal control group
(number of neutrophils: t=3.545, P=0.012; WBC, t=2.575, P=0.042;
lymphocytes: t=2.696, P=0.036; ratio of neutrophils: t=3.658,
P=0.011).
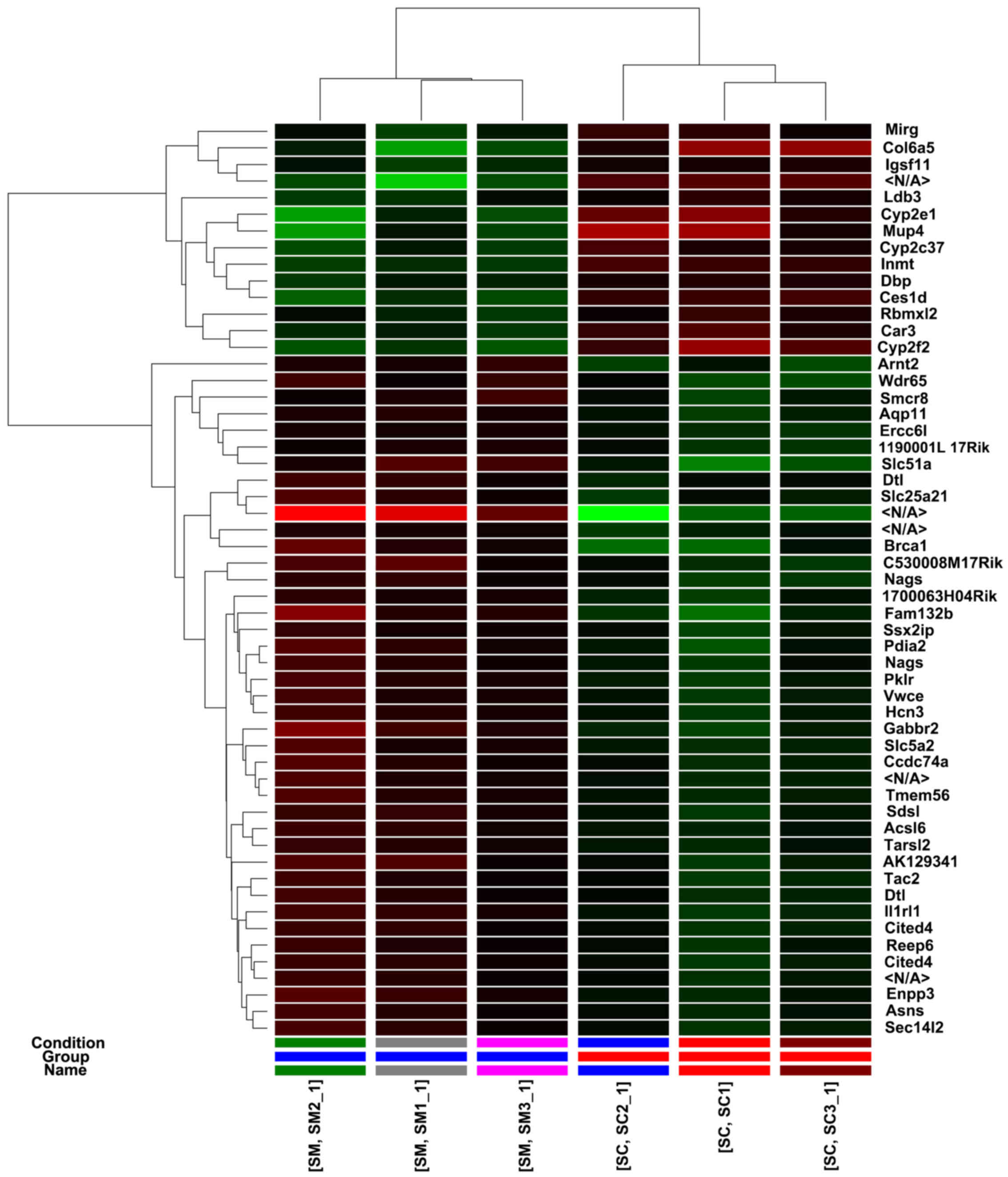
Differential expression of genes
The differential expression of genes in the spleen
was another area of focus in this study. We selected differentially
expressed genes whose value (P<0.05) and the gene expression
levels were increased >2-fold or decreased >2-fold when
comparing the depression model group with the normal control group.
The results indicated that there were 53 differentially expressed
mRNAs in the spleen when comparing the depression model group with
the normal control group, including miRNA containing gene
(Mirg), collagen, type VI, α5 (Col6a5),
immunoglobulin super-family, member 11 (Igsf11), LIM domain
binding 3 (Ldb3), cytochrome P450 family 2 subfamily E
member 1 (Cyp2e1), major urinary protein 4 (Mup4),
cytochrome P450, family 2, subfamily c, polypeptide 37
(Cyp2c37), indolethylamine N-methyltransferase
(Inmt), Dbp, carboxylesterase 1D (Ces1d), RNA binding
motif protein, X-linked-like 2 (Rbmxl2), carbonic anhydrase
3 (Car3), cytochrome P450 family 2 subfamily F member 1
(Cyp2f2), aryl hydrocarbon receptor nuclear translocator,
class E basic helix-loop-helix protein 2 (Arnt2), cilia and
flagella associated protein 57 (Cfap57, also known as
Wdr65), Smith-Magenis syndrome chromosome region, candidate
8 (Smcr8), aquaporin 11 (Aqp11), ERCC excision repair
6 like, spindle assembly checkpoint helicase (Ercc6l),
1190001L17Rik, solute carrier family 51, alpha subunit
(Slc51a), denticleless E3 ubiquitin protein ligase homolog
(Dtl), solute carrier family 25 member 21 (Slc25a21), breast
and ovarian cancer susceptibility protein 1 (Brca1),
C530008M17Rik, N-acetylglutamate synthase (Nags),
1700063H04Rik, erythroferrone (Erfe, also known as
Fam132b), SSX family member 2 interacting protein
(Ssx2ip), protein disulfide isomerase family A member 2
(Pdia2), pyruvate kinase, liver and RBC (Pklr), von
Willebrand factor C and EGF domain-containing protein
(Vwce), hyperpolarization activated cyclic nucleotide gated
potassium channel 3 (Hcn3), Gabbr2, solute carrier family 5
member 2 (Slc5a2), coiledcoil domain containing 74A
(Ccdc74a), transmembrane protein 56 (Tmem56), serine
dehydratase like (Sdsl), acyl-CoA synthetase long-chain
family member 6 (Acsl6), Threonyl-TRNA synthetase like 2
(Tarsl2), AK129341, tachykinin 2 (Tac2), interleukin
1 rReceptor like 1 (Il1rl1), Cbp/p300 interacting
transactivator with Glu/Asp rich carboxy-terminal domain 4
(Cited4), receptor accessory protein 6 (Reep6),
ectonucleotide pyrophosphatase/phosphodiesterase family member 3
(Enpp3), asparagine synthetase (Asns), SEC14 like
lipid binding 2 (Sec14l2) and 5 genes of unknown function
(Fig. 4). There were 14
upregulated mRNAs and 39 downregulated mRNAs among these genes.
The sieving captured 53 differentially expressed
genes annotated to the system of 'GO-pathway'. A GO enrichment
analysis of differentially expressed mRNAs was performed to
identify GOs with a higher confidence. Enrichment provides a
significant means of measuring function. The corresponding function
is more specific with the enrichment increasing, which helps us to
identify GOs with more concrete functional description in the
experiment (22). After this
process, the results revealed that the amino acid synthesis and the
clock gene were more significantly affected in two pathways
associated with depression among these genes. According to
microarray analysis and the literature data (23,24), we also found 11 of 55
differentially expressed genes in the spleen, which can influence
immune responses, inflammation, metabolism, collagen formation,
cell factor, protein transport and nerve myelin formation, were
closely linked to depression (Table
II).
| Table IIThe differently expressed genes in
spleen associated with depression (n=9). |
Table II
The differently expressed genes in
spleen associated with depression (n=9).
| No. | GenBank no. | Gene name | Gene
symbol | Fold change | Regulation | P-value |
|---|
| 1 | NM_001167923 | Collagen, type VI,
α5 | Col6a5 | 10.022 | Up | 0.027 |
| 2 | NM_170599 | Immunoglobulin
superfamily, member 11 | Igsf11 | 2.140 | Up | 0.039 |
| 3 | NM_016974 | D site albumin
promoter binding protein | Dbp | 2.280 | Up | 0.015 |
| 4 | NM_144520 | SEC14-like 2 (S.
cerevisiae) | Sec14l2 | 2.304 | Down | 0.034 |
| 5 | NM_010743 | Interleukin 1
receptor-like 1 | Il1rl1 | 2.632 | Down | 0.009 |
| 6 | NM_013631 | Pyruvate kinase
liver and red blood cell | Pklr | 2.577 | Down | 0.015 |
| 7 | NM_001033599 | Acyl-CoA synthetase
long-chain family member 6 | Acsl6 | 2.13 | Down | 0.020 |
| 8 | D14423 | Tachykinin 2 | Tac2 | 2.19 | Down | 0.038 |
| 9 | NM_001081070 | Protein disulfide
isomerase associated 2 | Pdia2 | 2.985 | Down | 0.039 |
| 10 | NM_134005 | Ectonucleotide
pyrophos-phatase/phosphodiesterase 3 | Enpp3 | 2.445 | Down | 0.043 |
| 11 | NM_001081141 | γ-aminobutyric acid
(GABA) B receptor, 2 | Gabbr2 | 3.717 | Down | 0.040 |
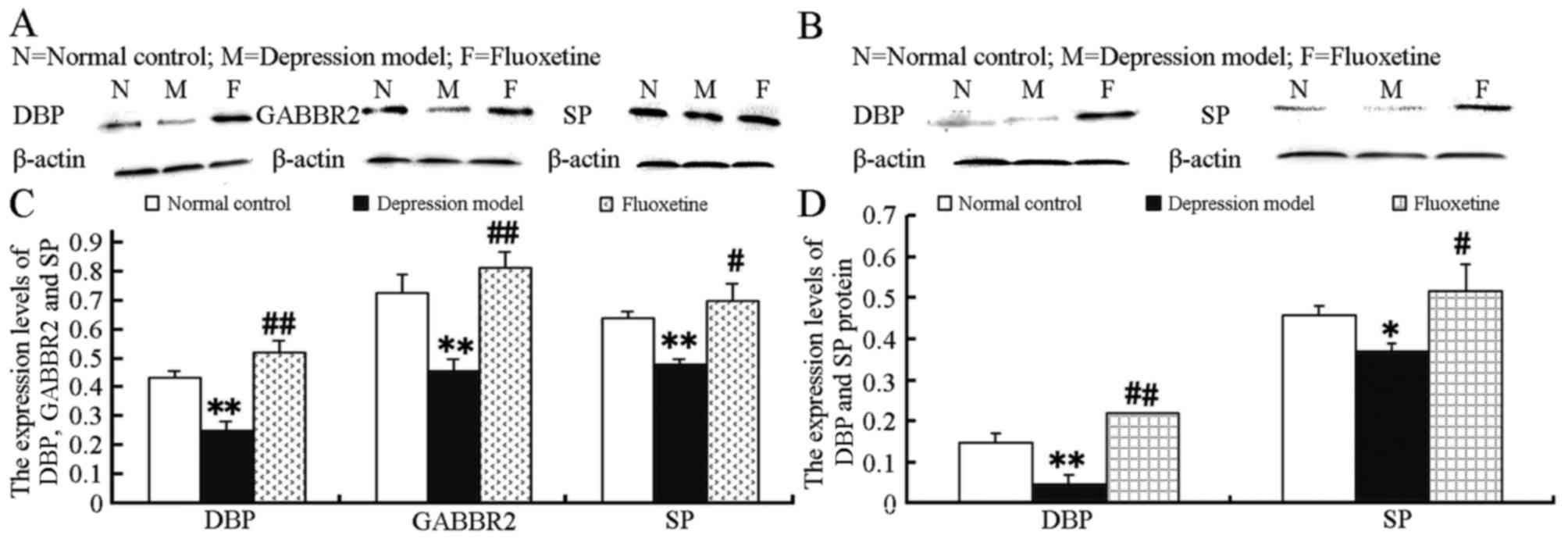
Expression levels of GABBR2, DBP and SP
related proteins
The DBP, GABBR2 and SP proteins in the spleen and
brain were further detected. As shown in Fig. 5A, the protein expression levels of
GABBR2, DBP and SP in the brain were significantly decreased in the
model group compared to those in the control group (GABBR2, t=6.61,
P=0.000; SP, t=6.045, P=0.001; DBP, t=6.956, P=0.000). GABBR2
proteins were not detected in the spleen (Fig. 5B), and the alterations in the DBP
and SP protein levels in the spleen were coincident with those in
the brain (DBP, t=7.496, P=0.00; SP, t=3.651, P=0.006). Moreover,
following treatment with fluoxetine, the protein expression levels
of DBP, SP or GABBR2 in the prefrontal cortex and the spleen were
markedly enhanced in the mice in the depression model group (in
prefrontal cortex: DBP, t=9.02, P=0.000; SP, t=3.587, P=0.007;
GABBR2, t=4.843, P=0.001; and in spleen: DBP, t=19.81, P=0.000; SP,
t=3.574, P=0.007).
Discussion
There is accumulating evidence to support a close
connection between stress and depression (25,26). Glucocorticoids are characteristic
mediators in stress responses, and the detrimental effects of
chronic stress on health have been proposed to be attributed to
their immunosuppressive properties (27). Thus, in this study, we examined
the histopathology of the spleen, the splenic function and gene
expression in mice with depressive-like behavior, as well as the
expression of certain proteins in the related pathways. To the best
of our knowledge, we demonstrate for the first time that splenic
function in mice with depressive-like behavior was markedly
impaired and we also found some differentially expressed genes in
the spleen. In addition, the levels of proteins in related pathways
were also altered in the mice with depressive-like behavior.
The present study had five main findings: first,
depression induces significant damage to the morphology and
function of the spleen in mice. Second, 53 genes exhibited a
differential response in the spleen of the mice in the depression
model group, 11 of which were more notable. Third, the amino acid
biosynthesis and the clock gene pathways were more markedly
affected in the spleen. Fourth, the related proteins in the
pathways were downregulated in the depressed mice. Fifth,
fluoxetine reversed the changes in GABBR2, DBP and SP protein
levels in the brain and spleen. The results obtained in the spleen
confirmed our hypothesis.
Counting the number of erythrocytes with HJB in
blood smear under a light microscope is a commonly used method to
assess splenic function (28).
When the spleen is absent or insufficient, the number of HJB
containing erythrocytes is increased (29). In the present study, the mice in
the depression model group exhibited higher counts of HJB, which
indicated that splenic function was compromised.
The numbers of neutrophils, WBC and lymphocytes and
the ratio of neutrophils were significantly decreased, while the
ratio of lymphocytes was elevated greatly in the depression model
group compared to the normal control group. These are highly
consistent with the clinical data that approximately 80% of
clinical patients exhibit a low neutrophil ratio and a high
lymphocytes ratio (unpublished data). The changes in lymphatic
structure and the increase in hemosiderosis suggested that the
splenic function and immune response declined (30). At the same time, this study found
the neurons and glial cells in the frontal cortex and hippocampus
were deformed or damaged, and the depressive behaviors of the mice
correlated well with low levels of NE, Ad, DA, Try and 5-HT in the
brain. These results are consistent with those of previous studies
on brain tissue in depression (31,32). This indicates that the function of
the spleen may play an important role in the pathogenesis of
depression.
Changes in gene expression are well documented in
depression and schizophrenia, and may contribute to the pathologic
phenotypes associated with these disorders (33). Although the onset of depression
cannot be attributed to a single gene, multiple small genetic
effects along with environmental factors have been shown to trigger
major depressive episodes (34,35). In this study, when analyzing gene
expression profiles in the spleen, we found 53 differentially
expressed mRNAs in the mice in the depression model group, and two
signaling pathways were more affected. A total of 11 of 53
differentially expressed genes in the spleen wre closely associated
with depression, of which many differentially expressed genes were
also screened in other experiments as differentially expressed
genes in the brain (21). Based
on these results, we can infer that the changes in the levels of
genes in the spleen may also be one of the pathological mechanisms
of depression.
Certain studies have reported that the decreased
protein levels of GABBR2 or elevated SP protein levels in the brain
are associated with depression (36,37), and DBP-knockout mice exhibit
bipolar disorder (38). We found
that although there was no GABBR2 protein expression in the spleen
(39), the expression of the
other two proteins in the spleen was generally consistent with that
in the brain: compared to the control group, the protein expression
of DBP and SP was significantly decreased. The results of GABBR2
and SP protein expression in the frontal cortex, and SP in the
spleen were coincident with those of microarray analysis. Of note,
although the Dbp gene in the spleen was upregulated, its
protein level was downregulated in the brains and spleens of the
mice in the depression model group. This result is basically
consistent with that of a previous stutdy (38); the effects of depression were
reversed by fluoxetine.
In addition, we also noted the discordant modulation
of SP in depression. Despite that vast majority of published
studies indicates that serum and cerebrospinal fluid SP
concentrations in depressed patients are significantly higher than
in healthy subjects (40,41), elevated SP levels have also been
reported in patients with major depressive disorder under
anti-depressant therapy (42),
which are in line with our results of reduced SP protein expression
in mice with depressive-like behavior, and the same reversed effect
of fluoxetine. Indirectly, this reflects a close association
between depression and changes in splenic gene expression.
As demonstrated in the present study, the
administration of corticosterone in mice affected behavioral and
neurobiological changes that mirror some of the core symptoms and
neurobiological changes associated with major depression. However,
the association between spleen and depression has not gained enough
attention. The results of this study suggest that the improvement
of splenic function and splenic gene expression may help patients
with depression; however, it cannot be determined whether
splenectomy or splenic injuries can lead to depression. A
cause-and-effect relationship between the changes in the spleen and
depression is not yet clear, and further research is required. In
conclusion, although preliminary, this study sheds light on splenic
changes in animal models of depression and indicate a strong
connection between depression and changes in splenic function,
pathomorphology and multiple genes, which may lay the ground work
for future studies on the pathogenesis in depression.
Acknowledgments
This study was supported by grants from the China
National Natural Science Funds (nos. 81470510 and 81170597), the
Science and Technology Planned Project of Henan Province (no.
152300410119) and the Natural Science Foundation of Henan Province
(no. 162300410219). We would like to thank Thomas Q. Zheng at
Maricopa Medical Center, University of Arizona College of
Medicine-Phoenix, for reviewing the manuscript, and for performing
language modifications and editing.
References
|
1
|
Lépine JP and Briley M: The increasing
burden of depression. Neuropsychiatr Dis Treat. 7(Suppl 1): 3–7.
2011.PubMed/NCBI
|
|
2
|
Arroll B, Macgillivray S, Ogston S, Reid
I, Sullivan F, Williams B and Crombie I: Efficacy and tolerability
of tricyclic antidepressants and SSRIs compared with placebo for
treatment of depression in primary care: A meta-analysis. Ann Fam
Med. 3:449–456. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kok RM, Heeren TJ and Nolen WA: Continuing
treatment of depression in the elderly: A systematic review and
meta-analysis of double-blinded randomized controlled trials with
antidepressants. Am J Geriatr Psychiatry. 19:249–255. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
McIntyre RS: When should you move beyond
first-line therapy for depression? J Clin Psychiatry. 71(Suppl 1):
16–20. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shrestha P, Mousa A and Heintz N: Layer
2/3 pyramidal cells in the medial prefrontal cortex moderate stress
induced depressive behaviors. Elife. 4:e087522015. View Article : Google Scholar :
|
|
6
|
Papadopoulou A, Siamatras T,
Delgado-Morales R, Amin ND, Shukla V, Zheng YL, Pant HC, Almeida OF
and Kino T: Acute and chronic stress differentially regulate
cyclin-dependent kinase 5 in mouse brain: Implications to
glucocorticoid actions and major depression. Transl Psychiatry.
5:e5782015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Blume J, Douglas SD and Evans DL: Immune
suppression and immune activation in depression. Brain Behav Immun.
25:221–229. 2011. View Article : Google Scholar :
|
|
8
|
Kiank C, Zeden JP, Drude S, Domanska G,
Fusch G, Otten W and Schuett C: Psychological stress-induced,
IDO1-dependent tryptophan catabolism: Implications on
immunosuppression in mice and humans. PLoS One. 5:e118252010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hernandez ME, Martinez-Mota L, Salinas C,
Marquez-Velasco R, Hernandez-Chan NG, Morales-Montor J, Pérez-Tapia
M, Streber ML, Granados-Camacho I, Becerril E, et al: Chronic
stress induces structural alterations in splenic lymphoid tissue
that are associated with changes in corticosterone levels in
wistar-kyoto rats. BioMed Res Int. 2013:8687422013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Park AJ, Collins J, Blennerhassett PA,
Ghia JE, Verdu EF, Bercik P and Collins SM: Altered colonic
function and microbiota profile in a mouse model of chronic
depression. Neurogastroenterol Motil. 25:733–e575. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Voorhees JL, Tarr AJ, Wohleb ES, Godbout
JP, Mo X, Sheridan JF, Eubank TD and Marsh CB: Prolonged restraint
stress increases IL-6, reduces IL-10, and causes persistent
depressive-like behavior that is reversed by recombinant IL-10.
PLoS One. 8:e584882013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ago Y, Yano K, Araki R, Hiramatsu N, Kita
Y, Kawasaki T, Onoe H, Chaki S, Nakazato A, Hashimoto H, et al:
Metabotropic glutamate 2/3 receptor antagonists improve behavioral
and prefrontal dopaminergic alterations in the chronic
corticosterone-induced depression model in mice. Neuropharmacology.
65:29–38. 2013. View Article : Google Scholar
|
|
13
|
Zhou Y, Lu L, Li Z, Gao X, Tian J, Zhang
L, Wu B and Qin X: Antidepressant-like effects of the fractions of
Xiaoyaosan on rat model of chronic unpredictable mild stress. J
Ethnopharmacol. 137:236–244. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee B, Sur B, Yeom M, Shim I, Lee H and
Hahm DH: L-tetrahydropalmatine ameliorates development of anxiety
and depression-related symptoms induced by single prolonged stress
in rats. Biomol Ther (Seoul). 22:213–222. 2014. View Article : Google Scholar
|
|
15
|
Bhatt S, Radhakrishnan M, Jindal A,
Devadoss T and Dhar AK: Neuropharmacological evaluation of a novel
5-HT3 receptor antagonist (6g) on chronic unpredictable mild
stress-induced changes in behavioural and brain oxidative stress
parameters in mice. Indian J Pharmacol. 46:191–196. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ren J, Fan C, Chen N, Huang J and Yang Q:
Resveratrol pretreatment attenuates cerebral ischemic injury by
upregulating expression of transcription factor Nrf2 and HO-1 in
rats. Neurochem Res. 36:2352–2362. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lv QQ, Wu WJ, Guo XL, Liu RL, Yang YP,
Zhou DS, Zhang JX and Liu JY: Antidepressant activity of astilbin:
Involvement of monoaminergic neurotransmitters and BDNF signal
pathway. Biol Pharm Bull. 37:987–995. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dor FJ, Gollackner B, Kuwaki K, Ko DS,
Cooper DK and Houser SL: Histopathology of spleen allograft
rejection in miniature swine. Int J Exp Pathol. 86:57–66. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Angst F, Stassen HH, Clayton PJ and Angst
J: Mortality of patients with mood disorders: Follow-up over 34–38
years. J Affect Disord. 68:167–181. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jangra A, Lukhi MM, Sulakhiya K, Baruah CC
and Lahkar M: Protective effect of mangiferin against
lipopolysaccharide-induced depressive and anxiety-like behaviour in
mice. Eur J Pharmacol. 740:337–345. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kohan G, Ocampo CG, Zandalazini HI,
Klappenbach R, Quesada BM, Porras LT, Rodriguez JA and Oria AS:
Changes in gastrosplenic circulation and splenic function after
distal pancreatectomy with spleen preservation and splenic vessel
excision. J Gastrointest Surg. 17:1739–1743. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Schlitt T, Palin K, Rung J, Dietmann S,
Lappe M, Ukkonen E and Brazma A: From gene networks to gene
function. Genome Res. 13:2568–2576. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lagus M, Gass N, Saharinen J, Savelyev S,
Porkka-Heiskanen T and Paunio T: Inter-tissue networks between the
basal forebrain, hippocampus, and prefrontal cortex in a model for
depression caused by disturbed sleep. J Neurogenet. 26:397–412.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li JZ, Bunney BG, Meng F, Hagenauer MH,
Walsh DM, Vawter MP, Evans SJ, Choudary PV, Cartagena P, Barchas
JD, et al: Circadian patterns of gene expression in the human brain
and disruption in major depressive disorder. Proc Natl Acad Sci
USA. 110:9950–9955. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guan XT, Lin WJ and Tang MM: Comparison of
stress-induced and LPS-induced depressive-like behaviors and the
alterations of central proinflammatory cytokines mRNA in rats.
Psych J. 4:113–122. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kim MH and Leem YH: Chronic exercise
improves repeated restraint stress-induced anxiety and depression
through 5HT1A receptor and cAMP signaling in hippocampus. J Exerc
Nutrition Biochem. 18:97–104. 2014. View Article : Google Scholar
|
|
27
|
Elenkov IJ: Glucocorticoids and the
Th1/Th2 balance. Ann NY Acad Sci. 1024:138–146. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lipson RL, Bayrd ED and Watkins CH: The
postsplenectomy blood picture. Am J Clin Pathol. 32:526–532. 1959.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Marques RG, Lucena SB, Caetano CE, de
Sousa VO, Portela MC and Petroianu A: Blood clearance of
Howell-Jolly bodies in an experimental autogenic splenic implant
model. Br J Surg. 101:820–827. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Paulo DN, Paulo IC, Kalil M, Vargas PM,
Silva AL, Baptista JF and Guerra AJ: Subtotal splenectomy
preserving the lower pole in rats: Technical, morphological and
functional aspects. Acta Cir Bras. 21:321–327. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jin ZL, Gao N, Li XR, Tang Y, Xiong J,
Chen HX, Xue R and Li YF: The antidepressant-like pharmacological
profile of Yuanzhi-1, a novel serotonin, norepinephrine and
dopamine reuptake inhibitor. Eur Neuropsychopharmacol. 25:544–556.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lima A, Sardinha VM, Oliveira AF, Reis M,
Mota C, Silva MA, Marques F, Cerqueira JJ, Pinto L, Sousa N and
Oliveira JF: Astrocyte pathology in the prefrontal cortex impairs
the cognitive function of rats. Mol Psychiatry. 19:834–841. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Raison CL and Miller AH: The
neuroimmunology of stress and depression. Semin Clin
Neuropsychiatry. 6:277–294. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Molteni R, Macchi F and Riva MA: Gene
expression profiling as functional readout of rodent models for
psychiatric disorders. Cell Tissue Res. 354:51–60. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shelton RC, Claiborne J,
Sidoryk-Wegrzynowicz M, Reddy R, Aschner M, Lewis DA and Mirnics K:
Altered expression of genes involved in inflammation and apoptosis
in frontal cortex in major depression. Mol Psychiatry. 16:751–762.
2011. View Article : Google Scholar
|
|
36
|
Fatemi SH, Folsom TD, Rooney RJ and Thuras
PD: Expression of GABAA α2-, β1- and ε-receptors are altered
significantly in the lateral cerebellum of subjects with
schizophrenia, major depression and bipolar disorder. Transl
Psychiatry. 3:e3032013. View Article : Google Scholar
|
|
37
|
Zuo HX, Li JQ, Han B, Ke CJ, Liu XD, Zhang
YC, Li L and Yang X: Di-(n-butyl)-phthalate-induced oxidative
stress and depression-like behavior in mice with or without
ovalbumin immunization. Biomed Environ Sci. 27:268–280.
2014.PubMed/NCBI
|
|
38
|
Le-Niculescu H, McFarland MJ, Ogden CA,
Balaraman Y, Patel S, Tan J, Rodd ZA, Paulus M, Geyer MA, Edenberg
HJ, et al: Phenomic, convergent functional genomic, and biomarker
studies in a stress-reactive genetic animal model of bipolar
disorder and co-morbid alcoholism. Am J Med Genet B Neuropsychiatr
Genet. 147B. pp. 134–166. 2008, View Article : Google Scholar
|
|
39
|
Bettler B, Kaupmann K, Mosbacher J and
Gassmann M: Molecular structure and physiological functions of
GABA(B) receptors. Physiol Rev. 84:835–867. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Rimón R, Le Grevés P, Nyberg F, Heikkilä
L, Salmela L and Terenius L: Elevation of substance P-like peptides
in the CSF of psychiatric patients. Biol Psychiatry. 19:509–516.
1984.PubMed/NCBI
|
|
41
|
Bondy B, Baghai TC, Minov C, Schüle C,
Schwarz MJ, Zwanzger P, Rupprecht R and Möller HJ: Substance P
serum levels are increased in major depression: Preliminary
results. Biol Psychiatry. 53:538–542. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cizza G, Marques AH, Eskandari F, Christie
IC, Torvik S, Silverman MN, Phillips TM and Sternberg EM; POWER
Study Group: Elevated neuroimmune biomarkers in sweat patches and
plasma of premenopausal women with major depressive disorder in
remission: The POWER study. Biol Psychiatry. 64:907–911. 2008.
View Article : Google Scholar : PubMed/NCBI
|