Introduction
Stem cell transplantation has emerged as a
promising, alternative treatment for ischemic heart disease. Of the
available stem cells, mesenchymal stem cells (MSCs) hae been widely
studied and are considered as a leading candidate with many
advantages, such as the capacity for self-renewal, a multilineage
differentiation potential, low immunogenicity, immunosuppressive
properties and low tumorigenicity (1,2).
Transdifferentiation and paracrine signaling are considered to be
the principal mechanisms of the effects of MSCs in the treatment of
ischemic heart disease (3).
It is evident that the first, vital step of cell
therapy is to enable the stem cells to reach the targeted ischemic
location and to successfully promote survival, which could be
partly responsible for the modest benefits of stem cell therapy
observed in a clinical trial setting (4). Undoubtedly, the improvement of the
engraftment and survival of the transplanted stem cells is vital.
Many efforts have been made in that regard, mainly focusing on the
improvement of the microenvironment and the enhancement of cellular
resistance to a hostile environment via pre-treatment with growth
factors or cytokines, hypoxic preconditioning, and genetic
modification with anti-apoptotic, angiogenetic and other genes
(5). An improved therapeutic
efficacy has been observed in animals with transplanted genetically
modified MSCs compared to animals that have been transplanted with
untreated MSCs. This process is known as cell-based gene therapy
(6). Recent data have
demonstrated that the modification of the angiogenetic gene
vascular endothelial growth factor (VEGF) in MSCs exerts a better
angiogenetic and paracrine effect (7), while the modification of the
anti-apoptotic gene, B-cell lymphoma-2 (Bcl-2), confers improved
survival (8). Importantly,
compared to the untreated MSCs, VEGF- and Bcl-2-modified cells have
been shown to improve cardiac function. Moreover, an interaction
which could promote the mutual expression of VEGF and Bcl-2 has
been observed in several studies (8–10).
Given the information described above, we
hypothesized that the combined overexpression of VEGF and Bcl-2
could confer much greater resistance to the MSCs in an adverse
environment.
Materials and methods
Recombinant lentiviral vector
construction
In order to confer stable overexpression, a
lentiviral vector was used for gene delivery (11,12). The plasmid,
pLV-CMV-EF1-fLuc-T2A-puro, purchased from HanBio Technology Co.,
Ltd. (Shanghai, China) was used to construct the recombinant
lentiviral vectors. The human VEGF gene (NM_001171626.1) and human
Bcl-2 gene (NM_000633) were polymerase chain reaction
(PCR)-amplified from a complementary deoxyribonucleic acid (cDNA)
library. A self-cleaving T2A peptide sequence was employed to link
and achieve the co-overexpression of VEGF and Bcl-2 (13). A Bcl-2-T2A-VEGF fragment was
obtained by overlapping PCR. PCR primers were designed and the
EcoR1 and Xho1 restriction endonuclease sites were
introduced into the plasmid as follows: VEGF,
5′-ACACGAATTCATGAACTTTCTGCTGTCTTGGGT-3′ (forward) and
5′-ATTGCTCGAGTCACCGCCTCGGCTTGTCACATC-3′ (reverse); Bcl-2,
5′-GAATTCATGGCGCACGCTGGGAGAACAGG-3′ (forward) and
5′-ATTGCTCGAGTCACTTGTGGCCCAGATAGGCACC-3′ (reverse); Bcl-2-T2A-VEGF,
5′-GAATTCATGGCGCACGCTGGGAGAACAGG-3′ (forward); Fusion-R1,
5′-CACGTCACCGCATGTTAGAAGACTTCCTCTGCCCTCCTTGTGGCCCAGATAG-3′ and
Fusion-F2,
5′-ACATGCGGTGACGTGGAGGAGAATCCCGGCCCTATGAACTTTCTGCTGTCTTG-3′ and
5′-TTGCTCGAGTTCACCGCCTCGGCTTGTCACATC-3′ (forward). Following the
addition of 10 μl of the PCR products onto a 1% agarose gel
with ethidium bromide (0.5 mg/ml) and electrophoresis, images were
captured by ultraviolet transillumination. The plasmid was doubly
digested with EcoR1 and Xho1 (NEB, Ipswich, MA, USA).
The PCR products and plasmid were purified, and they were then
ligated and the ligation mixture was transformed into competent
Escherichia coli DH5α cells (Invitrogen, Carlsbad, CA, USA).
Clones were selected for PCR validation, and the recombinant
plasmid was extracted for sequencing. The lentivirus packaging
system consists of 3 plasmids: pLV-CMV-EF1-fLuc-T2A-puro, pSPAX2
and pMD2G (HanBio Technology Co., Ltd.). These 3 plasmids were
co-transfected into the 293T cell line (HanBio Technology Co.,
Ltd.) according to the instructions of the manufacturer of
Lipofectamine 2000 (Invitrogen) after the endotoxin was removed.
The cells were transferred to a complete medium after 8 h of
co-transfection. The supernatant was harvested and concentrated
after 48 h of incubation. The viral titer was determined and
calibrated in the 293T cell line. The 293T cell culture medium was
then centri fuged and concentrated to obtain a lentivirus solution,
which was kept in a collection cup at −80°C. The 3 lentiviral
vectors were named Lv-VEGF, Lv-Bcl-2 and Lv-BV. The control
lentiviral vector encoding green fluorescent protein (GFP) was
purchased from HanBio Technology Co., Ltd.
MSC preparation, cell transfection and
the generation of cell lines with stable expression
OriCell™ Sprague-Dawley (SD) rat MSCs, which had
been primarily isolated, cultured and passaged no more than twice,
were purchased from Cyagen Biosciences Inc. (Santa Clara, CA, USA).
The MSCs were cultured according to the manufacturer's
instructions. Briefly, the MSCs were seeded in cell culture flasks
(Corning Inc., Corning, NY, USA) in complete medium containing low
glucose-Dulbecco's modified Eagle's medium (DMEM), 10% fetal bovine
serum (both from Gibco, Grand Island, NY, USA), 100 U/ml penicillin
and 100 μg/ml streptomycin sulphate (Gibco) at 37°C in a
humidified atmosphere in a 5% CO2 and 95% air incubator
(Thermo Fisher Scientific, San Jose, CA, USA). When 80% confluence
was reached, the cells were detached using 0.25% trypsin-EDTA
(Gibco) and subcultured at a 1:2 ratio. The 4th passage MSCs were
seeded into 6-well plates (Coster, Cambridge, MA, USA) at
1×104 cells/cm2. Lv-GFP was used as a
control. When the cells were 50% confluent, they were infected with
Lv-GFP at a multiplicity of infection (MOI) of 10, 20, 50,100 or
200. GFP expression was observed under a fluorescence microscope
(Leica, Wetzlar, Germany) after 48 h. An MOI=100 was selected for
subsequent experiments, which was adequate for sufficient genetic
overexpression in the infected cells with minimum damage. The 4
lentiviruses Lv-GFP, Lv-VEGF, Lv-Bcl-2 and Lv-BV were each added
separately into the culture medium at an MOI of 100. Transfections
were performed for 24 h at 37°C, and the virus-containing medium
was then replaced with fresh complete medium and the cells were
incubated for a further 48 h. Stably transfected cells were
selected by the addition of 2 μg/ml puromycin
(Sigma-Aldrich, St. Louis, MO, USA) to the medium, which was
changed every 2 days. Selection was terminated when the uninfected
control cells were completely dead. The 4 stably expressing cell
lines were designated as MSC-GFP, MSC-VEGF, MSC-Bcl-2 and
MSC-BV.
Cell proliferation assay
The proliferation of stably expressing cell lines
under normal culture conditions was assessed using a Cell Counting
kit-8 (CCK-8; Dojindo Laboratories, Kumamoto, Japan) according to
the manufacturer's instructions. For culturing, the cells were
plated in 96-well plates (Coster) at 5×103 cells/well.
CCK-8 solution (10 μl) was added to each well followed by
incubation at 37°C for 1 h. The optical density (OD) at 450 nm was
measured using a Synergy 2 multi-mode microplate reader (BioTek,
Winooski, VT, USA) at 7 days.
Reverse transcription-quantitative
(real-time) PCR (RT-qPCR)
RT-qPCR was performed to analyze the messenger
ribonucleic acid (mRNA) expression levels of VEGF and Bcl-2. TRIzol
reagent (Invitrogen) was used to extract the RNA from the MSCs and
then reverse transcribed into cDNA using the GoScript reverse
transcription system (Promega, Madison, WI, USA) following the
manufacturer's instructions. FastStart Universal SYBR-Green Master
(Rox) (Roche Applied Science, Mannheim, Germany) was used for
quantitative (real-time) PCR analysis. The primer sequences used in
this experiment were: VEGF forward, 5′-GCACTGGACCCTGGCTTTACT-3 and
reverse, 5′-ATGGGACTTCTGCTCTCCTTCTG-3′; Bcl-2 forward,
5′-GGGAGATCGTGATGAAGTACATACA-3′ and reverse,
5′-GCACAGCGGGCATTGGGTTG-3′; glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) forward, 5′-TCCACTCACGGCAAATTCAAC-3′ and
reverse, 5′-GTAG ACTCCACGACATACTCAGC-3′. The PCR-amplified mRNA was
quantified and the results were normalized against GAPDH
expression. The 2−ΔΔCq method was used to calculate the
relative mRNA expression level.
Western blot analysis
A western blot assay was performed. Briefly, the
cell lysate supernatants were collected. A Bradford assay was used
to measure the protein concentration. Protein samples (20–25
μg) were separated by sodium dodecyl sulfate polyacrylamide
gel electrophoresis (Boster, Wuhan, China) and transferred onto
polyvinylidene difluoride membranes (Millipore, Milford, MA USA).
The membranes were probed overnight with primary antibodies as
follows: mouse anti-VEGF (1:200 dilution, cat. no. Ab1316; Abcam,
Cambridge, UK), rabbit anti-Bcl-2 (1:1,000 dilution, cat. no. 2870;
Cell Signaling Technology, Danvers, MA, USA), rabbit anti-GAPDH
(1:1,000 dilution, cat. no. 2118; Cell Signaling Technology), mouse
anti-Bax (1:1,000 dilution, cat. no. sc-7480; Santa Cruz
Biotechnology, Santa Cruz, CA, USA), rabbit anti-cleaved-caspase-3
(1:1,000 dilution, cat. no. 9661), rabbit anti-LC3 (1:1,000
dilution, cat. no. 2775), rabbit anti-Beclin-1 (1:1,000, cat. no.
3495) and rabbit ant-p62 (1:1,000 dilution, cat. no. 8025, all from
Cell Signaling Technology). Diluted secondary antibodies [HRP goat
anti-mouse IgG (1:40,000 dilution, cat. no. 115-035-003) and HRP
goat anti-rabbit IgG (1:10,000 dilution, cat. no. 111-035-003; both
from Jackson ImmunoResearch Laboratory Inc., West Groove, PA, USA)]
were used to detect the corresponding primary antibodies. Further
analysis was carried out using ImageJ software (NIH, Bethesda, MD,
USA) to quantify the protein bands.
In vitro model of oxygen glucose
deprivation (OGD)
To mimic the ischemic microenvironment that the
transplanted cells encounter in vivo, the MSCs were exposed
to OGD conditions at 37°C, 95% N2, 5% CO2 in
glucose-free DMEM medium (Gibco). Four MSC lines of the same
passage were seeded into individual 6-well plates at a density of
5×104/cm2. Following 24 h of culture, the
cells were washed 3 times with phosphate-buffered saline (PBS)
gently and were then cultured under OGD conditions for 12 h. A
plate of MSC-GFP was simultaneously cultured under normal
conditions as a negative control (MSC-NC).
Flow cytometric analysis of
apoptosis
A cell apoptosis assay was performed using an
Annexin V-FITC/propidium iodide (PI) apoptosis detection kit
(Invitrogen) according to the manufacturer's instructions. Briefly,
the cells were collected and washed twice with PBS and then
suspended in 300 μl of binding buffer. Annexin V solution (5
μl) was added to the cell suspension and incubated for 15
min in the dark at room temperature. Subsequently, 200 μl of
blinding buffer and 5 μl of PI were added and the cell
suspension was immediately analyzed on a flow cytometer (Accuri™
C6; BD Biosciences, San Jose, CA, USA). Annexin
V−/PI− were viable cells, Annexin
V+/PI− cells were early apoptotic cells, and
Annexin V+/PI+ cells were late apoptotic or
dead cells.
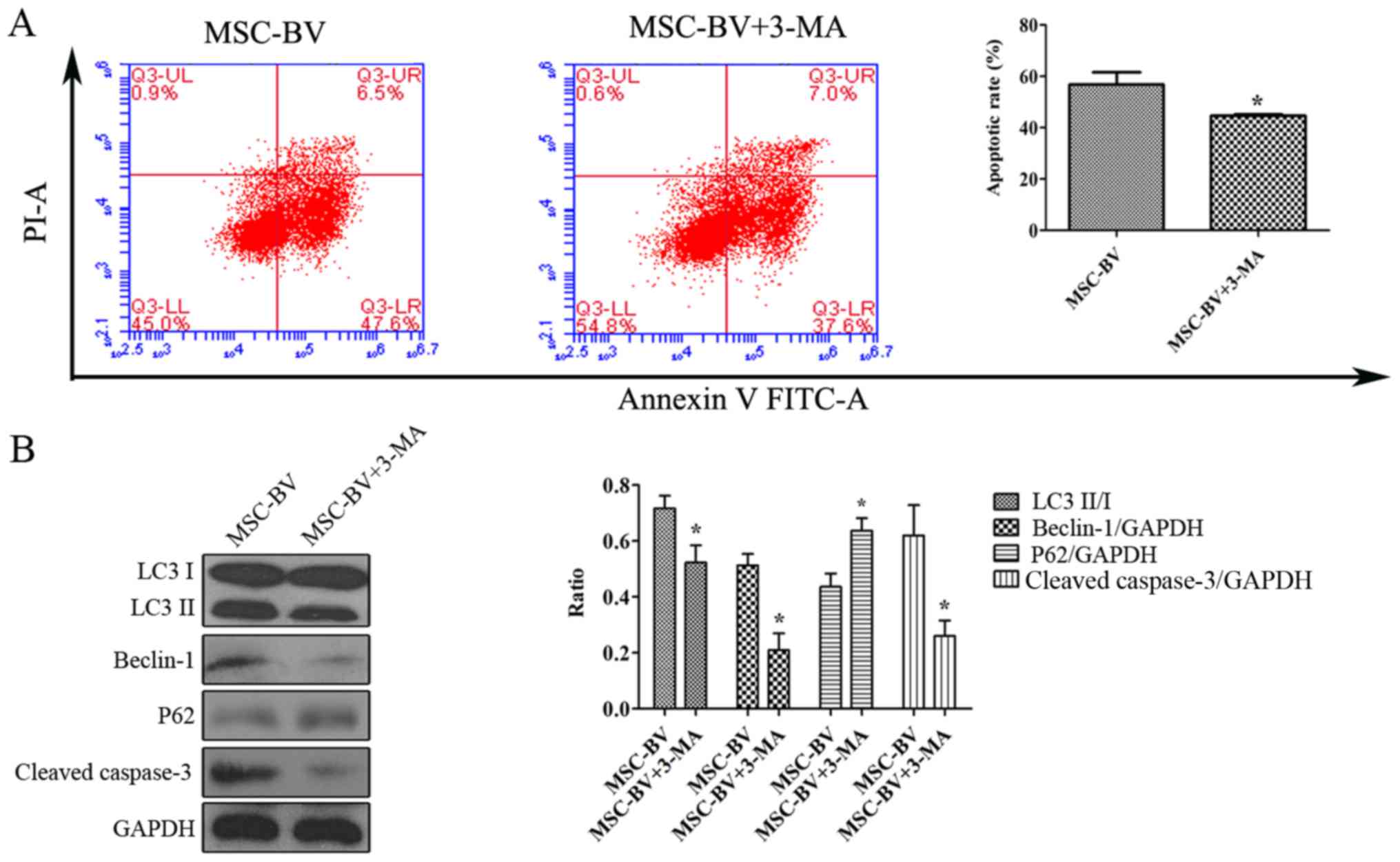
Suppression of autophagy in the MSC-BV
group under OGD conditions
The cells in the MSC-BV group were exposed to 12 h
of OGD conditions in the absence or presence of 5 mmol/l of
3-methyladenine (3-MA; Sigma-Aldrich), an autophagy inhibitor. Flow
cytometric analysis and western blot analysis for LC3, Beclin-1,
p62, cleaved-caspase-3 and GAPDH was carried out to assess the
effects of the suppression of autophagy on the apoptosis of the
cells in the MSC-BV group under OGD conditions.
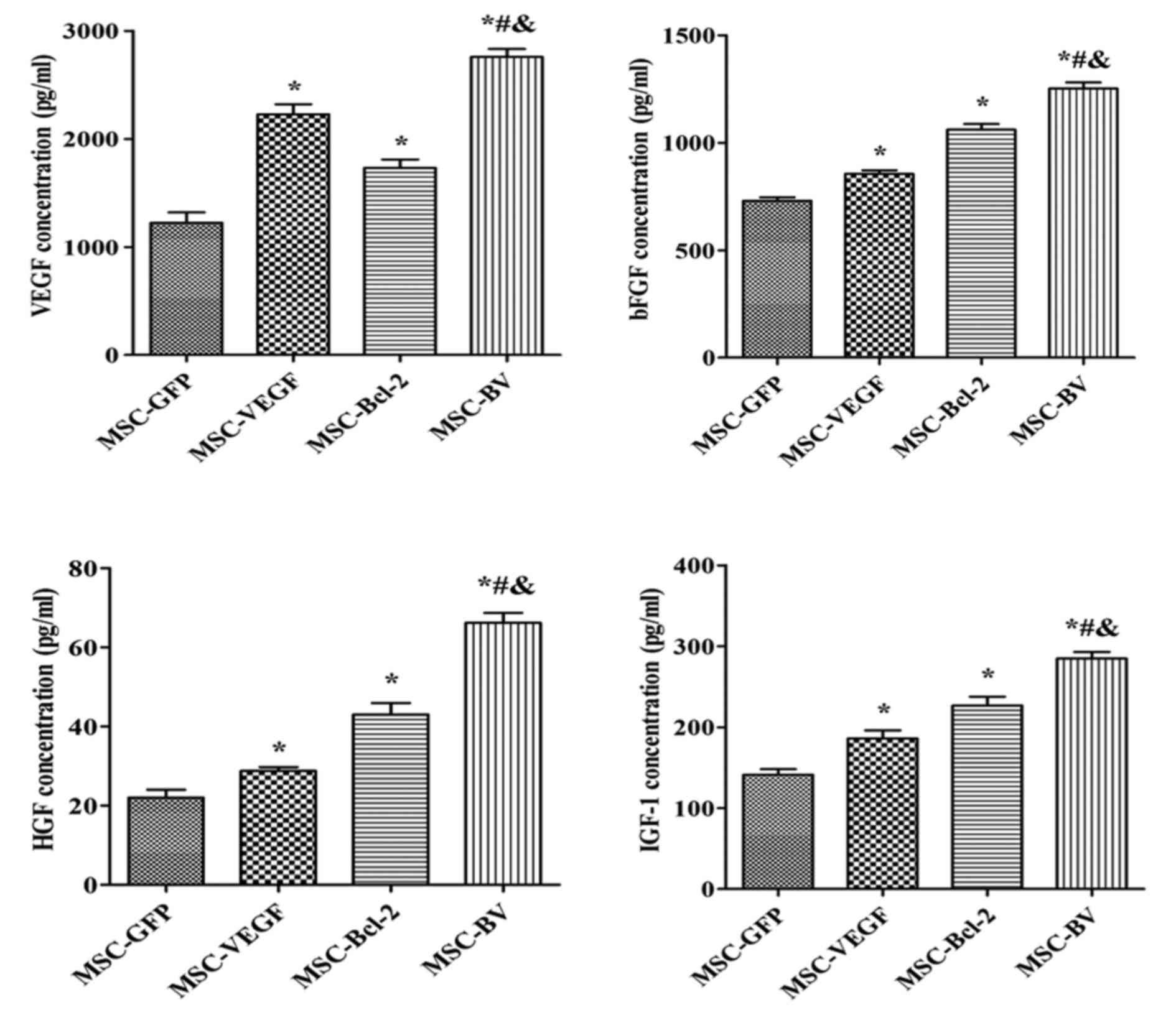
Enzyme-linked immunosorbent assay
(ELISA)
To determine whether the genetic modification of
MCSs affects paracrine signaling following culturing under OGD
conditions, an ELISA was performed to detect the concentration of
VEGF, basic fibroblast growth factor (bFGF), hepatocyte growth
factor (HGF) and insulin-like growth factor-1 (IGF-1) in the
culture supernatant using the appropriate ELISA kits (RapidBio,
Winnetka, CA, USA) according to the manufacturer's instructions.
Supernatant samples were collected from the 4 treatment groups. The
OD value at 450 nm was detected using an ELISA reader (BioTek).
Statistical analysis
The data are expressed as the means ± standard
deviation (SD). Statistical analysis was performed using SPSS for
Windows version 20.0 (SPSS, Inc., Chicago, IL, USA). Comparisons
between more than 2 groups were performed by a one-way analysis of
variance (ANOVA), followed by a post hoc Tukey test. Comparisons
between 2 groups were performed using the Student's t-text. A value
of P<0.05 was considered to indicate a statistically significant
difference.
Results
Recombinant lentiviral vector
construction
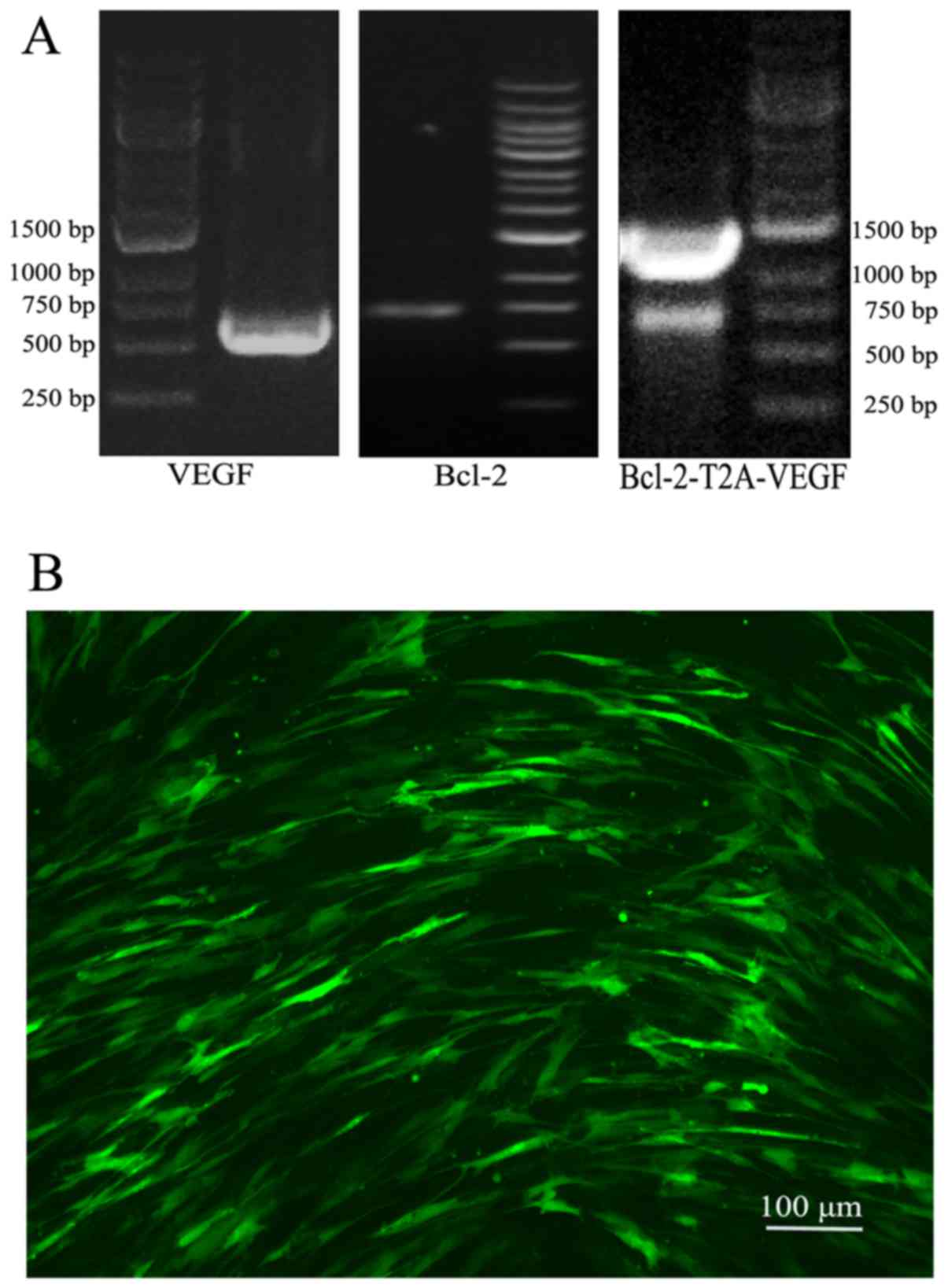
Following PCR amplification, agarose gel
electrophoresis revealed that the target gene fragments were in the
expected positions (Fig. 1A).
Sequencing confirmed that the gene fragments were correctly
subcloned into the vector (data not shown). The lentiviral vectors
were successfully constructed, and the viral titer was
1×108 TU/ml after packaging.
The 4 cell lines with stable gene
expression, target gene expression and cell proliferation
The infection rate of Lv-GFP in the MSCs at an MOI
of 100 was approximately 70% (Fig.
1B). Following 3 rounds of puromycin addition, stable cell
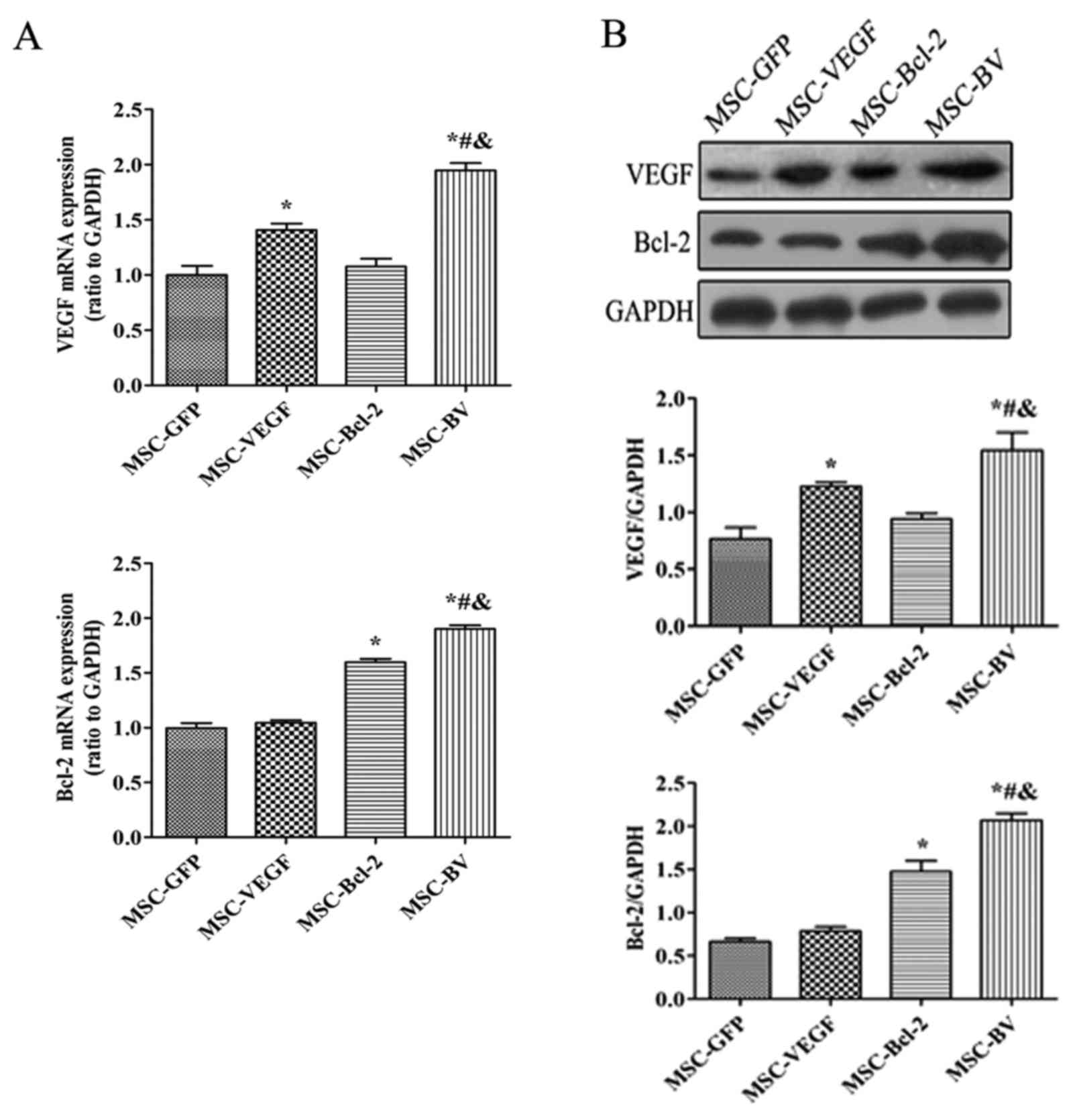
lines that expressed the target genes were selected. RT-qPCR
indicated that the mRNA expression levels of both VEGF and Bcl-2 in
the MSC-BV group were markedly higher than those in the other cell
groups (Fig. 2A). Western blot
analysis further confirmed the highest protein expression of both
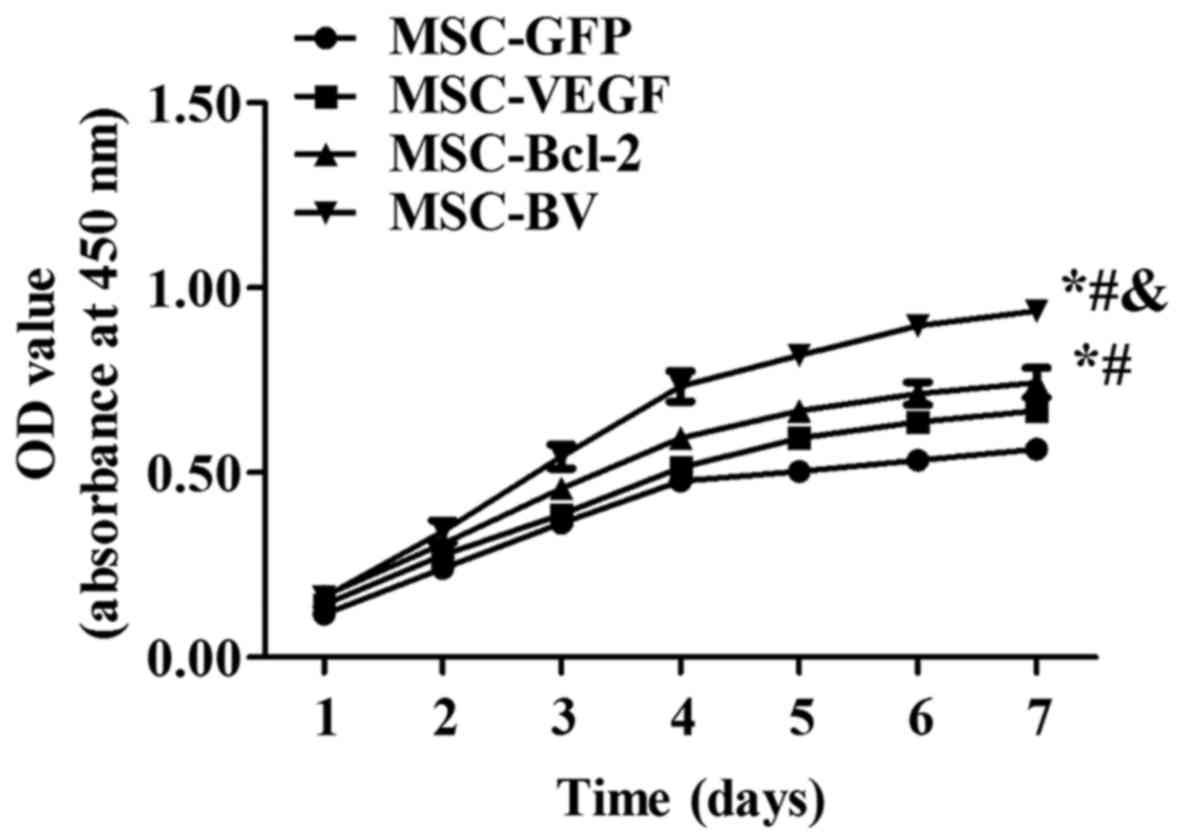
VEGF and Bcl-2 in the MSC-BV group (Fig. 2B). A CCK-8 assay revealed that the
cells in the MSC-BV group proliferated more rapidly than the 3
other cell lines over a period of 7 days under normal culture
conditions (Fig. 3).
Co-overexpression of VEGF and Bcl-2
inhibits the OGD-induced apoptosis of MSCs
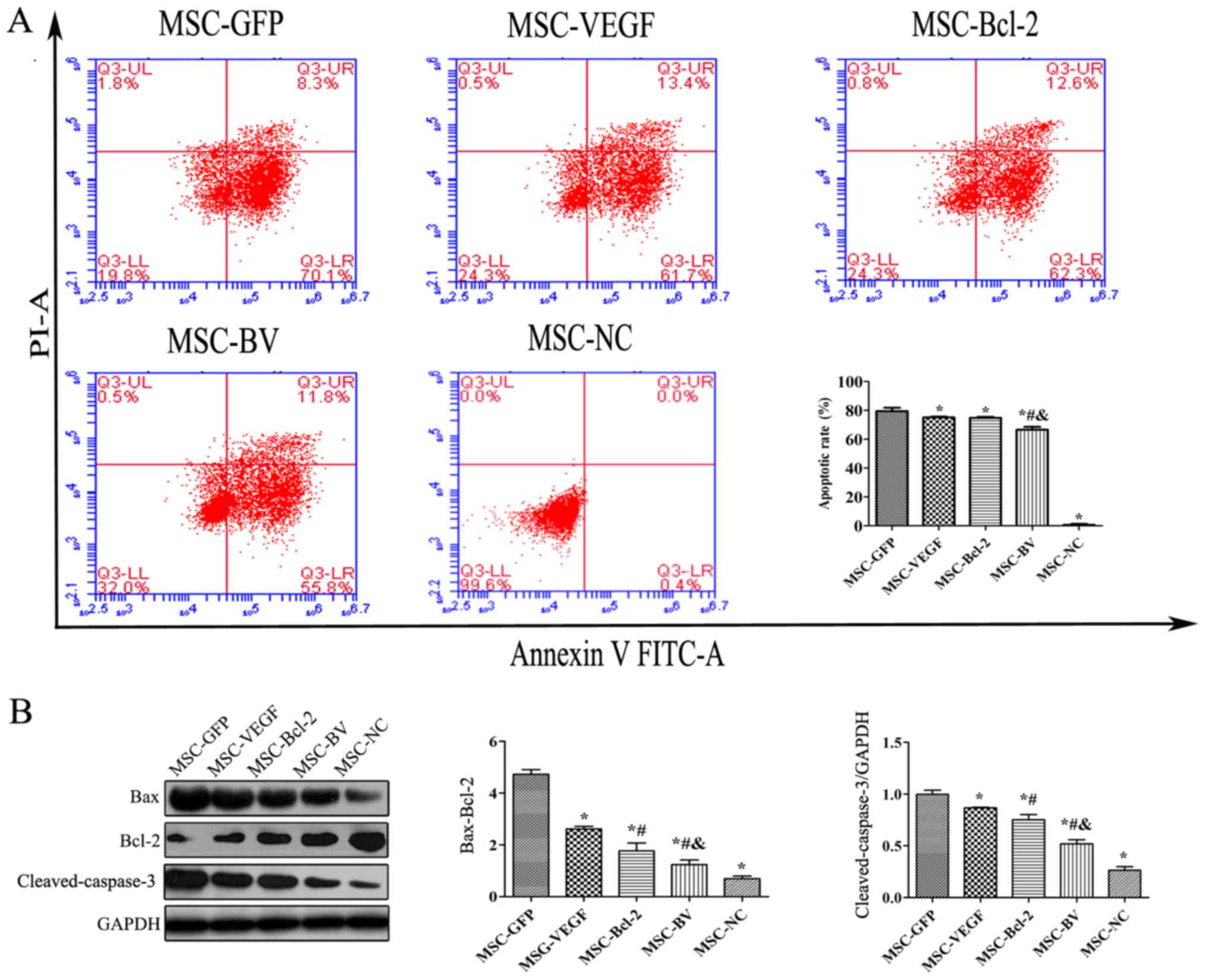
To examine the anti-apoptotic activity of MSCs
co-overexpressing VEGF and Bcl-2 under an adverse environment, the
cells in the MSC-GFP, MSC-VEGF, MSC-Bcl-2 and MSC-BV groups were
individually cultured under OGD conditions for 12 h. An Annexin
V-FITC/PI flow cytometric assay revealed that the apoptotic rate of
the cells in the MSC-BV group was significantly lower than that of
the 3 other cell groups (Fig.
4A). Western blot results also revealed that the lowest
Bax/Bcl-2 ratio and cleaved-caspase-3 protein expression were
observed in the cells in the MSC-BV group (Fig. 4B).
Co-overexpression of VEGF and Bcl-2
suppresses the OGD-induced autophagy of MSCs
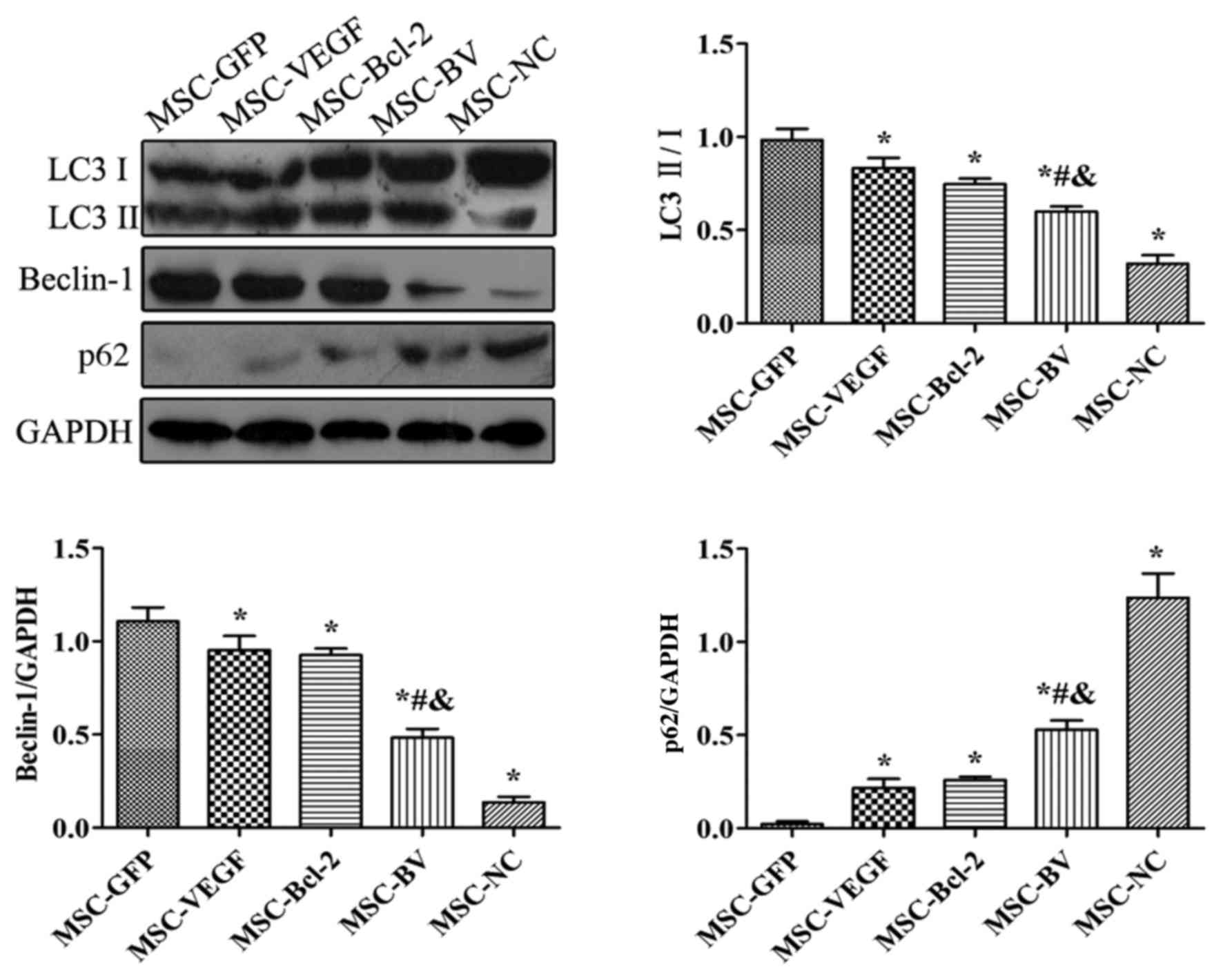
The results of western blot analysis revealed that
following exposure to OGD, the levels of LC3II/I and Beclin-1 in
the MSC-GFP group were markedly increased compared to the MSC-NC
group. However, their expression levels in the MSC-VEGF and
MSC-Bcl-2 groups were relatively lower than those in the MSC-GFP
group, while the expression levels were lowest in the MSC-BV group.
The expression of p62 in the MSC-GFP group was markedly decreased
compared to the level in the MSC-NC group. However, p62 expression
in the MSC-VEGF and MSC-Bcl-2 groups was relatively higher than
that in the MSC-GFP group, while the expression was the highest in
the MSC-BV group (Fig. 5).
Suppression of autophagy contributes to
the inhibition of the apoptosis of MSCs under OGD conditions
The Annexin V-FITC/PI flow cytometric assay revealed
that following exposure to OGD, the cells in the MSC-BV + 3-MA
group had a lower apoptotic rate than the cells in the MSC-BV group
(Fig. 6A). Western blot analysis
revealed that the levels of LC3II/I, Beclin-1 and cleaved-caspase-3
in the MSC-BV + 3-MA group were markedly decreased compared to
those in the MSC-BV group, while the expression of p62 was
increased (Fig. 6B).
Co-overexpression of VEGF and Bcl-2
enhances the paracrine effects of MSCs under OGD conditions
An ELISA was used to detect the concentration of
VEGF, bFGF, HGF and IGF-1. Consistent trends in the levels of these
growth factors were observed. The cells in the MSC-BV group had the
highest level of growth factors out of all the groups (Fig. 7).
Discussion
In this study, we first constructed a lentiviral
vector using a self-cleaving T2A peptide sequence to link and
achieve the co-overexpression of the VEGF and Bcl-2 genes, and we
then transfected the MSCs and generated cell lines that stably
overexpressed VEGF and Bcl-2. We subsequently confirmed that the
VEGF and Bcl-2 dual genetically modified MSCs had higher mRNA and
protein expression levels and a more rapid proliferation than the
MSCs in which single genes had been modified. Moreover, our study
also demonstrated that the dual genetic modification of VEGF and
Bcl-2 in the MSCs led to a more pronounced and additive
self-protection effect than the modification of single genes in
MSCs in an in vitro ischemic model (OGD), which was
indicated by the inhibition of apoptosis, the suppression of
autophagy and the enhancement of the paracrine effects. We even
found that the suppression of autophagy may have contributed to the
inhibition of apoptosis in MSCs under OGD conditions. To the best
of our knowledge, this is the first study that combines the
simultaneous modification of the angiogenic VEGF gene and the
anti-apoptotic Bcl-2 gene in MSCs. We confirmed that this combined
strategy endowed the MSCs with a better capacity to confront an
adverse environment than the modification of single genes, and an
even stronger paracrine effect.
Stem cell transplantation has been widely studied
and is considered a promising therapy for ischemic heart disease.
Although our results were encouraging, many preclinical and
clinical studies have shown that the poor survival of transplanted
cells can limit their efficacy. Therefore, measures aimed at
enhancing the tolerance of MSCs to a hostile environment are vital
for cell therapy, and gene modification may represent a potential
strategy. It has been demonstrated that the overexpression of
anti-apoptotic genes, such as Bcl-2, Akt, Hsp-27, Hsp-20 and
survivin is relatively effective in improving the survival of MSCs
in an adverse environment, improving cardiac function (8,14–17). VEGF has been used as a potent
therapeutic reagent in the treatment of ischemia via the induction
of angiogenesis in MSCs. VEGF-modified MSCs have been shown to
possess an enhanced repair capacity due to the induction of
angiogenesis and an enhanced paracrine effect (7,18–21). Of note, the expression of VEGF in
MSCs has been shown to be upregulated by Bcl-2 modification under
hypoxic conditions (8). In
addition, a similar regulation of Bcl-2 expression by VEGF in
hypoxic cancer cells has been observed (9). The involvement of the MAPK signaling
pathways may be partly responsible for the interaction VEGF and
Bcl-2 (10). Based on this
information, it is reasonable to hypothesize that combining the
VEGF and Bcl-2 genes in MSCs may enhance survival, and improve
angiogenesis and enhance the paracrine effects. In this study, our
results confirmed this hypothesis. Under normal culture conditions,
dual genetically modified MSCs had a higher overexpression level of
VEGF and Bcl-2. Under OGD conditions, dual genetically modified
MSCs exhibited higher survival rate and stronger paracrine
effect.
A previous study even confirmed that the strategy of
combining an anti-apoptotic and an angiogenetic gene in MSCs was
feasible (22). In that study,
the authors achieved the co-overexpression of Akt and Ang-1 using
two adenoviral vectors, which confirmed that the co-overexpression
led to better cytoprotection in the in vitro OGD model, and
showed that dually genetically modified MSCs transplanted into
animals with myocardial infarction could significantly improve
cardiac function by improving cellular survival and angiogenesis.
To a certain extent, our results from our in vitro
experiments are in accordance with the findings of that study.
However, compared to the previous study, our study has several
advantages. First, the interaction of VEGF and Bcl-2 was taken into
account in our selection of genes. It is not simply an additive
effect, but a mutual promotion. Second, our study used a lentiviral
vector to deliver the genes as this vector has a better
transfection efficiency and is less toxic to cells than the
adenoviral vector (23).
Importantly, the use of lentivirus enables final cell lines to
stably co-overexpress target genes, which allowed us to better
observe the effects of the genetic modification than with the
transient expression of the adenoviral vector. Moreover, a
self-cleaving T2A peptide sequence was used to link the two genes
and place them in a single lentiviral vector, which reduced the
viral dosage and ensured the equivalent expression level.
The Bcl-2 gene is also known to play an important
role in cellular autophagy (24);
thus, in this study, we also carried out simultaneous western blot
analyses to investigate several autophagy-related proteins in a
preliminary assessment of the effects of the dual genetic
modification under OGD conditions on autophagy in MSCs. Our results
suggested that the activation of autophagy in MSCs following 12 h
of exposure to OGD may be suppressed by dual genetic modification.
We then further confirmed that the suppression of autophagy
contributed to the inhibition of apoptosis in the OGD-exposed cells
in the MSC-BV group.
Therefore, the inhibition of apoptosis in the dually
modified cells may be partly achieved by the indirect suppression
of autophagy. The association between autophagy and apoptosis is
controversial. Initially, autophagy was found to favor cell
survival (25), but later it was
also found to play a role in cell death. Excessive and prolonged
autophagy may promote cell death (26,27). In this study, both the autophagy
and apoptosis of MSCs induced by OGD may contribute to cell death,
and was suppressed by the dual genetic modification.
The paracrine effect is an important mechanism in
the use of MSCs in the treatment of ischemic heart disease and has
been widely used as a target for improvement via various means,
including but not limited to, cellular factors. It has been shown
that VEGF-, bFGF-, HGF- and IGF-1-modified MSCs can significantly
enhance the paracrine effects (28–30). Our results from the ELISA
detection of four classical growth factors confirmed that the MSCs
dually modified with VEGF and Bcl-2 exhibited an enhanced paracrine
effect that improved MSC resistance to a harsh microenvironment,
further playing a protective role against ischemic disease.
Despite these encouraging results, some challenges
remain to be addressed further. First, the benefits from dually
modified VEGF and Bcl-2 MSCs must be further confirmed in an animal
study. Second, the signaling pathways involved in the inhibition of
apoptosis must be further clarified. Third, the association and
signaling pathways between VEGF and Bcl-2 in MSCs should be further
elucidated. Fourth, the role of autophagy in MSCs in a hostile
environment and its association with apoptosis must be further
investigated.
In conclusion, in this study, we constructed a
lentiviral vector that overexpressed the VEGF and Bcl-2 genes
simultaneously, and confirmed that the dual genetic modification of
the MSCs had a higher expression than singly modified MSCs. Such a
dual genetic modification strategy successfully made the MSCs much
more resistant to a hostile environment via the inhibition of
apoptosis, the suppression of autophagy and the enhancement of
paracrine signaling.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (grant no. 31271053).
References
|
1
|
Wang X, Zhang J, Zhang F, Li J, Li Y, Tan
Z, Hu J, Qi Y, Li Q and Yan B: The clinical status of stem cell
therapy for ischemic cardiomyopathy. Stem Cells Int.
2015:1350232015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang XJ and Li QP: The roles of
mesenchymal stem cells (MSCs) therapy in ischemic heart diseases.
Biochem Biophys Res Commun. 359:189–193. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Duran JM, Makarewich CA, Sharp TE,
Starosta T, Zhu F, Hoffman NE, Chiba Y, Madesh M, Berretta RM, Kubo
H, et al: Bone-derived stem cells repair the heart after myocardial
infarction through transdifferentiation and paracrine signaling
mechanisms. Circ Res. 113:539–552. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Young PP and Schäfer R: Cell-based
therapies for cardiac disease: A cellular therapist's perspective.
Transfusion. 55:441–451; quiz 440. 2015. View Article : Google Scholar
|
|
5
|
Lee S, Choi E, Cha MJ and Hwang KC: Cell
adhesion and long-term survival of transplanted mesenchymal stem
cells: A prerequisite for cell therapy. Oxid Med Cell Longev.
2015:6329022015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Penn MS and Mangi AA: Genetic enhancement
of stem cell engraftment, survival, and efficacy. Circ Res.
102:1471–1482. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Matsumoto R, Omura T, Yoshiyama M, Hayashi
T, Inamoto S, Koh KR, Ohta K, Izumi Y, Nakamura Y, Akioka K, et al:
Vascular endothelial growth factor-expressing mesenchymal stem cell
transplantation for the treatment of acute myocardial infarction.
Arterioscler Thromb Vasc Biol. 25:1168–1173. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li W, Ma N, Ong LL, Nesselmann C, Klopsch
C, Ladilov Y, Furlani D, Piechaczek C, Moebius JM, Lützow K, et al:
Bcl-2 engineered MSCs inhibited apoptosis and improved heart
function. Stem Cells. 25:2118–2127. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Baek JH, Jang JE, Kang CM, Chung HY, Kim
ND and Kim KW: Hypoxia-induced VEGF enhances tumor survivability
via suppression of serum deprivation-induced apoptosis. Oncogene.
19:4621–4631. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang D, Weng Q, Zhang L, He Q and Yang B:
VEGF and Bcl-2 interact via MAPKs signaling pathway in the response
to hypoxia in neuroblastoma. Cell Mol Neurobiol. 29:391–401. 2009.
View Article : Google Scholar
|
|
11
|
Shearer RF and Saunders DN: Experimental
design for stable genetic manipulation in mammalian cell lines:
Lentivirus and alternatives. Genes Cells. 20:1–10. 2015. View Article : Google Scholar
|
|
12
|
Sakuma T, Barry MA and Ikeda Y: Lentiviral
vectors: Basic to translational. Biochem J. 443:603–618. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ibrahimi A, Vande Velde G, Reumers V,
Toelen J, Thiry I, Vandeputte C, Vets S, Deroose C, Bormans G,
Baekelandt V, et al: Highly efficient multicistronic lentiviral
vectors with peptide 2A sequences. Hum Gene Ther. 20:845–860. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
McGinley LM, McMahon J, Stocca A, Duffy A,
Flynn A, O'Toole D and O'Brien T: Mesenchymal stem cell survival in
the infarcted heart is enhanced by lentivirus vector-mediated heat
shock protein 27 expression. Hum Gene Ther. 24:840–851. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang X, Zhao T, Huang W, Wang T, Qian J,
Xu M, Kranias EG, Wang Y and Fan GC: Hsp20-engineered mesenchymal
stem cells are resistant to oxidative stress via enhanced
activation of Akt and increased secretion of growth factors. Stem
Cells. 27:3021–3031. 2009.PubMed/NCBI
|
|
16
|
Mangi AA, Noiseux N, Kong D, He H, Rezvani
M, Ingwall JS and Dzau VJ: Mesenchymal stem cells modified with Akt
prevent remodeling and restore performance of infarcted hearts. Nat
Med. 9:1195–1201. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fan L, Lin C, Zhuo S, Chen L, Liu N, Luo
Y, Fang J, Huang Z, Lin Y and Chen J: Transplantation with
survivin-engineered mesenchymal stem cells results in better
prognosis in a rat model of myocardial infarction. Eur J Heart
Fail. 11:1023–1030. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tang YL, Zhao Q, Qin X, Shen L, Cheng L,
Ge J and Phillips MI: Paracrine action enhances the effects of
autologous mesenchymal stem cell transplantation on vascular
regeneration in rat model of myocardial infarction. Ann Thorac
Surg. 80:229–237. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Banai S, Jaklitsch MT, Shou M, Lazarous
DF, Scheinowitz M, Biro S, Epstein SE and Unger EF:
Angiogenic-induced enhancement of collateral blood flow to ischemic
myocardium by vascular endothelial growth factor in dogs.
Circulation. 89:2183–2189. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Deuse T, Peter C, Fedak PW, Doyle T,
Reichenspurner H, Zimmermann WH, Eschenhagen T, Stein W, Wu JC,
Robbins RC, et al: Hepatocyte growth factor or vascular endothelial
growth factor gene transfer maximizes mesenchymal stem cell-based
myocardial salvage after acute myocardial infarction. Circulation.
120(Suppl 11): S247–S254. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Locatelli P, Olea FD, Hnatiuk A, De
Lorenzi A, Cerdá M, Giménez CS, Sepúlveda D, Laguens R and
Crottogini A: Mesenchymal stromal cells overexpressing vascular
endothelial growth factor in ovine myocardial infarction. Gene
Ther. 22:449–457. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang S, Haider HK, Idris NM, Salim A and
Ashraf M: Supportive interaction between cell survival signaling
and angiocompetent factors enhances donor cell survival and
promotes angiomyo-genesis for cardiac repair. Circ Res. 99:776–784.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
McMahon JM, Conroy S, Lyons M, Greiser U,
O'shea C, Strappe P, Howard L, Murphy M, Barry F and O'Brien T:
Gene transfer into rat mesenchymal stem cells: A comparative study
of viral and nonviral vectors. Stem Cells Dev. 15:87–96. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lindqvist LM and Vaux DL: BCL2 and related
prosurvival proteins require BAK1 and BAX to affect autophagy.
Autophagy. 10:1474–1475. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Boya P, González-Polo RA, Casares N,
Perfettini JL, Dessen P, Larochette N, Métivier D, Meley D,
Souquere S, Yoshimori T, et al: Inhibition of macroautophagy
triggers apoptosis. Mol Cell Biol. 25:1025–1040. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Denton D, Xu T and Kumar S: Autophagy as a
pro-death pathway. Immunol Cell Biol. 93:35–42. 2015. View Article : Google Scholar
|
|
27
|
Fitzwalter BE and Thorburn A: Recent
insights into cell death and autophagy. FEBS J. 282:4279–4288.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Haider HK, Jiang S, Idris NM and Ashraf M:
IGF-1-overexpressing mesenchymal stem cells accelerate bone marrow
stem cell mobilization via paracrine activation of SDF-1alpha/CXCR4
signaling to promote myocardial repair. Circ Res. 103:1300–1308.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fiedler J, Brill C, Blum WF and Brenner
RE: IGF-I and IGF-II stimulate directed cell migration of
bone-marrow-derived human mesenchymal progenitor cells. Biochem
Biophys Res Commun. 345:1177–1183. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Song H, Song BW, Cha MJ, Choi IG and Hwang
KC: Modification of mesenchymal stem cells for cardiac
regeneration. Expert Opin Biol Ther. 10:309–319. 2010. View Article : Google Scholar : PubMed/NCBI
|