Introduction
Chemokines have been shown to control the migratory
behavior of several cell types, including lymphocytes. Helper
T-lymphocytes (Th1), cellular mediators of adaptive immune
responses, produce interferon-γ (IFN-γ) that can stimulate
macrophages, epithelial cells and tissue parenchymal cells to
express CXC chemokines, such as monokine induced by IFN-γ (MIG),
IFN-inducible protein of 10 kDa (IP-10) and IFN-inducible T-cell
alpha chemoattractant (I-TAC) (1–5).
It has also been shown that Th1 cells preferentially express CXC
chemokine receptor 3 (CXCR3), which binds IP-10, I-TAC and MIG with
high affinity, and the binding of CXCR3 ligands to the receptor
produces cellular signals important for chemotaxis and the
activation of T-cells (6,7), suggesting that the expression of
IP-10, MIG and I-TAC is important in T-lymphocyte recruitment and
host defense following various infections.
Immune responses that contribute to host defense are
also capable of causing tissue injury and disease under
pathological conditions. There is increasing evidence to indicate
that during the development of type 1 diabetes, autoreactive
T-cells transverse the endothelium and matrix barriers to
infiltrate pancreatic islets and these autoreactive T-cells target
a number of islet cell autoantigens, including insulin (8), glutamic acid decarboxylase (9) and protein tyrosine phosphatase
(10). The infiltration of immune
cells in islets, termed insulitis, increases progressively once it
begins and leads to β-cell destruction, insulin deficiency and
clinical type 1 diabetes (11).
It has been reported that insulitic lesions are characterized by
the presence of β-cells, elevated levels of chemokines, such as
IP-10, and the infiltration of lymphocytes expressing CXCR3
(12), which may suggest that
IFN-γ-inducible chemokines may play crucial roles in the initiation
of insulitis.
Sulforaphane
(1-isothiocyanato-4-methylsulfinylbutane, SFN) is a dietary
isothiocyanate found abundantly in cruciferous vegetables and has
been shown to possess anti-inflammatory and immune modulatory
activities (13,14). For instance, SFN has been shown to
suppress the bacterial lipopolysaccharide-mediated expression of
inducible nitric oxide synthase, cyclooxygenase 2, interleukin-1
and tumor necrosis factor-α (TNF-α) in RAW 264.7 and peritoneal
macrophages (13,15,16). It has also been reported that SFN
inhibits TNF-α-induced nuclear factor-κB (NF-κB) activation, which
leads to the reduced expression of NF-κB-regulated gene products,
such as matrix metalloproteinase-9 (17), further supporting its
anti-inflammatory activity. As MIG, IP-10 and I-TAC play an
important role in T-cell recruitment for the initiation of adaptive
immunity, therapies aiming at reducing the levels of these
chemokines are of great interest.
In this study, we primarily investigated the effects
of IFN-γ on the expression levels of MIG, IP-10 and I-TAC in INS-1
cells, a rat pancreatic β-cell line, and secondly determined
whether SFN modulates the IFN-γ-induced expression of MIG, IP-10
and I-TAC in INS-1 cells. To the best of our knowledge, we report
for the first time, the ability of SFN to strongly inhibit the
IFN-γ-induced mRNA expression of MIG, IP-10 and I-TAC in INS-1
cells through the modulation of the expression and/or
phosphorylation levels of interferon regulatory factor (IRF)-1,
signal transducer and activator of transcription-1 (STAT-1) and
protein kinase B (PKB).
Materials and methods
Materials
Recombinant rat IFN-γ was obtained from R&D
Systems (Minneapolis, MN, USA). JAK inhibitor I (a pan-JAK
inhibitor) and LY294002 (a PI3K/PKB inhibitor) were obtained from
Calbiochem (Billerica, MA, USA) and Biomol (Plymouth Meeting, PA,
USA), respectively. Anti-p-STAT-1 (#9171), anti-STAT-1 (#9175),
anti-p-PKB (#9271) and anti-PKB (#9272) antibodies were purchased
from Cell Signaling Technology (Boston, MA, USA). β-actin antibody
(A5441) was purchased from Sigma (St. Louis, MO, USA). PCR primers
were purchased from Bioneer (Daejeon, Korea). RPMI-1640 was
purchased from Gibco-BRL (Carlsbad, CA, USA). Other chemicals,
including SFN, were obtained from Sigma.
Cell culture
The INS-1 cells, an immortalized rat pancreatic
β-cell line (Korean Cell Line Bank, Seoul, Korea), were cultured in
RPMI-1640 medium containing 11.2 mM glucose, 2 mM l-glutamine, 10%
heat-inactivated fetal bovine serum, 1 mM pyruvate, 10 mM HEPES, 50
μM 2-mercaptoethanol and 100 μg/ml streptomycin.
Treatment of the INS-1 cells with IFN-γ,
SFN, and/or other agents
The INS-1 cells were treated with or without IFN-γ
at various concentrations (0, 0.25, 0.5, 1, 2, 5 or 10 ng/ml) and
for different periods of time (0, 0.5, 2, 6 or 24 h). The INS-1
cells were also treated with or without IFN-γ (10 ng/ml) in the
absence or presence of SFN at various concentrations (0, 5, 10 or
25 μM) for 6 h. For pharmacological inhibition experiments,
the INS-1 cells were treated with or without IFN-γ (10 ng/ml) in
the absence or presence of JAK inhibitor I (JI, 0.1 μM), a
pan-JAK inhibitor, or LY294002 (LY, 25 μM), a PI3K/PKB
inhibitor, for 6 h. For comparison, the INS-1 cells were treated
with or without IFN-γ (10 ng/ml) in the absence or presence of SFN
(25 μM), curcumin (10 μM), sanguinarine (500 nM),
resveratrol (25 μM), triptolide (40 nM) or EGCG (25
μM) for 6 h.
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total RNA was isolated using TRIzol reagent (Life
Technologies, Gaithersburg, MD, USA), and cDNA was prepared using
M-MLV reverse transcriptase (Gibco-BRL, Carlsbad, CA, USA)
according to the manufacturers' instructions. The following primers
were used for the amplification of rat MIG, IP-10, I-TAC, IRF-1,
IRF-2 and glyceraldehyde 3-phosphate dehydro-genase (GAPDH): MIG
sense, 5′-CAG CCA AGG CAC ATT CCA CT-3′ and antisense, 5′-GAT GCA
GAG CGC TTG TTG GT-3′; IP-10 sense, 5′-CAA GTG CTG CTG TCG TTC
TC-3′ and antisense, 5′-TCT CTC TGC TGT CCA TCG GT-3′; I-TAC sense,
5′-AGA TCA CCA GAG CCA CAG CA-3′ and antisense, 5′-ATC CGA CCT CCT
AGC GAG TT-3′; IRF-1 sense, 5′-AAG TGA AGG ACC AGA GCA GG-3′ and
antisense, 5′-CTG TTG CAG CTT CAG AGG TG-3′; IRF-2 sense, 5′-ACA
ACG CCT TCA GAG TCT AC-3′ and antisense, 5′-TGC ATA GGA AGA CAC AGG
AG-3′; and GAPDH sense, 5′-CCG TAT CGG ACG CCT GGT TA-3′ and
antisense, 5′-TGG TGG TGC AGG ATG TAT TG-3′. The PCR amplification
was carried out using the following cycling conditions: 94°C for 3
min followed by 17–25 cycles of 94°C for 45 sec, 58°C for 45 sec,
72°C for 45 sec, and a final extension at 72°C for 5 min. The
amplified products were separated by electrophoresis on a 1.5%
agarose gel and detected under UV light.
Preparation of whole cell lysates
The INS-1 cells were washed with cold
phosphate-buffered saline (PBS) and lysed in a modified RIPA buffer
(50 mM Tris-HCl pH 7.4, 1% NP-40, 0.25% Na-deoxycholate, 150 mM
NaCl, 1 mM Na3VO4 and 1 mM NaF) containing
protease inhibitors (100 μM phenyl-methylsulfonyl fluoride,
10 μg/ml leupeptin, 10 μg/ml pepstatin and 2 mM EDTA)
on ice for 15 min. The cell lysates were collected and centrifuged
at 10,000 × g at 4°C for 10 min. The supernatant was saved and
protein concentrations were determined using Bradford reagent
(Bio-Rad, Hercules, CA, USA).
Western blot analysis
Proteins (50 μg) were separated by sodium
dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE; 10%)
and transferred onto Immobilon-P membranes. The membranes were
washed with TBS (10 mM Tris, 150 mM NaCl) supplemented with 0.05%
(v/v) Tween-20 (TBST) followed by blocking with TBST containing 5%
(w/v) non-fat dried milk. The membranes were incubated overnight
with antibodies specific for p-STAT-1 (1:1,000), STAT-1 (1:1,000),
p-PKB (1:2,000) or PKB (1:1,000) at 4°C. The membranes were then
exposed to secondary antibodies [anti-rabbit IgG (sc-2004) and
anti-mouse IgG (sc-2005); purchased from Santa Cruz Biotechnology,
Delaware, CA, USA] coupled to horseradish peroxidase for 2 h at
room temperature. β-actin (dilution, 1:10,000) was used as an
internal control. The membranes were washed 3 times with TBST at
room temperature. Immunoreactivities were detected by ECL
reagents.
Results
Dose- and time-dependent increase in the
mRNA levels of the T-cell-specific chemokines, MIG, IP-10 and
I-TAC, in INS-1 cells treated with IFN-γ
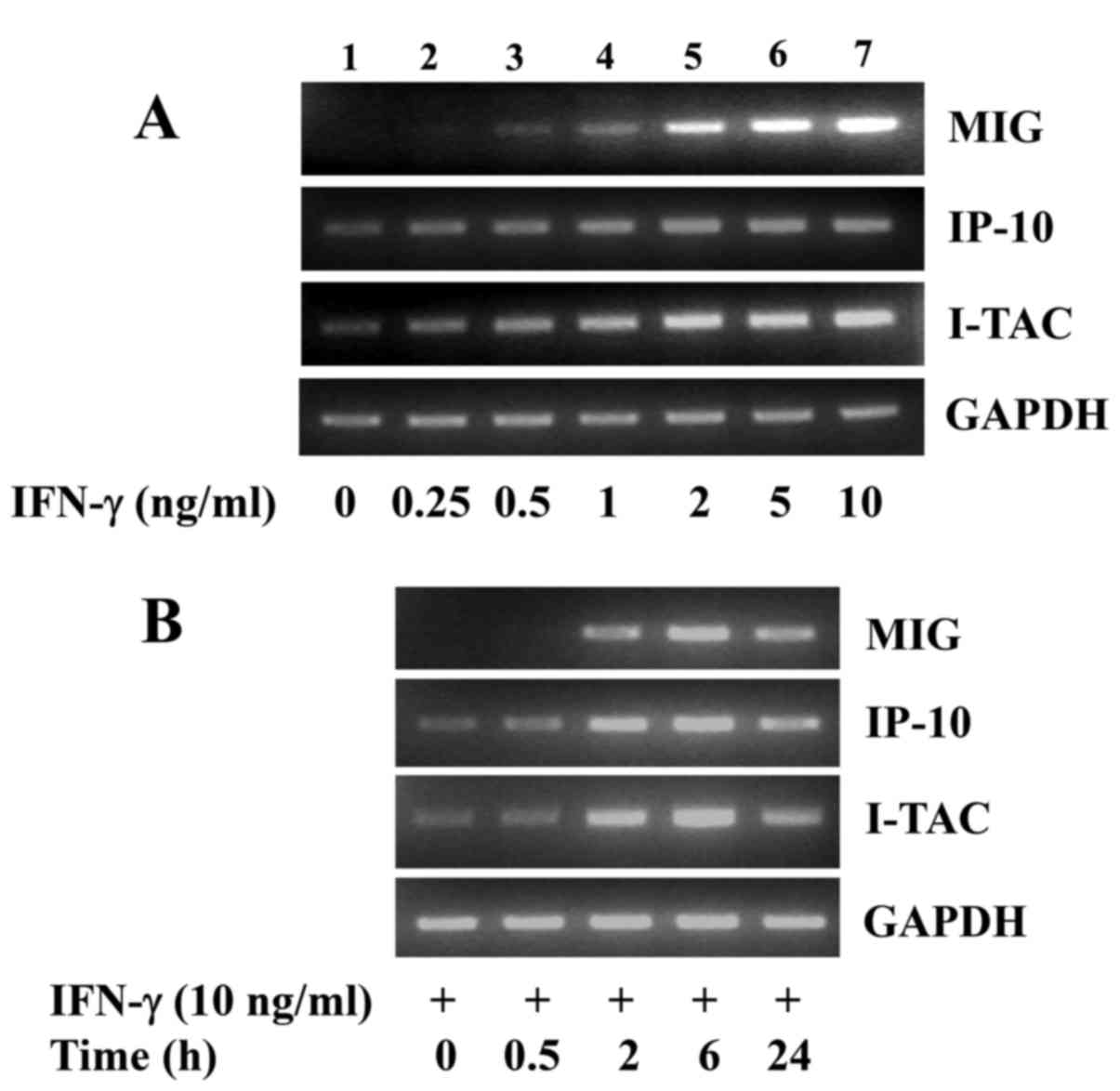
Initially, we investigated the effects of treatment
with various concentrations of IFN-γ for 6 h on the expression of
T-cell-specific chemokines, including MIG, IP-10 and I-TAC in INS-1
cells. As shown in Fig. 1A,
compared with the control (lane 1), 6 h of treatment with IFN-γ led
to a concentration-dependent increase in the mRNA levels of MIG,
IP-10 and I-TAC in the INS-1 cells (lanes 2–7). Due to the strong
induction of the MIG, IP-10 and I-TAC mRNA expression levels in
INS-1 cells, the concentration of 10 ng/ml of IFN-γ was selected
for use in further experiments. Time course experiments were then
carried out to determine the time of induction of chemokine
expression in INS-1 cells in response to IFN-γ (10 ng/ml). As shown
in Fig. 1B, treatment with IFN-γ
led to a time-dependent increase in the mRNA levels of MIG, IP-10
and I-TAC in the INS-1 cells, in which 6 h of treatment with IFN-γ
was maximal for the induction of the expression of these
chemokines. The mRNA expression of the control, GAPDH, remained
constant under these experimental conditions.
Blockage of the IFN-γ-induced expression
of MIG, IP-10 and I-TAC in INS-1 cells by SFN
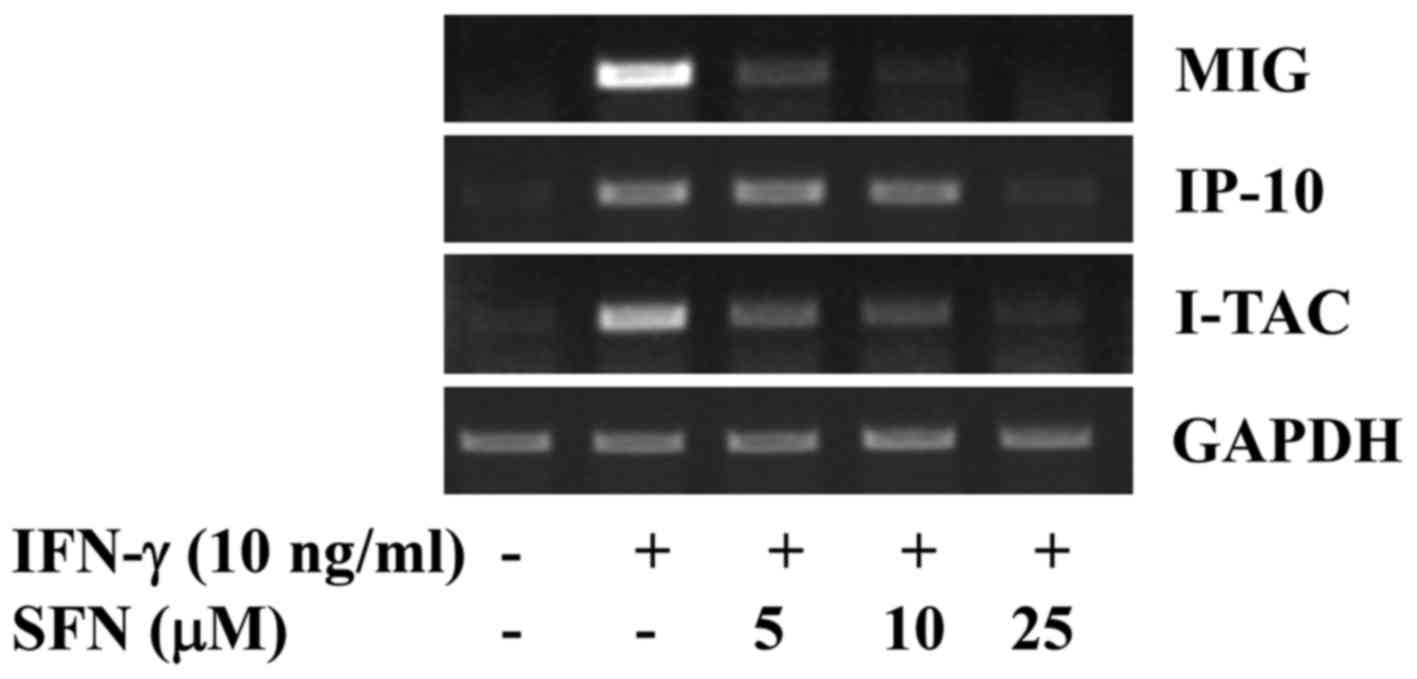
We then determined the effects of treatment with
various concentrations of SFN on the IFN-γ-induced expression of
MIG, IP-10 and I-TAC in INS-1 cells. As shown in Fig. 2, treatment with SFN resulted in a
dose-dependent blockage of the IFN-γ-induced expression of MIG,
IP-10 and I-TAC in the INS-1 cells. Of note, treatment with SFN at
25 μM was able to almost completely inhibit the
IFN-γ-induced expression of MIG, IP-10 and I-TAC. The mRNA
expression of the control, GAPDH, remained unaltered under these
experimental conditions.
Downregulation of the IFN-γ-induced mRNA
expression levels of IRF-1 and phosphorylation levels of STAT-1 and
PKB in INS-1 cells by SFN
It has been shown that IFN-γ activity is mediated
through the activation of the JAK/STAT signaling pathway, which
further triggers the activation of many downstream effectors, such
as PKB, and the activation of these signaling pathways or
components is necessary for the transcriptional induction of IFN-γ
target genes, including chemokines (18,19). There is also strong evidence to
indicate that the IFN-γ-induced expression of MIG, IP-10 and I-TAC
occurs through an STAT-1/IRF-1-dependent mechanism, which may play
an important role in the infiltration of leukocytes into tissue
(20). This promptly led us to
investigate the effects of IFN-γ and/or SFN on the activation
(phosphorylation) of STAT-1 and PKB, and on the expression of IRF-1
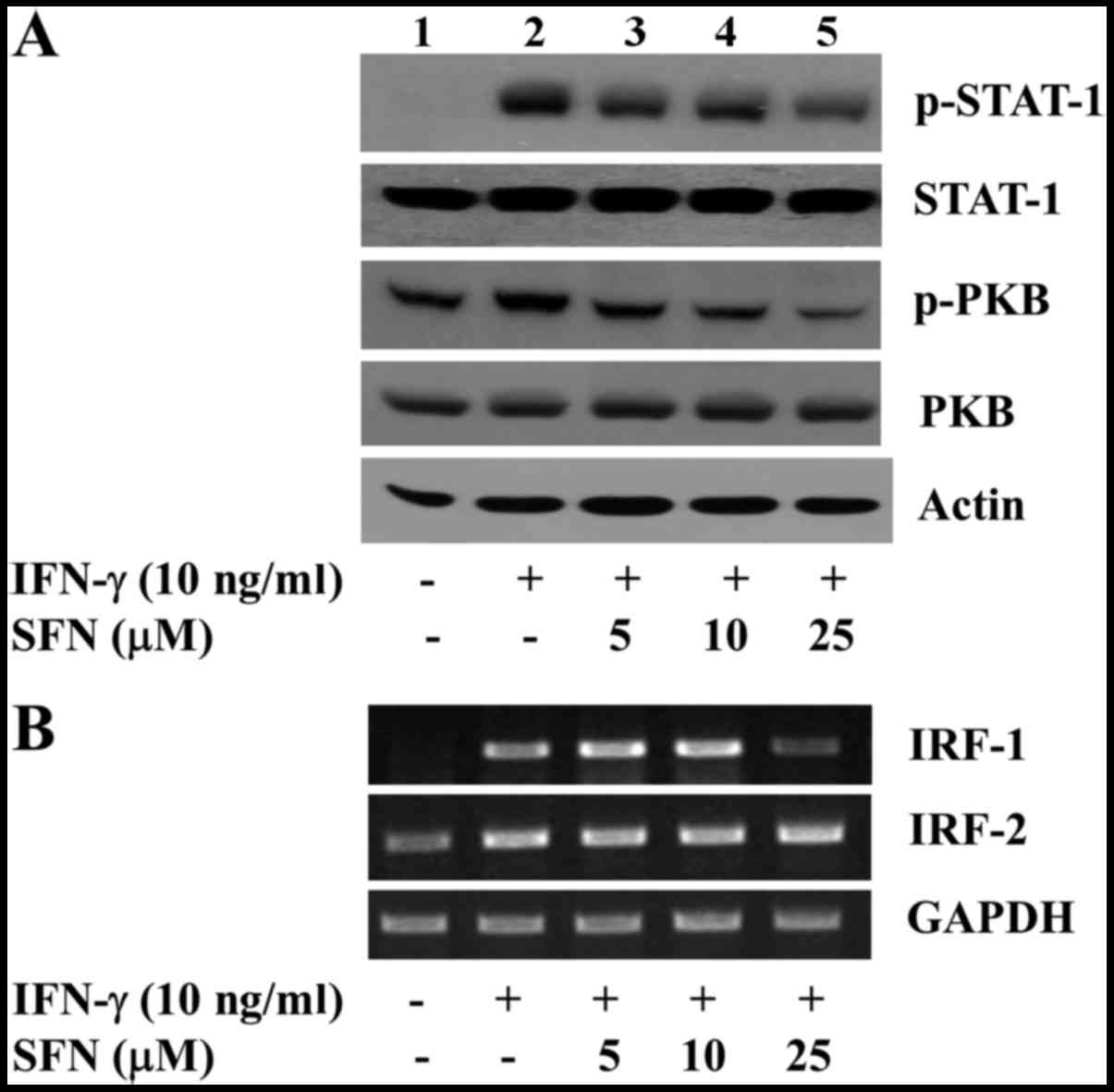
and IRF-2 in the INS-1 cells. As shown in Fig. 3A, compared with the control (lane
1), treatment with IFN-γ increased the phosphorylation levels of
both STAT-1 and PKB in the INS-1 cells (lane 2). However, treatment
with SFN led to a concentration-dependent reduction in the
phosphorylation levels of these proteins induced by IFN-γ in the
INS-1 cells (lanes 3–5). The maximal inhibition of the
IFN-γ-induced phosphorylation of STAT-1 and PKB was observed by
treatment with SFN at 25 μM. The total protein expression
levels of STAT-1 and PKB remained constant by treatment with or
without IFN-γ in the absence or presence of SFN, suggesting that
IFN-γ or SFN treatment alters the phosphorylation levels of
pre-existing STAT-1 and PKB in the INS-1 cells without leading to
de novo protein synthesis. As shown in Fig. 3B, compared with control (lane 1),
treatment with IFN-γ increased the mRNA expression levels of both
IRF-1 and IRF-2 in the INS-1 cells (lane 2). However, treatment
with SFN, particularly at 25 μM, strongly reduced the mRNA
levels of IRF-1 in the INS-1 cells (lane 5). SFN at the
concentrations tested had no effect on the mRNA expression of IRF-2
induced by IFN-γ in the cells. The mRNA expression of the control,
GAPDH, remained constant by treatment without or with IFN-γ in the
absence of presence of SFN.
Role of JAK/STAT-1 and PI3K/PKB in the
IFN-γ-induced expressions of MIG, IP-10 and I-TAC in the INS-1
cells
In order to determine the role of STAT-1 and/or PKB
in the IFN-γ-induced expression of MIG, IP-10 and I-TAC, as well as
in the suppressive effects of SFN on the IFN-γ-induced expression
of MIG, IP-10 and I-TAC in the INS-1 cells, we then performed
pharmacological inhibition experiments with the pan-JAK inhibitor,
JAK inhibitor I and the PI3K/PKB inhibitor, LY294002. As shown in
Fig. 4, treatment with JAK
inhibitor I strongly blocked the IFN-γ-induced expression of MIG
and I-TAC, but weakly inhibited the IFN-γ-induced expression of
IP-10 in the INS-1 cells. Treatment with LY294002 also slightly
reduced the mRNA levels of MIG and I-TAC, but not those of IP-10,
which were induced by IFN-γ in the INS-1 cells. The mRNA expression
of the control, GAPDH, remained constant by treatment with or
without IFN-γ in the absence of presence of JAK inhibitor I or
LY294002.
 | Figure 4Effect of interferon-γ (IFN-γ) and/or
JAK inhibitor I or LY294002 on the mRNA expression of monokine
induced by IFN-γ (MIG), IFN-inducible protein of 10 kDa (IP-10),
and IFN-inducible T cell alpha chemoattractant (I-TAC) in INS-1
cells. INS-1 cells were treated with or without IFN-γ (10 ng/ml) in
the absence or presence of JAK inhibitor I (JI, 0.1 μM), a
pan-JAK inhibitor, or LY294002 (LY, 25 μM), a PI3K/PKB
inhibitor, for 6 h. Total RNA was prepared and used for RT-PCR with
specific primers for MIG, IP-10, I-TAC or glyceraldehyde
3-phosphate dehydrogenase (GAPDH). The blots are representative of
3 independent experiments. |
Comparison of the effects of SFN and
other natural substances on the IFN-γ-induced expression of MIG,
IP-10 and I-TAC in INS-1 cells
To examine the specificity, we then compared the
effects of SFN and other natural substances, including curcumin,
sanguinarine, resveratrol, triptolide and EGCG, on IFN-γ-induced
expressions of MIG, IP-10 and I-TAC in INS-1 cells. As shown in
Fig. 5, as expected, treatment
with SFN strongly inhibited the mRNA expression levels of MIG,
IP-10 and I-TAC induced by IFN-γ in the INS-1 cells. However,
treatment with curcumin (Cur), sanguinarine (San), resveratrol
(Res), triptolide (Trip) or epigallocatechin gallate (EGCG; EG) did
not affect the IFN-γ-induced expressions of MIG, IP-10 and I-TAC in
the INS-1 cells.
 | Figure 5Effect of sulforaphane (SFN), curcumin
(Cur), sanguinarine (San), resveratrol (Res), triptolide (Trip) or
epigallocatechin gallate (EGCG; EG) on the interferon-γ
(IFN-γ)-induced expression of monokine induced by IFN-γ (MIG),
IFN-inducible protein of 10 kDa (IP-10) and IFN-inducible T cell
alpha chemoattractant (I-TAC) in INS-1 cells. INS-1 cells were
treated with or without IFN-γ (10 ng/ml) in the absence or presence
of SFN (25 μM), curcumin (10 μM), sanguinarine (500
nM), resveratrol (25 μM), triptolide (40 nM) or EGCG (25
μM) for 6 h. Total RNA was prepared and used for RT-PCR with
specific primers for MIG, IP-10, I-TAC or glyceraldehyde
3-phosphate dehydrogenase (GAPDH). The blots are representative of
3 independent experiments. |
Discussion
Evidence suggests elevated levels of CXC chemokines
in pathological conditions of the pancreas, including pancreatic
infection (21) or autoimmune
insulitis (22). The increased
expression of IP-10 and the infiltration of lymphocytes expressing
CXCR3 has been observed in all pancreatic lesions of patients with
type 1 diabetes, compared with no or low expression of IP-10 and
CXCR3 in the pancreas of non-diabetic control subjects (12). The enhanced expression of
chemokines within pancreatic islets is likely to contribute to
islet inflammation by controlling the recruitment and the
activation of macrophages, neutrophils and Th1. Thus, any compound
that can inhibit the excessive expression of chemokines in
pancreatic β-cells may have preventive and/or therapeutic potential
against islet inflammation and related diseases.
IFN-γ is a pro-inflammatory cytokine and mediates
inflammatory and immune responses. Reportedly, type I and II
interferons, including IFN-γ, are also associated with islet
inflammation and β-cell death and dysfunction (23–25). At present, the IFN-γ-mediated
induction of chemokine expression in pancreatic β-cells is not well
known. In this study, we demonstrated that treatment with IFN-γ
largely stimulated the mRNA expression levels of MIG, IP-10 and
I-TAC in INS-1 cells at the concentration of 10 ng/ml (Fig. 1). IFN-γ activity is mediated
through its cognate receptors, IFN-γR1 and IFN-γR2, which further
trigger the activation of the JAK-STAT signaling pathway (26). Upon IFN-γ binding to the IFN-γR1
and IFN-γR2, JAK1 and JAK2 associated with the receptors are
activated, leading to STAT-1 phosphorylation (on tyrosine 701) and
activation. Active STAT-1 then undergoes dimerization and the
dimeric complex translocates to the nucleus where it regulates gene
expression by binding to γ-activated sequence elements in the
promoters of IFN-γ-regulated genes, including chemokines (18,27,28). These results point out the
critical role of JAK-STAT-1 activity in the transcriptional
upregulation of chemokines in response to IFN-γ exposure. In
addition to JAK-STAT activity, the PI3K/PKB signaling pathway has
been reported to be important for IP-10 gene expression (19). In the present study, IFN-γ
treatment largely increased the phosphorylation levels of STAT-1
and PKB in the INS-1 cells (Fig.
3A, lane 2), but the blockage of JAK/STAT-1 activity by JAK
inhibitor I or PI3K/PKB activity by LY294002 strongly or weakly
abrogated the ability of IFN-γ to induce the mRNA expression of
MIG, IP-10 and I-TAC in INS-1 cells (Fig. 4, lanes 3 or 4). These results
suggest that the activation of the JAK/STAT-1 and PI3K/PKB
signaling proteins is critical for the IFN-γ-induced expression of
MIG, IP-10 and I-TAC in INS-1 cells.
SFN is an isothiocyanate substance abundantly found
in cruciferous vegetables, such as broccoli and brussel sprouts,
and has been shown to possess anti-inflammatory, anti-cancerous,
and immunomodulatory effects (13,14,29,30). Previously, the SFN-mediated
regulation of chemokines, such as CCL17 and CCL22, through heme
oxygenase-1 and NF-κB in human keratinocytes has been reported
(31). At present, neither the
inhibitory effect, nor the mechanisms of action of SFN as regards
the expression of T-cell chemokines induced by IFN-γ in pancreatic
β-cells are known. In this study, we demonstrated that SFN at the
25 μM concentration strongly inhibited not only the
IFN-γ-induced expression of MIG, IP-10 and I-TAC (Fig. 2), but also the IFN-γ-induced
activation (phosphorylation) of STAT-1 and PKB signaling proteins
(Fig. 3A, lane 5) in INS-1 cells.
Considering the positive role of STAT-1 and PKB activities in the
IFN-γ-induced expression of MIG, IP-10, and/or I-TAC in INS-1 cells
herein (Fig. 4), it is evident
that the suppressive effects of SFN on the IFN-γ-induced expression
of MIG, IP-10, and I-TAC in INS-1 cells is, at least in part,
attributable to the inhibition of STAT-1 and PKB signaling
proteins.
The activation of STAT-1 induced by IFN-γ leads to
the transcriptional induction of a number of genes, including
IRF-1, a transcription factor involved in the transcription of many
antiviral and anti-apoptotic genes (32). There is of interest recent
evidence suggesting that the transcriptional upregulation of MIG,
IP-10 and I-TAC genes occurs through an STAT-1/IRF-1-dependent
mechanism (20). The present
study revealed that IFN-γ treatment largely upregulated the
transcript levels of IRF-1 and IRF-2, while SFN treatment,
particularly at 25 μM strongly suppressed the IFN-γ-induced
mRNA expression of IRF-1, but not that of IRF-2 in the INS-1 cells
(Fig. 3B). It is thus likely that
the downregulation of the STAT-1/IRF-1 signaling pathways further
contributes to the suppressive effects of SFN on the IFN-γ-induced
expression of MIG, IP-10 and I-TAC in INS-1 cells.
An interesting finding of the present study is the
specificity of SFN in inhibiting the IFN-γ-induced expression of
MIG, IP-10 and I-TAC in INS-1 cells, as evidenced by that unlike
SFN, other natural substances with known anti-inflammatory and/or
immune modulatory activities, such as curcumin (33), sanguinarine (34), resveratrol (35), triptolide (36) or EGCG (37), had no effect on IFN-γ-induced
expressions of MIG, IP-10 and I-TAC in INS-1 cells (Fig. 5). These results indicate that SFN
may have a unique structural moiety leading to a strong inhibitory
effect on the IFN-γ-induced expression of MIG, IP-10 and I-TAC in
INS-1 cells.
In conclusion, the findings of our study
collectively demonstrate the ability of SFN to strongly inhibit the
IFN-γ-induced expression of MIG, IP-10 and I-TAC in INS-1 cells
through the reduced expression and phosphorylation levels of IRF-1,
STAT-1 and PKB. Although there are still important issues that
remain to be resolved, including the suppressive effects of SFN on
the expression of chemokines in animal models of islet
inflammation, the present findings suggest that SFN may be a
promising natural product for use against pancreatic islet
inflammation in which the overexpression of T-cell chemokines is
problematic.
Acknowledgments
This study was supported by the research promoting
grant from the Keimyung University Dongsan Medical Center in
2009.
References
|
1
|
Salgame P, Abrams JS, Clayberger C,
Goldstein H, Convit J, Modlin RL and Bloom BR: Differing lymphokine
profiles of functional subsets of human CD4 and CD8 T cell clones.
Science. 254:279–282. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Neville LF, Mathiak G and Bagasra O: The
immunobiology of interferon-gamma inducible protein 10 kD (IP-10):
A novel, pleiotropic member of the C-X-C chemokine superfamily.
Cytokine Growth Factor Rev. 8:207–219. 1997. View Article : Google Scholar
|
|
3
|
Farber JM: Mig and IP-10: CXC chemokines
that target lymphocytes. J Leukoc Biol. 61:246–257. 1997.PubMed/NCBI
|
|
4
|
Cole KE, Strick CA, Paradis TJ, Ogborne
KT, Loetscher M, Gladue RP, Lin W, Boyd JG, Moser B, Wood DE, et
al: Interferon-inducible T cell alpha chemoattractant (I-TAC): A
novel non-ELR CXC chemokine with potent activity on activated T
cells through selective high affinity binding to CXCR3. J Exp Med.
187:2009–2021. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu MT, Armstrong D, Hamilton TA and Lane
TE: Expression of Mig (monokine induced by interferon-gamma) is
important in T lymphocyte recruitment and host defense following
viral infection of the central nervous system. J Immunol.
166:1790–1795. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bonecchi R, Bianchi G, Bordignon PP,
D'Ambrosio D, Lang R, Borsatti A, Sozzani S, Allavena P, Gray PA,
Mantovani A, et al: Differential expression of chemokine receptors
and chemotactic responsiveness of type 1 T helper cells (Th1s) and
Th2s. J Exp Med. 187:129–134. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Groom JR and Luster AD: CXCR3 in T cell
function. Exp Cell Res. 317:620–631. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nakayama M, Abiru N, Moriyama H, Babaya N,
Liu E, Miao D, Yu L, Wegmann DR, Hutton JC, Elliott JF, et al:
Prime role for an insulin epitope in the development of type 1
diabetes in NOD mice. Nature. 435:220–223. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Michelsen BK, Petersen JS, Boel E, Møldrup
A, Dyrberg T and Madsen OD: Cloning, characterization, and
autoimmune recognition of rat islet glutamic acid decarboxylase in
insulin-dependent diabetes mellitus. Proc Natl Acad Sci USA.
88:8754–8758. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bonifacio E, Lampasona V, Genovese S,
Ferrari M and Bosi E: Identification of protein tyrosine
phosphatase-like IA2 (islet cell antigen 512) as the
insulin-dependent diabetes-related 37/40K autoantigen and a target
of islet-cell antibodies. J Immunol. 155:5419–5426. 1995.PubMed/NCBI
|
|
11
|
Graham KL, Krishnamurthy B, Fynch S,
Ayala-Perez R, Slattery RM, Santamaria P, Thomas HE and Kay TW:
Intra-islet proliferation of cytotoxic T lymphocytes contributes to
insulitis progression. Eur J Immunol. 42:1717–1722. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Roep BO, Kleijwegt FS, van Halteren AG,
Bonato V, Boggi U, Vendrame F, Marchetti P and Dotta F: Islet
inflammation and CXCL10 in recent-onset type 1 diabetes. Clin Exp
Immunol. 159:338–343. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Heiss E, Herhaus C, Klimo K, Bartsch H and
Gerhäuser C: Nuclear factor kappa B is a molecular target for
sulforaphane-mediated anti-inflammatory mechanisms. J Biol Chem.
276:32008–32015. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rose P, Huang Q, Ong CN and Whiteman M:
Broccoli and watercress suppress matrix metalloproteinase-9
activity and invasiveness of human MDA-MB-231 breast cancer cells.
Toxicol Appl Pharmacol. 209:105–113. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lin W, Wu RT, Wu T, Khor TO, Wang H and
Kong AN: Sulforaphane suppressed LPS-induced inflammation in mouse
peritoneal macrophages through Nrf2 dependent pathway. Biochem
Pharmacol. 76:967–973. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhao J, Moore AN, Redell JB and Dash PK:
Enhancing expression of Nrf2-driven genes protects the blood brain
barrier after brain injury. J Neurosci. 27:10240–10248. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Annabi B, Rojas-Sutterlin S, Laroche M,
Lachambre MP, Moumdjian R and Béliveau R: The diet-derived
sulforaphane inhibits matrix metalloproteinase-9-activated human
brain microvascular endothelial cell migration and tubulogenesis.
Mol Nutr Food Res. 52:692–700. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Schroder K, Hertzog PJ, Ravasi T and Hume
DA: Interferon-gamma: An overview of signals, mechanisms and
functions. J Leukoc Biol. 75:163–189. 2004. View Article : Google Scholar
|
|
19
|
Lu X, Masic A, Liu Q and Zhou Y:
Regulation of influenza A virus induced CXCL-10 gene expression
requires PI3K/Akt pathway and IRF3 transcription factor. Mol
Immunol. 48:1417–1423. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jaruga B, Hong F, Kim WH and Gao B:
IFN-gamma/STAT1 acts as a proinflammatory signal in T cell-mediated
hepatitis via induction of multiple chemokines and adhesion
molecules: A critical role of IRF-1. Am J Physiol Gastrointest
Liver Physiol. 287:G1044–G1052. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Capua I, Mercalli A, Pizzuto MS,
Romero-Tejeda A, Kasloff S, De Battisti C, Bonfante F, Patrono LV,
Vicenzi E, Zappulli V, et al: Influenza A viruses grow in human
pancreatic cells and cause pancreatitis and diabetes in an animal
model. J Virol. 87:597–610. 2013. View Article : Google Scholar :
|
|
22
|
Christen U, McGavern DB, Luster AD, von
Herrath MG and Oldstone MB: Among CXCR3 chemokines,
IFN-gamma-inducible protein of 10 kDa (CXC chemokine ligand (CXCL)
10) but not monokine induced by IFN-gamma (CXCL9) imprints a
pattern for the subsequent development of autoimmune disease. J
Immunol. 171:6838–6845. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Westwell-Roper C, Nackiewicz D, Dan M and
Ehses JA: Toll-like receptors and NLRP3 as central regulators of
pancreatic islet inflammation in type 2 diabetes. Immunol Cell
Biol. 92:314–323. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Burke SJ and Collier JJ: Insulitis and
diabetes: A perspective on islet inflammation. Immunome Res.
S2:e0022014.
|
|
25
|
Baker RG, Hayden MS and Ghosh S: NF-κB,
inflammation, and metabolic disease. Cell Metab. 13:11–22. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Darnell JE Jr, Kerr IM and Stark GR:
Jak-STAT pathways and transcriptional activation in response to
IFNs and other extracellular signaling proteins. Science.
264:1415–1421. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Aaronson DS and Horvath CM: A road map for
those who don't know JAK-STAT. Science. 296:1653–1655. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
O'Shea JJ, Gadina M and Schreiber RD:
Cytokine signaling in 2002: New surprises in the Jak/Stat pathway.
Cell. 109(Suppl): S121–S131. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Guerrero-Beltrán CE, Mukhopadhyay P,
Horváth B, Rajesh M, Tapia E, García-Torres I, Pedraza-Chaverri J
and Pacher P: Sulforaphane, a natural constituent of broccoli,
prevents cell death and inflammation in nephropathy. J Nutr
Biochem. 23:494–500. 2012. View Article : Google Scholar
|
|
30
|
Devi JR and Thangam EB: Mechanisms of
anticancer activity of sulforaphane from Brassica oleracea in HEp-2
human epithelial carcinoma cell line. Asian Pac J Cancer Prev.
13:2095–2100. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jeong SI, Choi BM and Jang SI:
Sulforaphane suppresses TARC/CCL17 and MDC/CCL22 expression through
heme oxygenase-1 and NF-κB in human keratinocytes. Arch Pharm Res.
33:1867–1876. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Saha B, Jyothi Prasanna S, Chandrasekar B
and Nandi D: Gene modulation and immunoregulatory roles of
interferon gamma. Cytokine. 50:1–14. 2010. View Article : Google Scholar
|
|
33
|
Zhong F, Chen H, Han L, Jin Y and Wang W:
Curcumin attenuates lipopolysaccharide-induced renal inflammation.
Biol Pharm Bull. 34:226–232. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Niu X, Fan T, Li W, Xing W and Huang H:
The anti-inflammatory effects of sanguinarine and its modulation of
inflammatory mediators from peritoneal macrophages. Eur J
Pharmacol. 689:262–269. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Park DW, Kim JS, Chin BR and Baek SH:
Resveratrol inhibits inflammation induced by heat-killed Listeria
monocytogenes. J Med Food. 15:788–794. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hongqin T, Xinyu L, Heng G, Lanfang X,
Yongfang W and Shasha S: Triptolide inhibits IFN-γ signaling via
the Jak/STAT pathway in HaCaT keratinocytes. Phytother Res.
25:1678–1685. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yang J, Han Y, Chen C, Sun H, He D, Guo J,
Jiang B, Zhou L and Zeng C: EGCG attenuates high glucose-induced
endothelial cell inflammation by suppression of PKC and NF-κB
signaling in human umbilical vein endothelial cells. Life Sci.
92:589–597. 2013. View Article : Google Scholar : PubMed/NCBI
|