Introduction
In recent decades, the incidence of diabetes
mellitus has increased rapidly, and by 2025, the number of
individuals with diabetes is estimated to be 330–380 million.
Diabetes is recognized as a potent and prevalent risk factor for
ischemic heart disease. Among the 3.8 million deaths each year,
~2/3 are attributable to heart disease associated with diabetes
(http://www.eatlas.idf.org/). Diabetic
cardiomyopathy (DCM), associated with both type 1 and 2 diabetes,
represents one of the major cardiovascular complications in
diabetic patients and is characterized by both early-onset
diastolic and late-onset systolic dysfunctions that may lead to
heart failure (1,2). Several mechanisms have been
implicated in the pathogenesis of DCM. Among them, oxidative stress
is involved in the etiology of diabetes-induced downregulation of
heart function (3). The
beneficial effects of several molecules on DCM have been attributed
to the alleviation of oxidative stress (4).
Oxidative stress is defined as a serious imbalance
between the production of endogenous reactive oxygen species (ROS)
and antioxidants, and in favor of the former (5). Hyperglycemia, a major etiological
component in the development of DCM, is known to promote the
production of reactive nitrogen species (RNS) and ROS and/or to
deplete antioxidant systems in many cell types. Overproduction of
ROS is a causative factor in the development of DCM (6–8).
Blocking ROS formation has been shown to prevent hyperglycemic
damage (7). Even though
antioxidant prevention or therapy has been explored for many years,
there are few exogenous antioxidants that efficiently prevent DCM
in the clinic to date. Accordingly, activation of the endogenous
antioxidant pathways in the heart and the underlying molecular
mechanisms have been proposed as an attractive strategy (9).
Morus alba L., a plant of the Moraceous
family, has more than 4,000 years of history in China. Mulberry is
cultivated for fruit production, and its leaf is traditionally used
as the food of silkworms (10).
Among the various nutritional components of mulberry, flavonoids
are known to have effects as antioxidants or free radical
scavengers (11). It also
contains many phenolic antioxidants that can reduce cardiovascular
disease. Several recent studies have shown that mulberry has
antioxidant, anti-hyperglycemic, anti-inflammatory,
neuroprotective, and hypolipidemic and non-toxic activities
(12–14). Dietary mulberry has also been
reported to have hypoglycemic, hypolipidemic and antioxidant
effects (15). Based on LDL
oxidation assay, mulberry showed relatively high antioxidant
activity in comparison with 52 types of edible plant products in
Japan (16). Because of these
findings, the food of mulberry was developed into a type of
granules, named mulberry granules (MLD), which is convenient to
eat. MLD controls blood glucose levels in diabetes patients, and we
also found their beneficial effects on DCM in our preliminary
experiments. However, the beneficial effects and possible mechanism
remain largely unknown. This study was carried out to explore the
potential of MLD in the management of experimental myocardial
ischemia/reperfusion (MI/R) injury in diabetes mice. This
investigation may promote an understanding of the mechanisms
underlying the protective effects of MLD against
diabetes-associated cardiomyopathy.
Materials and methods
Chemicals and herbal materials
The fruits of the black mulberry (Morus
nigra) were purchased from Xi'an Zaolutang Pharmaceutical Co,
Ltd. (Xi'an, China) and were identified botanically by Professor
Haifeng Tang (Department of Natural Medicine, School of Pharmacy,
The Fourth Military Medical University, Xi'an, China). A voucher
specimen (no. 1509252) was deposited at the Department of Natural
Medicine, School of Pharmacy, The Fourth Military Medical
University.
MLD was obtained from Shaanxi Junbisha
Pharmaceutical Co., Ltd. (Xianyang, China). Chlorogenic acid (with
a purity >98% by HPLC analysis) was purchased from National
Institute for the Control of Pharmaceutical and Biological Products
(Beijing, China). HPLC-grade methanol and acetonitrile were
obtained both from Thermo Fisher Scientific, Inc. (Waltham, MA,
USA). Deionized water was purified using the Milli-Q Reagent Water
system (Millipore, Bedford, MA, USA). Streptozotocin (STZ) was
obtained from Sigma-Aldrich (St. Louis, MO, USA). Polyclonal rabbit
anti-mouse AMP-activated protein kinase (AMPK; 2532), p-AMPK
(2535), nuclear factor erythroid 2-related factor 2 (Nrf2; 12721)
and β-actin (4970) antibodies were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA). Lactate dehydrogenase (LDH),
creatinine kinase (CK)-MB, SOD, GSH, CAT and GR detection kit were
purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing,
China).
HPLC analysis
For phytochemical analysis, MLD (~0.50 g) was
extracted with 50 ml of 75% (w/v) ethanol in a water bath at 80°C
for 30 min with one shaking every 15 min. After adjusting to
ambient temperature, ethanol was then added to compensate for the
lost weight during the extraction. Finally, the supernatant was
filtered through a 0.22-mm membrane filter and injected into the
HPLC system for analysis.
HPLC analysis was performed on a Shimadzu LC-20AD
system (Shimadzu, Kyoto, Japan). The analytes were separated on a
Kromasil C18 (250 mm × 4.6 mm, 5 µm). The mobile phase
consisted of A (acetonitrile) and B (water containing 0.3%
phosphoric acid). The flow rate was 0.8 ml/min at 30°C. UV
absorbance was monitored at 254 nm for finger printing analysis and
quantitative analysis. Chromatographic data were collected and
processed by an Empower™ chromatographic working station (Waters,
Milford, MA, USA).
Determination of
1,1-diphenyl-2-picryl-hydrazyl (DPPH) radical scavenging
activity
The free radical scavenging activity of MLD was
measured by DPPH assay as we previously described (17). DPPH-free radical agent was
prepared as 0.0158 g dissolved in 50 ml 95% (v:v) ethanol, and
mixed with MLD solution with a ratio of 8:1. The mixture was shaken
and kept in the dark at 37°C for 30 min. Vitamin C (Vc) was
employed as a control. The quantity of DPPH remaining in the mixed
solution was measured at 517 nm.
Superoxide anion radical-scavenging
activity
Xanthine oxidase and xanthine-induced luminol
chemiluminescence measurements were carried out as previously
described (18) with some
modifications. The concentration gradient of the sample and Vc were
9.6, 19.2, 28.8, 38.4 and 48 µg/ml. The mixture, sample or
Vc and 4.5 ml of 50 mM Tris-HCl buffer (pH 8.2) were mixed in a 10
ml centrifuge tube, and then shaken and incubated at 25°C for 20
min. Preheated (25°C) 0.3 ml of 3 mM pyrogallol solution was then
added. After standing for 5 min, 0.2 ml of 10 M of HCl was added to
stop the reaction. The absorbance was measured at 325 nm. As a
control, 0.3 ml of 10 mM HCl replaced the pyrogallol solution.
Animals and type 2 diabetic mouse
model
The mice were obtained from the Experimental Animal
Center of the Fourth Military Medical University. The experimental
protocol was approved by the Ethics Committee for Animal
Experimentation and was performed according to the Guidelines for
Animal Experimentation of the Fourth Military Medical University
and the National Institute of Health Guide for the Care and Use of
Laboratory Animals (NIH publications no. 80–23) revised in 1996.
The animals were housed under a 12-h light/dark cycle and
temperature was kept at 25°C.
For the STZ diabetic model, 6- to 8-week-old male
C57BL/6 mice, weighing 23–25 g were used. They were randomly
grouped into diabetic (model) or non-diabetic (control) mice as
they received, for 5 consecutive days, an intraperitoneal injection
of STZ (Sigma-Aldrich) at a dose of 50 mg/kg or vehicle (0.1 mol/l
citrate buffer, pH 4.5) alone. After 4 weeks, blood glucose levels
were measured using Bayer BREEZE® 2 meter (Bayer
HealthCare LLC, Mishawaka, IN, USA) by tail vein blood sampling.
Mice with blood glucose levels >11.1 mM were used as the model
group in the present study.
MLD was dissolved in physiological saline before
use. Animals were randomly divided into five groups: sham, I/R, I/R
+ MLD (30 mg/kg), I/R + MLD (20 mg/kg) and I/R + MLD (10 mg/kg)
groups. MLD was administered through oral administration for one
month. Then, the animals were exposed to 30 min ischemia followed
by 24 h of reperfusion using a previously described technique
(19). After these operations,
infarct size and heart function were estimated.
Blood analyses
Glycemic parameters including fasting blood glucose
(FBG) and fasting blood insulin (FBI) were measured by enzymatic
end procedure and enzyme-linked immunosorbent assay (ELISA)
according to the manufacturer's instructions respectively.
Homeostasis model of insulin resistance (HOMA-IR) was calculated as
described: HOMA-IR (mass units) = fasting insulin (mIU/ml) ×
fasting glucose (mg/dl)/405. High-density lipoprotein cholesterol
(HDL-C) and low-density lipoprotein cholesterol (LDL-C), total
cholesterol (TC) and triglyceride (TG) in serum were determined
using of the Bayer 1650 blood chemistry analyzer (Bayer, Tarrytown,
NY, USA).
Cardiac function measurement
After induction of anesthesia with 5% chloral
hydrate by intraperitoneal injection, cardiac parameters were
detected by a biofunction experiment system MP100-CE (Biopac
Systems Inc., Santa Barbara, CA, USA). Heart rate (HR), left
ventricular systolic pressure (LVSP), left ventricular maximum
upstroke and descent velocity
(+dP/dtmax/−dP/dtmax) were measured by
computer algorithms and an interactive videographics program
(Po-Ne-Mah Physiology Platform P3 Plus). MI size was determined by
the Evans blue/TTC double-staining method as previously described
after completion of functional determination (20).
Detection of LDH and CK-MB levels
Blood samples were collected from the abdominal
aorta after 24 h of reperfusion and centrifuged at 3,250 × g for 10
min. LDH and CK-MB levels in plasma were measured to evaluate
myocardial damage using commercially available assay kits (Nanjing
Jiancheng Bioengineering Institute).
Detection of GSH, GSSG, CAT, SOD and
MDA
GSH, GSSG, CAT, SOD and MDA levels in cardiac muscle
tissues or myocardial cell were detected by the corresponding kit
according to the manufacturer's instructions.
Western blotting
After reperfusion, ~50 mg of myocardial tissue was
collected and stored at −80°C. Radioimmunoprecipitation assay
(RIPA) lysis buffer was used to extract the whole protein, and the
concentration was determined by the BCA protein kit according to
the manufacturer's instructions, and the mean concentration values
were computed. The assessment was carried out twice for each
sample, and the values were averaged. Then the samples were kept at
−80°C.
Equal amount of protein was separated by 10%
Tris-glycine SDS-PAGE polyacrylamide gel and transferred to
polyvinylidene fluoride membranes (Invitrogen Life Technologies,
Carlsbad, CA, USA). After blocking for 1.5 h with a 5% solution of
skim milk, the membranes were incubated with the primary antibodies
at 4°C overnight, followed by the horseradish peroxidase
(HRP)-linked anti-goat antibody for 60 min at 37°C. After
washing with Tris-buffered saline, the relative intensity of the
protein signals was normalized to the corresponding β-actin
intensity and was quantified by densitometric analysis with the use
of Quantity One software (Bio-Rad Laboratories, Inc., Hercules, CA,
USA).
Statistical analysis
The experiment was performed at least three times.
Data are expressed as means ± standard deviation (SD), and were
analyzed with SPSS statistical software (SPSS, Inc., Chicago, IL,
USA). Data were compared by one-way analysis of variance (ANOVA).
P<0.05 was considered statistically significant.
Results
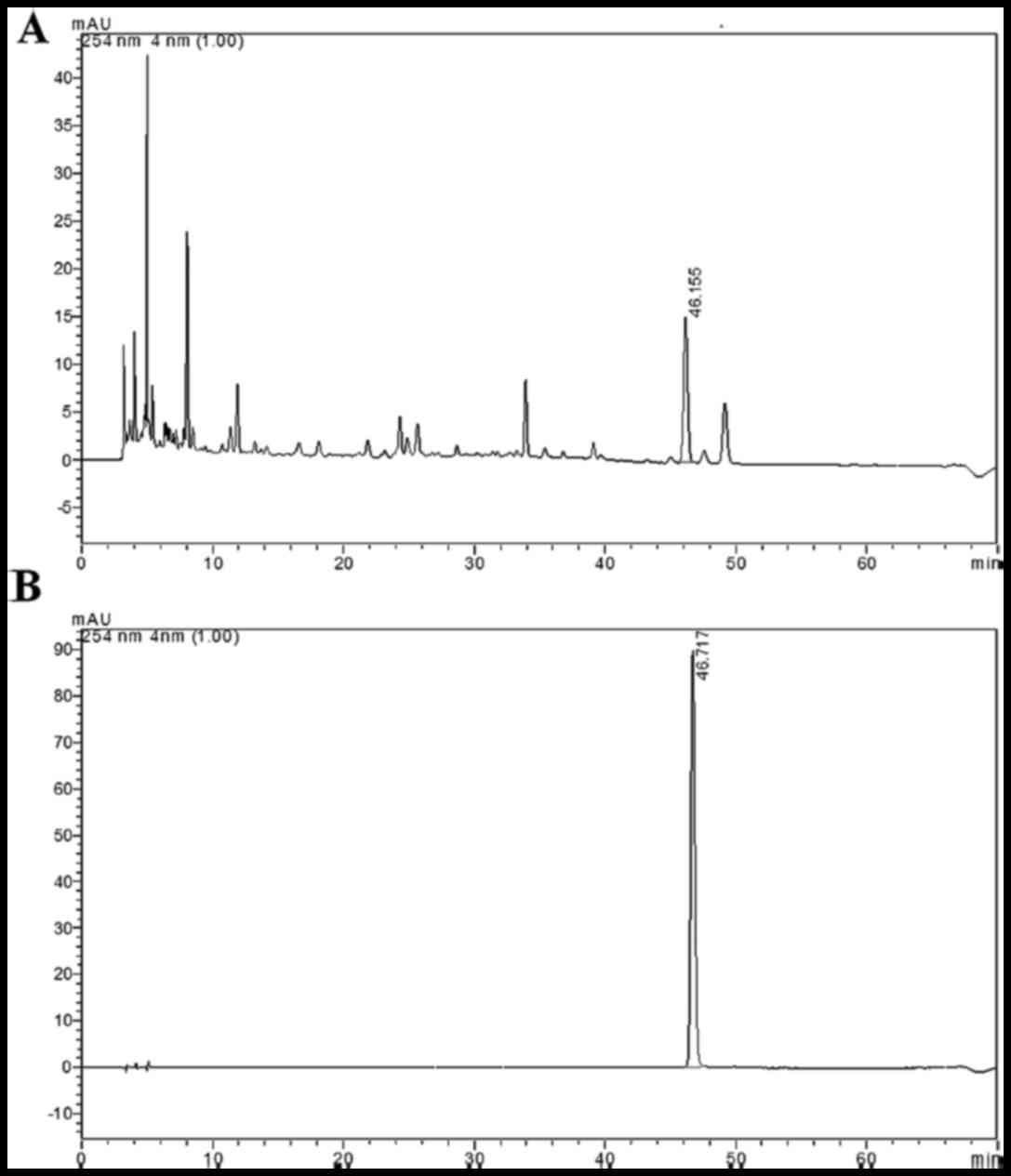
HPLC method validation and sample
analysis
To confirm the accuracy of the equipment, a sample
solution was successive injected for 5 times, and then the
correlation coefficients between the reference HPLC fingerprint and
the sample HPLC fingerprints were calculated. The values obtained
were 0.997, indicating that the accuracy of our measurements was
satisfactory. To determine the stability of the sample solutions
during the measurement period, at different time-points (0, 3, 6, 9
and 12 h), five injections of the same sample solution were
analyzed and the correlation coefficients were calculated. The
values obtained were 0.972 (0 h), 0.969 (3 h), 0.971 (6 h), 0.971
(9 h) and 0.972 (12 h), which confirmed that during the measurement
periods the sample solutions were stable. We also investigated the
repeatability of the analytical method with 5 solutions from the
same sample, and the values obtained were 0.997 (0 h), 0.998 (3 h),
0.999 (6 h), 0.998 (9 h), and 0.999 (12 h), which confirmed that
the method was repeatable. Using the method described above, the
MLD sample was analyzed. The HPLC fingerprinting profiles are shown
in Fig. 1. Chlorogenic acid was
used for chemical standardization of the MLD preparation. The
similarity between the values obtained for the 10 samples was
higher than 0.998. This indicate a high consistency and stability
between the batches of MLD tested.
Antioxidant activities
To measure radical scavenging activities, several
in vitro antioxidant assays were tested. DPPH is a stable
free radical and is often used to evaluate the antioxidant activity
of natural compounds. As shown in Fig. 2A, the activity of MLD in reducing
the DPPH free radical was as effective as Vc. Superoxide anion
radical and hydroxyl radical are two types of the most important
free radicals in organisms. Luminol chemiluminescence models
producing hydroxyl and superoxide anion radicals were also used to
evaluate the free radical-scavenging activity of MLD. Fig. 2B shows a high degree of hydroxyl
radical activity. Superoxide anion radical-scavenging activity of
Vc was more efficient than that of MLD (Fig. 2C). The results showed that MLD had
high antioxidant activity in vitro.
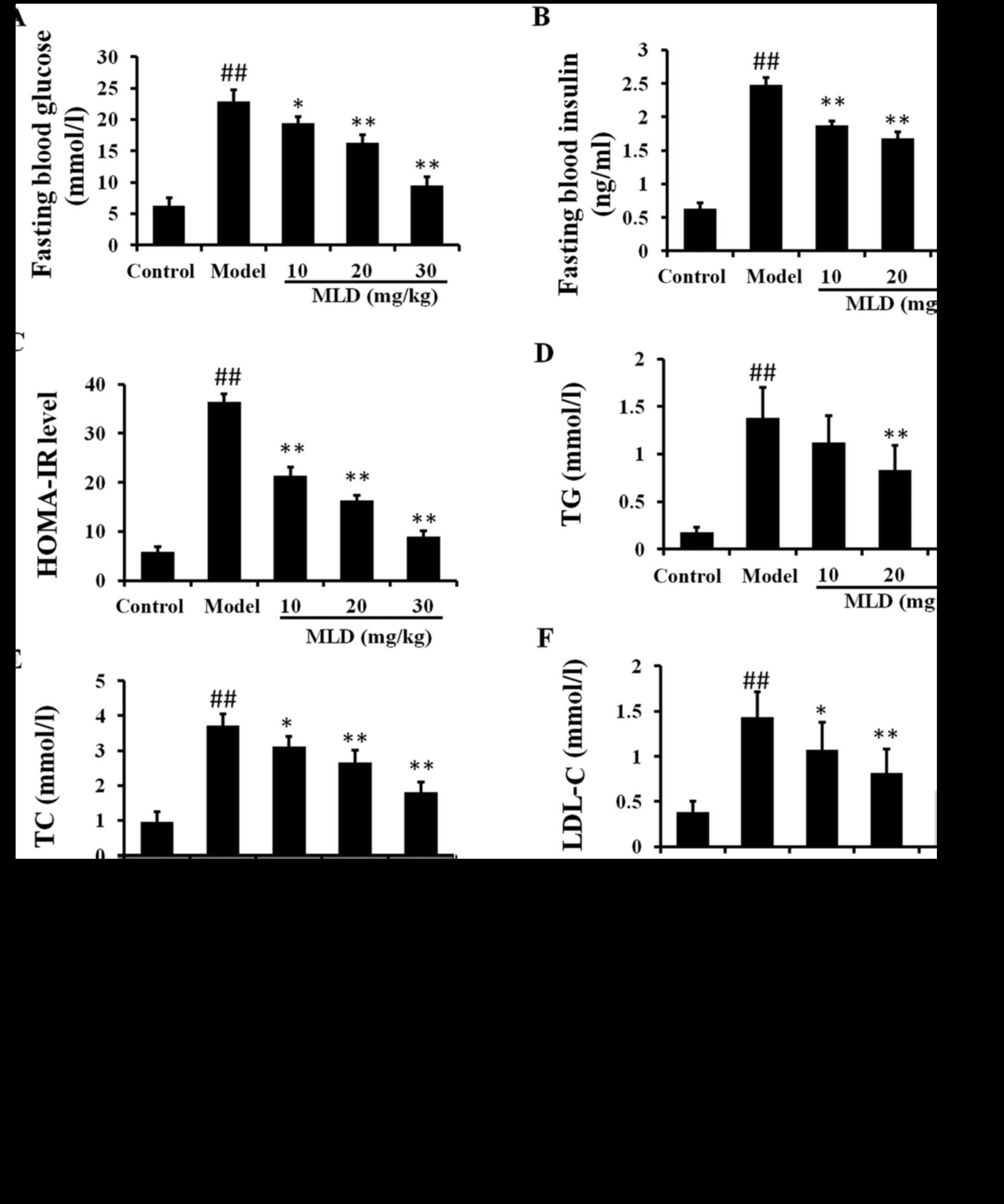
Effects of MLD on FBG, FBI and
lipids
Several symptoms characteristic of hyperglycemia,
hyperlipidemia, polydipsia and polyphagia were observed in the type
2 diabetic mice. There was no statistically significant difference
in FBG, FBI and lipids among the group at the beginning of the
experiment (P>0.05). The levels of FBG and FBI in the type 2
diabetic model mice were higher than those in the control mice at
the end of 4 weeks (P<0.01). FBG and FBI were significantly
lower in the MLD groups than levels in the diabetic mice at 4 weeks
(P<0.01) (Fig. 3A and B). The
HOMA-IR levels were also significantly decreased in the MLD groups,
as compared with the diabetic model group (Fig. 3C; P<0.01). These results
indicated that administration of MLD in diabetic mice resulted in a
significant improvement in the level of insulin resistance when
compared to the diabetic group.
In the type 2 diabetic model group, TG, TC and LDL-C
levels were significantly increased compared to these levels in the
control group (Fig. 3D–F;
P<0.01). There was no significant difference in HDL-C level
between the control and diabetic model group (Fig. 3G). After a 4-week treatment with
MLD, the TG, TC and LDL-C levels were significantly decreased
compared to levels in the diabetic group (P<0.01; 20 and 30
mg/kg MLD groups). After treatment with 20 and 30 mg/kg MLD, HDL-C
was significantly increased (P<0.01). These results showed that
type 2 diabetes was accompanied by insulin resistance and lipid
metabolism disorder and that MLD improved the negative influences
to some degree.
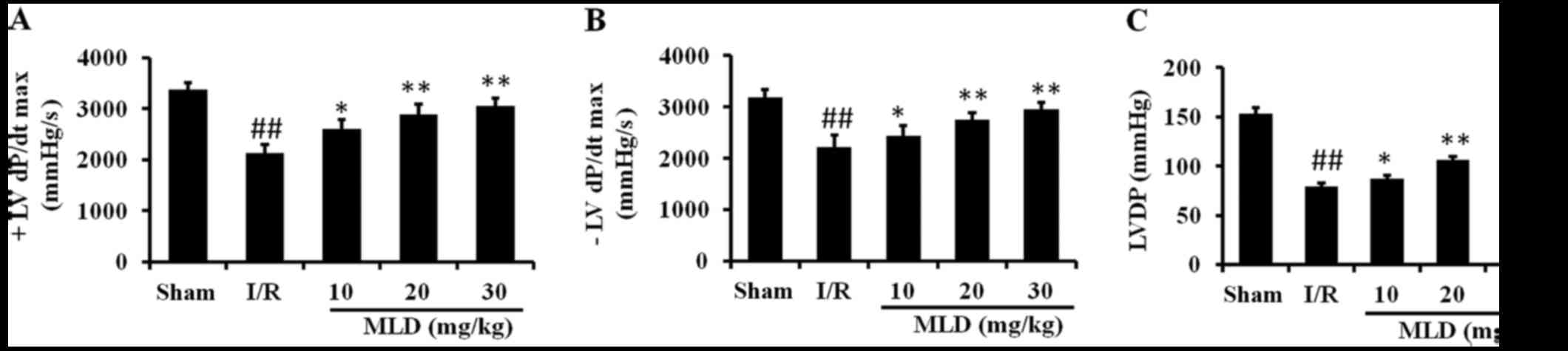
MLD improves the cardiac function after
MI/R in the diabetic mice
Cardiac function was examined to study whether MLD
has cardioprotective effects against I/R injury in diabetic mice.
As shown in Fig. 4, the LVDP, +LV
dP/dtmax and −LV dP/dtmax were significantly
decreased after I/R in the diabetic mice (P<0.01). Compared to
the sham group, I/R injury caused a significant decrease in +LV
dP/dtmax, −LV dP/dtmax and LVDP. In the MLD
treatment groups these cardiac function parameters were improved
significantly at the end of 3 h of reperfusion compared with the
I/R group (P<0.01; 20 and 30 mg/kg MLD groups). These results
showed that MLD improved cardiac functional recovery in the
diabetic mice subjected to I/R.
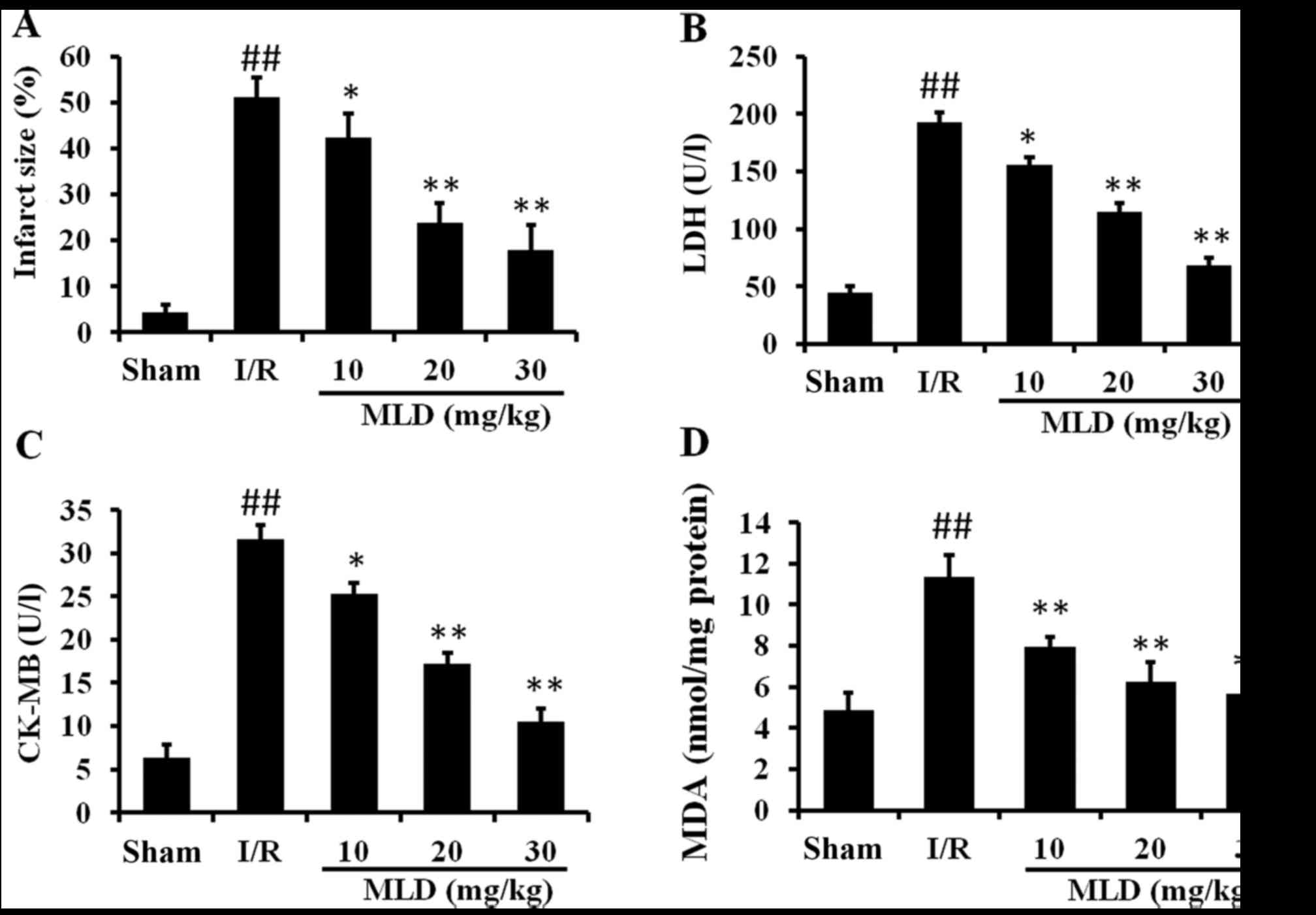
MLD inhibits myocardial injury following
MI/R in diabetic mice
As myocardial function is improved by MLD, the
infarct size was measured and is represented as a percentage of
infarct area/area at risk. As shown in Fig. 5A, I/R caused significant
myocardial infarction in the diabetic mice exposed to I/R compared
with the sham group (P<0.01). However, MLD significantly reduced
the infarct size compared to the I/R group (P<0.01; 20 and 30
mg/kg MLD groups).
The activities of CK-MB and LDH contents in serum
are commonly used to monitor damage of the myocardium. Relative to
the sham group, CK-MB and LDH contents were significantly increased
by I/R in the diabetic mice (P<0.01) (Fig. 5B and C). MLD significantly
reversed these abnormal changes compared with the I/R group
(P<0.01; 20 and 30 mg/kg MLD groups).
MDA, a marker of lipid peroxidation and free radical
activity, was also assessed in this study. As shown in Fig. 5D, I/R caused a significantly
increase in MDA levels in the diabetic model mice (P<0.01),
indicating that oxidative stress was induced by I/R and diabetes.
After pretreatment with MLD for 4 weeks, the MDA level was
significantly decreased in a dose-dependent manner (P<0.01).
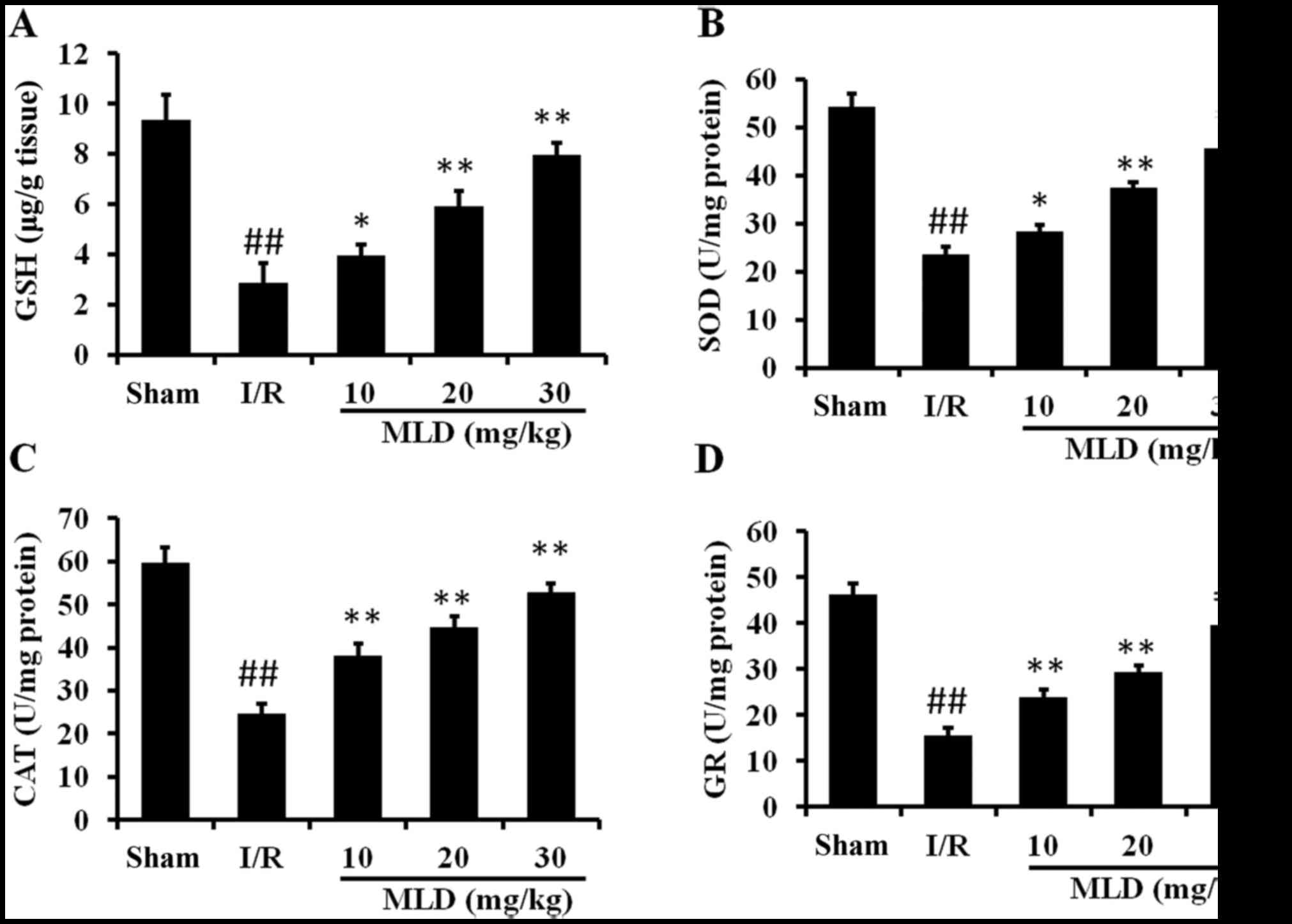
MLD enhances the antioxidant capacity in
cardiac tissues
To test whether the protective effects of MLD
against I/R-induced heart injury are due to its antioxidative
activity, several antioxidant proteins were assessed. As shown in
Fig. 6, the activities of GSH,
SOD, CAT and GR were significantly decreased by I/R in the diabetes
mice (P<0.01). Pretreatment with MLD before I/R reversed these
effects, which significantly included upregulation of actions of
these enzymes (P<0.01; 20 and 30 mg/kg MLD groups). These
findings suggest that MLD considerably improves cellular
antioxidative defense capacity against oxidative stress.
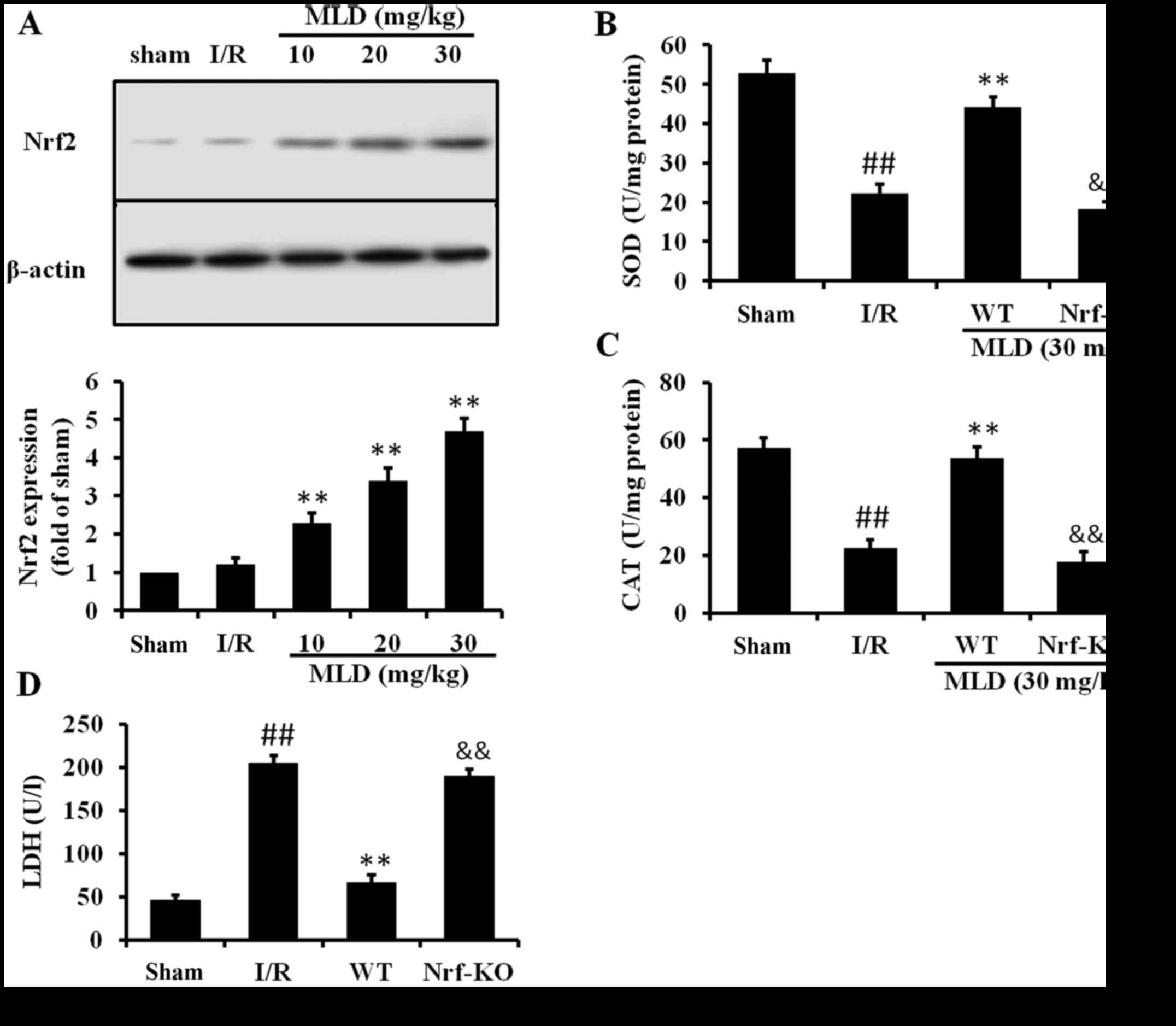
MLD activates the Nrf2 pathway
Nrf2 is a major regulator transcription factor
against oxidative stress and the transcription factor Nrf2 plays a
pivotal role in the induction of several antioxidant enzymes,
including GSH, SOD, and CAT. Therefore, we examined the effect of
MLD on Nrf2 expression to assess the mechanism responsible for the
antioxidative effect of MLD. As the results show, I/R caused a
slight increase in Nrf2 expression in the diabetic model mice
(Fig. 7A). Pretreatment with MLD
before I/R significantly increased expression of Nrf2 (P<0.01).
Subsequently, we compared the severity of heart and antioxidant
protein levels in the WT and Nrf2-knockout (KO) mice to determine
the role of Nrf2 in MLD. The results of the ELISA assay revealed an
increase in the levels of SOD and CAT in the WT diabetic mice
subjected to I/R. However, the levels of these two cytokines were
significantly lower in the Nrf2-KO mice compared with the WT mice
(Fig. 7B and C). Moreover, the
LDH level in the Nrf2-KO mice was significantly higher than that in
the WT mice (Fig. 7D). These
results demonstrated that the cardioprotective effects of MLD may
be through the Nrf2 pathway.
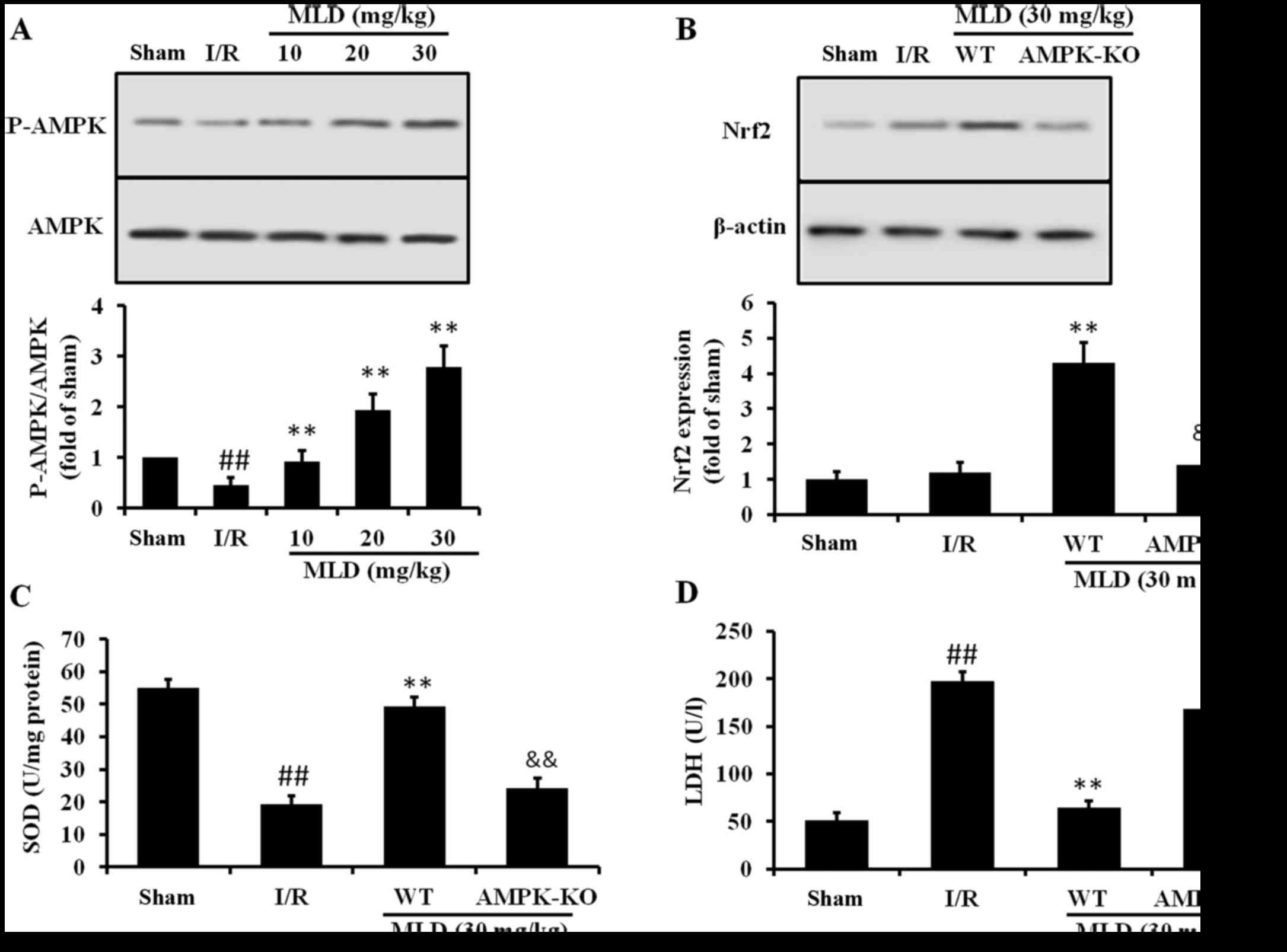
MLD activates the Nrf2 pathway through
AMPK
To investigate whether AMPK may be responsible for
the protective effect of MLD, protein kinase activities in cardiac
tissues were assessed by determining the phosphorylated form of
AMPK. As shown in Fig. 8A, MLD
administered p.o. for 4 weeks significantly (P<0.01) reversed
the decreased phosphorylation of AMPK caused by I/R. To determine
whether activation of Nrf2 is due to increased AMPK mediated by
MLD, AMPK-KO mice were used. As the results show, AMPK-KO abolished
the Nrf2 expression and cardioprotective effects (Fig. 8B and C) induced by AMPK,
suggesting that the AMPK increase is upstream and is required for
Nrf2 activation mediated by MLD.
Discussion
Diabetic patients have a 2- to 5-fold increased risk
of developing cardiovascular diseases compared with non-diabetic
patients, which include hypertensive heart disease, atherosclerosis
and DCM (20). DCM is
particularly difficult to manage due to its asymptomatic nature and
its ability to progress into HR (21,22). Several cardiovascular pathological
consequences of diabetes such as hypertension affect the heart to
varying degrees. However, hyperglycemia, as an independent risk
factor, directly causes cardiac damage and leads to DCM (6). Hyperglycemia induces overproduction
of reactive oxygen and nitrogen species, and then an outbreak of
oxidative stress in the heart. Oxidative stress causes abnormal
expression of genes or proteins, altered signal transduction and
programmed myocardial cell death (23). Thus, myocardial cell loss plays a
critical role in the onset and development of DCM. For the
treatment of DCM, advances in the application of drugs against
hyperglycemia-induced oxidative myocardial injury may be
useful.
The edible product of mulberry, which contains
flavonoids and phenols, is delicious and popular in many countries.
It has been used to treat diabetes mellitus in ancient
prescriptions and folk medicine. However, the food from mulberry
physically deteriorates quickly following maturation, and does not
favor preservation. Thus, mulberry was developed into a type of
granules, MLD, which can be preserved for a long time. We found
that it could protect diabetics from cardiovascular disease. But
the cardioprotective effects and possible mechanism are largely
unknown. In this study, HPLC was used for fingerprinting analysis
and MI/R injury in a diabetic mouse model was used to evaluate its
efficacy and possible mechanism.
There are several mechanisms involved in the
function of antioxidants, one of which is the scavenging of ROS and
free radicals. To evaluate the activity of MLD in scavenging free
radical, two free radical-generating models were used. DPPH, a
stable free radical, is commonly used in testing free
radical-scavenging activity. Any molecule that can transfer
electrons or hydrogen atoms to DPPH may react with it and reduce
the absorbance at 517 nm (24).
When MLD was tested for DPPH-scavenging ability, it showed a strong
DPPH-scavenging ability, which was similar to that of Vc. In this
study, we also used xanthine oxidase-induced luminol
chemiluminescence model. Compared with Vc, MLD showed a certain
degree of free radical-scavenging activity. These results
demonstrated that MLD could scavenge free radical directly in
vitro.
To evaluate the cardioprotective effects of MLD,
MI/R was induced in diabetic mouse. We found that MLD reduced FBG,
FBI and lipids which were induced by STZ in the diabetic mice. We
also found that MLD significantly improved cardiac dysfunction,
including +LV dP/dtmax, −LV dP/dtmax and
LVDP. Cytosolic enzymes, such as LDH and CK-MB, serve as diagnostic
markers of myocardial ischemia injury. Upon injury, the cell
membrane becomes permeable or ruptures, and then the enzymes leak
out from the damaged myocardial tissues to the blood stream. Thus,
they are considered as important indicators to determine reversible
vs. irreversible damage to cells. In this study, LDH and CK-MB were
increased by I/R in the diabetic mice, suggesting that I/R and
diabetes cause serious injury to the heart. However, MLD reversed
those changes. To the best of our knowledge, this was the first
study to demonstrate the protective role of MLD in diabetic mice
with MI/R injury.
Chronic hyperglycemia has been reported to trigger
oxidative stress either by direct generation of ROS or by altering
the redox balance (25). The
ability of antioxidants to inhibit hyperglycemia-derived injury has
raised the possibility of new therapeutic treatments for diabetic
heart diseases. In the present study, the ischemic heart of the
diabetic mice showed a significant increase in the lipid
peroxidation marker, MDA, with concomitant decreased GSH, SOD, CAT
and GSR. MLD supplementation markedly attenuated I/R-induced
oxidative stress by preventing GSH depletion and enhancement of the
enzymatic antioxidants in the diabetic mice. These results together
with the in vitro results indicated that the
cardioprotective effects of MLD may occur by increasing the
expression of endogenous antioxidant enzymes and scavenging free
radical in the body directly.
Nrf2, a key transcriptional regulatory protein,
activates antioxidant enzyme genes in response to oxidative stress
(26). It is broadly expressed in
multiple tissues but is only activated in response to various
electrophilic agents and oxidative stress including ROS, some
antioxidants and certain disease processes. Upon activation, Nrf2
through its interaction with the antioxidant response element
(ARE), mediates the induction of a series of cytoprotective
proteins including phase II enzymes, such as SOD, GSH and CAT
(27,28). These studies suggested that
activation of the Nrf2/ARE pathway may be involved in the protein
expression of SOD, CAT, GSH and GSR induced by MLD treatment. In
the present study, the results showed that MLD upregulated Nrf2
expression in the hearts of the diabetic mice subjected to I/R, and
also showed that expression of antioxidant proteins in Nrf2-KO mice
was lower than that in WT mice. Under the condition of the same
treatment, the level of injury in Nrf2-KO mice was much more
serious than that in the WT mice. These results demonstrated that
the cardioprotective effects of MLD may be through the activation
of Nrf2 and its downstream proteins.
AMPK, a key factor in fat and carbohydrate
metabolism that acts as a master switch of cellular metabolism,
plays an important role in mediating the change between anabolic
and catabolic states. Several factors activate the kinase, mainly
cellular energy stress during physiological (exercise) or
pathological (ischemia) situations, leading to a rise in AMP/ATP
and ADP/ATP ratios (29). Once
activated, AMPK induces several cellular signaling cascades and
then activates metabolic key enzymes, and gene transcription, thus
protecting the cell from I/R injury or other injury factors. A
previous study also showed that defects in AMPK signaling result in
the metabolic abnormalities of type 2 diabetes (30). In the present study, we
demonstrated that MLD induced the phosphorylation of AMPK. To
further clarify the role of AMPK in activating Nrf2, AMPK-KO mice
were used. We found that the effect of MLD on expression of Nrf2
was abolished by AMPK-KO and also the cardioprotective effects.
In conclusion, a simple and rapid HPLC method was
developed and validated the active compounds in MLD. MLD improved
insulin resistance, decreased blood glucose, TG, TC and LDL levels
in a diabetic mouse model. MLD has a protective effect on DCM by
inhibiting hyperglycemia-induced oxidative stress through
activation of the AMPK/Nrf2 pathway. Given the key role of
oxidative stress in the progression of DCM, MLD can be potentially
used as an effective drug for the treatment of type 2 diabetes
mellitus and cardiovascular diseases.
Acknowledgments
This study was supported by the '13115' Technology
Innovation Project of Engineering Research Center of Shaanxi
Province (no. 2013ZDGC-01).
References
|
1
|
Pappachan JM, Varughese GI, Sriraman R and
Arunagirinathan G: Diabetic cardiomyopathy: pathophysiology,
diagnostic evaluation and management. World J Diabetes. 4:177–189.
2013.PubMed/NCBI
|
|
2
|
Galderisi M: Diastolic dysfunction and
diabetic cardiomyopathy: evaluation by Doppler echocardiography. J
Am Coll Cardiol. 48:1548–1551. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ayaz M, Ozdemir S, Ugur M, Vassort G and
Turan B: Effects of selenium on altered mechanical and electrical
cardiac activities of diabetic rat. Arch Biochem Biophys.
426:83–90. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ceriello A: New insights on oxidative
stress and diabetic complications may lead to a 'causal'
antioxidant therapy. Diabetes Care. 26:1589–1596. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Vassort G and Turan B: Protective role of
antioxidants in diabetes-induced cardiac dysfunction. Cardiovasc
Toxicol. 10:73–86. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cai L and Kang YJ: Oxidative stress and
diabetic cardiomyopathy: a brief review. Cardiovasc Toxicol.
1:181–193. 2001. View Article : Google Scholar
|
|
7
|
Nishikawa T, Edelstein D, Du XL, Yamagishi
S, Matsumura T, Kaneda Y, Yorek MA, Beebe D, Oates PJ, Hammes HP,
et al: Normalizing mitochondrial superoxide production blocks three
pathways of hyperglycaemic damage. Nature. 404:787–790. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Inoguchi T, Li P, Umeda F, Yu HY, Kakimoto
M, Imamura M, Aoki T, Etoh T, Hashimoto T, Naruse M, et al: High
glucose level and free fatty acid stimulate reactive oxygen species
production through protein kinase C-dependent activation of NAD(P)H
oxidase in cultured vascular cells. Diabetes. 49:1939–1945. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cai L: Diabetic cardiomyopathy and its
prevention by metallothionein: experimental evidence, possible
mechanisms and clinical implications. Curr Med Chem. 14:2193–2203.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Arabshahi-D S, Vishalakshi Devi D and
Urooj A: Evaluation of antioxidant activity of some plant extracts
and their heat, pH and storage stability. Food Chem. 100:1100–1105.
2007. View Article : Google Scholar
|
|
11
|
Shibata Y, Kume N, Arai H, Hayashida K,
Inui-Hayashida A, Minami M, Mukai E, Toyohara M, Harauma A,
Murayama T, et al: Mulberry leaf aqueous fractions inhibit
TNF-alpha-induced nuclear factor kappaB (NF-kappaB) activation and
lectin-like oxidized LDL receptor-1 (LOX-1) expression in vascular
endothelial cells. Atherosclerosis. 193:20–27. 2007. View Article : Google Scholar
|
|
12
|
Du J, He ZD, Jiang RW, Ye WC, Xu HX and
But PP: Antiviral flavonoids from the root bark of Morus alba L.
Phytochemistry. 62:1235–1238. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pan G and Lou C: Isolation of an
1-aminocyclopropane-1-carboxylate oxidase gene from mulberry (Morus
alba L.) and analysis of the function of this gene in plant
development and stresses response. J Plant Physiol. 165:1204–1213.
2008. View Article : Google Scholar
|
|
14
|
Sun F, Shen L and Ma Z: Screening for
ligands of human aromatase from mulberry (Mori alba L.) leaf by
using high-performance liquid chromatography/tandem mass
spectrometry. Food Chem. 126:1337–1343. 2011. View Article : Google Scholar
|
|
15
|
El-Beshbishy HA, Singab AN, Sinkkonen J
and Pihlaja K: Hypolipidemic and antioxidant effects of Morus alba
L. (Egyptian mulberry) root bark fractions supplementation in
cholesterol-fed rats. Life Sci. 78:2724–2733. 2006. View Article : Google Scholar
|
|
16
|
Enkhmaa B, Shiwaku K, Katsube T, Kitajima
K, Anuurad E, Yamasaki M and Yamane Y: Mulberry (Morus alba L.)
leaves and their major flavonol quercetin 3-(6-malonylglucoside)
attenuate atherosclerotic lesion development in LDL
receptor-deficient mice. J Nutr. 135:729–734. 2005.PubMed/NCBI
|
|
17
|
Gulcin I: Antioxidant properties of
resveratrol: a structure-activity insight. Innov Food Sci Emerg.
11:210–218. 2010. View Article : Google Scholar
|
|
18
|
Banu S, Greenway GM and Wheatley RA:
Luminol chemiluminescence inducedby immobilised xanthine oxidase.
Anal Chim Acta. 541:89–95. 2005. View Article : Google Scholar
|
|
19
|
Huang XL, Wang W and Zhou YW: Protective
effect of epimedium flavonoids injection on experimental myocardial
infarction rats. Zhongguo Zhong Xi Yi Jie He Za Zhi. 26:68–71.
2006.In Chinese. PubMed/NCBI
|
|
20
|
Trachanas K, Sideris S, Aggeli C,
Poulidakis E, Gatzoulis K, Tousoulis D and Kallikazaros I: Diabetic
cardiomyopathy: from pathophysiology to treatment. Hellenic J
Cardiol. 55:411–421. 2014.PubMed/NCBI
|
|
21
|
Aneja A, Tang WH, Bansilal S, Garcia MJ
and Farkouh ME: Diabetic cardiomyopathy: insights into
pathogenesis, diagnostic challenges, and therapeutic options. Am J
Med. 121:748–757. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Arrington CB, Dowse BR, Bleyl SB and
Bowles NE: Non-synonymous variants in pre-B cell leukemia homeobox
(PBX) genes are associated with congenital heart defects. Eur J Med
Genet. 55:235–237. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ustinova EE, Barrett CJ, Sun SY and
Schultz HD: Oxidative stress impairs cardiac chemoreflexes in
diabetic rats. Am J Physiol Heart Circ Physiol. 279:H2176–H2187.
2000.PubMed/NCBI
|
|
24
|
Gao X, Ohlander M, Jeppsson N, Björk L and
Trajkovski V: Changes in antioxidant effects and their relationship
to phytonutrients in fruits of sea buckthorn (Hippophae rhamnoides
L.) during maturation. J Agric Food Chem. 48:1485–1490. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rains JL and Jain SK: Oxidative stress,
insulin signaling, and diabetes. Free Radic Biol Med. 50:567–575.
2011. View Article : Google Scholar
|
|
26
|
Itoh K, Wakabayashi N, Katoh Y, Ishii T,
Igarashi K, Engel JD and Yamamoto M: Keap1 represses nuclear
activation of antioxidant responsive elements by Nrf2 through
binding to the amino-terminal Neh2 domain. Genes Dev. 13:76–86.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ma Q, Kinneer K, Bi Y, Chan JY and Kan YW:
Induction of murine NAD(P)H:quinone oxidoreductase by
2,3,7,8-tetrachlorodibenzo-p-dioxin requires the CNC (cap 'n'
collar) basic leucine zipper transcription factor Nrf2 (nuclear
factor erythroid 2-related factor 2): cross-interaction between AhR
(aryl hydrocarbon receptor) and Nrf2 signal transduction. Biochem
J. 377:205–213. 2004. View Article : Google Scholar
|
|
28
|
He X, Chen MG, Lin GX and Ma Q: Arsenic
induces NAD(P)H quinone oxidoreductase I by disrupting the
Nrf2•Keap1•Cul3 complex and recruiting Nrf2•Maf to the antioxidant
response element enhancer. J Biol Chem. 281:23620–23631. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hardie DG: AMP-activated protein kinase:
an energy sensor that regulates all aspects of cell function. Genes
Dev. 25:1895–1908. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sriwijitkamol A, Ivy JL, Christ-Roberts C,
DeFronzo RA, Mandarino LJ and Musi N: LKB1-AMPK signaling in muscle
from obese insulin-resistant Zucker rats and effects of training.
Am J Physiol Endocrinol Metab. 290:E925–E932. 2006. View Article : Google Scholar
|