Introduction
Astrocytes are the most abundant glial subtype in
the central nervous system (CNS) and play important roles in
interacting with neurons and communicating with other neural cell
types (1–4). In spinal cord injury (SCI),
astrocytes proximal to lesion zones become reactive. The process of
reactive astrogliosis involves the migration and proliferation of
astrocytes, contributing to glial scar formation, which is a major
impediment to axonal regeneration. Overcoming this physical and
biochemical barrier may be crucial for axonal regeneration and
functional compensation during the progression of SCI (5,6).
Although extensive investigations have been conducted on SCI
(7), no effective therapy has yet
been established. The lack of effective interventions is at least
partially attributed to the complex process and the failure of
axonal regeneration due to glial scaring (7). There is therefore a need to develop
therapeutic strategies with which to prevent reactive
astrogliosis.
Fisetin (3,3′,4′,7-tetrahydroxyflavone) is a dietary
flavonoid found in fruits and vegetables, including strawberries,
persimmons, grapes, cucumbers and onions (8). It has extensive physiological and
pharmacological activities, including antioxidant,
anti-inflammatory, anti-neoplastic and neuroprotective effects
(8–10). It has also been shown that fisetin
enhances behavioral performance and attenuates reactive gliosis and
inflammation during aluminum chloride–induced neurotoxicity
(9). Furthermore, fisetin
suppresses oxidative and neuroinflammatory responses in microglial
cells (10). However, the effects
of fisetin on reactive astrocytes remain unclear. In the present
study, we examined whether fisetin inhibits reactive astrogliosis
in an in vitro scratch-wound model, and then characterized
its underlying mechanisms of action.
Materials and methods
Materials and reagents
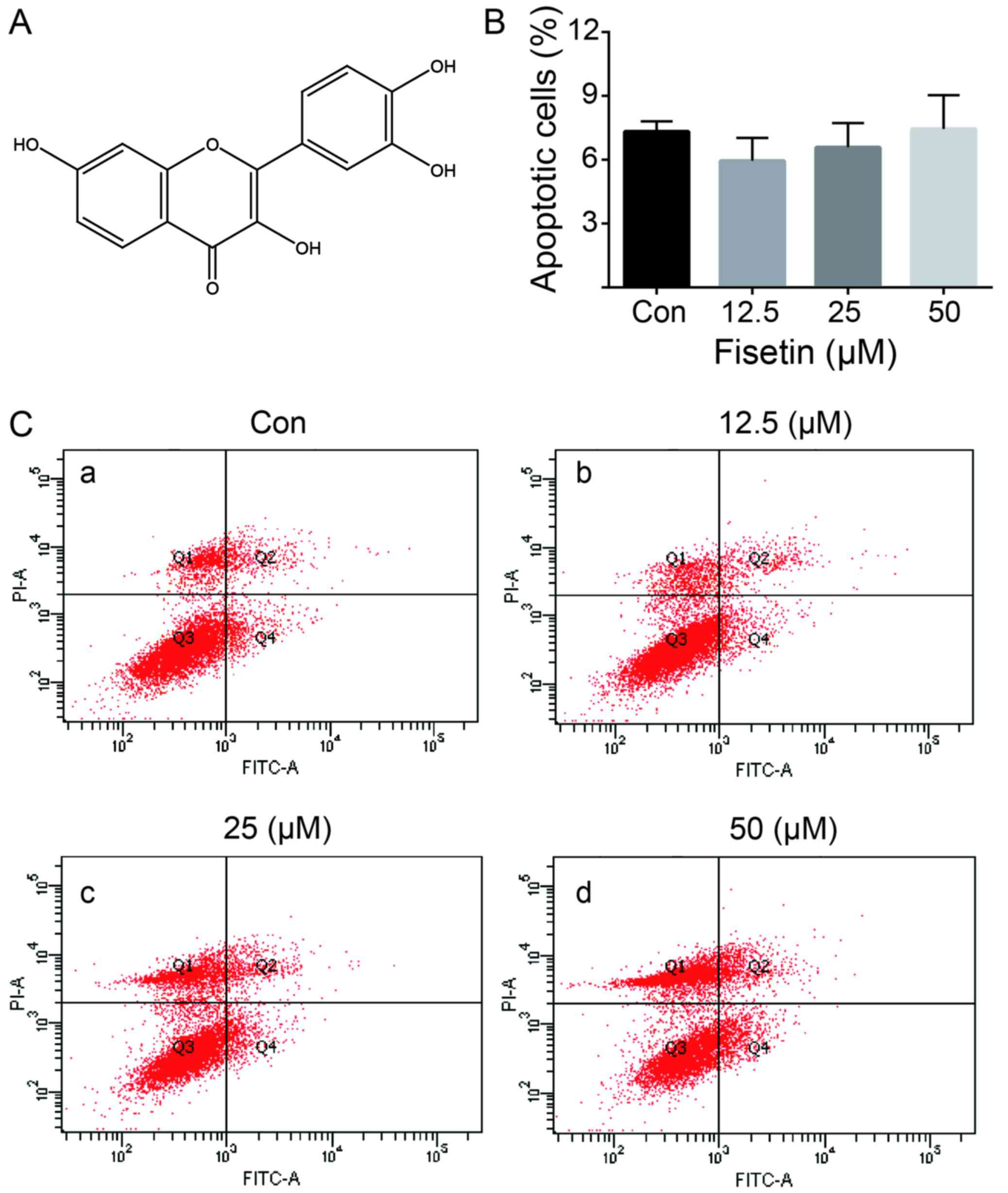
Fisetin (Fig. 1A)
(molecular weight, 286.24; Sigma-Aldrich, St. Louis, MO, USA) was
used at a purity of >98%. The molecular structure has been
deposited in a publicly accessible database in PubChem with
research number: CID 5281614. Stock solutions of fisetin were
prepared in dimethyl sulfoxide (DMSO; Sigma-Aldrich) and stored at
−20°C.
Cell culture
Primary astrocyte cultures were prepared from
1-day-old Sprague-Dawley rat brains (Animal Experiment Center,
Southern Medical University, Guangzhou, China), as previously
described (11,12). Ethics approval was granted by the
Medical Ethics Committee of Southern Medical University, Guangzhou,
China. Briefly, a total of 12 rats (n=3/group) were deeply
anesthetized with a mixture of ketamine and xylazine (70:6 mg/kg,
intramuscular injection) and then decapitated, the striatums were
dissected, and the meninges were removed. The cerebral hemisphere
was freed of the meninges and cut into 1 mm3 cubes in
Dulbecco's modified Eagle's medium (DMEM; Gibco Life Technologies,
Fresno, CA, USA). The tissue was dissociated by vortex mixing for
60 sec, and the cell suspension was passed through 70 and 20
µm sterile mesh nylon filters. A volume of cell suspension
containing approximately 4.5×105 cells was seeded in a
35 mm2 Falcon tissue culture dish (Corning, Albany, NY,
USA). Primary astrocytes were cultured in DMEM containing 10% fetal
bovine serum (FBS) and 0.15% penicillin-streptomycin reagent (both
from Gibco Life Technologies) at 37°C with 5% CO2. The
medium was changed twice a week. Cultures of at least 4 weeks were
used in the experiments.
Scratch-wound assay
The capacity of the astrocytes to migrate was
examined by scratching confluent astrocyte monolayers to induce
natural cell migration, as previously reported (12,13). Astrocyte monolayers were wounded
by dragging a sterile 200-µl pipette tip across the surface.
The detached cells and debris were removed immediately with 3
washes of phosphate-buffered saline (PBS). The cells were then
maintained for an additional 24 h in culture medium with 1% FBS and
various concentrations of fisetin (25 and 50 µM) or 0.1%
DMSO as the control group. Images of the closing wound were
acquired under an inverted microscope (Olympus TH4-200; Olympus,
Tokyo, Japan) and analyzed using NIS Elements D3.2 (Nikon NT-88-V3;
Nikon, Tokyo, Japan).
Cell apoptosis assay
The astrocytes were pre-treated with fisetin (12.5,
25 and 50 µM) or 0.1% DMSO as the control group for 24 h.
The cells were then digested with 0.125% EDTA-free trypsin and
collected by centrifugation at 2,000 rpm for 5 min. The cells were
washed twice with PBS, resuspended in 500 µl binding buffer,
and incubated in the dark for 10 min at room temperature with 5
µl of Annexin V-Fluor 488 and 5 µl of propidium
iodide (PI) (product code: KGAV103; KeyGen, Nanjing, China). The
samples were assessed by flow cytometry (BD Biosciences, Mountain
View, CA, USA).
Click-iT 5-ethynyl-2′-deoxyuridine (EdU)
test
The astrocytes were treated simultaneously with
various concentrations of fisetin (12.5–50 µM) or 0.1% DMSO
as the control group and EdU (10 µM), using the Cell-Light™
EdU Apollo 567 in vitro flow cytometry kit (product code:
C10338-3; Guangzhou RiboBio Co., Ltd., Guangzhou, China) for 24 h.
The cells were then stained according to the following protocol:
the cells were collected by centrifugation at 1,500 rpm for 5 min,
4% paraformaldehyde was added at room temperature for 30 min to fix
the cells, followed by washing with PBS, then the addition of 0.5%
Triton X-100 for 15 min, 2 washes with PBS, and incubation with
click reaction buffer (100 mM Tris-HCl, pH 8.5; 1 mM
CuSO4; 100 µM Apollo 550 fluorescent azide; and
100 mM ascorbic acid) for 30 min in the dark, washing with 0.5%
Triton X-100, and a final addition of 500 µl PBS for the
flow cytometry analysis, within 1 h.
Cell cycle assay
The astrocytes were synchronized by starved for 12 h
in serum-free medium and treated subsequently with different
concentrations of fisetin (12.5–50 µM) or 0.1% DMSO as the
control group for 24 h prior to analysis. Cell cycle analysis was
performed by PI staining. The astrocytes were trypsinized and
washed with ice-cold PBS (pH 7.4) and fixed in ice-cold 75% ethanol
for >2 h. The cells were then washed with PBS, treated with 100
µl RNase A at 37°C for 30 min, and finally stained with 400
µl PI in the dark at 4°C for 30 min. Flow cytometric
analysis was then carried out within 1 h. The analysis was
performed using a Cell Cycle Detection Test kit (product code:
KGA512; KeyGen).
Western blot analysis
Western blot analysis was conducted as previously
described (6). The astrocytes
were treated simultaneously with various concentrations of fisetin
(12.5–50 µM) or 0.1% DMSO as the control group for 24 h
prior to analysis. The cells were then washed with ice-cold PBS,
and total proteins were extracted using the
radioimmunoprecipitation assay (RIPA; Sigma-Aldrich) buffer
supplemented with phosphatase inhibitors. The concentration of
proteins was determined by the bicinchoninic acid (BCA) method with
bovine serum albumin (BSA) as a standard using the protein
quantification kit (product code, 51254; Sigma-Aldrich). A total of
30 µg of protein from each sample was analyzed by SDS/PAGE
and transferred by electrophoresis onto polyvinylidene fluoride
(PVDF) membranes (Millipore Corp., Billerica, MA, USA). The
membranes were blocked for 1 h at room temperature by using 5%
(w/v) BSA Tris-buffered saline with Tween-20 (TBST) buffer (20 mM
Tris pH 7.5 and 150 mM NaCl). The membranes were then incubated
with primary antibodies to the following: phosphorylated (p-)focal
adhesion kinase (FAK; Tyr 576/577; product code: 3281; diluted
1:1,000), FAK (D2R2E; product code: 13009; diluted 1:1,000),
p-paxillin (Tyr118; product code: 2541; diluted 1:1,000), paxillin
(product code: 12065; diluted 1:1000), cyclin D1 (product code:
2922; diluted 1:1,000), p-Akt (s473; product code: 4060; diluted
1:2,000), Akt (pan; product code: 4691; diluted 1:1,000), p-p44/42
mitogen-activated protein kinase [MAPK; extracellular
signal-regulated kinase (Erk1/2); Thr202/Tyr204; product code:
4370, diluted 1:2,000), p44/42 MAPK (Erk1/2; 137F5; product code:
4695, diluted 1:1,000), p-p38 MAPK (Thr180/Tyr182; D3F9; product
code: 4511, diluted 1:1,000), p38 MAPK (D13E1; product code: 8690,
diluted 1:1,000) and glyceraldehyde 3-phosphate dehydrogenase
(GAPDH; product code: 5174, diluted 1:1,000. All antibodies were
from Cell Signaling Technology, Inc., (Beverly, MA, USA), and were
used at 4°C overnight. The blots were then incubated with
horseradish peroxidase-conjugated anti-rabbit IgG (product code:
7071; all diluted 1:4,500; Cell Signaling Technology, Inc.) for 1 h
at room temperature. Finally, the signals were developed using the
enhanced chemiluminescence (ECL) kit (Millipore Corp.), and
analyzed using Quantity One software (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). Protein levels were expressed as gray values
(GVs) relative to the background controls.
Quantitative analysis
The evaluation of the western blot analysis data was
carried out using the following steps: i) The GV of the western
blot bandings was examined using Image-Pro Plus (IPP) software in
the control and experimental group; ii) in each group, the GV of
the protein of interest was divided by the GV of the internal
reference; iii) the results of the experimental group from 'step
ii' were divided by the results of the control group from 'step
ii'; and iv) the final results were presented as the 'fold
change'.
Statistical analysis
Data are representative of at least 3 independent
experiments. Each experiment was carried out in triplicate. Data
were plotted using GraphPad 'PRISM' soft ware (version 3.0;
GraphPad Software, Inc., La Jolla, CA, USA). The significance of
the differences was evaluated by one-way ANOVA followed by the
Student-Newman-Keuls post hoc test. Error bars represent the
standard error of the mean (SEM) and values of P<0.05 and
P<0.01 were considered to indicate statistically significant
differences.
Results
Effect of fisetin on astrocyte
apoptosis
As shown in Fig. 1B
and C, we examined the effect of fisetin on astrocyte cell
apoptosis using flow cytometry analysis. Fisetin, at concentrations
ranging from 12.5–50 µM, did not significantly affect cell
apoptosis following treatment for 24 h.
Fisetin suppresses astrocyte
migration
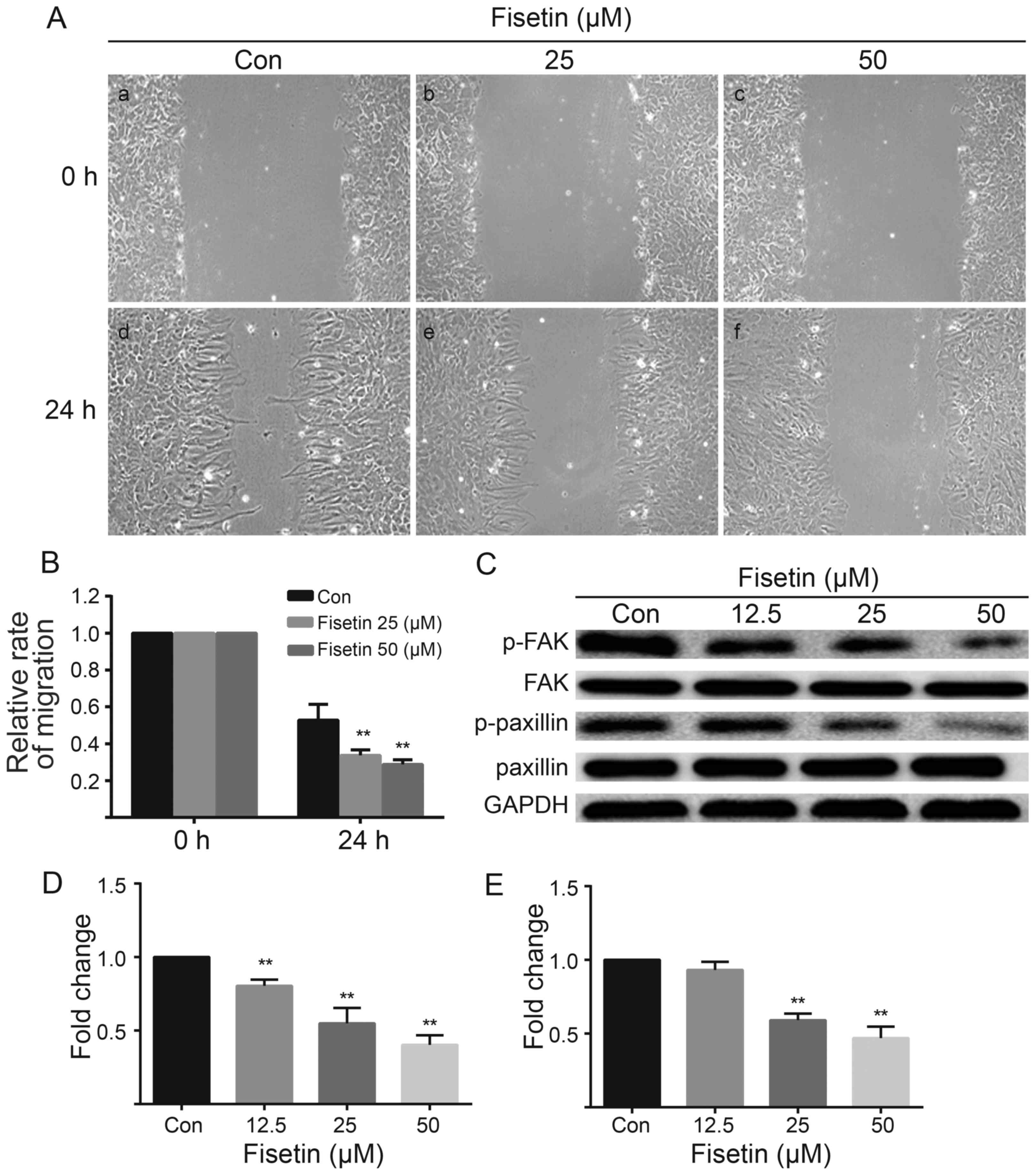
The effect of fisetin on astrocyte migration was
determined using an in vitro scratch-wound assay, in which
the loss of cell contact induces local chemokine release, which in
turn stimulates the cells to close the scratch (12,13). Our results revealed that fisetin
significantly decreased the ability of astrocytes to migrate into
the empty space (Fig. 2A and B).
In the control astrocytes, the wound gap was reduced at 24 h
(Fig. 2A, panel d), while the gap
was larger at 24 h in the fisetin-treated astrocytes (Fig. 2A, panels e and f). Examination of
the leading edge of the migrating cells revealed multiple
protrusions and extensions in the control astrocytes, but a
relatively uniform flat edge in the astrocytes in the fisetin
group. To gain further insight into the effects of fisetin on the
migration of primary cultured astrocytes, we examined the
expression of p-FAK and p-paxillin in vitro. The results of
western blot analysis revealed that the expression levels of both
p-FAK (Fig. 2C and D) and
p-paxillin (Fig. 2C and E) were
decreased in the fisetin-treated astrocytes. Taken together, these
results indicate that fisetin suppresses astrocyte migration.
Fisetin inhibits astrocyte proliferation
and cell cycle progression
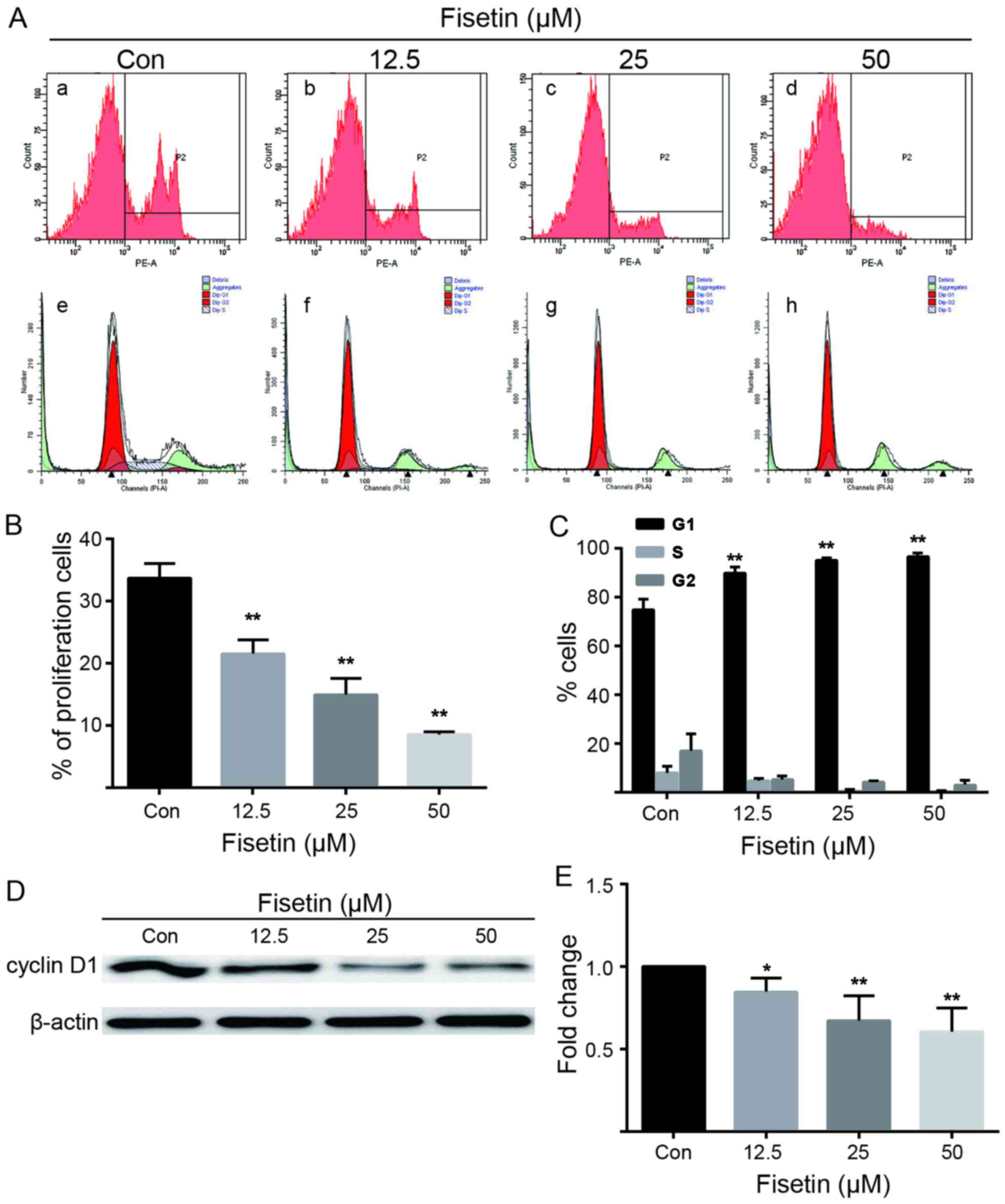
Astrocyte proliferation was detected using EdU as a
marker of dividing cells (Fig.
3A). The proportion of EdU(+) cells was markedly reduced in the
fisetin-treated group (Fig. 3A,
panels b–d) compared with the control group (Fig. 3A, panel a), in a dose-dependent
manner (Fig. 3A and B).
Similarly, fisetin increased the percentage of cells in the G1/G0
phase and decreased the percentage of cells in the G2 and S phase
(Fig. 3A, panels f–h) compared
with the control group (Fig. 3A,
panel e), as demonstrated by flow cytometry, and also in a
dose-dependent manner (Fig. 3A and
C). Accordingly, the results of western blot analysis revealed
that fisetin downregulated cyclin D1 protein expression (Fig. 3D and E). Taken together, these
data indicated that treatment with fisetin inhibited astrocyte
proliferation and arrested the cell cycle at the G1 phase.
Fisetin suppresses the Akt/Erk signaling
pathway
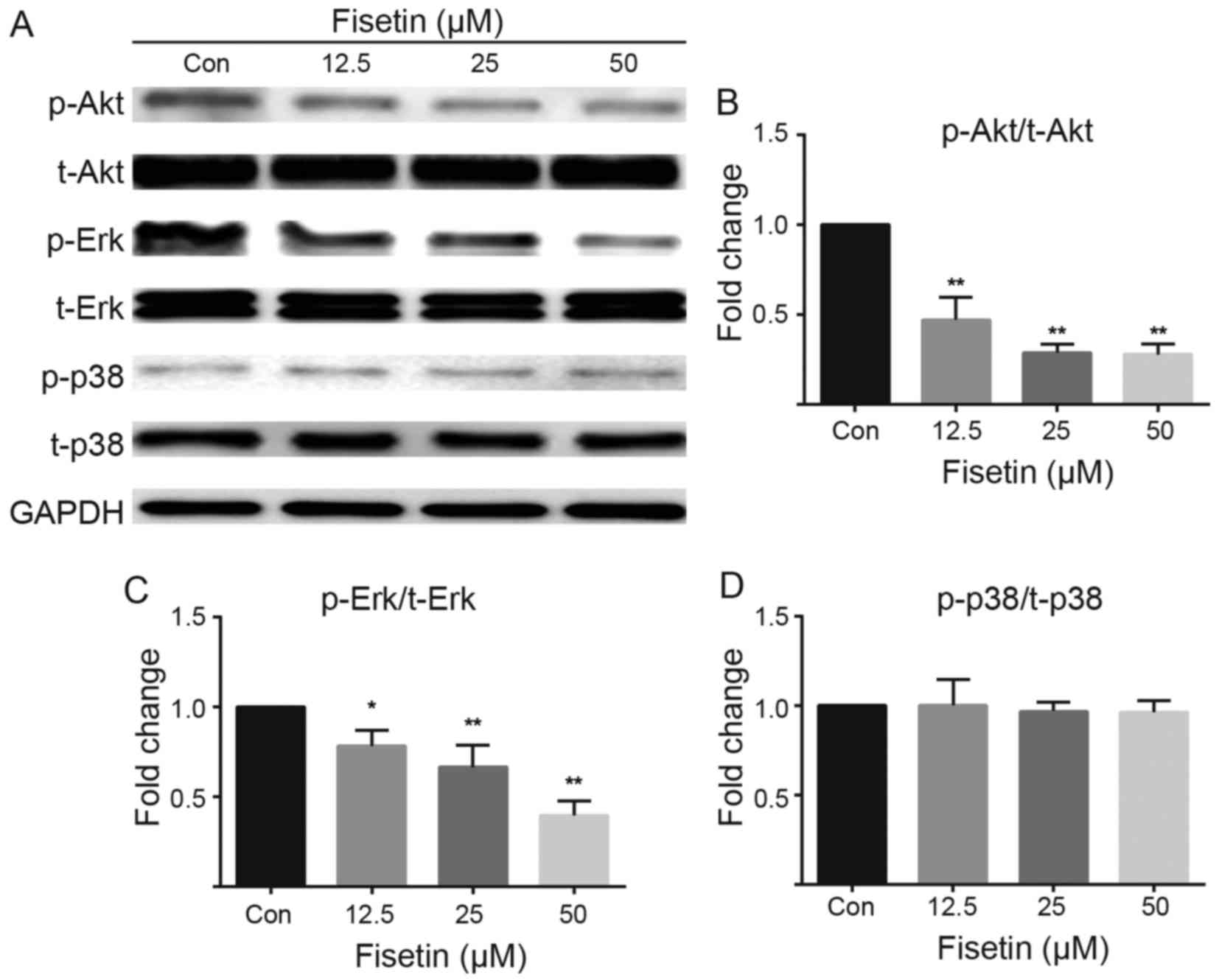
Previous studies have demonstrated that fisetin
regulates the phosphoinositide 3-kinase (PI3K)/Akt and Erk
signaling pathways (14–16). It has also been demonstrated that
astrocyte migration is modulated by Rho GTPase/p21-activated kinase
(PAK)/Erk-dependent signaling (17). In the present study, the results
of western blot analysis revealed that fisetin downregulated the
phosphorylation of Akt (Fig. 4A and
B) and Erk (Fig. 4A and C) in
a dose-dependent manner, but had no effect on total levels of Akt
and Erk. Furthermore, fisetin had no effect on the expression of
p38 MAPK or p38 phosphorylation levels compared with the control
group (Fig. 4A and D).
Discussion
Astrocytes are ubiquitous glia with finely branching
processes that envelope all the cellular components throughout the
CNS (5). Under normal
physiological conditions, astrocytes provide essential support for
neuronal activities, such as maintaining the homeostasis of the
extracellular ionic environment and pH, the clearance and release
of extracellular glutamate (18),
and provisions for metabolic substrates for neurons (19). SCI is a common and devastating CNS
insult that leads to the destruction of the cord microenvironment,
followed by limited neuronal regeneration and poor functional
recovery in adult mammals. Following severe SCI, and in response to
changes in the local microenvironment, astrocytes, the most
abundant glial cells in the CNS, transform into reactive astrocytes
(20). Reactive astrogliosis
involves cell division, migration, hypertrophy, proliferation and
changes in gene expression that contribute to glial scar formation.
Compelling evidence suggests that reactive astrogliosis has
beneficial effects, including restricting the spread of
inflammation, protecting nerves from secondary injuries, and
restoring tissue integrity in the acute phase of CNS insults
(21,22). However, studies have also
demonstrated the detrimental effects of reactive astrogliosis in
terms of interfering with the function of residual neuronal
circuits by preventing axonal remyelination and inhibiting axonal
regeneration (23,24), thus blocking functional
recovery.
Fisetin is a flavonoid and an important natural
product derived from many fruits and herbal sources. It has low
cytotoxicity and has antitumor effects in various cancer cells, in
addition to inducing cell cycle arrest (25). Although previous studies have
revealed that fisetin exerts both antioxidant and anti-inflammatory
effects in astrocytes (9,10,26), its other activities in astrocytes
are unclear.
Astrocyte migration and proliferation in SCI is a
fundamental component of glial scarring. In the present study, we
demonstrated that fisetin inhibited astrocyte migration in an in
vitro scratch-wound model and decreased FAK and paxillin
phosphorylation levels in a dose-dependent manner. FAK is a widely
expressed cytoplasmic protein tyrosine kinase involved in
integrin-mediated signal transduction. It plays an important role
in the control of several biological processes, including cell
spreading, migration and survival (27). FAK serves as a unique regulator of
focal adhesion (FA) assembly and disassembly, processes that are
fundamental for efficient directional cell movement (28,29). FAK is a leading edge organizer.
Nascent FAs are formed at the cell periphery by integrin and
extracellular matrix (ECM) interactions. Paxillin is another
important cytoskeletal and scaffolding protein recruited early to
nascent FAs at cell fronts, and is necessary for FA turnover (e.g.,
adhesion disassembly at cell fronts) during cell migration
(29,30). Our results indicated that fisetin
inhibited healing in the scratch wound model of primary astrocytes
and decreased the phosphorylation levels of FAK and paxillin, thus
indicating that is plays an essential role in the migratory ability
of astrocytes. Furthermore, emerging data indicate that fisetin
possesses potent anti-proliferative activity against various cancer
cells (25,31,32). Another study reported that fisetin
perturbed spindle checkpoint signaling, which may contribute to the
anti-proliferative effects of the compound (33). In a previous study, using a
cell-based high throughput screen for small molecules that can
override chemically-induced mitotic arrest, fisetin was identified
as an anti-mitotic compound. It was demonstrated that fisetin
rapidly compromised the microtubule drug-induced mitotic block in a
proteasome-dependent manner in several human cell lines (33). In this study, we found that
fisetin suppressed astrocyte proliferation by inhibiting DNA
synthesis, which was shown by EdU, and induced cell cycle arrest in
the G1 phase. We also examined the cell cycle-associated protein
cyclin D1 by western blot analysis. Cyclin D1 is expressed in the
G1 phase and binds to and activates cyclin-dependent kinase (CDK)4
and 6, followed by the activation of the cyclin E-dependent kinase,
CDK2.
The MAPK and Akt signaling pathways are crucial for
cellular responses including proliferation, differentiation,
survival and apoptosis. Accumulating evidence suggests an important
role for the MAPK and PI3K/Akt pathways in promoting these
biological functions; however, the findings are complex and
controversial, and depend on the specific characteristics of the
cell type (34,35). A recent study found that the
Erk1/2 signaling pathway modulated the cytoskeleton dynamics of
astrocytes in a scratch-wound model (17). Fisetin has been shown to play an
important role in regulating the PI3K/Akt and Erk pathways
(14,16,36). In the present study, fisetin
downregulated the Akt and Erk phosphorylation levels in astrocytes
in a dose-dependent manner, but had no effect on p38 expression and
phosphorylation. The p38 protein has been implicated as a key
regulator of apoptosis (37), and
these results were consistent with the apparent lack of effect of
fisetin on apoptosis.
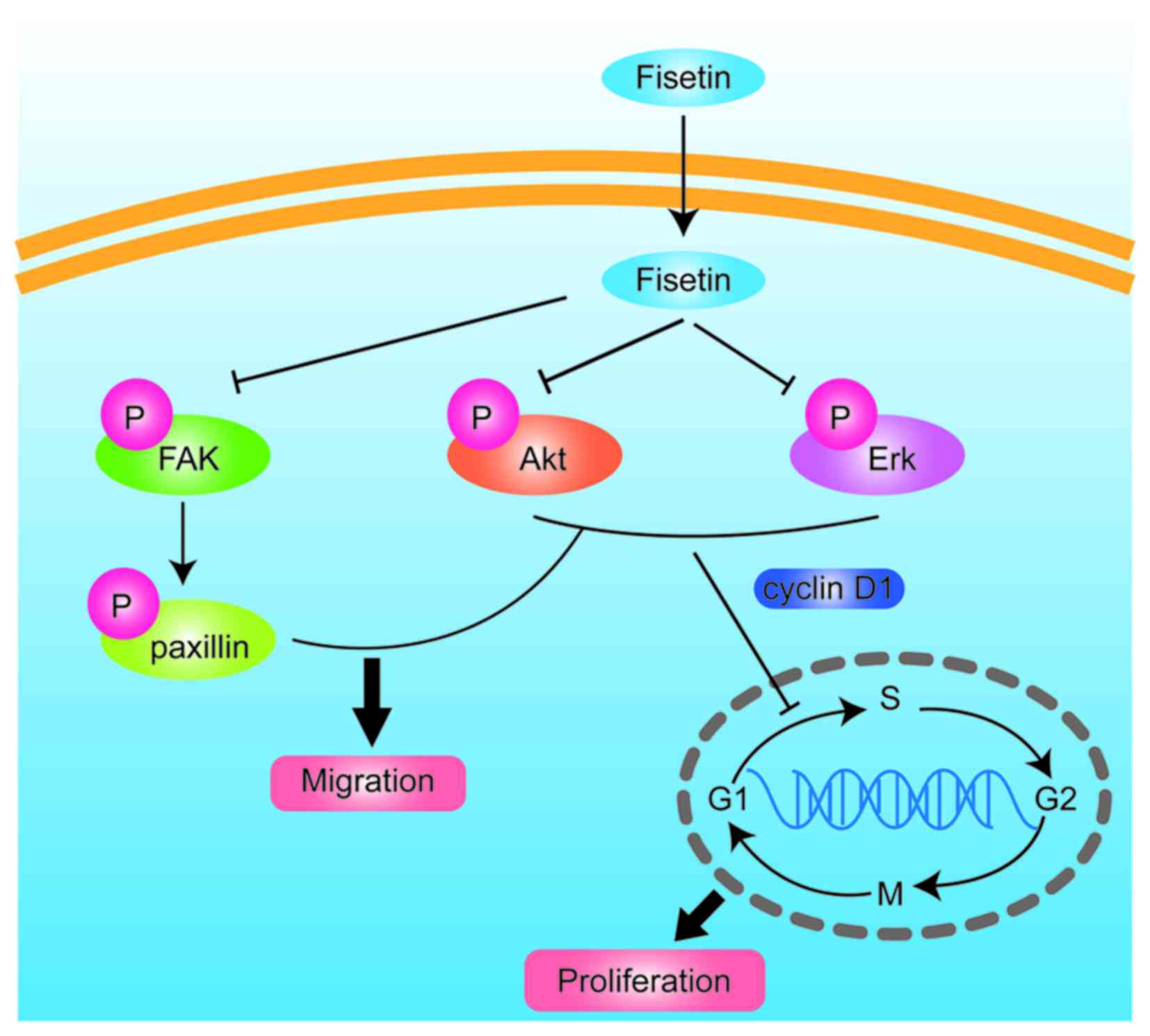
In conclusion, the findings of the present study
indicate that fisetin inhibits astrocyte migration, as shown by a
scratch-wound model and induced cell cycle arrest in the G1 phase.
Fisetin inhibited astrocyte proliferation via the PI3K/Akt- and
Erk-dependent signaling pathways, and further decreased glial scar
formation. The mechanisms involved ares shown in Fig. 5. Fisetin may therefore have
potential for use in the treatment of glial scar formation in
SCI.
Acknowledgments
The present study was supported by the Presidential
Foundation of Nan Fang Hospital, Southern Medical University, China
(grant no. 2013C003)
References
|
1
|
Yang Q, Wang EY, Huang XJ, Qu WS, Zhang L,
Xu JZ, Wang W and Tian DS: Blocking epidermal growth factor
receptor attenuates reactive astrogliosis through inhibiting cell
cycle progression and protects against ischemic brain injury in
rats. J Neurochem. 119:644–653. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Binder DK and Steinhäuser C: Functional
changes in astroglial cells in epilepsy. Glia. 54:358–368. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fuller S, Münch G and Steele M: Activated
astrocytes: a therapeutic target in Alzheimer's disease? Expert Rev
Neurother. 9:1585–1594. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Holley JE, Gveric D, Newcombe J, Cuzner ML
and Gutowski NJ: Astrocyte characterization in the multiple
sclerosis glial scar. Neuropathol Appl Neurobiol. 29:434–444. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sofroniew MV: Reactive astrocytes in
neural repair and protection. Neuroscientist. 11:400–407. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li CY, Li X, Liu SF, Qu WS, Wang W and
Tian DS: Inhibition of mTOR pathway restrains astrocyte
proliferation, migration and production of inflammatory mediators
after oxygen-glucose deprivation and reoxygenation. Neurochem Int.
83–84:9–18. 2015. View Article : Google Scholar
|
|
7
|
Wang YF, Zu JN, Li J, Chen C, Xi CY and
Yan JL: Curcumin promotes the spinal cord repair via inhibition of
glial scar formation and inflammation. Neurosci Lett. 560:51–56.
2014. View Article : Google Scholar
|
|
8
|
Adhami VM, Syed DN, Khan N and Mukhtar H:
Dietary flavonoid fisetin: a novel dual inhibitor of PI3K/Akt and
mTOR for prostate cancer management. Biochem Pharmacol.
84:1277–1281. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Prakash D, Gopinath K and Sudhandiran G:
Fisetin enhances behavioral performances and attenuates reactive
gliosis and inflammation during aluminum chloride-induced
neurotoxicity. Neuromolecular Med. 15:192–208. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chuang JY, Chang PC, Shen YC, Lin C, Tsai
CF, Chen JH, Yeh WL, Wu LH, Lin HY, Liu YS and Lu DY: Regulatory
effects of fisetin on microglial activation. Molecules.
19:8820–8839. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu BY and Yu AC: Quercetin inhibits c-fos,
heat shock protein, and glial fibrillary acidic protein expression
in injured astrocytes. J Neurosci Res. 62:730–736. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yuan Z, Yao F, Hu Z, Sun S and Wu B:
Quercetin inhibits the migration and proliferation of astrocytes in
wound healing. Neuroreport. 26:387–393. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cory G: Scratch-wound assay. Methods Mol
Biol. 769:25–30. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen PY, Ho YR, Wu MJ, Huang SP, Chen PK,
Tai MH, Ho CT and Yen JH: Cytoprotective effects of fisetin against
hypoxia-induced cell death in PC12 cells. Food Funct. 6:287–296.
2015. View Article : Google Scholar
|
|
15
|
Maher P, Dargusch R, Bodai L, Gerard PE,
Purcell JM and Marsh JL: ERK activation by the polyphenols fisetin
and resveratrol provides neuroprotection in multiple models of
Huntington's disease. Hum Mol Genet. 20:261–270. 2011. View Article : Google Scholar
|
|
16
|
Chiang HM, Chan SY, Chu Y and Wen KC:
Fisetin ameliorated photodamage by suppressing the
mitogen-activated protein kinase/matrix metalloproteinase pathway
and nuclear factor-κB pathways. J Agric Food Chem. 63:4551–4560.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lichtenstein MP, Carriba P, Baltrons MA,
Wojciak-Stothard B, Peterson JR, García A and Galea E:
Secretase-independent and RhoGTPase/PAK/ERK-dependent regulation of
cytoskeleton dynamics in astrocytes by NSAIDs and derivatives. J
Alzheimers Dis. 22:1135–1155. 2010.PubMed/NCBI
|
|
18
|
Mazzanti M, Sul JY and Haydon PG:
Glutamate on demand: astrocytes as a ready source. Neuroscientist.
7:396–405. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pellerin L and Magistretti PJ:
Neuroenergetics: calling upon astrocytes to satisfy hungry neurons.
Neuroscientist. 10:53–62. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yuan YM and He C: The glial scar in spinal
cord injury and repair. Neurosci Bull. 29:421–435. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rolls A, Shechter R and Schwartz M: The
bright side of the glial scar in CNS repair. Nat Rev Neurosci.
10:235–241. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Silver J and Miller JH: Regeneration
beyond the glial scar. Nat Rev Neurosci. 5:146–156. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sofroniew MV: Molecular dissection of
reactive astrogliosis and glial scar formation. Trends Neurosci.
32:638–647. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang CX and Shuaib A: Involvement of
inflammatory cytokines in central nervous system injury. Prog
Neurobiol. 67:161–172. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen CM, Hsieh YH, Hwang JM, Jan HJ, Hsieh
SC, Lin SH and Lai CY: Fisetin suppresses ADAM9 expression and
inhibits invasion of glioma cancer cells through increased
phosphorylation of ERK1/2. Tumour Biol. 36:3407–3415. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Currais A, Prior M, Dargusch R, Armando A,
Ehren J, Schubert D, Quehenberger O and Maher P: Modulation of P25
and inflammatory pathways by fisetin maintains cognitive function
in Alzheimer's disease transgenic mice. Aging Cell. 13:379–390.
2014. View Article : Google Scholar
|
|
27
|
Schaller MD, Hildebrand JD, Shannon JD,
Fox JW, Vines RR and Parsons JT: Autophosphorylation of the focal
adhesion kinase, pp125FAK, directs SH2-dependent binding of P60s
rc. Mol Cell Biol. 14:1680–1688. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lim Y, Lim ST, Tomar A, Gardel M,
Bernard-Trifilo JA, Chen XL, Uryu SA, Canete-Soler R, Zhai J, Lin
H, et al: PyK2 and FAK connections to p190Rho guanine nucleotide
exchange factor regulate RhoA activity, focal adhesion formation,
and cell motility. J Cell Biol. 180:187–203. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hu YL, Lu S, Szeto KW, Sun J, Wang Y,
Lasheras JC and Chien S: FAK and paxillin dynamics at focal
adhesions in the protrusions of migrating cells. Sci Rep.
4:60242014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Deakin NO and Turner CE: Paxillin comes of
age. J Cell Sci. 121:2435–2444. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Syed DN, Suh Y, Afaq F and Mukhtar H:
Dietary agents for chemoprevention of prostate cancer. Cancer Lett.
265:167–176. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Khan N, Syed DN, Ahmad N and Mukhtar H:
Fisetin: a dietary antioxidant for health promotion. Antioxid Redox
Signal. 19:151–162. 2013. View Article : Google Scholar :
|
|
33
|
Salmela AL, Pouwels J, Varis A, Kukkonen
AM, Toivonen P, Halonen PK, Perälä M, Kallioniemi O, Gorbsky GJ and
Kallio MJ: Dietary flavonoid fisetin induces a forced exit from
mitosis by targeting the mitotic spindle checkpoint.
Carcinogenesis. 30:1032–1040. 2009. View Article : Google Scholar :
|
|
34
|
Kim EK and Choi EJ: Pathological roles of
MAPK signaling pathways in human diseases. Biochim Biophys Acta.
1802:396–405. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Engelman JA: Targeting PI3K signalling in
cancer: opportunities, challenges and limitations. Nat Rev Cancer.
9:550–562. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Barbakadze T, Natsvlishvili N and
Mikeladze D: Thyroid hormones differentially regulate
phosphorylation of ERK and Akt via integrin αvβ3 receptor in
undifferentiated and differentiated PC-12 cells. Cell Biochem
Funct. 32:282–286. 2014. View
Article : Google Scholar
|
|
37
|
Liu Y, Zhang S, Su D, Liu J, Cheng Y, Zou
L, Li W and Jiang Y: Inhibiting (pro)renin receptor-mediated P38
MAPK signaling decreases hypoxia/reoxygenation-induced apoptosis in
H9c2 cells. Mol Cell Biochem. 403:267–276. 2015. View Article : Google Scholar : PubMed/NCBI
|