Introduction
Gene- and cell-based hybrid treatment has received
increased attention in the past decade with continuous refinement.
In hybrid therapy, a human MSC loaded with the pacemaker gene
hyperpolarization-activated cyclic nucleotide-gated channel 2
(HCN2) is implanted in the myocardium to induce the pace-maker
function. The gene encoding transcription factor TBX18 has also
been used to engineer phenotypic and functional pacemaker cells
from myocardial cells in situ (1,2).
Stem cells possess the capacity to differentiate into numerous
cellular phenotypes and are readily accessible; therefore, they are
considered optimal candidates for cardiac pacing (3,4).
Bone mesenchymal stem cells (BMSCs) are a type of adult stem cell
that have been widely used as cytoreagents for gene therapy
(5).
BMSCs are heterogeneous cells derived from bone
marrow cavities, which have various advantages compared with other
types of stem cell. Notably, BMSCs are able to differentiate into
various cell types in vitro. Based on these characteristics,
mesenchymal stem cells can be used to deliver biological pace-maker
genes, in order to induce the formation of functional and
phenotypic pacemaker cells (6-9).
During cardiogenesis, human T-box transcription
factors, including T-Box protein 3 (TBX3), TBX5 and TBX18 serve a
prominent role in development of the sinoatrial node (SAN),
particularly TBX18, which is a gene necessary for early SAN
specification. Mesenchymal progenitor cells that express the TBX18
transcription factor are associated with generation of the SAN,
particularly formation of the head region, which not only accounts
for ~75% of SAN volume, but also produces pacemaker activity in the
early phase of embryonic heart formation (10-12). TBX18 gene knock-out mice exhibit a
marked reduction in the SAN head region (13). In addition, a proof-of-concept
study successfully converted postnatal rat ventricular myocytes to
induced pacemaker cells via TBX18 expression (14). These findings may provide
information regarding a novel method for generation of a biological
pacemaker.
To the best of our knowledge, direct evidence that
indicates the transformation of BMSCs to biological pacemaker cells
via the TBX18 transcription factor is currently lacking. In the
present study, TBX18 was expressed via adenovirus vectors to
identify whether a single transcription factor is sufficient to
induce differentiation of phenotypic and functional pace-makers.
The present study aimed to provide an increased array of treatment
options for gene and cell therapies.
Materials and methods
Animals
Adult male Sprague-Dawley rats (n=30; age, 2 months;
weight, 160-200 g) were purchased from the Center for Disease
Control and Prevention of Hubei Province (Hubei, China). The rats
were housed in micro-isolators under specific pathogen-free
conditions, in a climate-controlled enrivonment with an ambient
temperature of 24°C and a 12:12 h light/ dark cycle. They were fed
with standard laboratory chow and given sterilized water. The
present study was approved by the Experimental Animal Committee of
Wuhan University (Hubei, China). All animals received care in
accordance with the guidelines for animal care published by the
United States National Institutes of Health (NIH) (Guide for the
Care and Use of Laboratory Animals, Department of Health and Human
Services, NIH Publication no. 86-23, revised 1985).
Isolation and purification of BMSCs
BMSCs were isolated and cultured as described by
Huang et al (15) with
some modifications. In addition, all attempts were made to minimize
rat suffering. Briefly, following sacrifice, the femurs and tibias
of rats were quickly stripped, and muscle and extraossial tissue
were trimmed. A 5 ml syringe equipped with complete culture medium
[Dulbecco's modified Eagle's medium/nutrient mixture F-12
supplemented with 10% fetal bovine serum (both Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), 100 U/ml penicillin and 100
U/ml streptomycin] was inserted into the bone marrow cavity, so as
to flush target bone marrow cells into culture dishes, which were
cultured in an atmosphere containing 5% CO2 at 37°C. The
medium was initially substituted at 48 h and was then changed every
3 days. Once the cells reached 80% confluence, adherent cells were
trypsinized with 0.25% trypsin solution and passaged. Cells from
passages 3-5 were available for use in the following
experiments.
Characterization of BMSCs by flow
cytometry
BMSCs within passages 3-5 were harvested by
trypsinization, and the detached cells were resuspended in PBS.
Subsequently, approximately 1×106 cells were stained
with the following antibodies: Alexa Fluor® 647 Hamster
Anti-Rat cluster of differentiation (CD)29 (562153, 1:100),
phycoerythrin (PE)-Cy™7 Mouse Anti-Rat CD90 (561404, 1:100) and
fluorescein isothiocyanate (FITC) Mouse Anti-Rat CD45 (561867,
1:100) (all BD Biosciences, San Jose, CA, USA). Control samples
were stained with Alexa Fluor® 647-conjugated hamster
immunoglobulin (Ig)M isotype anti-body (562110, 1:100) or
PE-Cy™7-conjugated mouse IgG1 isotype antibody (557872, 1:100) and
FITC-conjugated mouse IgG1 isotype antibody (550616, 1:100) (all
from BD Biosciences). Whole incubations were performed at 4°C for
20 min. After incubation, the cells were analyzed using a
FACSCalibur flow cytometer (BD Biosciences).
Construction and purification of human
TBX18 gene adeno-virus vector
pHBAd-MCMV-GFP (HanBio Biotechnology Co., Ltd.,
Shanghai, China) was digested with BamHI and NotI.
The ORF sequence of the human TBX18 gene (GenScript,
Nanjing, China) was amplified by polymerase chain reaction (PCR).
After enzyme digestion, gel extraction was performed. The digested
fragment and vector were ligated to form pHBAd-MCMV-GFP-TBX18,
which was then transformed into competent DH5α cells (Tiangen,
Beijing, China). Positive clones were identifed by liquid
sequencing. Bacteria in liquid in the logarithmic growth phase were
incubated at 37°C in LB culture medium with shaking at 300 × g
overnight. Large scale preperation of recombinant plasmid was
conducted using the Plasmid Midi Preparation kit (Beijing CW
Biotech Co., Ltd., Beijing, China). 293 cells (from our laboratory)
were transfected with pHBAd-MCMV-GFP-TBX18 and the backbone vector
pHBAd-BHG using Lipoflter™ (both from HanBio Biotechnology Co.,
Ltd.). The supernatant was harvested after virus amplifcation.
Ad-GFP and Ad-TBX18 were measured as 1 1010 PFU/ml and
were preserved at −80°C.
Transduction of BMSCs with
hTBX18-expressing adeno-virus vector
About (5-8)x105 BMSCs were infected
with pHBAd-MCMV-GFP-TBX18 or the pHBAd-MCMV-GFP empty vector at a
multiplicity of infection (MOI) of 20, 50, 80 and 100 for 2 h at
37°C, after which the medium was replaced with complete culture
medium. Transduction efficiency was estimated according to the
proportion of GFP-positive cells. After 24 and 48 h, inverted
fluorescence microscopy (IX51; Olympus Corporation, Tokyo, Japan)
was used to detect GFP expression. A total of 2 days postinfection,
cells were accumulated for evaluation of hTBX18 expression by
western blotting and reverse transcription-quantitative PCR
(RT-qPCR).
Total RNA isolation and RT-qPCR
Total cellular RNA was extracted from BMSCs using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. Subsequently, RT was
conducted using the PrimeScript™ RT reagent kit (Takara
Biotechnology, Ltd., Dalian, China) in a 20 µl mixture.
RT-qPCR was performed using SYBR® Premix Ex Taq™kit
(Takara Biotechnology, Ltd.) as follows: 95°C for 1 min followed by
40 cycles of 95°C for 15 sec, 58°C for 20 sec and 72°C for 45 sec
and a final extension at 60°C for 10 min. GAPDH was used as a
reference gene. The results of RT-qPCR were analyzed using the
2−ΔΔCq method (16).
To ensure accuracy, each sample was analyzed ≥3 times. The primers
used in this experiment are presented in Table I.
 | Table IPrimer sequences used in the
study. |
Table I
Primer sequences used in the
study.
| Gene | Primer sequences
(5′-3′) | Accession no. | Size (bp) |
|---|
| R-α-actin | Forward:
GAGCACGGCATTATCACCAAC | NM_019183.1 | 246 |
| Reverse:
CAGAACAATGCCTGTGGTTCTC | | |
| R-HCN4 | Forward:
CACTAAGGGCAACAAGGAGACC | NM_021658.1 | 281 |
| Reverse:
GGTAGTTGAAGACGCCTGAGTTG | | |
| R-CX43 | Forward:
GCTGGTGGTGTCCTTGGTGT | NM_012567.2 | 213 |
| Reverse:
GGAGGAGACATAGGCGAGAGTG | | |
| R-GAPDH | Forward:
CGCTAACATCAAATGGGGTG | NM_017008.3 | 201 |
| Reverse:
TTGCTGACAATCTTGAGGGAG | | |
| H-TBX18 | Forward:
ACGTCATCCGTAAAGACTGTGG | NM_001080508.2 | 251 |
| Reverse:
AGTCCGTAGTGATGGTCGCC | | |
| H-GAPDH | Forward:
GGTCGGAGTCAACGGATTTG | NM_002046.3 | 218 |
| Reverse:
GGAAGATGGTGATGGGATTTC | | |
Western blot analysis
The cells were harvested using RIPA lysis buffer
(Beyotime Institute of Biotechnology, Haimen, China). Equal amounts
of protein (40 µg) were loaded onto a gel for 15% sodium
dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE), and
the seperated proteins were transferred to a nitrocellulose
membrane, and were subsequently incubated with primary antibodies
overnight at 4°C. Goat-anti-TBX18 polyclonal primary antibody
(sc-17869, 1:200; Santa Cruz Biotechnology, Inc., Dallas, TX, USA)
was applied to detect TBX18 protein expression 48 h
post-trans-duction. Rabbit anti-α-actin monoclonal antibody
(ab156302, 1:3,000), mouse anti-cardiac troponin I (cTnI)
monoclonal antibody (ab19615, 1:1,000), rat
anti-hyperpolarization-activated cyclic nucleotide-gated channel 4
(HCN4) monoclonal antibody (ab32675, 1:500) and rabbit
anti-connexin 43 (CX43) polyclonal antibody (ab11370, 1:2,000) (all
Abcam, Cambridge, MA, USA) were applied to detect biological
pacemaker induction of BMSCs after 1 week of culture. After
washing, the membranes were incubated with 1:10,000 horseradish
peroxidase (HRP) -rabbit anti-goat (14-13-06), HRP-goat anti-rabbit
(074-1506), anti-rat (14-16-06) or anti-mouse IgG (074-1806) (all
KPL, Inc., Gaithersburg, MD, USA) secondary antibodies for 30 min
at room temperature. Enhanced chemiluminescence detection (ECL;
Beyotime Institute of Biotechnology) was then performed. GAPDH was
used as a loading control. All western blot analyses were repeated
>3 times.
Immunofluorescence staining
A total of 1 week post-transduction,
immunofluorescence staining was performed on cultured cells.
Briefly, approximately (5-8)x105
cells grown on glass cover-slips were fixed with 4%
paraformaldehyde for 20 min at room temperature. Following
permeabilization with 0.1% Triton X-100, cells were incubated
overnight with rat anti-HCN4 monoclonal primary antibody (ab32675,
1:50; Abcam) at 4°C and were then incubated with Cy3-labeled goat
anti-rat secondary antibody (AS-1111, 1:50; Aspen Biotechnology,
Hubei, China) for 50 min at room temperature. Nuclei were also
stained with 4′,6-diamidino-2-phenylindole for 5 min. Fluorescent
images were acquired using an inverted fluorescent microscopy.
Statistical analysis
The reported data are expressed as the means ± SD.
The statistical signifcance of the differences between two groups
was determined using the Student's t-test. Comparisons among 3
groups were made using one-way analysis of variance (ANOVA). A
P-value <0.05 was considered to indicate a statistically
significant difference.
Results
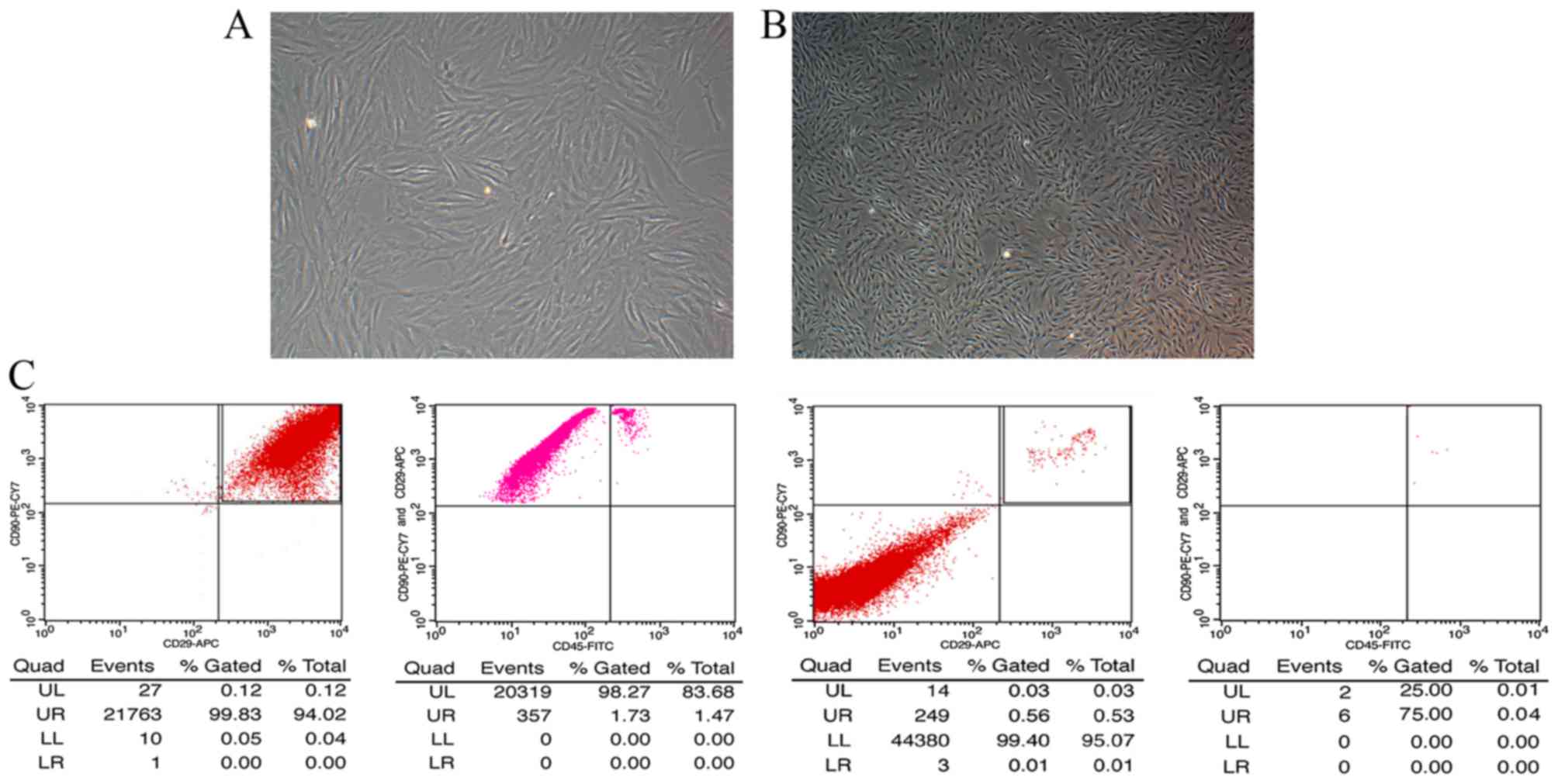
Morphological characteristics and
identification of BMSCs
When initially seeded, BMSCs appeared round in
shape. After 48 h, cells became adherent, and exhibited triangular,
fusiform or spindle shapes. These colonies fused after 6-8 days of
culture; the majority of cultured cells exhibited uniform
spindle-like morphology. As the number of passages increased,
sub-cultured cells were more purified, homogenous and spindle-like
(Fig. 1A and B).
Flow cytometric analysis
To identify BMSC phenotypes, flow cytometry was
conducted. The results indicated that BMSCs expressed stromal cell
markers CD29 and CD90 (≤99%); however, almost no expression of the
hematopoietic marker, CD45, was detected (Fig. 1C). These findings suggested that
the cells tested were purified BMSCs.
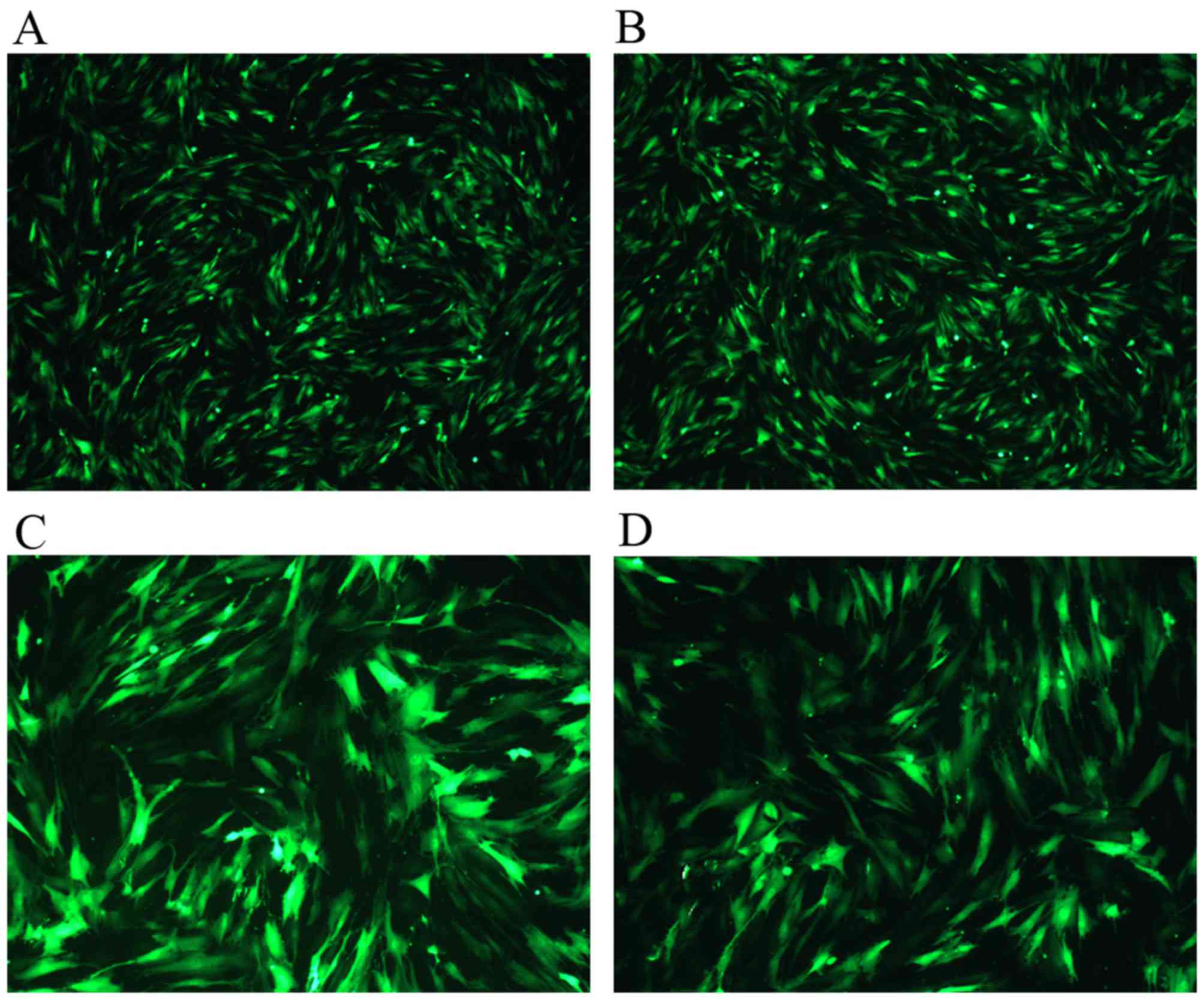
Transduction of BMSCs with
pHBAd-MCMV-GFP-TBX18 or pHBAd-MCMV-GFP
Through continuous attempts, the present study
confirmed that the appropriate MOI was 100, which was the lowest
adenovirus titer that generated optimal infection efficiency.
Post-transduction for 24 h, green fluorescence could be detected
under fluorescent microscopy. Under this condition, the
transduction efficiency was 97-99% (Fig. 2A and B). The expression levels and
high infection efficiency peaked at 48 h and then faded gradually
(Fig. 2C and D). These findings
confirmed that the plasmids had been successfully transduced into
the BMSCs.
Identification of target protein
expression by western blotting and RT-qPCR
A total of 48 h after lipofection, transgene
expression was examined by RT-qPCR and western blot-ting. The
protein expression levels of TBX18 in BMSCs are presented in
Fig. 3A; the results indicated
that the transduced cells successfully expressed TBX18 protein. The
protein expression levels of TBX18 were normalized to GAPDH
(Fig. 3B). The results of RT-qPCR
detection of TBX18 mRNA, relative to GAPDH, are presented in
Fig. 3C. These results indicated
that pHBAd-MCMV-GFP-TBX18 was successfully introduced into
BMSCs.
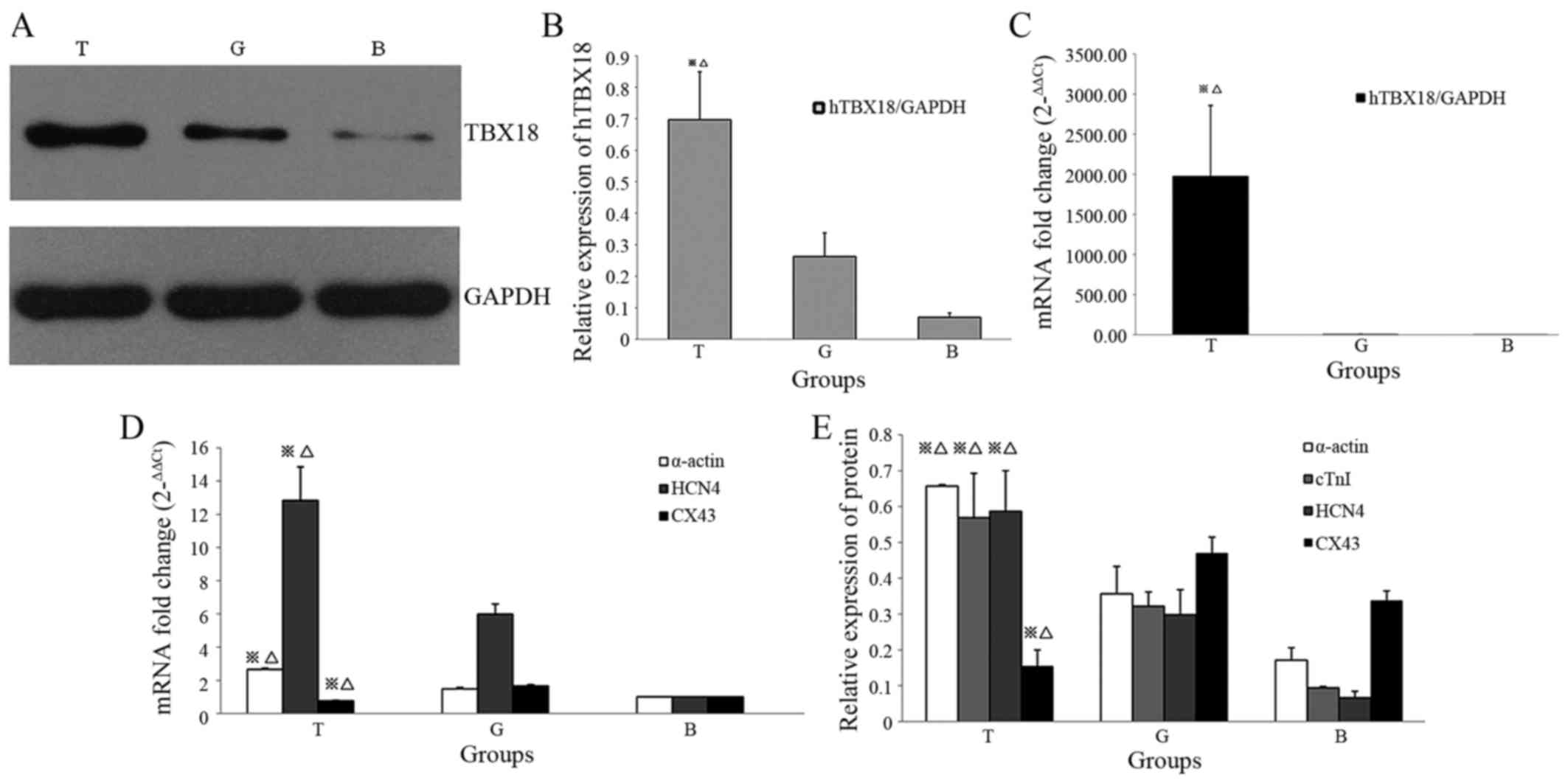 | Figure 3Western blotting and reverse
transcription-quantitative polymerase chain reaction analysis of
target protein and mRNA expression post-transduction. (A)
pHBAd-MCMV-GFP-TBX18-transduced BMSCs displayed exogenous TBX18
expression, as determined by western blotting. (B) Protein
expression levels of TBX18 were verified in differentiated cells by
western blotting. (C) Alterations in the relative mRNA expression
levels of TBX18 were compared between the T group and the G and B
groups. (D) Quantitative analysis of the mRNA expression levels of
α-actin, HCN4 and CX43. (E) Western blotting detected increased
α-actin, cTnI and HCN4 protein expression, and reduced CX43 protein
expression, in the T group compared with in the G and B groups.
※P<0.05, T group vs. G group; △P<0.05,
T group vs. B group. BMSCs, bone mesenchymal stem cells; B, blank
group; cTnI, cadiac troponin I; CX43, connexin 43; G, GFP
group/empty plasmid group; GFP, green fluorescent protein; HCN4,
hyperpolarization-activated cyclic nucleotide-gated channel 4;
hTBX18, human TBX18; T, TBX18 group; TBX18, T-box protein 18. |
RT-qPCR analysis of genes associated with
cardiac pacing in BMSCs
The expression levels of target genes associated
with biological pacemaker development were detected in BMSCs
infected with pHBAd-MCMV-GFP-TBX18. The genes detected in the
present study included α-actin, HCN4 and CX43. As shown in Fig. 3D, the mRNA expression levels of
α-actin and HCN4 were significantly increased in BMSCs infected
with TBX18 adenovirus (T group) compared with in the empty plasmid
group (G group) and the blank group (B group) (P<0.05).
Conversely, the expression levels of CX43 were significantly lower
in the T group compared with in the G and B groups (P<0.05). No
significant differences were detected between the G and B
groups.
Western blot analysis of target protein
expression
A total of 1 week post-transduction, total proteins
were extracted from cells in the T, G and B groups. Western
blotting was used to detect the protein expression levels of
α-actin, cTnI, HCN4 and CX43. The results indicated that α-actin,
cTnI and HCN4 expression levels in the T group were significantly
upregulated compared with in the G and B groups (P<0.05).
Conversely, CX43 expression was significantly downregulated in the
T group compared with in the G and B groups (P<0.05; Fig. 3E). No significant differences were
detected between the G and B groups.
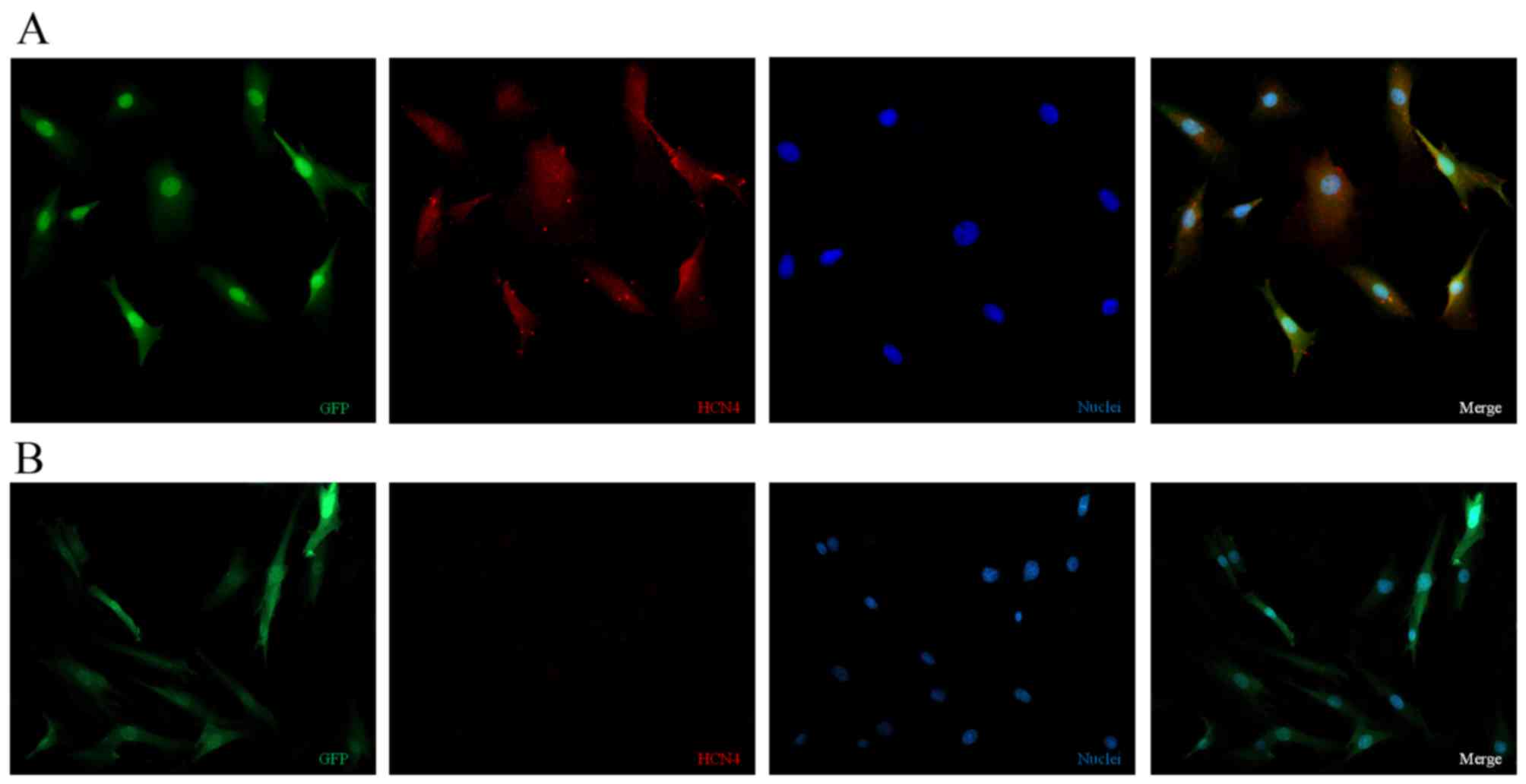
Immunocytochemical analysis of BMSCs
following transgene induction
After being cultured for 1 week, some
differenti-ated cells developed the biological pacemaker phenotype.
As demonstrated in Fig. 4A,
TBX18-transduced BMSCs co-expressed HCN4 (red) and GFP (green)
in vitro, indicating that the TBX18 adenovirus vector
induced biological pacemaker differentiation of BMSCs. However, in
the GFP-transduced group HCN4 protein expression (red) was barely
detectable (Fig. 4B).
Discussion
As a consequence of the advanced progress in the
field of molecular biology, gene reprogramming techniques have been
used in various aspects of biology. BMSCs exhibit numerous
differentiative abilities in vivo and in vitro
(17,18). Notably, mesenchymal stem cells can
inhibit T-cell proliferation, so as to restrain immunoreactivity of
the host (19). BMSCs loaded with
the biological pacemaker genes HCN4 or HCN2 have been successfully
implanted into the myocardium of large animals to induce pacemaker
function (20-23).
BMSCs are easy to modify at the genetic level. The
present study introduced the TBX18 transcription factor into BMSCs
to determine whether this gene is able to accomplish the direct
conversion of BMSCs to biological pacemaker cells. TBX18 can not
only induce the direct conversion of neonatal rat ventricular
myocytes to pacemaker cells, but also produces stable pacemaker
activity for ~2 weeks via injection of TBX18 adenovirus in pigs,
along with complete heart block (24). These findings suggested that TBX18
may be an optimal candidate gene in the biological pacemaker
process. In the present study, on the 7th day of culture, positive
results were detected, and the outcomes were evaluated with western
blotting, RT-qPCR and immunofluorescence.
To identify whether TBX18 may induce the myocardial
phenotype of BMSCs, the cells were incubated with antibodies
against cardiomyocyte-associated proteins, including cTnI and
α-actin, which are myocardium-specific markers. cTnI is one of
three subunits of troponin (cTnI, cTnT and cTnC), which is located
in the thin filaments (25). cTnI
inhibits the connection between myosin and actin via intracellular
Ca2+ to regulate muscle contraction and relaxation;
mutations in cTnI result in hypertrophic cardiomyopathy (26). α-actin is known as cardiac
structural gene, which is expressed in mouse cardiac tissue during
embryogenesis and can provide redundant function (27). In the present study, western
blotting and RT-qPCR detected cTnI and α-actin expression in
TBX18-transduced BMSCs, which is consistent with the features of
the SAN.
Notably, HCN4 expression was detected in the induced
cells in the present study. The SAN is located in the right atrium
adjacent to the superior vena cava, and is responsible for
heartbeat initiation (28). HCN4
is an important HCN isoform, which accounts for >80% of the
total amount of HCN mRNA (29,30). Furthermore, HCN4 expression is the
most typical characteristic of pace-maker cells. Furthermore, HCN4
expression is the most typical characteristic of pacemaker cells.
In the present study, HCN4 expression was detected through western
blotting, RT-qPCR and immunofluorescence. The present study
demonstrated that the expression levels of cTnI, α-actin and HCN4
were significantly increased in TBX18-transduced cells. However,
CX43 expression levels were markedly downregulated in the
TBX18-transduced groups compared with in the GFP-transduced and
blank groups. These findings were consistent with those of Kapoor
et al (31). TBX18 may
repress the CX43 promoter directly, further to the transcript and
protein levels, other connexins like CX30.2, CX40 and CX45 promotes
the slow action potential propagation. The characteristics of the
TBX18-transduced BMSCs were similar to the functional features of
the SAN, which include slow contractibility and slow-propagating
connexins compared with the surrounding myocardium.
For high-efficiency transduction, adenovirus vectors
were selected as ideal gene therapy vehicles in the present study.
In previous studies, adenovirus vectors have been used for a large
amount of transgene therapies, due to their ease of production,
high titers, stable inheritance, sufficient transduction efficiency
and early peak expression compared with other vectors (32-35). However, adenovirus-mediated gene
expression may not last a long period of time; peaking at ~1 week,
declining at 3-5 weeks and vanishing at ~10 weeks (36).
Consequently, adenovirus vectors are appropriate for temporary
applications that have no demand for long-term gene expression. In
addition, adenovirus vectors are likely to induce inflammatory and
immune responses; therefore, their permanent safety and efficacy
remains to be improved.
In conclusion, the present study indicated that
expression of the transcription factor TBX18 is capable of
initiating differentiation of BMSCs into biological pacemaker
cells. This discovery represents a possible therapeutic alternative
to electronic devices. However, long-term practice in large animals
is required to evaluate the safety prior to application in patients
with sinus dysfunction.
Acknowledgments
Flow cytometry conducted in the present study was
supported by Central Laboratory, Renmin Hospital, Wuhan University
(Wuhan, China). The present study was supported by the Fundamental
Research Funds for the Central Universities of China (grant nos.
2042015kf0229 and 2042014 kf0306).
References
|
1
|
Rosen MR: Gene therapy and biological
pacing. N Engl J Med. 371:1158–1159. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chauveau S, Brink PR and Cohen IS: Stem
cell-based biological pacemakers from proof of principle to
therapy: A review. Cytotherapy. 16:873–880. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cai B, Li J, Wang J, Luo X, Ai J, Liu Y,
Wang N, Liang H, Zhang M, Chen N, et al: microRNA-124 regulates
cardiomyocyte differentiation of bone marrow-derived mesenchymal
stem cells via targeting STAT3 signaling. Stem Cells. 30:1746–1755.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Psaltis PJ, Zannettino AC, Worthley SG and
Gronthos S: Concise review: mesenchymal stromal cells: potential
for cardiovascular repair. Stem Cells. 26:2201–2210. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hamada H, Kobune M, Nakamura K, Kawano Y,
Kato K, Honmou O, Houkin K, Matsunaga T and Niitsu Y: Mesenchymal
stem cells (MSC) as therapeutic cytoreagents for gene therapy.
Cancer Sci. 96:149–156. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou YF, Yang XJ, Li HX, Han LH and Jiang
WP: Mesenchymal stem cells transfected with HCN2 genes by LentiV
can be modified to be cardiac pacemaker cells. Med Hypotheses.
69:1093–1097. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yang XJ, Zhou YF, Li HX, Han LH and Jiang
WP: Mesenchymal stem cells as a gene delivery system to create
biological pacemaker cells in vitro. J Int Med Res. 36:1049–1055.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ma J, Zhang C, Huang S, Wang G and Quan X:
Use of rats mesenchymal stem cells modified with mHCN2 gene to
create biologic pacemakers. J Huazhong Univ Sci Technolog Med Sci.
30:447–452. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nong Y, Zhang C, Wei L, Zhang Z, Cheng J,
Wen L and Song Z: In situ investigation of allografted mouse HCN4
gene-transfected rat bone marrow mesenchymal stromal cells with the
use of patch-clamp recording of ventricular slices. Cytotherapy.
15:905–919. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wiese C, Grieskamp T, Airik R, Mommersteeg
MT, Gardiwal A, de Gier-de Vries C, Schuster-Gossler K, Moorman AF,
Kispert A and Christoffels VM: Formation of the sinus node head and
differentiation of sinus node myocardium are independently
regulated by Tbx18 and Tbx3. Circ Res. 104:388–397. 2009.
View Article : Google Scholar
|
|
11
|
McNally EM and Svensson EC: Setting the
pace: Tbx3 and Tbx18 in cardiac conduction system development. Circ
Res. 104:285–287. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cho HC: Pacing the heart with genes:
Recent progress in biological pacing. Curr Cardiol Rep. 17:652015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Greulich F, Trowe MO, Leffler A, Stoetzer
C, Farin HF and Kispert A: Misexpression of Tbx18 in cardiac
chambers of fetal mice interferes with chamber-specific
developmental programs but does not induce a pacemaker-like gene
signature. J Mol Cell Cardiol. 97:140–149. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kapoor N, Liang W, Marbán E and Cho HC:
Direct conversion of quiescent cardiomyocytes to pacemaker cells by
expression of Tbx18. Nat Biotechnol. 31:54–62. 2013. View Article : Google Scholar
|
|
15
|
Huang XP, Sun Z, Miyagi Y, McDonald
Kinkaid H, Zhang L, Weisel RD and Li RK: Differentiation of
allogeneic mesenchymal stem cells induces immunogenicity and limits
their long-term benefits for myocardial repair. Circulation.
122:2419–2429. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)). Method Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
17
|
Karaoz E, Aksoy A, Ayhan S, Sariboyaci AE,
Kaymaz F and Kasap M: Characterization of mesenchymal stem cells
from rat bone marrow: Ultrastructural properties, differentiation
potential and immunophenotypic markers. Histochem Cell Biol.
132:533–546. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ye NS, Chen J, Luo GA, Zhang RL, Zhao YF
and Wang YM: Proteomic profiling of rat bone marrow mesenchymal
stem cells induced by 5-azacytidine. Stem Cells Dev. 15:665–676.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tokcaer-Keskin Z, Akar AR, Ayaloglu-Butun
F, Terzioglu-Kara E, Durdu S, Ozyurda U, Ugur M and Akcali KC:
Timing of induction of cardiomyocyte differentiation for in vitro
cultured mesenchymal stem cells: A perspective for emergencies. Can
J Physiol Pharmacol. 87:143–150. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Potapova I, Plotnikov A, Lu Z, Danilo P
Jr, Valiunas V, Qu J, Doronin S, Zuckerman J, Shlapakova IN, Gao J,
et al: Human mesenchymal stem cells as a gene delivery system to
create cardiac pacemakers. Circ Res. 94:952–959. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Plotnikov AN, Shlapakova I, Szabolcs MJ,
Danilo P Jr, Lorell BH, Potapova IA, Lu Z, Rosen AB, Mathias RT,
Brink PR, et al: Xenografted adult human mesenchymal stem cells
provide a platform for sustained biological pacemaker function in
canine heart. Circulation. 116:706–713. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cheng J, Zhang Z, Wei L, Nong Y, Wen L,
Qin Y and Song Z: Canine bone marrow mesenchymal stromal cells with
lentiviral mHCN4 gene transfer create cardiac pacemakers.
Cytotherapy. 14:529–539. 2012. View Article : Google Scholar
|
|
23
|
Wei L, Nong Y, Boli R, Cheng J, Zhang C,
Zhou Y and Song Z: mHCN4 genetically modified canine mesenchymal
stem cells provide biological pacemaking function in complete dogs
with atrioventricular block. Pacing Clin Electrophysiol.
36:1138–1149. 2013. View Article : Google Scholar
|
|
24
|
Hu YF, Dawkins JF, Cho HC, Marbán E and
Cingolani E: Biological pacemaker created by minimally invasive
somatic reprogramming in pigs with complete heart block. Sci Transl
Med. 6:245ra942014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Takahashi-Yanaga F, Morimoto S, Harada K,
Minakami R, Shiraishi F, Ohta M, Lu QW, Sasaguri T and Ohtsuki I:
Functional consequences of the mutations in human cardiac troponin
I gene found in familial hypertrophic cardiomyopathy. J Mol Cell
Cardiol. 33:2095–2107. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ruan Z, Zhu L, Yin Y and Chen G:
Overexpressing NKx2.5 increases the differentiation of human
umbilical cord drived mesenchymal stem cells into
cardiomyocyte-like cells. Biomed Pharmacother. 78:110–115. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu M, Wani M, Dai YS, Wang J, Yan M, Ayub
A and Ashraf M: Differentiation of bone marrow stromal cells into
the cardiac phenotype requires intercellular communication with
myocytes. Circulation. 110:2658–2665. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Husse B and Franz WM: Generation of
cardiac pacemaker cells by programming and differentiation. Biochim
Biophys Acta. 1948–1952.2016:1863PubMed/NCBI
|
|
29
|
Robinson RB and Siegelbaum SA:
Hyperpolarization-activated cation currents: From molecules to
physiological function. Annu Rev Physiol. 65:453–480. 2003.
View Article : Google Scholar
|
|
30
|
Verkerk AO and Wilders R: Pacemaker
activity of the human sinoatrial node: Effects of HCN4 mutations on
the hyperpolarization-activated current. Europace. 16:384–395.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kapoor N, Galang G, Marbán E and Cho HC:
Transcriptional suppression of connexin43 by TBX18 undermines
cell-cell electrical coupling in postnatal cardiomyocytes. J Biol
Chem. 286:14073–14079. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
French BA, Mazur W, Geske RS and Bolli R:
Direct in vivo gene transfer into porcine myocardium using
replication-deficient adenoviral vectors. Circulation.
90:2414–2424. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Brenner M: Gene transfer by adenovectors.
Blood. 94:3965–3967. 1999.PubMed/NCBI
|
|
34
|
Wright MJ, Wightman LM, Lilley C, de Alwis
M, Hart SL, Miller A, Coffin RS, Thrasher A, Latchman DS and Marber
MS: In vivo myocardial gene transfer: Optimization, evaluation and
direct comparison of gene transfer vectors. Basic Res Cardiol.
96:227–236. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li RA: Gene- and cell-based bio-artificial
pacemaker: What basic and translational lessons have we learned?
Gene Ther. 19:588–595. 2012. View Article : Google Scholar : PubMed/NCBI
|