Introduction
Prostate cancer (PCa) is a major cause of morbidity
and mortality among men worldwide, with recent trends indicating
that younger men are being affected and that the incidence rates
are increasing annually (1). PCa
is also associated with other diseases, including metabolic
syndrome and diabetes mellitus. As with other types of cancer,
tumor metastasis is the main cause of mortality in patients with
PCa; and cell migration and invasion are two important features of
metastasis and the spread of tumor cells. The processes that
control migration, invasion and ultimately tumor metastasis are not
yet fully understood (2,3). At the molecular and cellular level,
metastasizing prostate tumor cells undergo numerous alterations,
including cell-cell and cell-matrix adhesion, cytoskeletal
reorganization and activation of various signaling pathways.
Although numerous studies have investigated cell migration and
invasion, their underlying mechanisms are not yet fully understood
(4,5).
Prohibitin 2 (PHB2), also known as BAP37, REA and
prohibitone, is one of two highly homologous inhibitins. As a
multifunctional protein, PHB2 participates in numerous important
cellular processes, including cell survival, apoptosis, metabolism,
inflammation, gene transcription and signal transduction (6,7).
PHB2 is predominantly located in the cell nucleus, mitochondria and
cell membrane, and is highly expressed in PCa, liver cancer,
pancreatic carcinoma and breast cancer compared with in
corresponding normal tissues (8-11).
These observations may be associated with the fact that PHB2 serves
prominent roles in maintaining mitochondrial function, and
promoting cell proliferation and survival (12–14). Furthermore, PHB2 is involved in
cell growth, resistance to chemotherapy and tumor metastasis by
modulating the transcriptional activity of transforming growth
factor-β in the Ras-cRaf-mitogen-activated protein kinase
kinase-extracellular signal-regulated kinase signaling pathway
(15,16). In addition, the expression of PHB2
has been revealed to be upregulated in early invasive stages but
downregulated in later stages in hepatocellular carcinoma (17). To the best of our knowledge, a
single report demonstrated that knockdown of PHB2 abolished
insulin-like growth factor-binding protein-6 (IGFBP-6)002Dinduced
Rh30 cell migration (18). In
addition, relatively little is known about PHB2 and migration,
which is the focus of the present study.
As a serine protein kinase, AKT serine/threonine
kinase (AKT) modulates various cellular processes, including
metabolism, proliferation, migration, apoptosis and angiogenesis by
phosphorylating downstream substrates (19). AKT2 was the first gene
identified in the AKT family that could change at the genomic level
in diabetes patients and serve a key role in the malignant process
(20–22). In PC3 PCa cells, AKT2 has been
reported to inhibit cell migration and invasion by activating
β-integrin. Conversely, silencing of AKT2 resulted in an increase
in focal adhesion size, upregulation of microRNA-200 family
expression and improvement of integrin activity, which may
eventually enhance cell motility (23). In addition, AKT2 knockout enhanced
cell migration by regulating the Rac/Pak signaling pathway in mouse
embryonic fibroblast cells (24).
One possible reason for the participation of AKT2 in cell migration
is that AKT2 upregulates K18 and vimentin expression by increasing
mRNA stability, which may affect intermediate filament expression
(25). Similarly, as a
multifunctional protein, AKT2 is located in the mitochondria,
nuclei and cell membrane, and is involved in several cellular
processes, including cell cycle regulation, mitochondrial stability
and cell migration. Notably, there is some overlap in the cellular
processes regulated by AKT2 and PHB2, thus indicating a potential
functional correlation between them.
There are many similarities in the cellular
localization and function of AKT2 and PHB2. Both are located in the
mitochondria, nucleus and cell membrane, which may be associated
with their signal transduction function (11,23,26). Localization of PHB2 in the nucleus
allows it to partially interact with AKT2 in differentiated
fibroblasts. In addition, there appears to be a negative
correlation between AKT2 and PHB2 protein levels; AKT2-silenced
cells exhibited increased levels of PHB2, whereas AKT2
overexpression resulted in decreased PHB2 expression, thus
suggesting overlapping functions between the two proteins (27). In myoblasts, AKT2 negatively
modulates the expression of PHB2, and exerts negative feedback
regulation (28,29).
Previous studies have indicated that AKT2 is a
negative regulator of cell migration in PCa, whereas PHB2 may be
positively correlated with the malignant progression of PCa
(23). Therefore, it may be
hypothesized that a regulatory interaction between AKT2 and PHB2
exists, which is associated with migration of PCa cells. The
present study demonstrated that PHB2 could significantly accelerate
migration of PCa cells and negatively modulate AKT2 expression.
Furthermore, knockdown of PHB2 increased the stability of AKT2
protein expression, whereas co-overexpression of PHB2 and AKT2
inhibited cell migration. These results indicated that PHB2 may
promote the migration of PCa cells by inhibiting AKT2
expression.
Materials and methods
Cell culture and treatment
RWPE-1 and BPH-1 cell lines were provided as gifts
from Professor Helmut Klocker (Innsbruck University School of
Medicine (Innsbruck, Austria). The human PCa cell line PC3 was
purchased from the Deutsche Sammlung fuer Mikroorganismen und
Zellkulturen (Braunschweig, Germany) and the PCa cell line DU145
was obtained from the American Type Culture Collection (ATCC,
Manassas, VA, USA). RWPE-1 cells were cultured in keratinocyte
serum-free medium (ATCC) supplemented with 0.05 mg/ml bovine
pituitary extract (cat. no. 0713; ScienCell, San Diego, CA, USA)
and 5 ng/ml human recombinant epidermal growth factor (cat. no.
10784-015; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). The other cell lines were cultured in RPMI-1640 medium
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) supplemented with
100 mg/ml penicillin/streptomycin (HyClone; GE Healthcare Life
Sciences, Logan, UT, USA) and 10% fetal bovine serum (FBS; Gibco;
Thermo Fisher Scientific, Inc.). Cells were routinely cultured in a
5% CO2 incubator at 37°C.
Oncomine data analysis
Oncomine is freely available to the academic
research community at http://www.oncomine.org. The method was performed as
described previously (30,31).
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from PC3 and DU145 cells
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.).
RT was performed for 2 h at 37°C as described previously (32). RT-PCR reagents were purchased from
Invitrogen; Thermo Fisher Scientific, Inc. (cat. no. AM1005M). The
relative gene expression levels were determined using the
comparative Cq method (33) and
were normalized to the housekeeping gene hypoxanthine
phosphoribosyltransferase 1 (HPRT). Primer sequences were as
follows: HPRT, sense, 5′-TGACACTGGCAAAACAATGCA-3′ and
antisense, 5′-GGTCCTTTTCACCAGCAAGCT-3′; PHB2, sense,
5′-ACAGAGCCATCTTCTTCAATC-3′ and antisense,
5′-CTCGTTCCTCGTAGTCCAG-3′; and AKT2, sense, 5′-CCC
GGTTTTATGGTGCAGAGAT-3′ and antisense, 5′-GGCCGCACATCATCTCGTACAT-3′.
SYBR-Green I-based real-time quantitative PCR was performed on an
MJ Research DNA Engine Opticon continuous fluorescence detection
system (Opticon Monitor II; MJ Research, Inc.). SYBR™-Green PCR
Master Mix was obtained from Invitrogen; Thermo Fisher Scientific,
Inc. (cat. no. 4309155). The thermocycling conditions were as
follows: step 1, 95°C for 5 min; step 2, 95°C for 30 sec; step 3,
58–65°C for 30 sec; step 4, 72°C for 30 sec; step 5, repeat from
step 2 for 39 cycles; step 6, 72°C for 2 min.
Plasmids, small interfering (si)RNA and
transient transfection
PHB2 or AKT2 fragments were obtained
by PCR amplification of cDNA collected from PC3 cells, and were
cloned into the expression vector pcDNA3.1 (Addgene, Cambridge, MA,
USA). Specific siRNA oligonucleotides targeting PHB2 and
AKT2 mRNA were designed by Shanghai GenePharma Co., Ltd.
(Shanghai, China). The oligonucleotide sequences were as follows:
PHB2, 5′-CUGAACCCCUCUUGGAUUAAGTT-3′; PHB2 siNC,
5′-UUCUCCGAACGUGUCACGUTT-3′ and AKT2,
5′-AAGAGTGGATGCGGGCTATCC-3′ (34). Cell transfection was performed
using Lipofectamine™ 3000 (Invitrogen; Thermo Fisher Scientific,
Inc.). PC3 and DU145 were seeded into 6-well plates at
approximately 70–90% confluence and transduced with Lipofectamine
3000 and plasmid/siRNA, at a ratio of 1:1. Transfected cells were
harvested after 72 h of transfection for RNA or protein extraction.
The plasmid concentration was 2.5 µg for a 6-well, and siRNA
was 75 pmol.
Western blot analysis
Cells were lysed with radioimmunoprecipitation assay
buffer (cat. no. 89900; Thermo Fisher Scientific, Inc.) for 30 min
at ice to extract the total protein, then centrifuged for 10 min
and the supernatant was aspirated. Protein was quantified using the
bicinchoninic acid method.
Proteins extracted from PC3 and DU145 cells (30
µg protein) were separated by 10% SDS-PAGE and were
transferred onto polyvinylidene fluoride membranes. The membranes
were blocked in 4% skim milk powder for 1 h at room temperature.
Antibodies against PHB2 (sc-67045; 1:1,000), AKT2 (sc-5270;
1:1,000) and GAPDH (sc-47724; 1: 5,000) (all from Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) were used. The membranes were
incubated overnight at 4°C. Proteins were detected using
appropriate horseradish peroxidase-conjugated secondary antibodies
(Bio-Rad Laboratories, Inc., Hercules, CA, USA) followed by
enhanced chemiluminescence detection (Amersham; GE Healthcare Life
Sciences, Little Chalfont, UK). The secondary antibodies were
incubated for 1 h at room temperature and the antibodies
information was as follows: goat anti-rabbit IgG-HRP (sc-2004;
1:5,000); goat anti-mouse IgG-HRP (sc-2005; 1:5,000) (both from
Santa Cruz Biotechnology, Inc.). ImageJ 1.47v software (National
Institutes of Health, Bethesda, MD, YSA) was used for densitometry
quantification.
Wound healing assay
A wound healing assay was performed as previously
described (35). Briefly, DU145
and PC3 PCa cells were seeded into 6-well plates and were
transiently transfected with the appropriate plasmids or siRNAs
once cell density reached 80–90%. Subsequently, mitomycin (10
ng/ml) was added, and after 2 h scratch wounds were made by
scraping the cell layer in each culture plate using the tip of a
sterile pipette. The cells were then washed with 1X D-Hank's
buffered saline. The wounded cultures were incubated with medium
containing mitomycin (10 ng/ml). ImageJ software (1.47v; National
Institutes of Health, Bethesda, MD, USA) was used to calculate cell
migration distance. Cell images were captured using Olympus CX41
microscope (Olympus, Tokyo, Japan) 24 h later. The experiments were
performed in triplicate.
Transwell migration assay
Transwell inserts (8 µm pore size; BD
Biosciences, Franklin Lakes, NJ, USA) were used for migration
assays and were placed in 24-well plates. DU145 and PC3 transfected
with PHB2 and/or AKT2 were resuspended in serum-free RPMI-1640
medium in the upper chamber at 40,000 cells/well 48 h after
transfection. RPMI-1640 medium containing 10% FBS was added to the
bottom chamber. Plates were incubated for 24 h, after which
migrated cells were fixed with 4% paraformaldehyde and stained with
10% crystal violet. The stained cells were mounted and visualized
in the bright field with an Olympus CX41 microscope. Positively
stained cells were counted in at least 9 randomly selected
microscopic fields and counted by ImageJ 1.47v software (National
Institutes of Health).
Statistical analysis
Data are presented as the means ± standard
deviation. Significance was assessed using Student's paired t-test.
Data between multiple groups were compared using two-tailed
unpaired t-test or analysis of variance. All data analyses were
performed using SPSS 16.0 software (SPSS Inc., Chicago, IL, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
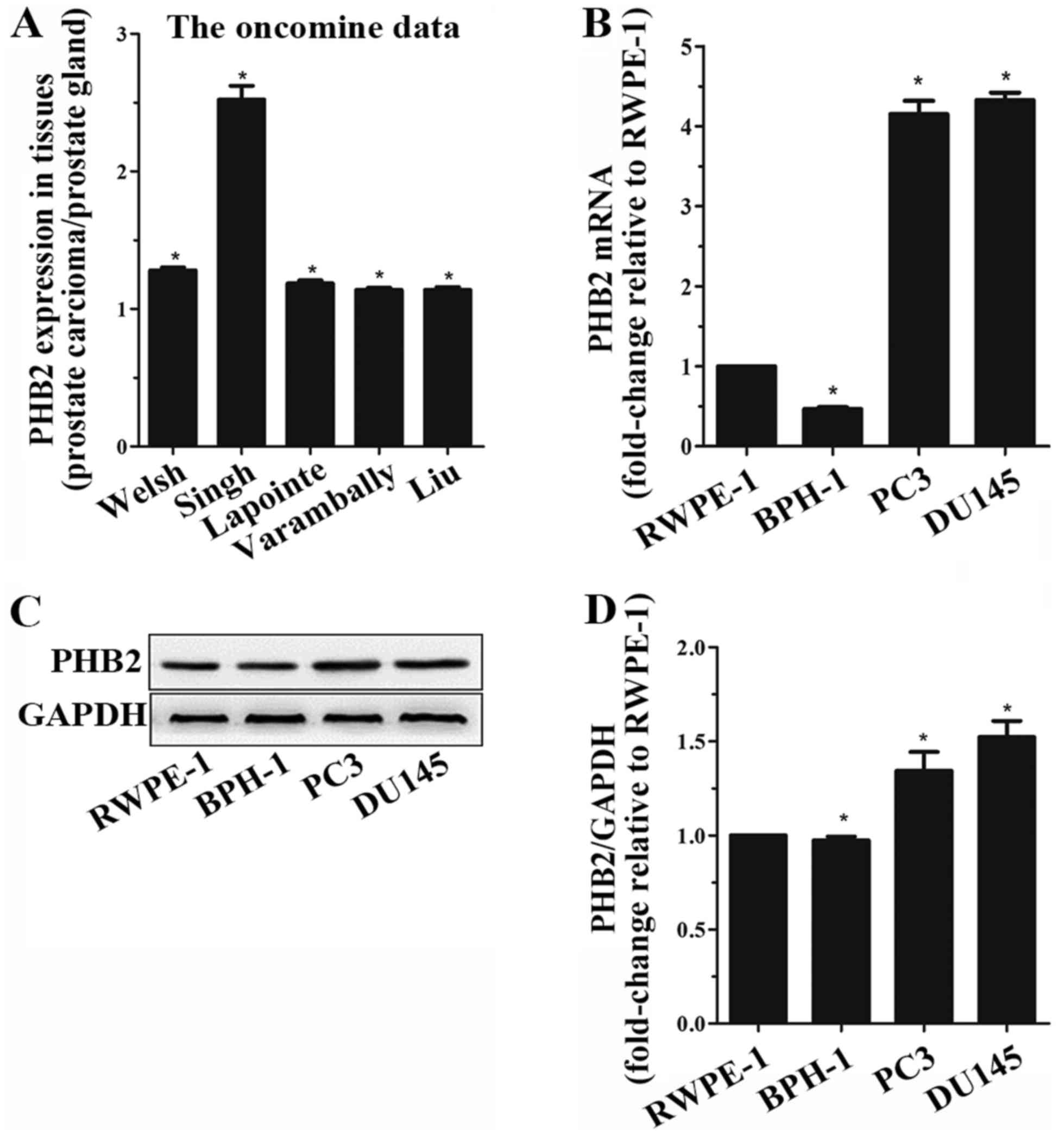
PHB2 expression in prostate tissues
Oncomine data demonstrated that PHB2 is highly
expressed in PCa tissues compared with in normal tissues. Oncomine
data demonstrated the same trend (Fig. 1A). In addition, RT-qPCR and
western blot analysis were used to detect the expression levels of
PHB2 in four prostate epithelial cell lines with various degrees of
malignancy: RWPE-1, BPH-1, DU145 and PC3. PHB2 expression in the
PCa cells, DU145 and PC3, was higher than in the RWPE-1 and BPH-1
cells (Fig. 1B-D). It is well
known that the degree of malignancy is higher in DU145 and PC3
cells compared with in RWPE-1 and BPH-1 cells. These results
indicated that PHB2 expression was increased in PCa cells with
higher malignancy.
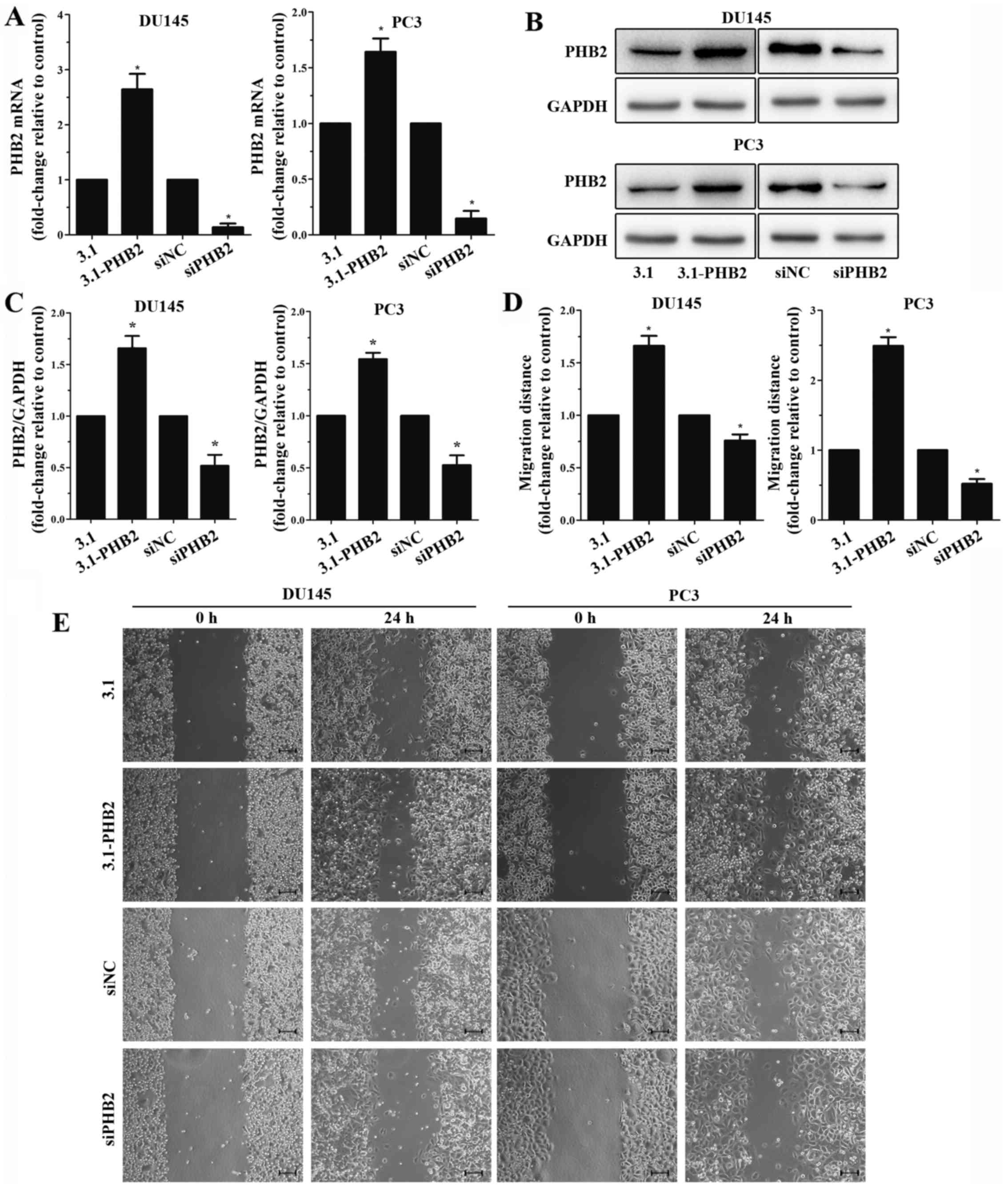
PHB2 promotes PCa cell migration
To further investigate the association between PHB2
and PCa cell migration, pcDNA3.1-PHB2 plasmid or siPHB2 were
transiently transfected into PC3 and DU145 cells, in order to
examine the effects on cell migration. Transfection efficiency was
assessed by RT-qPCR and western blot analysis (Fig. 2A–C). In order to determine the
association between PHB2 and PCa migration, a wound healing assay
was conducted in PC3 and DU145 cells with PHB2 overexpression and
PHB2 knockdown. Wound healing was examined at 0 and 24 h. The
migratory ability of PC3 and DU145 cells with PHB2 overexpression
was significantly enhanced, whereas PHB2 knockdown significantly
reduced migration compared with in the control group within 24 h
(Fig. 2D and E). These results
suggested that PHB2 may promote the migration of PC3 and DU145 PCa
cells, whereas PHB2 knockdown had no effect. Therefore, PHB2 may be
a positive regulator of migration in PC3 and DU145 cells. In
addition, PHB2 overexpression was induced in BPH-1 cells, which
have relatively lower expression levels of PHB2 compared with PC3
and DU145; the results indicated that PHB2 overexpression could
promote migration of BPH-1 cells (Fig. 3).
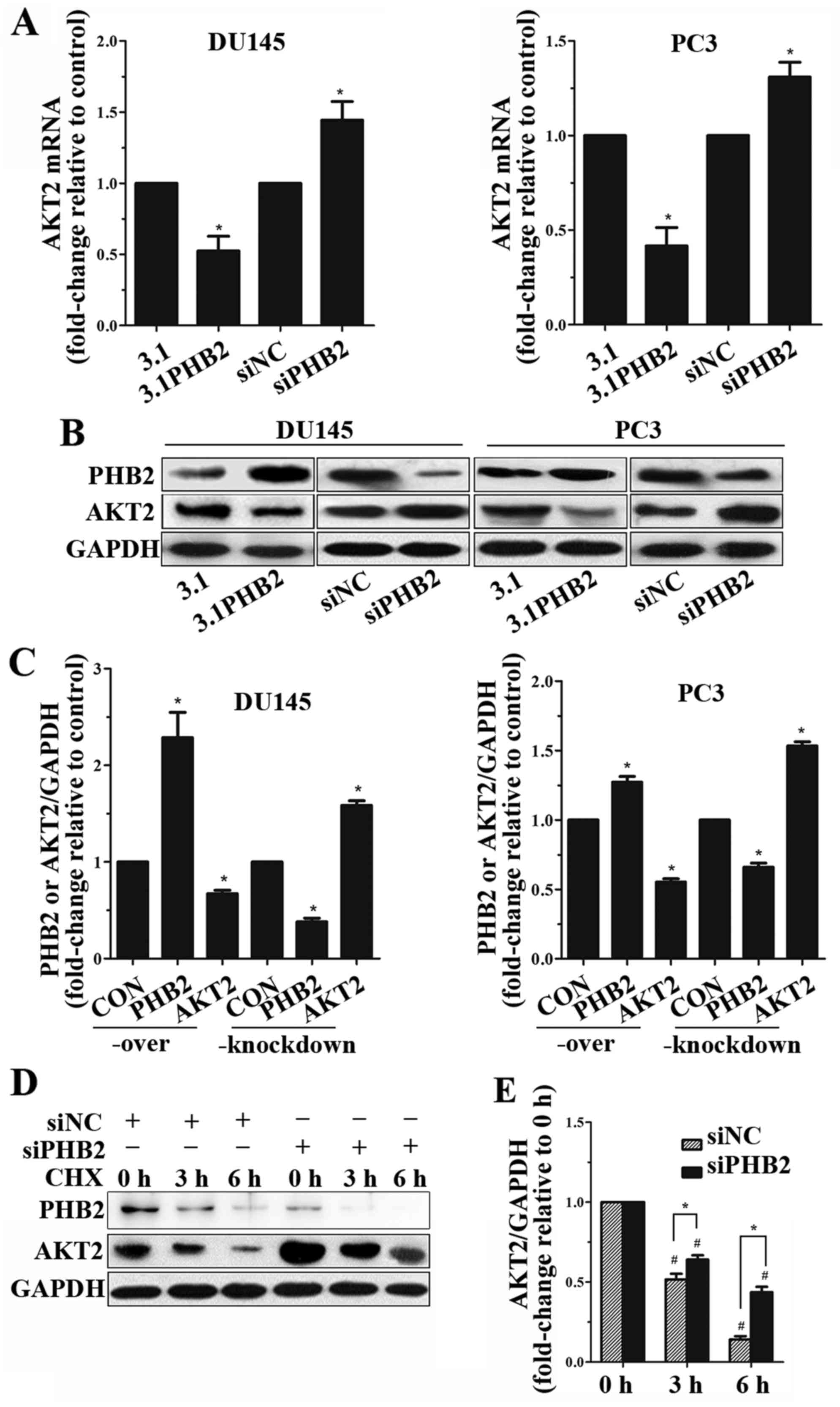
PHB2 inhibits AKT2 expression
To determine whether PHB2 could affect the
expression of AKT2, the expression levels of AKT2 were detected by
RT-qPCR and western blot analysis in DU145 and PC3 cells. As shown
in Fig. 4, the mRNA expression
levels of AKT2 were decreased in PC3 and DU145 cells with
PHB2 overexpression; however, they were increased in response to
PHB2 knockdown for 24 h (Fig.
4A). Notably, western blotting indicated that AKT2 expression
was markedly decreased in cells with PHB2 overexpression (Fig. 4B and C); these results suggested
that AKT2 expression was regulated by PHB2. In addition,
overexpression of PHB2 could down-regulate the protein expression
levels of AKT2 in BPH-1 cells (Fig.
3A and B).
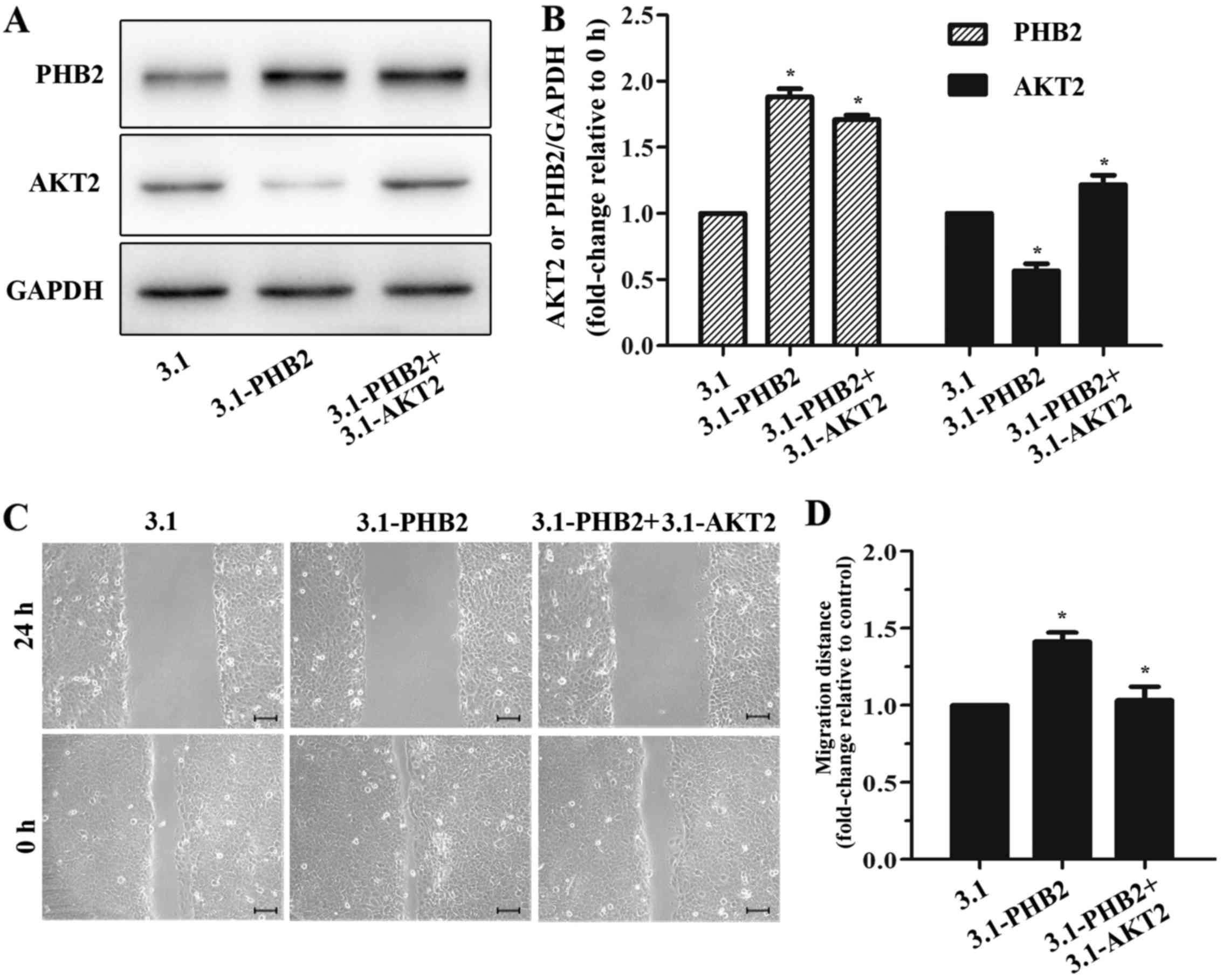
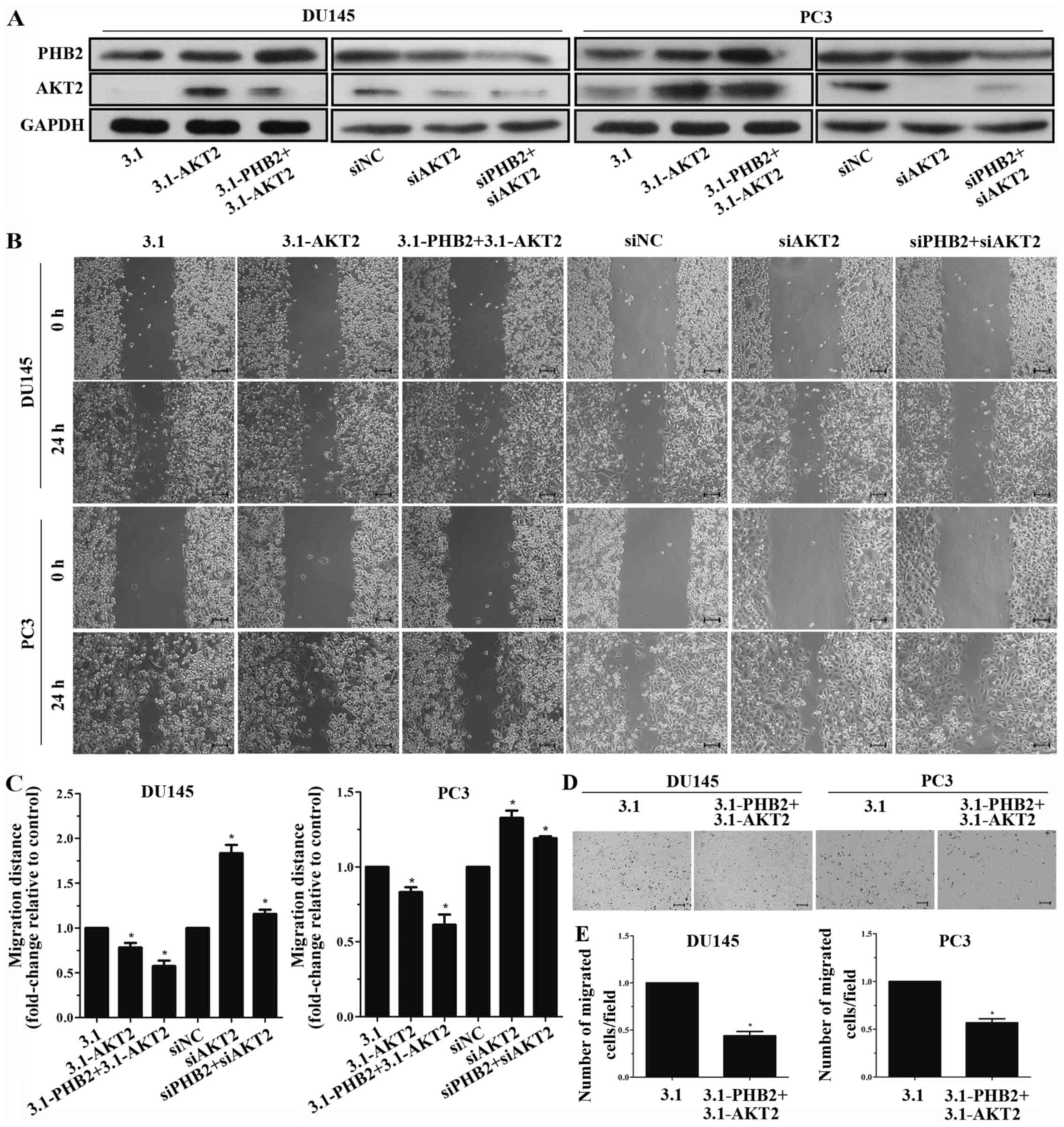
Since PHB2 may promote cell migration and inhibit
AKT2 expression, it was hypothesized that overexpression of AKT2
may reduce PHB2-induced migration. Therefore, cells were
cotransfected with PHB2 and AKT2 vectors/siRNAs, and the effects on
migration were detected. The protein expression levels of PHB2 and
AKT2 were significantly increased following cotransfection with
overexpression vectors, and were decreased following cotransfection
with siRNAs (Fig. 5A). Results of
the wound healing assay indicated that the migration of PC3 and
DU145 cells with PHB2 and AKT2 co-overexpression was significantly
decreased after 24 h compared with in the control group; however,
knockdown of both AKT2 and PHB2 enhanced migration compared with in
the control group within 24 h (Fig.
5B and C). The results of the wound healing assay were
confirmed by a Transwell migration assay (Fig. 5D and E). In addition, these
findings were verified in BPH-1 cells (Fig. 3C and D). In conclusion, AKT2 may
reduce PHB2-induced migration, thus suggesting that PHB2 promotes
the migration of PC3 and DU145 PCa cells by inhibiting the
expression of AKT2.
Discussion
Previous studies have reported that PHB2 is highly
expressed in various types of cancer, including liver, breast and
bladder cancer; and likely promotes or inhibits neoplastic
progression depending on PHB2 transcription, post-translational
modifications and translation in various tumor types (6,10,36). PHB2 is a multifunctional protein
that serves diverse functions depending on cellular localization.
The transportation of PHB2 into the nucleus and cytoplasm may be
crucial for its 'functional shift'. It has previously been reported
that knockdown of PHB2 abolishes IGFBP-6-induced Rh30 cell
migration (18); however, how
PHB2 participates in the progression of PCa remains unclear, and
reports that examine the association between PHB2 and cell
migration are currently lacking. We found that PHB2 expression was
increased in PCa cells with higher malignancy (37). Therefore, in the present study,
wound healing assays were used to examine cell migration; the
results indicated that cell migration was enhanced upon
transfection with a PHB2 overexpression plasmid in DU145, PC3 and
BPH-1 cells, whereas it was reduced upon PHB2 siRNA transfection.
These data suggested that PHB2 may be considered a positive
regulatory factor in PCa cell migration.
AKT2 is a promigratory kinase that is well defined
in breast cancer (38). However,
unlike in breast cancer, AKT2 functions as a negative regulator of
cell migration and invasion in PC3 PCa cells (23,24,39). Cariaga-Martinez et al
reported that AKT2 ablation increased migration of PC3 cells
(26). The present study
hypothesized that a relationship between PHB2 and AKT2 may exist.
The results indicated that overexpression of PHB2 reduced the
protein expression levels of AKT2. Conversely, knockdown of PHB2
increased the expression of AKT2 in DU145 and PC3 cells. In
addition, the stability of AKT2 expression was increased in PCa
cells with PHB2 knockdown. These findings suggested that
PHB2-mediated cell migration may be regulated by AKT2. Potentially,
AKT2 may counteract PHB2-induced migration, whereas PHB2 may
promote cell migration by inhibiting the expression of AKT2 in PC3
and DU145 cells. Furthermore, the present study indicated that
co-overexpression of PHB2 and AKT2 significantly decreased
migration of PCa cells, which supports our hypothesis. These
results are in agreement with those reported in muscle cells, in
which feedback regulation between AKT2 and PHB2 maintained a
dynamic balance (27).
At present, the mortality rate of PCa continues to
rise each year in China, thus underscoring the importance of
understanding the pathogenesis of PCa. PCa morbidity is affected by
complications that arise from tumor invasion, including bone and
lymphatic metastases, which can ultimately lead to mortality.
Therefore, understanding the process of PCa is of clinical
significance. While PHB2 is known to promote PCa progression, its
role in PCa migration has not been well studied. The present study
demonstrated that PHB2 could promote cell migration, and revealed
that AKT2 was negatively regulated by PHB2 in PCa. The present
study provides information regarding the function of PHB2 in the
pathological process of PCa migration. These findings may
potentially lay the foundation for designing targeted drugs that
have clinical value in the treatment of PCa.
Acknowledgments
The authors would like to thank Professor Helmut
Klocker (Innsbruck University School of Medicine) for the gift of
RWPE-1 and BPH-1 cells. The present study was funded by the
National Natural Science Foundation of China (nos. 81370859 and
81572784) and the Tianjin Municipal Science and Technology
Commission (nos. 12ZCDZSY17000 and 15JCYBJC50900).
References
|
1
|
Friedl P and Wolf K: Plasticity of cell
migration: A multiscale tuning model. J Cell Biol. 188:11–19. 2010.
View Article : Google Scholar :
|
|
2
|
Martin NE, Mucci LA, Loda M and Depinho
RA: Prognostic determinants in prostate cancer. Cancer J.
17:429–437. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kim SH, Hwang KA, Shim SM and Choi KC:
Growth and migration of LNCaP prostate cancer cells are promoted by
triclosan and benzophenone-1 via an androgen receptor signaling
pathway. Environ Toxicol Pharmacol. 39:568–576. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Quail DF and Joyce JA: Microenvironmental
regulation of tumor progression and metastasis. Nat Med.
19:1423–1437. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yilmaz M and Christofori G: Mechanisms of
motility in metastasizing cells. Mol Cancer Res. 8:629–642. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Thuaud F, Ribeiro N, Nebigil CG and
Désaubry L: Prohibitin ligands in cell death and survival: Mode of
action and therapeutic potential. Chem Biol. 20:316–331. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kowno M, Watanabe-Susaki K, Ishimine H,
Komazaki S, Enomoto K, Seki Y, Wang YY, Ishigaki Y, Ninomiya N,
Noguchi TA, et al: Prohibitin 2 regulates the proliferation and
lineage-specific differentiation of mouse embryonic stem cells in
mitochondria. PLoS One. 9:e815522014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yoshimaru T, Komatsu M, Matsuo T, Chen YA,
Murakami Y, Mizuguchi K, Mizohata E, Inoue T, Akiyama M, Yamaguchi
R, et al: Targeting BIG3-PHB2 interaction to overcome tamoxifen
resistance in breast cancer cells. Nat Commun. 4:24432013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chowdhury I, Garcia-Barrio M, Harp D,
Thomas K, Matthews R and Thompson WE: The emerging roles of
prohibitins in folliculogenesis. Front Biosci (Elite Ed).
4:690–699. 2012. View
Article : Google Scholar
|
|
10
|
Sievers C, Billig G, Gottschalk K and
Rudel T: Prohibitins are required for cancer cell proliferation and
adhesion. PLoS One. 5:e127352010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yurugi H and Rajalingam K: A role for
prohibitin in mast cell activation: Location matters. Sci Signal.
6:pe292013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cheng J, Gao F, Chen X, Wu J, Xing C, Lv
Z, Xu W, Xie Q, Wu L, Ye S, et al: Prohibitin-2 promotes
hepatocellular carcinoma malignancy progression in hypoxia based on
a label-free quantitative proteomics strategy. Mol Carcinog.
53:820–832. 2014. View
Article : Google Scholar
|
|
13
|
Pabona JM, Velarde MC, Zeng Z, Simmen FA
and Simmen RC: Nuclear receptor co-regulator Krüppel-like factor 9
and prohibitin 2 expression in estrogen-induced epithelial cell
proliferation in the mouse uterus. J Endocrinol. 200:63–73. 2009.
View Article : Google Scholar
|
|
14
|
Van Aken O, Whelan J and Van Breusegem F:
Prohibitins: Mitochondrial partners in development and stress
response. Trends Plant Sci. 15:275–282. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Polier G, Neumann J, Thuaud F, Ribeiro N,
Gelhaus C, Schmidt H, Giaisi M, Köhler R, Müller WW, Proksch P, et
al: The natural anticancer compounds rocaglamides inhibit the
Raf-MEK-ERK pathway by targeting prohibitin 1 and 2. Chem Biol.
19:1093–1104. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhu B, Zhai J, Zhu H and Kyprianou N:
Prohibitin regulates TGF-β induced apoptosis as a downstream
effector of Smad-dependent and -independent signaling. Prostate.
70:17–26. 2010. View Article : Google Scholar
|
|
17
|
Chen RX, Song HY, Dong YY, Hu C, Zheng QD,
Xue TC, Liu XH, Zhang Y, Chen J, Ren ZG, et al: Dynamic expression
patterns of differential proteins during early invasion of
hepatocellular carcinoma. PLoS One. 9:e885432014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fu P, Yang Z and Bach LA: Prohibitin-2
binding modulates insulin-like growth factor-binding protein-6
(IGFBP-6)-induced rhabdomyosarcoma cell migration. J Biol Chem.
288:29890–29900. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vivanco I, Chen ZC, Tanos B, Oldrini B,
Hsieh WY, Yannuzzi N, Campos C and Mellinghoff IK: A
kinase-independent function of AKT promotes cancer cell survival.
eLife. 3:e037512014. View Article : Google Scholar
|
|
20
|
Kang W, Tong JHM, Lung RWM, Dong Y, Yang
W, Pan Y, Lau KM, Yu J, Cheng AS and To KF: let-7b/g silencing
activates AKT signaling to promote gastric carcinogenesis. J Transl
Med. 12:2812014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liang JW, Shi ZZ, Shen TY, Che X, Wang Z,
Shi SS, Xu X, Cai Y, Zhao P, Wang CF, et al: Identification of
genomic alterations in pancreatic cancer using array-based
comparative genomic hybridization. PLoS One. 9:e1146162014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chin YR, Yuan X, Balk SP and Toker A:
PTEN-deficient tumors depend on AKT2 for maintenance and survival.
Cancer Discov. 4:942–955. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Virtakoivu R, Pellinen T, Rantala JK,
Perälä M and Ivaska J: Distinct roles of AKT isoforms in regulating
β1-integrin activity, migration, and invasion in prostate cancer.
Mol Biol Cell. 23:3357–3369. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhou GL, Tucker DF, Bae SS, Bhatheja K,
Birnbaum MJ and Field J: Opposing roles for Akt1 and Akt2 in
Rac/Pak signaling and cell migration. J Biol Chem. 281:36443–36453.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fortier AM, Van Themsche C, Asselin E and
Cadrin M: Akt isoforms regulate intermediate filament protein
levels in epithelial carcinoma cells. FEBS Lett. 584:984–988. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cariaga-Martinez AE, López-Ruiz P,
Nombela-Blanco MP, Motiño O, González-Corpas A, Rodriguez-Ubreva J,
Lobo MV, Cortés MA and Colás B: Distinct and specific roles of AKT1
and AKT2 in androgen-sensitive and androgen-independent prostate
cancer cells. Cell Signal. 25:1586–1597. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Héron-Milhavet L, Mamaeva D, Rochat A,
Lamb NJ and Fernandez A: Akt2 is implicated in skeletal muscle
differentiation and specifically binds Prohibitin2/REA. J Cell
Physiol. 214:158–165. 2008. View Article : Google Scholar
|
|
28
|
Gardner S, Anguiano M and Rotwein P:
Defining Akt actions in muscle differentiation. Am J Physiol Cell
Physiol. 303:C1292–C1300. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Matheny RW Jr and Adamo ML: Effects of
PI3K catalytic subunit and Akt isoform deficiency on mTOR and
p70S6K activation in myoblasts. Biochem Biophys Res Commun.
390:252–257. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rhodes DR, Kalyana-Sundaram S, Mahavisno
V, Varambally R, Yu J, Briggs BB, Barrette TR, Anstet MJ,
Kincead-Beal C, Kulkarni P, et al: Oncomine 3.0: genes, pathways,
and networks in a collection of 18,000 cancer gene expression
profiles. Neoplasia. 9:166–180. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rhodes DR, Yu J, Shanker K, Deshpande N,
Varambally R, Ghosh D, Barrette T, Pandey A and Chinnaiyan AM:
ONCOMINE: a cancer microarray database and integrated data-mining
platform. Neoplasia. 6:1–6. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang Z, Wang L, Mei M, Zhu Y, Du X, Lee
C, Park I, Zhang J and Shi J: Both nongenomic and genomic effects
are involved in estradiol's enhancing the phenotype of smooth
muscle cells in cultured prostate stromal cells. Prostate.
70:317–332. 2010.
|
|
33
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
34
|
Héron-Milhavet L, Franckhauser C, Rana V,
Berthenet C, Fisher D, Hemmings BA, Fernandez A and Lamb NJ: Only
Akt1 is required for proliferation, while Akt2 promotes cell cycle
exit through p21 binding. Mol Cell Biol. 26:8267–8280. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yarrow JC, Perlman ZE, Westwood NJ and
Mitchison TJ: A high-throughput cell migration assay using scratch
wound healing, a comparison of image-based readout methods. BMC
Biotechnol. 4:21. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Artal-Sanz M and Tavernarakis N:
Prohibitin and mitochondrial biology. Trends Endocrinol Metab.
20:394–401. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Du XL, Yu L, Ning ZC, et al: Estrogen
receptor inhibitor, REA, have an effect on proliferation and
migration in prostate cancer cells. Acta Scientiarum Naturalium
Universitatis Nankaiensis. 46:36–43. 2013.
|
|
38
|
Irie HY, Pearline RV, Grueneberg D, Hsia
M, Ravichandran P, Kothari N, Natesan S and Brugge JS: Distinct
roles of Akt1 and Akt2 in regulating cell migration and
epithelial-mesenchymal transition. J Cell Biol. 171:1023–1034.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dillon RL, Marcotte R, Hennessy BT,
Woodgett JR, Mills GB and Muller WJ: Akt1 and akt2 play distinct
roles in the initiation and metastatic phases of mammary tumor
progression. Cancer Res. 69:5057–5064. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Welsh JB, Sapinoso LM, Su AI, Kern SG,
Wang-Rodriguez J, Moskaluk CA, Frierson HF Jr and Hampton GM:
Analysis of gene expression identifies candidate markers and
pharmacological targets in prostate cancer. Cancer Res.
61:5974–5978. 2001.PubMed/NCBI
|
|
41
|
Singh D, Febbo PG, Ross K, Jackson DG,
Manola J, Ladd C, Tamayo P, Renshaw AA, D'Amico AV, Richie JP, et
al: Gene expression correlates of clinical prostate cancer
behavior. Cancer Cell. 1:203–209. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lapointe J, Li C, Higgins JP, van de Rijn
M, Bair E, Montgomery K, Ferrari M, Egevad L, Rayford W, Bergerheim
U, Ekman P, et al: Gene expression profiling identifies clinically
relevant subtypes of prostate cancer. Proc Natl Acad Sci USA.
101:811–816. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Varambally S, Yu J, Laxman B, Rhodes DR,
Mehra R, Tomlins SA, Shah RB, Chandran U, Monzon FA, Becich MJ, Wei
JT, et al: Integrative genomic and proteomic analysis of prostate
cancer reveals signatures of metastatic progression. Cancer Cell.
8:393–406. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Liu P, Ramachandran S, Ali Seyed M,
Scharer CD, Laycock N, Dalton WB, Williams H, Karanam S, Datta MW,
Jaye DL and Moreno CS: Sex-determining region Y box 4 is a
transforming oncogene in human prostate cancer cells. Cancer Res.
66:4011–4019. 2006. View Article : Google Scholar : PubMed/NCBI
|