Introduction
Erythropoietin (Epo) has been established to be an
important growth factor that can promote the recruitment
mesenchymal stem cells (MSCs) and angiogenesis. Previously, studies
have suggested that Epo can trigger bone formation from MSCs. Epo
can improve osteogenic differentiation of MSCs in various ways,
including by increasing the expression of vascular endothelial
growth factor and bone morphogenetic protein 2 (1), and regulating receptor activator of
nuclear factor-κB ligand signaling (2). In vivo, MSCs induced by Epo
can facilitate enhanced bone regeneration in a rat alveolar bone
defect model and a cranial defect model (3,4).
Periodontitis is a common inflammatory bone disease
that can lead to periodontal tissue destruction and tooth loss
(5,6). To date, conventional therapies have
succeeded in controlling periodontal inflammation but have failed
to restore damage of periodontal tissues (7). Previously, tissue engineering based
on MSCs has been reported to be an effective approach for
periodontal regeneration (8).
Human periodontal ligament tissue-derived mesenchymal stem cells
(hPDLSCs) are one type of MSCs that can be isolated from
periodontal ligament tissue. These cells have multi-directional
differentiation capacity and can differentiate into various tissue
types, including bone, cartilage, fat and nerves (9-13).
Because of the tissue of origin and multi-lineage potential of
hPDLSCs, these cells are considered to be a promising cell line for
achieving alveolar bone regeneration. Previous studies have
established that combining hPDLSCs and biomaterials can achieve
partial periodontal regeneration by forming cementum/periodontal
ligament-like structures (14-16).
Considering that periodontal hard tissue
regeneration is generally the aim of the periodontitis treatment
and that periodontal regeneration is difficult to achieve,
osteogenic differentiation of hPDLSCs is potentially very
important. Various factors can impact the osteogenesis of hPDLSCs,
among which inflammation is one of the most researched causative
factors (17–22). Studies have established that
inflammation can inhibit osteogenic differentiation potential via
the canonical Wnt and p38 mitogen-activated protein kinase (MAPK)
pathways (21,23).
The findings for the current study indicate that Epo
can enhance the osteogenesis and proliferation of hPDLSCs and
periodontitis mesenchymal stem cells (pPDLSCs). To further
investigate the mechanism of these processes, the p38 MAPK pathway
was focused on, which has been previously been demonstrated to be
important in osteogenic differentiation of hPDLSCs and pPDLSCs
(23). Furthermore, previous
studies reported that p38 MAPK signaling is activated by Epo
(24-26). In the current study, it was
demonstrated that Epo can regulate the osteogenic differentiation
of hPDLSCs and pPDLSCs via activating the MAPK pathway. When the
p38 MAPK signaling was inhibited, the positive effects of Epo on
osteogenesis were attenuated. Overall, the findings demonstrate
that Epo can enhance bone formation in hPDLSCs and pPDLSCs via the
p38 MAPK pathway.
Materials and methods
Cell culture
Primary hPDLSC cultures were obtained from 10
individuals, 5 male and 5 female, aged 35-45 years, undergoing
routine premolar procedures for orthodontic reasons or third molar
extractions. For every single experiment, cells from at least 3
different individuals were tested. The pPDLSCs were obtained from 7
individuals, 4 male and 3 female, aged 27-52 years, who were
diagnosed with stable phase periodontitis with two-thirds alveolar
bone destruction or at least one periodontal pocket (depth, >5
mm). For every single experiment, cells from at least 3 different
individuals were tested. None of these selected subjects had recent
periodontal infection or systemic disease, a history of smoking or
histories of maxillofacial surgery, radiotherapy or chemo-therapy.
All samples were collected at the Dental Clinic of the Fourth
Military Medical University (Xi'an, China). Each participant
provided written informed consent, and the study was approved by
the Hospital's Ethics Committee (license no. IRB-REV-2015038). The
tissue were obtained from the periodontal ligament of the root
surface and using type 1 collagenase digestion (0.66 mg/ml;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at 37°C for >20
min. Single-cell suspensions were cultured in α-minimum essential
medium (α-MEM) (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo
Fisher Scientific, Inc.), glutamine, penicillin and streptomycin.
At passage 3, hPDLSCs and pPDLSCs were isolated using
immunomagnetic beads (M2450; Dynal Biotech, Wirral, UK) with STRO-1
antibodies (340106; Biolegend, San Diego, CA, USA), as previously
described (19). Cells at
passages 3-5 were used in this study.
Epo treatment
hPDLSCs and pPDLSCs at passage 3
(1×105/well) were cultured in α-MEM (10% FBS) until they
reached 80% confluence. Then, cells were incubated with Epo medium
(Epo medium contains 20 IU/ml Epo in α-MEM with 10% FBS) and the
medium was changed in 2 days. Subsequently, the cells were cultured
in osteogenic-inducing media for 7 or 21 days to induce osteogenic
differentiation.
SB203580 treatment
HPDLSCs and pPDLSCs at passage 3
(1×105/well) were cultured in α-MEM (10% FBS) until they
reached 80% confluence. Then, cells were incubated with SB203580
medium (SB203580 medium contained 10 µmol/l SB203580 in
α-MEM with 10% FBS) and the medium was changed in 2 days. Then the
cells were cultured in osteogenic-inducing media for 7 or 21 days
to induce osteogenic differentiation
Osteogenic differentiation
Cells (1×105/well) were cultured were
cultured until they reached 80% of the culture flask. Then, media
were changed with osteogenic medium (100 nM dexamethasone, 50 mg/ml
ascorbic acid, and 5 mM β-glycerophosphate; Sigma-Aldrich; Merck
KGaA) cells were cultured for 7 or 21 days. The alkaline
phosphatase (ALP) activity assay was performed following osteogenic
induction for 7 days using an ALP kit according to the
manufacturer's instructions (Nanjing Jiancheng Bioengineering
Institute, Nanjing, China). ALP staining was performed using a
BCIP/NBT ALP Color Development kit according to the manufacturer's
instructions (Beyotime Institute of Biotechnology, Haimen, China).
Following inducing in osteogenic medium for 21 days, Alizarin Red
staining was performed. Cells were washed with 10% FBS in PBS
twice. Then cells were fixed with 60% isopropanol for 1 min.
Subsequently, cells were washed with distilled water for 3 min and
stained using 1% Alizarin Red (Sigma-Aldrich; Merck KGaA) at room
temperature for 30 min. The Alizarin Red-stained nodules were
visualized under an Olympus BX51 light microscope equipped with an
Olympus DP70 camera (Olympus, Co., Tokyo, Japan). To quantify
Alizarin staining, mineralized nodules were dissolved in 0.5 N HCl
with 0.5 ml 5% SDS for 30 min.
To quantify Alizarin Red-stained nodules, the stain
was solubilized with 0.5 ml 5% SDS in 0.5 N HCl for 30 min at room
temperature. Subsequently, 0.15 ml of the liquid was transferred to
a 96-well plates and absorbance value were measured at 405 nm using
a microplate reader (Bio-Tek Instruments, Winooski, VT,, USA). All
assays were repeated three times.
Western blot analyses
hPDLSCs and pPDLSCs were lysed in
radioimmunoprecipitation assay buffer and protein content of the
lysate was determined using a protein assay kit (Beyotime Institute
of Biotechnology) according to the manufacturer's instructions.
Then, 20 mg cell protein lysate was boiled for 10 min and was
resolved using 10% SDS-PAGE. The proteins were transferred to a
polyvinylidene difluoride membrane (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). Membranes were blocked in 5% bovine serum
albumin (Solarbio, Beijing, China) at room temperature for 2 h and
were then incubated with primary antibodies at room temperature for
4 h. Subsequently, membranes were incubated with anti-rabbit or
anti-mouse IgG antibodies at room temperature for 2 h.
Immunodetection was performed using the Western-Light
Chemiluminescent Detection system (JS-1070P; Peiqing P&Q
Science and Technology, Shanghai, China). We then performed
densitometry using Image J software (National Institutes of Health,
Bethesda, MD, USA). All assays were repeated three times.
The following primary antibodies were used: p38
(1:1,000; cat. no. 9212S) and phospho (p)-p38 (1:1,000; cat. no.
4511; both from Cell Signaling Technology, Inc.); β-actin (1:800;
cat. no. CW0096A; CWBio, Co., Ltd., Beijing, China); secondary
antibodies, anti-rabbit and anti-mouse IgG anti-bodies (1: 10,000;
cat. nos. 115-035-003 and 111-035-003; Jackson ImmunoResearch
Laboratories, Inc., West Grove, PA, USA)
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated using TRIzol reagent (Life
Technologies; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions and converted into cDNA using a
PrimeScript RT reagent kit (Takara Biotechnology Co., Ltd., Dalian,
China). For RT of mRNA, random-primed cDNA was synthesized from 2
mg total RNA. qPCR analysis was performed using the SYBR Premix Ex
Taq II kit (Takara Biotechnology Co., Ltd.) and detected on the ABI
Prism 7500 HT sequence detection system (Applied Biosystems; Thermo
Fisher Scientific, Inc.). β-actin was used for quantitation of
mRNAs. The data were analyzed using the 2−ΔΔCq (27) relative expression method. All
assays were repeated three times. Primer pairs were as follows:
Runt related transcription factor 2 (Runx2), 5′-CCC GTG GCC TTC
AAGGT-3′, 5′-CGT TAC CCG CCA TGA CAGTA-3′; ALP, 5′-GGA CCA TTC CCA
CGT CTT CAC-3′, 5′-CCT TGT AGC CAG GCC CATTG-3′; osteocalcin (OCN),
5′-CCC AGG CGC TAC CTG TAT CAA-3′, 5′-GGT CAG CCA ACT CGT CAC
AGTC-3′; and β-actin, 5′-TGG CAC CCA GCA CAA TGAA-3′, 5′-CTA AGT
CAT AGT CCG CCT AGA AGCA-3′.
Flow cytometric analysis of the cell
cycle
Cell cycle was analyzed by measuring the amount of
propidium iodide (PI) in ethanol fixed cells. Cells were routine
cultured for 5 days. Cells (2×105 cells) were washed
with PBS three times and fixed with cold 70% ethanol for 24 h.
Then, cells were washed with PBS three times and resuspended in 1
ml permeabilizing solution (Triton X-100, sodium azide 0.01% and
RNase A 100 µg/µl) in PBS for 10 min. Following one
wash in PBS, cells were stained with 1 ml PBS with PI (2.5 mg/ml)
and incubated for 15 min at 4°C. Finally, the cell cycle was
measured using a flow cytometer. Cells in G2 and S
phases were considered to be in the proliferation phase. All assays
were repeated three times.
Statistical analyses
All experiments in this study were repe ated at
least three times. Data were analyzed using an independent samples
t-test, and presented as the mean ± standard deviation. The
Bonferroni correction was applied for multiple comparisons.
P<0.05 was considered to indicate a statistically significant
difference.
Results
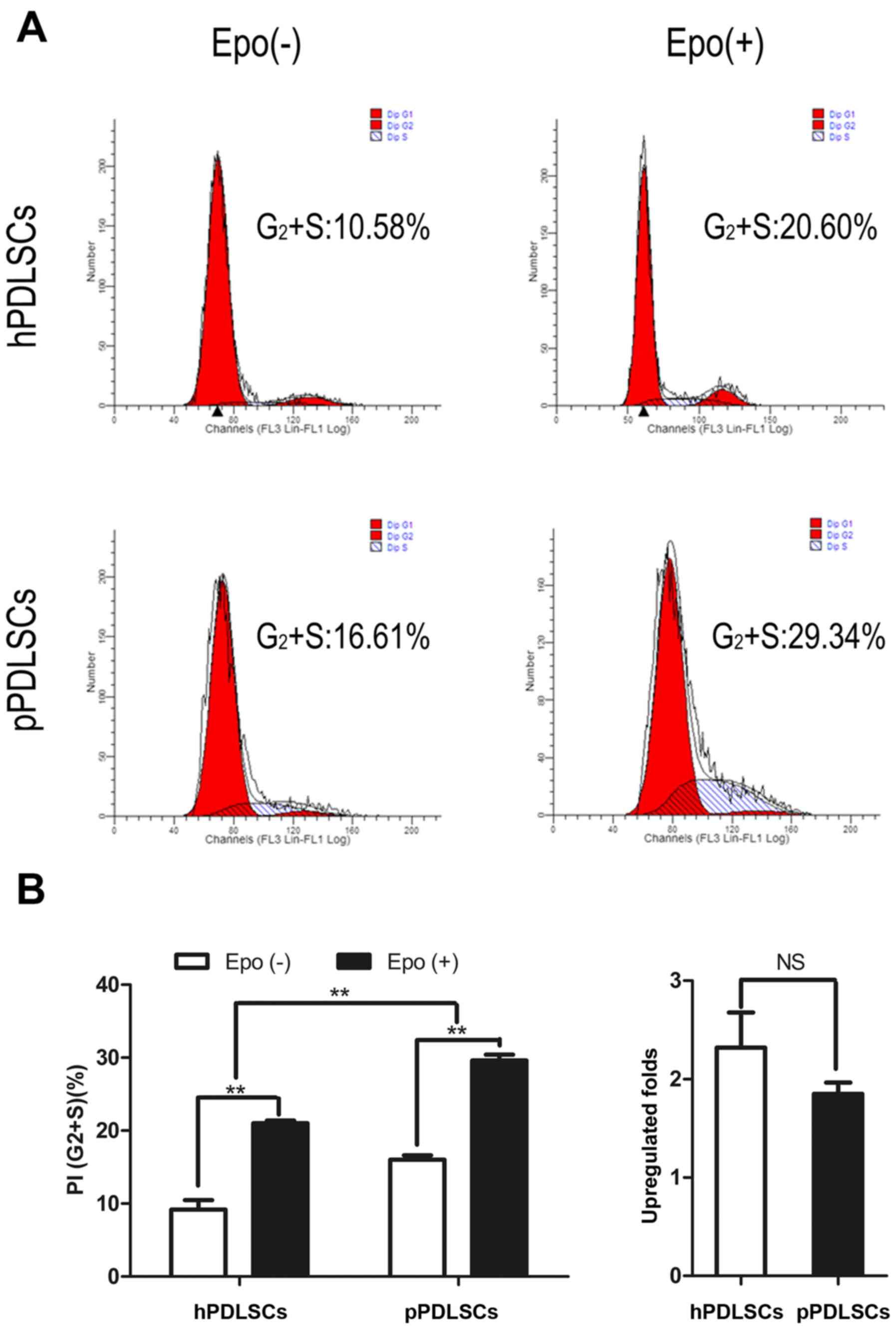
Effect of Epo on the proliferation of
hPDLSCs and pPDLSCs
hPDLSCs and pPDLSCs were cultured in normal culture
medium and Epo-induced culture medium (with 20 IU/ml Epo). Cell
proliferation capacities were assessed using cell cycle analysis.
The percentages of cells in the proliferation phase were considered
as the proliferation index (PI). The PI of hPDLSCs and pPDLSCs
induced by Epo was increased compared with untreated hPDLSCs and
pPDLSCs (Fig. 1A). Notably,
regarding proliferation, hPDLSCs exhibited greater sensitivity to
Epo based on the PI; however, pPDLSCs exhibited greater
proliferation ability than hPDLSCs (Fig. 1B).
Effects of Epo on osteogenic
differentiation of hPDLSCs and pPDLSCs
The effects of Epo on the osteogenic differentiation
of hPDLSCs and pPDLSCs were analyzed by ALP staining (Fig. 2A) and ALP activity assay (Fig. 2B), and the mRNA transcript levels
of genes Runx2, ALP and OCN (Fig.
2C) were measured. Furthermore, to characterize the effect of
Epo, mineralized nodule formation was analyzed by Alizarin Red
staining following induction of cells in osteogenic media for 21
days. Epo promoted the osteogenesis of hPDLSCs and pPDLSCs
(Fig. 2D and E). We also found
that inflammation could affect osteogenesis in hPDLSCs (Fig. 2A–E).
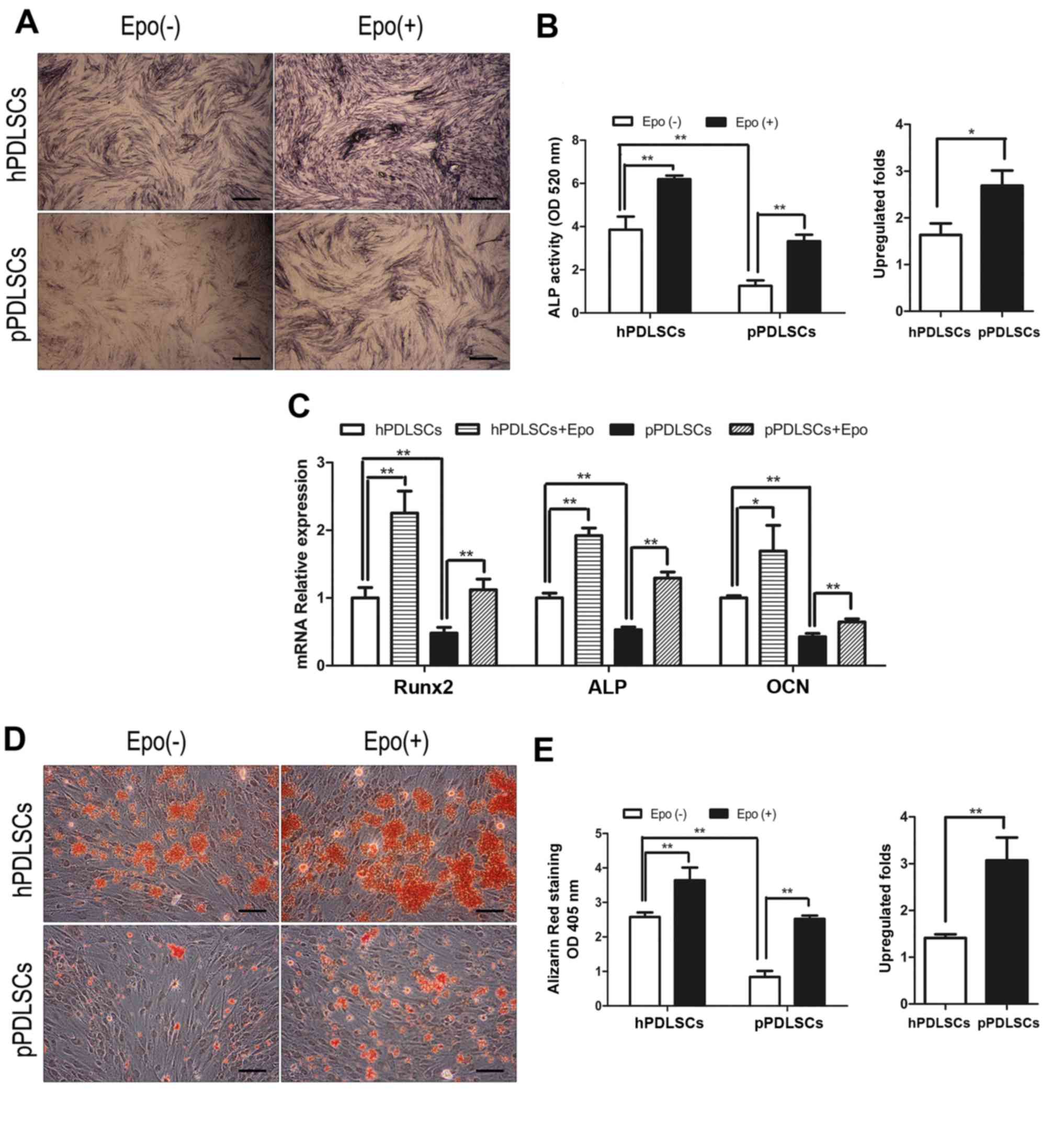 | Figure 2Effects of Epo on the osteogenic
differentiation of hPDLSCs and pPDLSCs. (A) Osteogenic
differentiation was determined by ALP staining at day 7 after
osteogenic differentiation induced. (B) ALP activity was measured
by ALP activity assay at day 7 after osteogenic differentiation
induced and fold upregulation of ALP activity in hPDLSCs and
pPDLSCs induced by Epo. (C) Expression levels of the osteogenic
genes ALP, Runx2 and OCN were measured by reverse
transcription-quantitative polymerase chain reaction at day 7 after
osteogenic differentiation induced. (D) Osteogenic differentiation
was determined by Alizarin Red S staining at day 21 after
osteogenic differentiation induced. (E) Calcium concentration
determined by Alizarin Red S and fold upregulation of calcium
concentration in hPDLSCs and pPDLSCs induced by Epo. Data are
presented as the mean ± standard deviation.*P<0.05,
**P<0.01. Scale bar, 100 µm. Epo(-), cells
cultured without Epo; Epo(+), cells cultured with Epo; Epo,
erythropoietin; hPDLSC, human periodontal ligament tissue-derived
mesenchymal stem cells; pPDLSCs, periodontitis mesenchymal stem
cells; Runx2, runt related transcription factor 2; ALP, alkaline
phosphatase; OCN, osteocalcin; OD, optical density. |
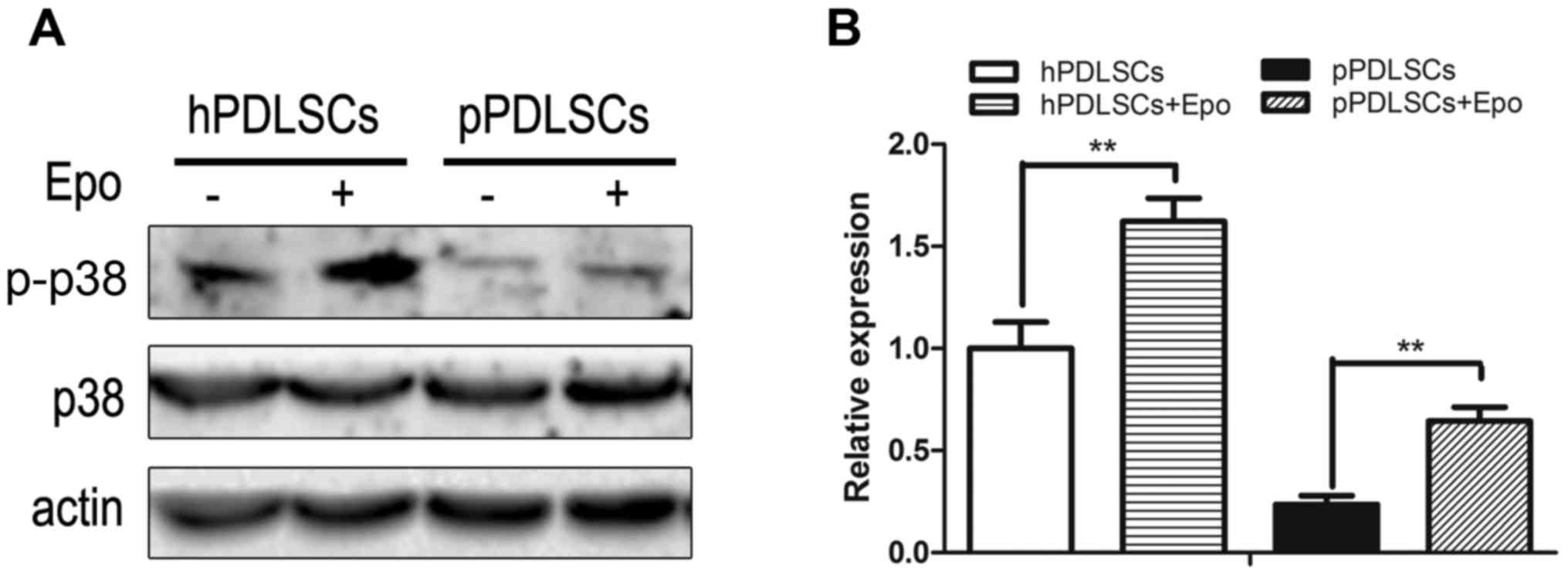
Effects of Epo on the p38 MAPK
pathways
To identify the mechanism by which Epo regulates the
bone formation capacity of hPDLSCs and pPDLSCs, the effects of Epo
on the p38 MAPK pathways in hPDLSCs and pPDLSCs were analyzed based
on the phosphorylation of p38. hPDLSCs and pPDLSCs induced by Epo
exhibited increased p-p38 level compared with the untreated control
groups (Fig. 3). However, the
expression of total p38 protein was not altered.
Role of p38 MAPK pathways in Epo-mediated
regulation of osteogenic differentiation of hPDLSCs and
pPDLSCs
To characterize the role of the p38 MAPK pathways on
the processes whereby Epo regulates the osteogenic differentiation
of hPDLSCs and pPDLSCs, this pathway was inhibited using the
pathway-specific inhibitor SB203580. Following treatment with
SB203580, the osteogenic capacities of hPDLSCs and pPDLSCs with or
without Epo were determined. The mineralized nodule formation
(Fig. 4A and B) and ALP activity
(Fig. 4C and D) induced by Epo
were also decreased significantly when cells were co-treated with
SB203580. The expression level of osteogenic genes was measured
using RT-qPCR. SB203580 treatment led to a significant decrease in
Epo-induced Runx2, ALP and OCN expression (Fig. 4E). In conclusion, the effects of
Epo on the osteogenic differentiation of hPDLSCs and pPDLSCs were
reduced when the MAPK pathway was inhibited by SB203580.
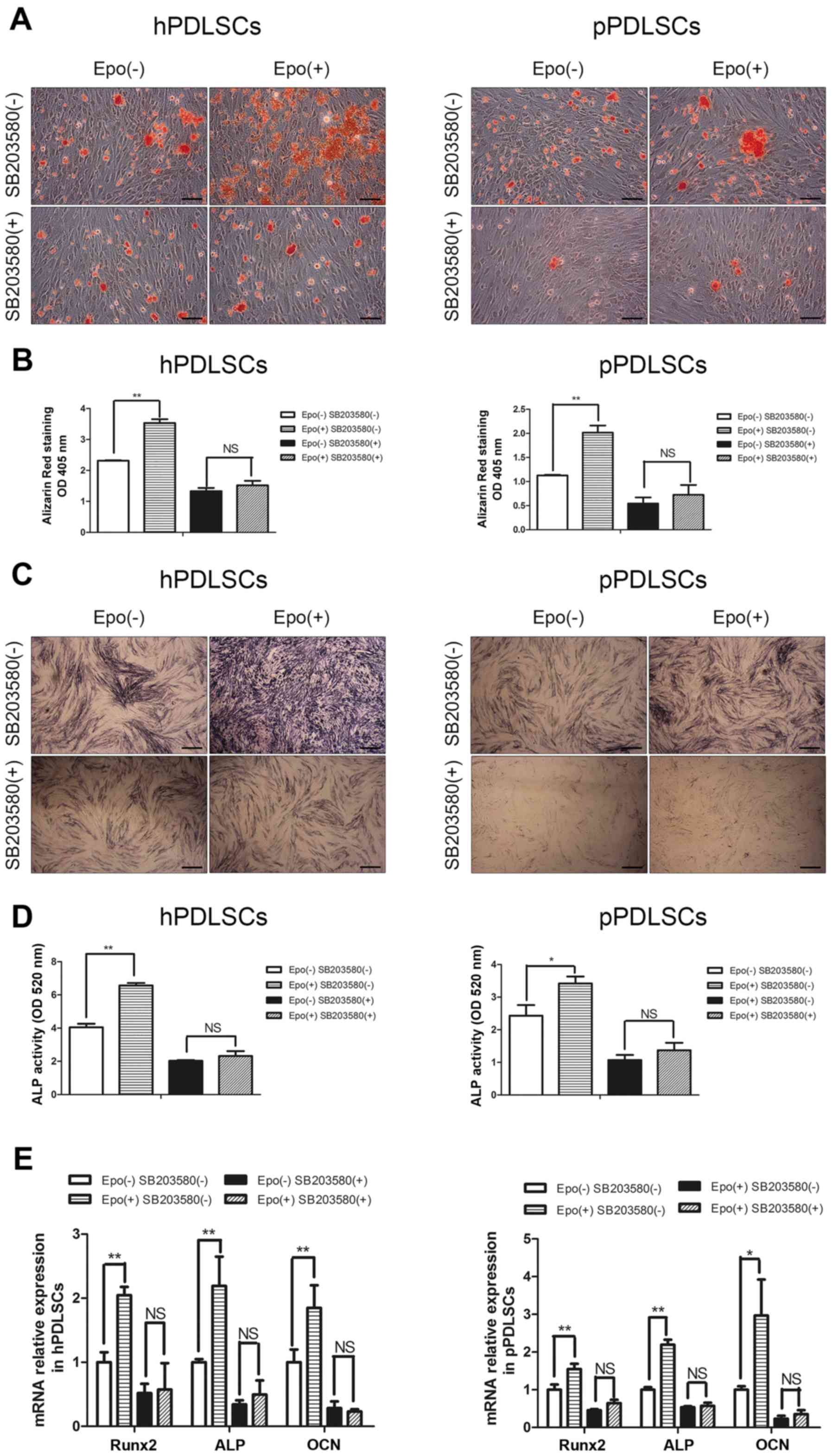 | Figure 4Effects of the p38 mitogen-activated
protein kinase pathway on the processes of Epo regulating the
osteogenic differentiation of hPDLSCs and pPDLSCs. (A) Osteogenic
differentiation was determined by Alizarin Red S staining at day 21
after osteogenic differentiation induced. (B) Alizarin Red S
staining was used to determine calcium levels. (C) Osteogenic
differentiation was determined by alkaline phosphatase (ALP)
staining at day 7 after osteogenic differentiation induced. (D) ALP
activity was measured by ALP activity assay at day 7 after
osteogenic differentiation induced. (E) Expression levels of the
osteogenic genes ALP, Runx2 and OCN were measured by reverse
transcription-quantitative polymerase chain reaction at day 7 after
osteogenic differentiation induced. All all analyses were performed
after 7 days in culture. Data are presented as the mean ± standard
deviation. *P<0.05, **P<0.01. Scale
bar, 100 µm. hPDLSC, human periodontal ligament
tissue-derived mesenchymal stem cells; pPDLSCs,
periodontitis mesenchymal stem cells; Epo, erythropoietin; Epo(-),
cells cultured without Epo; Epo(+), cells cultured with Epo;
SB203580(-), cells cultured without SB203580; SB203580(+), cells
cultured with SB203580; ALP, alkaline phosphatase; OD, optical
density; NS, not significant; Runx2, runt related transcription
factor 2; OCN, osteocalcin. |
Discussion
Epo, which was first discovered as a regulator of
erythropoiesis, has been used as a therapeutic for certain red
blood cell disorders (28).
Furthermore, it has been reported to have multiple biological
functions, including repair of neuronal injury, improve the
proliferation and differentiation in endothelial progenitor cells,
and promotion of wound healing (29). Recently, the effects of Epo on the
regulation of osteogenic differentiation of MSCs have attracted
significant attention. Epo may induce the osteogenesis of MSCs by
promoting cell proliferation, migration and differentiation
(30). Kim et al (3) reported that Epo can regulate
differentiation of both osteoblasts and osteoclasts through
mechanistic target of rapamycin kinase (mTOR) signaling. In this
process, Epo improved bone formation of MSCs. Furthermore, Epo also
increases nuclear factor of activated T-cells cytoplasmic 1
expression and decreases cathepsin K expression in an
mTOR-independent manner, resulting in an increase of osteoclast
numbers and a decrease in resorption activity (3). However, the effects of Epo on
osteogenic differentiation of hPDLSCs remain unknown.
In the present study, Epo increased the osteogenic
differentiation of hPDLSCs, as indicated by the expression of
osteogenic genes, Alizarin Red and ALP staining, and an ALP
activity assay. Osteogenic differentiation was defined based on the
differentiation of a sufficient number of cells. Thus, the effect
of Epo on the proliferation of hPDLSCs was also assessed, which was
determined by cell cycle analyses. Epo increased the proliferation
of hPDLSCs.
Periodontitis is a type of inflammatory disease
characterized by the destruction of periodontal tissues that
contain alveolar bone, periodontal ligament and root cementum.
However, the regeneration of periodontal hard tissues is always an
essential problem in such studies.
It has previously been established that hPDLSCs
present in an inflammatory microenvironment for a long period will
exhibit negative effects on osteogenesis (23). Such exposure may lead to
periodontal bone tissue defects. Therefore, determining how to
restore the normal capacity of osteogenic differentiation in
pPDLSCs is an important goal for studies of periodontal
regeneration. In the present study, Epo upregulated the
osteogenesis of pPDLSCs by inducing the expression of osteogenic
genes. Alizarin Red and ALP staining, and ALP activity were also
increased by Epo. In addition, among the osteogenesis genes (Runx2,
ALP and OCN), Runx2 was changed most markedly. Runx2 is an
osteoblast differentiation factor that is often expressed in
mesenchymal cell types. Runx2 often promotes the level of bone
morphogenic protein and expressed in terminally differentiated
osteoblasts (31). Notably,
proinflammatory T cells may inhibit MSC-mediated bone formation via
tumor necrosis factor (TNF)-α-induced downregulation of Runx2
(32). Therefore, the promotion
of bone regeneration by Epo-induced pPDLSCs may exert an
anti-inflammatory effect that reduces levels of TNF-α expression.
Compared with hPDLSCs, pPDLSCs have increased capacities for
proliferation. We also demonstrated that the proliferation of
pPDLSCs was further improved when induced by Epo.
The MAPK pathway is involved in various cellular
processes, including cell proliferation, survival and
differentiation (33-35). Among the mediators of the MAPK
pathway, p38 was reported to be involved in early and late bone
formation of osteoblasts, MSCs and MC3T3-E1 cells (36). In this process, p38 could increase
bone homeostasis and osteogenesis through Runx2, the key
transcription factor of osteogenic differentiation (37). Furthermore, the findings of the
current study indicate that the p38 MAPK pathway may also promote
bone regeneration of hPDLSCs and pPDLSCs. Chang et al
(38) reported that activating
p38 MAPK signaling can enhance the bone formation ability of
hPDLSCs. When the p38 level was reduced, the osteogenic capacities
of hPDLSCs were inhibited (39).
In an inflammatory environment, the activation of p38 MAPK was
altered in PDLSCs during the osteogenic differentiation, and the
osteogenesis ability of pPDLSCs was damaged (23).
The present study identified that the p38
phosphorylation level was decreased in pPDLSCs, compared with
hPDLSCs, which is consistent with a previous study (23). Additionally, the results
demonstrated that Epo promoted the phosphorylation of p38, which
demonstrated that p38 MAPK can be activated by Epo in hPDLSCs and
pPDLSCs. Previous studies have demonstrated that Epo can positively
regulate the p38 MAPK pathway in multiple cell lines, including
smooth muscle cells, heart cells and MSCs (24-26). However, whether this process is
involved in osteogenesis has not been thoroughly investigated. In
the present study, the osteogenic effects of Epo in hPDLSCs and
pPDLSCs were attenuated when the p38 signaling pathway was
inhibited. All these findings strongly demonstrate that Epo can
induce osteogenesis of hPDLSCs and pPDLSCs by activating the p38
MAPK pathway.
In conclusion, the present study established that
Epo upregulates osteogenesis and the proliferation of hPDLSCs and
pPDLSCs. The underlying mechanism may involve p38 MAPK signaling.
Further animal studies are required to verify the function and
safety of Epo in promoting the osteogenesis capacity of hPDLSCs and
pPDLSCs in vivo.
Acknowledgments
This study was supported by grants from the National
Natural Science Foundation of China (grant no. 81271176)
References
|
1
|
Holstein JH, Orth M, Scheuer C, Tami A,
Becker SC, Garcia P, Histing T, Mörsdorf P, Klein M, Pohlemann T,
et al: Erythropoietin stimulates bone formation, cell
proliferation, and angiogenesis in a femoral segmental defect model
in mice. Bone. 49:1037–1045. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shiozawa Y, Jung Y, Ziegler AM, Pedersen
EA, Wang J, Wang Z, Song J, Wang J, Lee CH, Sud S, et al:
Erythropoietin couples hematopoiesis with bone formation. PLoS One.
5:e108532010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kim J, Jung Y, Sun H, Joseph J, Mishra A,
Shiozawa Y, Wang J, Krebsbach PH and Taichman RS: Erythropoietin
mediated bone formation is regulated by mTOR signaling. J Cell
Biochem. 113:220–228. 2012. View Article : Google Scholar
|
|
4
|
Chen S, Li J, Peng H, Zhou J and Fang H:
Administration of erythropoietin exerts protective effects against
glucocorticoid-induced osteonecrosis of the femoral head in rats.
Int J Mol Med. 33:840–848. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pihlstrom BL, Michalowicz BS and Johnson
NW: Periodontal diseases. Lancet. 366:1809–1820. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nanci A and Bosshardt DD: Structure of
periodontal tissues in health and disease. Periodontol. 40:11–28.
2006. View Article : Google Scholar
|
|
7
|
Chen FM, Zhang J, Zhang M, An Y, Chen F
and Wu ZF: A review on endogenous regenerative technology in
periodontal regenerative medicine. Biomaterials. 31:7892–7927.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Izumi Y, Aoki A, Yamada Y, Kobayashi H,
Iwata T, Akizuki T, Suda T, Nakamura S, Wara-Aswapati N, Ueda M, et
al: Current and future periodontal tissue engineering. Periodontol.
56:166–187. 2011. View Article : Google Scholar
|
|
9
|
Seo BM, Miura M, Gronthos S, Bartold PM,
Batouli S, Brahim J, Young M, Robey PG, Wang CY and Shi S:
Investigation of multi-potent postnatal stem cells from human
periodontal ligament. Lancet. 364:149–155. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Akiyama K, Chen C, Gronthos S and Shi S:
Lineage differentiation of mesenchymal stem cells from dental pulp,
apical papilla, and periodontal ligament. Methods Mol Biol.
887:111–121. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Washio K, Iwata T, Mizutani M, Ando T,
Yamato M, Okano T and Ishikawa I: Assessment of cell sheets derived
from human periodontal ligament cells: a pre-clinical study. Cell
Tissue Res. 341:397–404. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gault P, Black A, Romette JL, Fuente F,
Schroeder K, Thillou F, Brune T, Berdal A and Wurtz T:
Tissue-engineered ligament: implant constructs for tooth
replacement. J Clin Periodontol. 37:750–758. 2010.PubMed/NCBI
|
|
13
|
Yang H, Gao LN, An Y, Hu CH, Jin F, Zhou
J, Jin Y and Chen FM: Comparison of mesenchymal stem cells derived
from gingival tissue and periodontal ligament in different
incubation conditions. Biomaterials. 34:7033–7047. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ma Z, Li S, Song Y, Tang L, Ma D, Liu B
and Jin Y: The biological effect of dentin noncollagenous proteins
(DNCPs) on the human periodontal ligament stem cells (HPDLSCs) in
vitro an in vivo. Tissue Eng Part A. 14:2059–2068. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen FM, Zhao YM, Wu H, Deng ZH, Wang QT,
Zhou W, Liu Q, Dong GY, Li K, Wu ZF, et al: Enhancement of
periodontal tissue regeneration by locally controlled delivery of
insulin-like growth factor-I from dextran-co-gelatin microspheres.
J Control Release. 114:209–222. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ramseier CA, Abramson ZR, Jin Q and
Giannobile WV: Gene therapeutics for periodontal regenerative
medicine. Dent Clin North Am. 50:245–263. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu RX, Bi CS, Yu Y, Zhang LL and Chen FM:
Age-related decline in the matrix contents and functional
properties of human periodontal ligament stem cell sheets. Acta
Biomater. 22:70–82. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang J, An Y, Gao LN, Zhang YJ, Jin Y and
Chen FM: The effect of aging on the pluripotential capacity and
regenerative potential of human periodontal ligament stem cells.
Biomaterials. 33:6974–6986. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhou Y, Fan W and Xiao Y: The effect of
hypoxia on the stemness and differentiation capacity of PDLC and
DPC. BioMed Res Int. 890675:2014. View Article : Google Scholar
|
|
20
|
Kim SY, Kang KL, Lee JC and Heo JS:
Nicotinic acetylcholine receptor α7 and β4 subunits contribute
nicotine-induced apoptosis in periodontal ligament stem cells. Mol
Cells. 33:343–350. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu W, Konermann A, Guo T, Jäger A, Zhang
L and Jin Y: Canonical Wnt signaling differently modulates
osteogenic differentiation of mesenchymal stem cells derived from
bone marrow and from periodontal ligament under inflammatory
conditions. Biochim Biophys Acta. 1840:1125–1134. 2014. View Article : Google Scholar
|
|
22
|
Zheng W, Wang S, Wang J and Jin F:
Periodontitis promotes the proliferation and suppresses the
differentiation potential of human periodontal ligament stem cells.
Int J Mol Med. 36:915–922. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu N, Shi S, Deng M, Tang L, Zhang G, Liu
N, Ding B, Liu W, Liu Y, Shi H, et al: High levels of β-catenin
signaling reduce osteogenic differentiation of stem cells in
inflammatory microenvironments through inhibition of the
noncanonical Wnt pathway. J Bone Miner Res. 26:2082–2095. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu N, Tian J, Cheng J and Zhang J: Effect
of erythropoietin on the migration of bone marrow-derived
mesenchymal stem cells to the acute kidney injury microenvironment.
Exp Cell Res. 319:2019–2027. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Park SL, Won SY, Song JH, Kambe T, Nagao
M, Kim WJ and Moon SK: EPO gene expression promotes proliferation,
migration and invasion via the p38M/APK/AP-1/MMP-9 pathway by
p21WAF1 expression in vascular smooth muscle cells. Cell Signal.
27:470–478. 2015. View Article : Google Scholar
|
|
26
|
Rafiee P, Shi Y, Su J, Pritchard KA Jr,
Tweddell JS and Baker JE: Erythropoietin protects the infant heart
against ischemia-reperfusion injury by triggering multiple
signaling pathways. Basic Res Cardiol. 100:187–197. 2005.
View Article : Google Scholar
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)). Method Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
28
|
Wu H, Liu X, Jaenisch R and Lodish HF:
Generation of committed erythroid BFU-E and CFU-E progenitors does
not require erythropoietin or the erythropoietin receptor. Cell.
83:59–67. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Broxmeyer HE: Erythropoietin: multiple
targets, actions, and modifying influences for biological and
clinical consideration. J Exp Med. 210:205–208. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li C, Shi C, Kim J, Chen Y, Ni S, Jiang L,
Zheng C, Li D, Hou J, Taichman RS, et al: Erythropoietin promotes
bone formation through EphrinB2/EphB4 signaling. J Dent Res.
94:455–463. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ducy P, Zhang R, Geoffroy V, Ridall AL and
Karsenty G: Osf2/Cbfa1: a transcriptional activator of osteoblast
differentiation. Cell. 89:747–754. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu Y, Wang L, Kikuiri T, Akiyama K, Chen
C, Xu X, Yang R, Chen W, Wang S and Shi S: Mesenchymal stem
cell-based tissue regeneration is governed by recipient T
lymphocytes via IFN-γ and TNF-α. Nat Med. 17:1594–1601. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu DJ, Zhao YZ, Wang J, He JW, Weng YG and
Luo JY: Smads, p38 and ERK1/2 are involved in BMP9-induced
osteogenic differentiation of C3H10T1/2 mesenchymal stem cells. BMB
Rep. 45:247–252. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhao Y, Song T, Wang W, Wang J, He J, Wu
N, Tang M, He B and Luo J: P38 and ERK1/2 MAPKs act in opposition
to regulate BMP9-induced osteogenic differentiation of mesenchymal
progenitor cells. PLoS One. 7:e433832012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang Y, Ma J, Du Y, Miao J and Chen N:
Human amnion-derived mesenchymal stem cells protect human bone
marrow mesenchymal stem cells against oxidative stress-mediated
dysfunction via ERK1/2 MAPK signaling. Mol Cells. 39:186–194. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Greenblatt MB, Shim JH, Zou W, Sitara D,
Schweitzer M, Hu D, Lotinun S, Sano Y, Baron R, Park JM, et al: The
p38 MAPK pathway is essential for skeletogenesis and bone
homeostasis in mice. J Clin Invest. 120:2457–2473. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Higuchi C, Myoui A, Hashimoto N, Kuriyama
K, Yoshioka K, Yoshikawa H and Itoh K: Continuous inhibition of
MAPK signaling promotes the early osteoblastic differentiation and
mineralization of the extracellular matrix. J Bone Miner Res.
17:1785–1794. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chang J, Sonoyama W, Wang Z, Jin Q, Zhang
C, Krebsbach PH, Giannobile W, Shi S and Wang CY: Noncanonical
Wnt-4 signaling enhances bone regeneration of mesenchymal stem
cells in craniofacial defects through activation of p38 MAPK. J
Biol Chem. 282:30938–30948. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ye G, Li C, Xiang X, Chen C, Zhang R, Yang
X, Yu X, Wang J, Wang L, Shi Q, et al: Bone morphogenetic protein-9
induces PDLSCs osteogenic differentiation through the ERK and p38
signal pathways. Int J Med Sci. 11:1065–1072. 2014. View Article : Google Scholar : PubMed/NCBI
|