Introduction
DNA damage is an important etiological factor in
lung cancer (1), and various
types of DNA damage exist, including nucleotide alterations
(deletions, insertions and substitutions of nucleotides),
single-strand breaks and double-strand breaks (2). When DNA damage occurs, a set of DNA
repair pathways are activated (3–5).
Among these pathways, the base excision repair (BER) pathway is
responsible for repairing the majority of DNA damage caused by
alkylation and oxidative stress (4,6).
Human apurinic/apyrimidinic (AP) endonuclease 1
(APE1), also known as APE, APEX and Ref-1, is a ubiquitous
multifunctional protein, which is associated with BER and redox
activities. APE1 is a rate-limiting enzyme in the BER pathway
(7–9). When base damage occurs, glycosylases
in the cell excise the damaged base to generate an AP, or abasic,
site. Unlike other glycosylases, APE1 is the only enzyme that can
process the BER abasic site; APE1 hydrolyzes the phosphodiester
backbone 5′ to the AP site, creating a normal 3′ hydroxyl terminus
and an abasic 5′ deoxyribose phosphate terminus. During this
process, APE1 also recruits various polymerases and ligases to
participate in the repair. Mammalian APE1 also affects numerous DNA
transcription factors, including hypoxia inducible factor (HIF)-1α,
activator protein (AP)-1, nuclear factor (NF)-κB and p53 (10), and by doing so, indirectly
contributes to DNA repair.
APE1 is the only DNA repair protein known to have
redox regulatory activity (8,10),
and is responsible for 95% of cellular endonuclease activity
(7,8). APE1 is able to regulate the reducing
state of amino acid residues at key sites of transcription factors,
and therefore participates in various basic cellular events,
including proliferation, differentiation, apoptosis and
transformation. The expression levels, aberrant subcellular
localization and the patterns of post-translational modification of
APE1 have been implicated in chemo- and radioresistance (11,12), and are associated with poor
prognosis in numerous types of cancer, including non-small cell
lung cancer (NSCLC) (13–22). Serum APE1 autoantibody has been
proposed as a potential tumor marker and predictor of responses to
chemotherapy in patients with NSCLC (23).
Lung cancer accounts for 13% of all new cases of
malignant tumors and 19.4% of cases of malignant tumor-associated
mortality (24). The latest
cancer statistics have revealed increasing morbidity and mortality
of lung cancer in developed and developing countries (25,26). Approximately 85% of all patients
with lung cancer have NSCLC. In patients who are not a candidate
for surgery, therapeutic approaches mainly include chemotherapy and
targeted therapy (27–29). However, despite advances in
treatment, the five-year survival rate remains <17.4% (30).
Under the theoretical framework of traditional
Chinese medicine (TCM), lung cancer is believed to reflect external
invasion, which may lead to disharmonious lung functions, and
ultimately results in the obstruction of lung Qi and blood stasis.
Previous studies regarding TCM and lung cancer (31–36) have suggested that supplementing
the center, and boosting Qi and anti-inflammatory function, may
prolong patient survival and improve the quality of life (QOL) for
patients with NSCLC. The major ingredients of original Bu-Fei
decoction (BFD) are Milkvetch root (Huang-Qi), Radix Asteris
(Zi-Yuan) and Cortex Mori Radicis (Sang-Bai-Pi). In addition,
modified BFD (MBFD) lso contains Hedyotic diffusa
(Bai-Hua-She-She-Cao), Duchesnea (She-Mei) and Scutellaria
barbata (Ban-Zhi-Lian). He et al demonstrated that BFD
attenuated TGF-β1-mediated epithelial-mesenchymal transition in
lung cancer A549 cells via decreasing canonical Smad signaling
pathway (37). BFD has been shown
to exhibit immunomodulatory effects and possess anti-tumor
activities (38). The present
study aimed to examine the potential antitumor effects of BFD and
MBFD on representative NSCLC cell lines and a mouse xenograft
model.
Materials and methods
Cell culture
The H1975 and H292 human lung cancer cell lines, and
the BEAS-2B human lung bronchial epithelial cell line, were
purchased from American Type Culture Collection (Manassas, VA,
USA). The cells were grown in RPMI-1640 medium (Thermo Fisher
Scientific, Inc.) containing 10% fetal bovine serum (FBS; Gibco;
Thermo Fisher Scientific, Inc.) and maintained at 37°C in an
atmosphere containing 5% CO2.
BFD and MBFD
The BFD decoction is composed of 6 medicinal herbs:
milkvetch root (Huang-Qi), Radix asteris (Zi-Yuan),
Cortex Mori Radicis (Sang-Bai-Pi), Rehmannia
glutinosa (Di-Huang), Codonopsis pilosula (Dang-shen)
and Schisandra chinensis (Wu-Wei-Zi), Hedyotic
diffusa (Bai-Hua-She-She-Cao), Duchesnea (She-Mei) and
Scutellaria barbata (Ban-Zhi-Lian) were added to make MBFD.
The components of BFD and MBFD were converted into formula granules
by Beijing Tcmages Pharmaceuticals Co., Ltd. (Beijing, China) at a
receiving rate of 18.8 and 14.1%, respectively. The quality of the
BFD and MBFD granules was monitored by Fourier transform infrared
spectroscopy (model IRPRestige-21; Shimadzu Corp., Kyoto, Japan).
Prior to use, the formula granules were dissolved in deionized
water at about 50°C, centrifuged at 13,800 × g for 30 min to remove
drug sediment, and sterilized by filtration through a 0.22
µm membrane (EMD Millipore, Billerica, MA, USA); finally,
the solutions were stored at −80°C. The concentrations of BFD and
MBFD indicated in the subsequent text denote crude drug
concentrations.
Cell proliferation assay
Cells were resuspended in RPMI-1640 at a density of
1×104 cells/well in 96-well plates (Costar; Corning
Incorporated, Corning, NY, USA). Drug exposure was conducted after
culturing for 24 h and were treated with the drugs alongside 1% FBS
in an atmosphere containing 5% CO2. H1975, H292 and
BEAS-2B cells were treated with BFD (0–30 mg/ml) or MBFD (0–15
mg/ml) for 24 h. β-actin (#4790; dilution, 1:10,000; Cell Signaling
Technology, Inc., Danvers, MA, USA) was used as a loading control
in cell proteins and GAPDH (#5174; dilution, 1:10,000; Cell
Signaling Technology, Inc.) in tissue proteins. Cells were then
incubated with MTT at a final concentration of 0.5 mg/ml
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at 37°C for 4 h,
then MTT was replaced by 150 µl dimethyl sulfoxide,
incubated for 15 min at room temperature. Absorbance (optical
density) was measured at 570 nm using a microplate reader (Model
680; Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Flow cytometry and cell cycle
analysis
Cells were resuspended in RPMI-1640 at a density of
3×105 cells/well in 6-well plates (Costar; Corning
Incorporated). Drug exposure was conducted after culturing for 24 h
and at 1% FBS. H1975 cells were treated with BFD (0–30 mg/ml) or
MBFD (0–15 mg/ml) for 24 h; H292 cells were treated with BFD (0–30
mg/ml) or MBFD (0–20 mg/ml) for 24 h at 37°C in an atmosphere
containing 5% CO2. Cells were then collected, fixed with
cold 75% ethanol and stored at −20°C for ≥24 h. Subsequently, after
washing twice with cold PBS, cells were incubated with propidium
iodide (PI)/RNase staining buffer (BD Biosciences, San Diego, CA,
USA) for 15 min at room temperature. DNA content was analyzed by
flow cytometry using BD Accuri C6 (version 1.0.264.15; BD
Biosciences). The percentage of cells in the various phases of the
cell cycle was determined using ModFit LT 4.1 software (BD
Biosciences).
Annexin V/PI staining assay
Annexin V/PI staining was performed using an
apoptosis detection kit (Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan). Following treatment, cells were collected and
dissolved in 100 µl Annexin V binding buffer
(5×105 cells/ml). To 100 µl of each sample, 5
µl Annexin V and 5 µl PI were added, and the samples
were incubated for 15 min at room temperature. Analysis of
apoptosis was performed using BD Accuri C6. Annexin
V-negative/PI-negative cells were identified as viable cells,
Annexin V-positive/PI-negative cells were recognized as early
apoptotic cells, and Annexin V-positive/PI-positive cells were
identified as late apoptotic/dead cells.
Comet assay
Cells were suspended in ice-cold 1X PBS
(Ca2+- and Mg2+-free) to a concentration of
2×105 cells/ml. DNA strand breaks were evaluated using
Trevigen CometAssay® kit (Trevigen, Gaithersburg, MD,
USA). Cells (2×105 cells/ml) were mixed with molten
LMAgarose at a volume ratio of 1:10 and the mixture was immediately
evenly spread onto comet slides, which were incubated at 4°C in the
dark for 10 min. The slides were then transferred to prechilled
lysis solution for 60 min at 4°C.
For the alkaline comet assay, slides were incubated
in 300 mM NaOH containing 1 mM EDTA (pH >13) for 20 min at room
temperature in the dark, and were electrophoresed at 1 V/cm and 300
mA for 40 min. After extensive rinsing, slides were incubated in
70% ethanol for 5 min and dried. Finally, DNA was stained with
SYBR-Green I dye (1:10,000 in Tris-EDTA buffer, pH 7.5; Trevigen)
for 20 min at 4°C. Images were captured using a fluorescence
microscope (TCS SP5; Leica Microsystems GmbH, Wetzlar,
Germany).
For the neutral comet assay, samples were immersed
in 50 ml 1X neutral electrophoresis buffer for 30 min at 4°C,
separated by electrophoresis at 1 V/cm for 40 min, and transferred
to DNA precipitation solution for 30 min at room temperature. After
drying, DNA was stained with SYBR-Green I dye (1:10,000 in
Tris-EDTA buffer, pH 7.5; Trevigen) for 20 min at 4°C. Images were
captured using a fluorescence microscope (TCS SP5; Leica
Microsystems GmbH).
Comet analysis
The percentage of DNA in the tail (the percentage of
total cell DNA found in the tail) and tail moment (the amount of
DNA in the tail combined with the distance of migration) are common
descriptors of DNA damage determined by alkaline comet and neutral
comet assays, respectively. At least 50 randomly selected cells
were analyzed for each slide using TriTek CometScore™ Freeware v1.5
image analysis software (TriTek Corporation, Sumerduck, VA,
USA).
Small interfering (si)RNA and plasmid
transfection
Cells were transfected with APE1 siRNA [(5′→3′):
GTTGGCGCCTTGATTACTT] or a scrambled RNA control (Guangzhou Ribobio
Co., Ltd., Guangzhou, China) at a final concentration of 80 pM
using Lipofectamine® 2000 (Invitrogen, Carlsbad, CA,
USA). pReceiver-M98 plasmid containing the APE1 gene (cat. no.
EX-C0496-M98-5) and an empty plasmid were synthesized by
GeneCopoeia (Rockville, MD, USA). Transfection efficiency was
confirmed by western blot analysis. Cells were re-suspended in
RPMI-1640 at a density of 2×105 cells/well in 24-well
plates (Costar; Corning Incorporated), cultured for 24 h at 37°C in
an atmosphere of 5% CO2. Before transfection, dilution
of 0.8 µg DNA in 50 µl Opti-MEM medium without serum
(Gibco, Grandsland, NY, USA) and dilution of 2 µl
Lipofectamine® 2000 in 50 µl Opti-MEM Medium
without serum was made, and incubation for 5 min at room
temperature followed. The diluted DNA and diluted
Lipofectamine® 2000 (total volume =100 µl) were
combined and incubation for 20 min at room temperature followed. A
total of 100 µl of complexes was then added to each well
containing cells and 400 µl RPMI-1640 medium; cells were
incubated at 37°C in an atmosphere of 5% CO2 for 24 h.
Same as the plasmid transfection, 40 pmol siRNA and 1 µl
Lipofectamine® 2000 were used in the siRNA transfection
protocol. Transfection was conducted 48 h prior to drug treatment
with BFD or MBFD.
RNA isolation, polymerase chain reaction
(PCR) and reverse transcription-quantitative PCR (RT-qPCR)
analysis
Total RNA was extracted from cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). RT was performed using the TransScript First-Strand cDNA
synthesis supermix (Beijing Transgen Biotech Co., Ltd., Beijing,
China) according to the manufacturer’s protocol. The concentration
and quality of extracted RNA were verified using a NanoDrop 2000
(NanoDrop; Thermo Fisher Scientific, Inc., Wilmington, DE, USA). RT
was performed using the TransScript First-Strand cDNA synthesis
supermix (Beijing Transgen Biotech Co., Ltd., Beijing, China). PCR
was conducted according to the standard protocol of EasyTag
supermix (Beijing Transgen Biotech Co., Ltd.) and DNA amplification
was performed using a PCR thermocycler (S1000TM, Thermal Cycler;
Bio-Rad Laboratories, Inc.). The cycling conditions were as
follows: One cycle at 95°C for 5 min, followed by 30 cycles at 95°C
for 30 sec, 60°C for 30 sec and 72°C for 35 sec. The amplification
products were separated by 1% agarose gel electrophoresis and were
analyzed using an ultraviolet transilluminator (GelDoc™ EZ; Bio-Rad
Laboratories, Inc.). RT-qPCR was performed using a SYBR-Green qPCR
Supermix (Applied Biosystems; Thermo Fisher Scientific, Inc.) on an
ABI Prism 7500 sequence detection system (Applied Biosystems;
Thermo Fisher Scientific, Inc.). The cycling conditions were as
follows: One cycle at 35°C for 10 min, followed by 40 cycles at
95°C for 30 sec, 60°C for 30 sec and 72°C for 35 sec. The specific
primers designed for PCR were synthesized by Sangon Biotech Co.,
Ltd. (Shanghai, China); the sequences were as follows (5′→3′):
APE1, forward GAGTAAGACGGCCGCAAAGAAAAA, reverse
CCGAAGGAGCTGACCAGTATTGAT; and GAPDH, forward GGTGAAGGTCGGTGTGAACG
and reverse CTCGCTCCTGGAAGATGGTG. Cells were resuspended in
RPMI-1640 at a density of 3×105 cells/well in 6-well
plates (Costar; Corning Incorporated). Drug exposure was conducted
after culture for 24 h and at 1% FBS. H1975 cells were treated with
BFD (0–30 mg/ml) or MBFD (0–15 mg/ml) for 24 h; H292 cells were
treated with BFD (0–30 mg/ml) or MBFD (0–20 mg/ml) for 24 h.
Following treatment, the cells were washed with ice-cold PBS and
lysed with RIPA buffer (CWBIO, Beijing, China). Protein
concentration was detected using a BCA protein assay kit (Thermo
Fisher Scientific, Inc.). The specificity of amplification was
confirmed via melting curve analysis. The mRNA expression levels of
APE1 relative to GAPDH were calculated using the ΔΔCq method, as
described previously (39).
Western blot analysis
Samples (15 µg protein) were separated by 12%
SDS-PAGE and were transferred to polyvinylidene difluoride
membranes (EMD Millipore). The membranes were then blocked with 5%
skim milk for 1 h at room temperature, and were incubated with APE1
antibody (ab92744; dilution, 1:20,000; Abcam, Cambridge, MA, USA)
overnight at 4°C. The membranes were washed with Tris-buffered
saline containing 0.05% Tween and were then incubated with
horseradish peroxidase-conjugated secondary antibody (cat. no.
129736; dilution, 1:10,000; ZSGB-BIO Co., Ltd, Beijing, China) for
1 h at room temperature. Finally the blots were visualized using
chemiluminescence (EMD Millipore). β-actin (#4790; dilution,
1:10,000; Cell Signaling Technology, Inc.) was used as a loading
control. Semi-quantitative analysis of the blots was performed
using ImageJ2x (National Institutes of Health, Bethesda, MD, USA).
Protein concentration was determined using the bicinchoninic acid
method prior to western blotting.
Nude mouse xenograft model
Female BALB/c nude mice (n=50; age, 6–8 weeks;
weight, 22 g; National Institutes for Food and Drug Control,
Beijing, China) were housed in environmentally controlled cabinets
(temperature, 23±2°C; 12-h light/dark cycle; relative humidity,
50%; normal food and water) under specific pathogen-free conditions
for 7 days prior to experimentation. Protocols were performed in
accordance with the US Animal Welfare guidelines (40), and the present study was approved
by the Institutional Animal Care and Use Committee of the First
Hospital Affiliated to PLA General Hospital. H1975 cells
(3×106 density) were suspended in cold PBS mixed with
Matrigel (BD Biosciences) at a volume ratio of 1:1, and were
subcutaneously inoculated into the right flank of nude mice.
The weight of tumor-bearing mice was measured and
tumor volume was calculated according to the following formula:
Tumor volume (mm3) = (length × width2)/2. The
daily dosage for nude mice experiments (BFD and MBFD at 15.17 and
24.42 g/kg, respectively) was based on human use (1.23 and 1.98
g/kg, respectively), as described previously (41). When tumor volume reached 50–100
mm3, mice were randomly divided into five groups
(n=10/group): Control, BFD (30 g/kg), BFD (60 g/kg), MBFD (48 g/kg)
and MBFD (96 g/kg). Drugs were orally administered twice daily for
21 consecutive days. After 21 days, the mice were sacrificed. The
blood samples (0.5 ml) were collected from foss orbital veins into
heparinized polythene tubes. Tumors were resected and weighed after
the mice were sacrificed. Tumor tissues were fixed in formalin and
in ice-cold PIPA lysate buffer for protein detection.
ELISA assay
The protein concentration of APE1 was determined in
mouse plasma samples using a commercial ELISA kit (Beijing
Keyingmei Technology, Beijing, China). The blood samples (0.5 ml)
were collected from foss orbital veins into heparinized polythene
tubes. After placement at 4°C for 2 h, and centrifugation at 2,500
rpm for 15 min, the supernatant was collected to a new tube for
ELISA.
Statistical analysis
Data are presented as the means ± standard error of
mean of at least three independent experiments. Half maximal
inhibitory concentration (IC50) was calculated using
GraphPad Prism 5.0.1 software (GraphPad Software, Inc., La Jolla,
CA, USA). The analysis of variance (ANOVA) was used to compare gene
expression levels between the different groups; other comparisons
between two groups were analyzed using Student’s t-test. All data
analyses were conducted using SPSS statistical software 19.0 (IBM
Corp., Armonk, NY, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Effects of BFD and MBFD on cell
proliferation
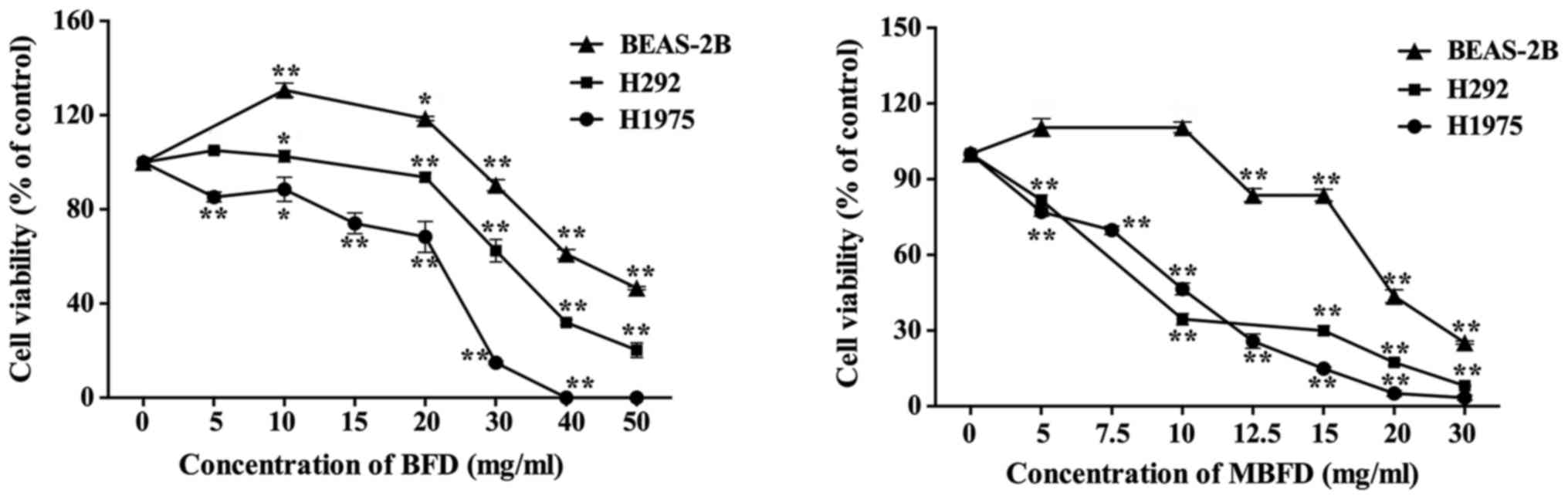
BFD and MBFD inhibited the proliferation of H1975
and H292 cells in a dose-dependent manner (Fig. 1). The IC50 values of
BFD were 21.66±1.01, 36.84±0.56 and 46.02±0.89 mg/ml in H1975, H292
and BEAS-2B cells, respectively. The IC50 values of MBFD
were 9.12±0.96, 12.66±0.56 and 18.1±1.08 mg/ml in H1975, H292 and
BEAS-2B cells, respectively. These results indicated that BEAS-2B
cells are less sensitive to BFD and MBFD compared with the two
cancer cell lines (P<0.01).
Effects of BFD and MBFD on cell cycle
progression and cell apoptosis
The results of flow cytometry experiments
demonstrated that BFD and MBFD have no effects on cell cycle arrest
and apoptosis of H1975 and H292 cells (data not shown).
Effects of BFD and MBFD on DNA
repair
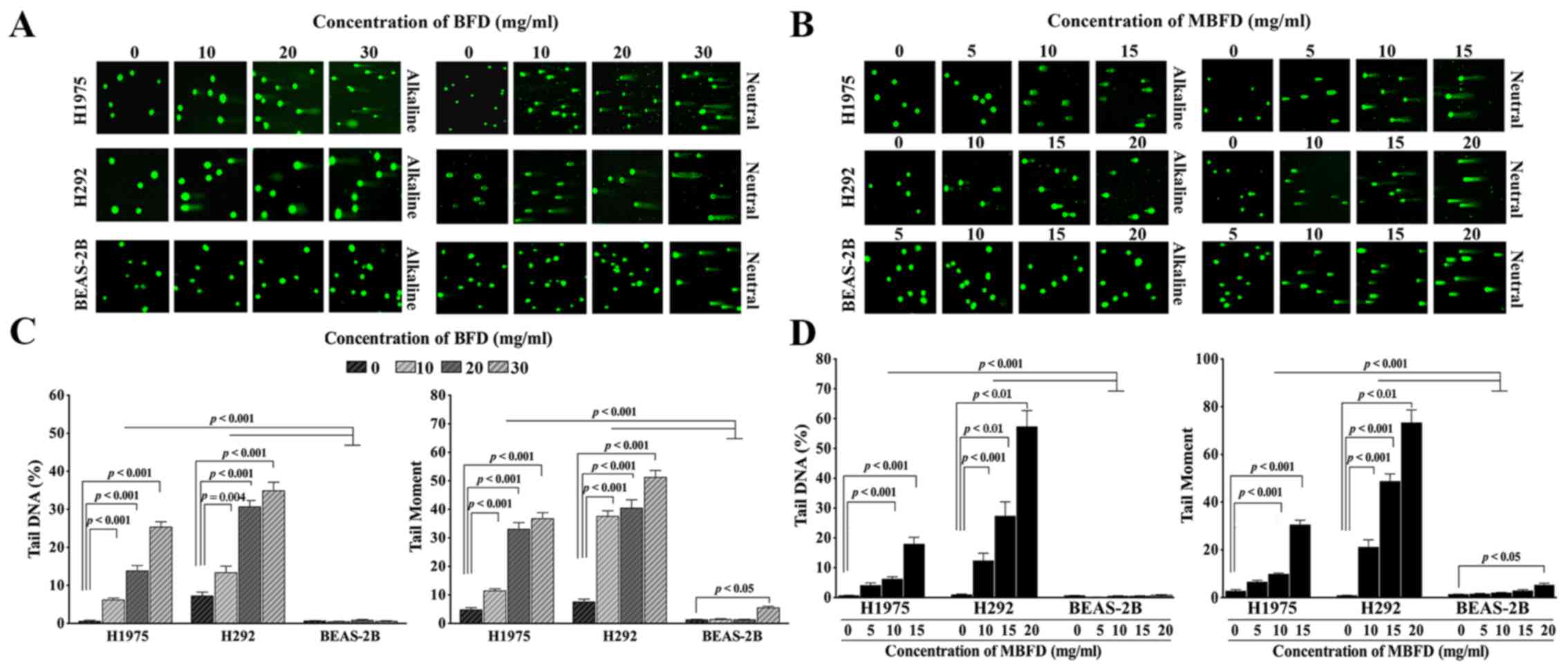
Treatment with BFD and MBFD increased DNA migration
from the nucleus in a concentration-dependent manner, thus
suggesting that DNA repair is suppressed. The migration of DNA from
the nucleus formed a ‘comet tail’ (Fig. 2A and B). Tail DNA increased by
11.14-, 25.04- and 45.93-fold, and 1.85-, 4.25- and 4.85-fold in
H1975 and H292 cells, respectively, by BFD at 10, 20 and 30 mg/ml
(P<0.001 vs. control) (Fig.
2C). BFD did not affect tail DNA in BEAS-2B cells at this
concentration range. Tail moment increased by 2.38-, 6.86- and
7.62-fold, and 4.98-, 5.36- and 6.79-fold in H1975 and H292 cells,
respectively, by BFD at 10, 20 and 30 mg/ml (P<0.001). In
BEAS-2B cells, BFD increased tail moment only when used at the
highest concentration (30 mg/ml).
MBFD also increased the extent of DNA damage in
H1975 and H292 cells. Tail DNA was increased by 6.37-, 9.72- and
28.21-fold in H1975 cells in response to 5, 10 and 15 mg/ml MBFD,
respectively (P<0.001) (Fig.
2D). In H292 cells, tail DNA was increased by 13.77-, 30.56-
and 64.07-fold in response to 10 (P<0.001), 15 (P<0.01) and
20 mg/ml (P<0.01) MBFD, respectively (P<0.001). BEAS-2B cells
were not affected by MBFD. In H1975 cells, tail moment was
increased by 2.44-, 3.69- and 11.5-fold in response to 5, 10 and 15
mg/ml MBFD, respectively. In H292 cells, tail moment was increased
by 28.14-, 64.89- and 97.68-fold in response to 10, 15 and 20 mg/ml
MBFD, respectively. In BEAS-2B cells, tail moment was increased
only when cells were treated with the highest concentration of
MBFD.
Effects of APE1 manipulation
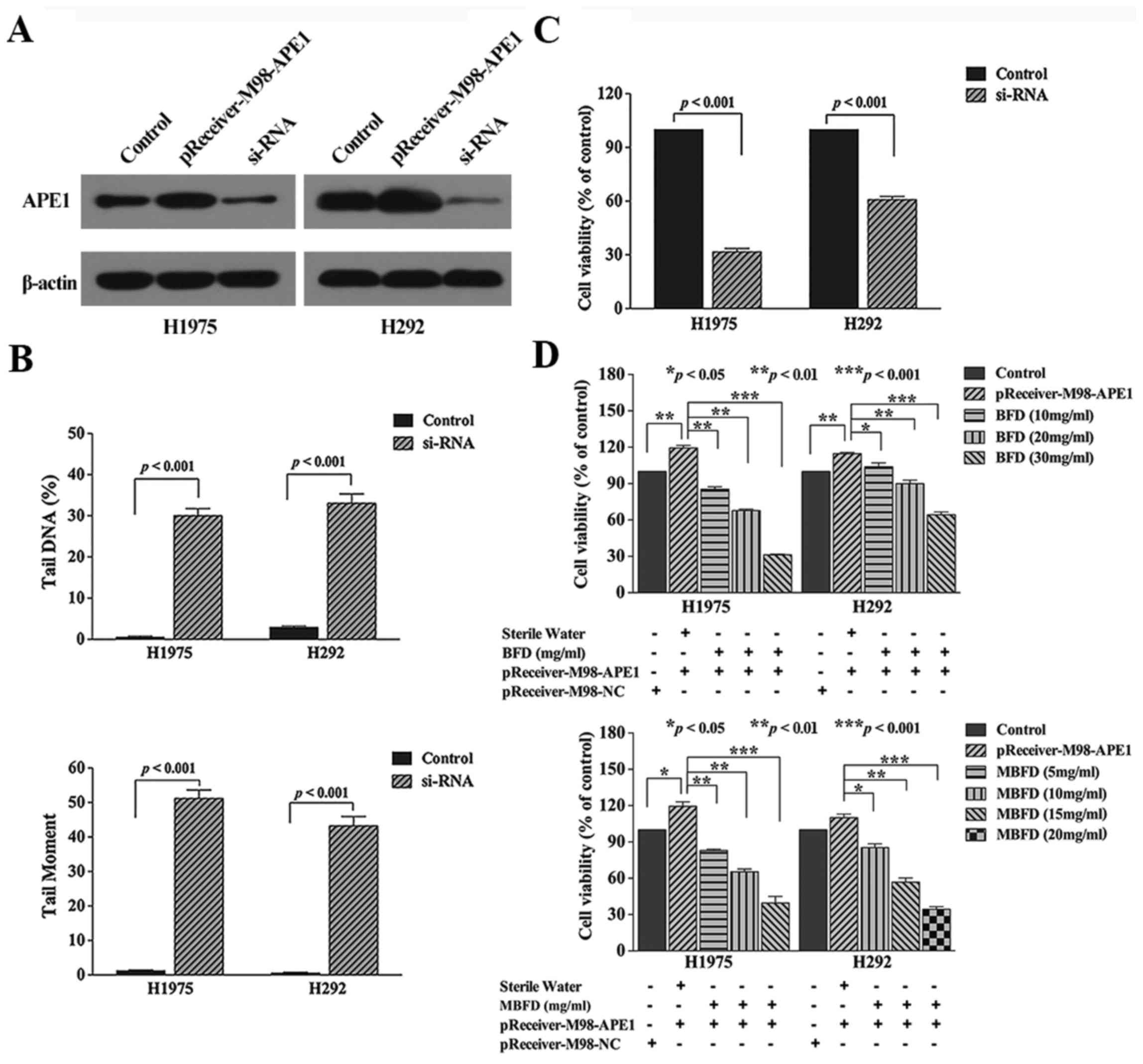
Knockdown of APE1 induced DNA damage and inhibited
proliferation of the two cancer cell lines (Fig. 3B, and C). Conversely,
overexpression of APE1 with a pReceiver-M98 plasmid increased cell
proliferation (Fig. 3D). Compared
with in the transfection group, treatment with BFD at 10, 20 and 30
mg/ml concentration inhibited H1975 cell proliferation by 27.96
(P<0.01), 41.54 (P<0.01) and 73.4% (P<0.001),
respectively, and inhibited H292 cell proliferation by 13.6
(P<0.05), 25.2 (P<0.01) and 46.16% (P<0.001),
respectively. In addition, treatment with MBFD at 5, 10 and 15
mg/ml concentrations inhibited H1975 cell proliferation by 29.01
(P<0.01), 47.21 (P<0.01) and 66.68% (P<0.001),
respectively, and MBFD at 10, 15 and 20 mg/ml concentrations
inhibited H292 cell proliferation by 24.06 (P<0.05), 48.76
(P<0.01) and 66.49% (P<0.001), respectively.
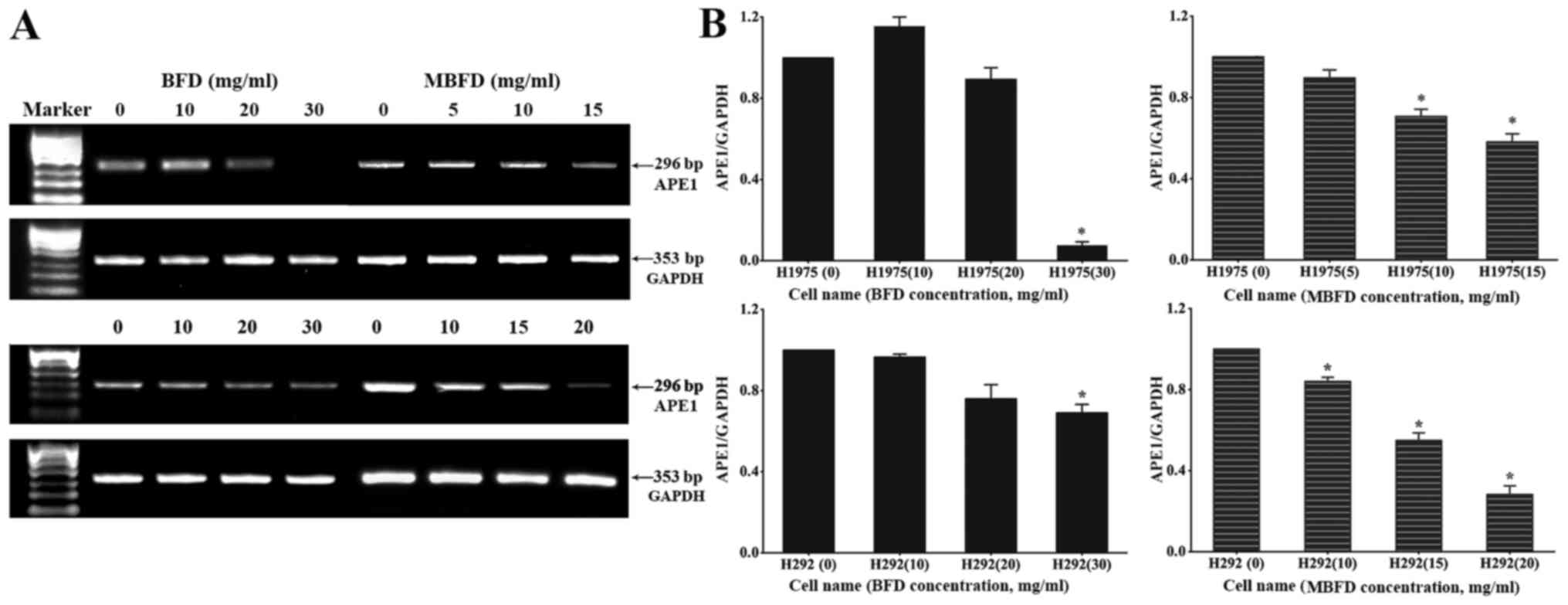
BFD and MBFD inhibit APE1 mRNA expression
in H1975 and H292 cells
The mRNA expression levels of APE1 (296 bp) and
GAPDH (353 bp) were subsequently detected (Fig. 4A). Compared with in the control
H1975 and H292 cells (untreated cells), the mRNA expression levels
of APE1 were decreased following treatment with BFD and MBFD
(P<0.05) (Fig. 4B); however,
10 mg/ml BFD slightly enhanced APE1 mRNA expression in H1975 cells
(P>0.05).
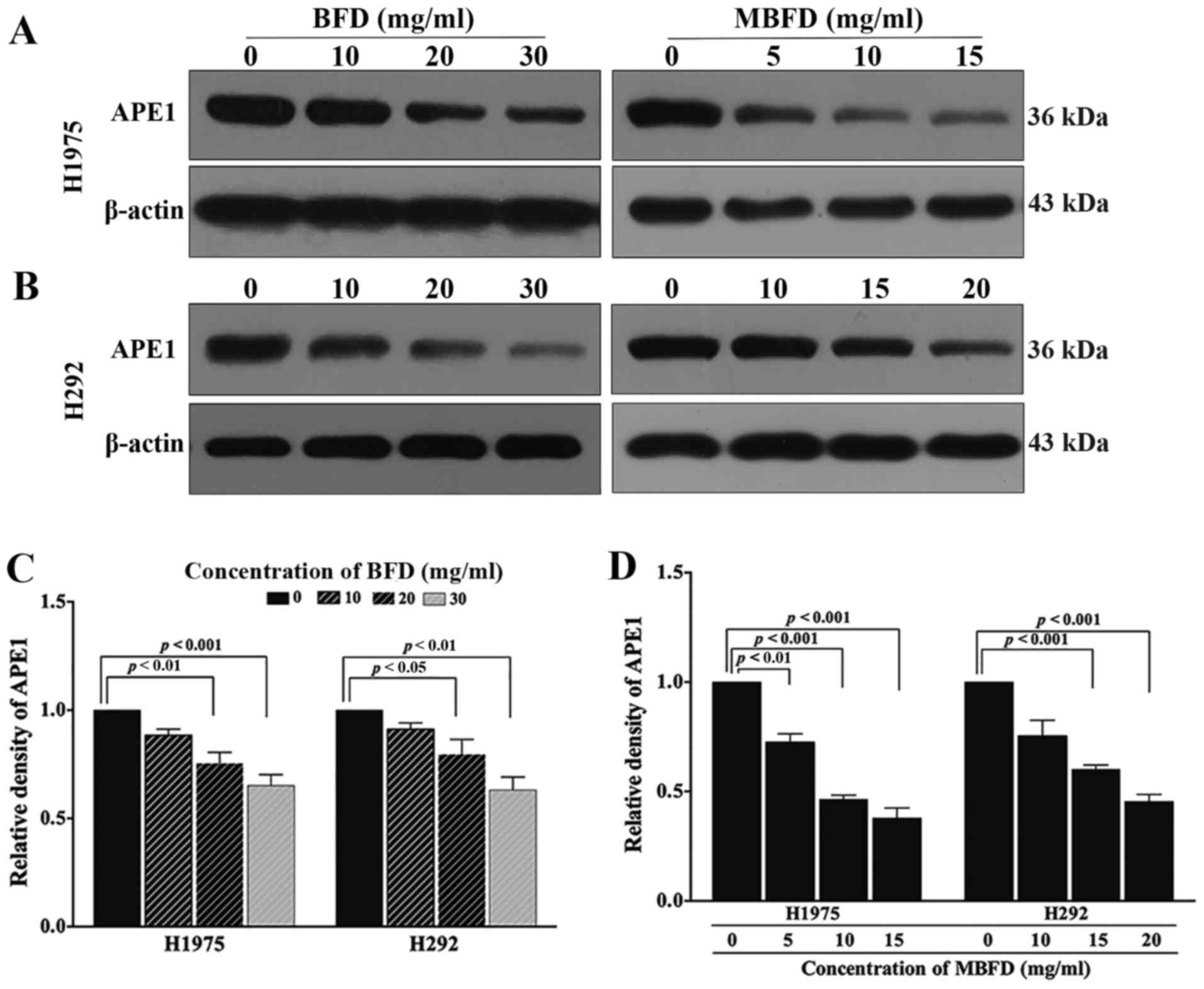
BFD and MBFD inhibit APE1 protein
expression in H1975 and H292 cell lines
As presented in Fig.
5, treatment with BFD or MBFD decreased APE1 protein expression
in H1975 and H292 cells. In H1975 cells, APE1 protein expression
levels were reduced by 11.53, 24.65 (P<0.01) and 34.74%
(P<0.001) in response to treatment with BFD at 10, 20 and 30
mg/ml, respectively; and by 18.42 (P<0.01), 53.51 (P<0.001)
and 62.17% (P<0.001) in response to treatment with MBFD at 5, 10
and 15 mg/ml, respectively. In H292 cells, APE1 protein expression
levels were reduced by 8.74, 20.57 (P<0.05) and 36.84%
(P<0.01) in response to treatment with BFD at 10, 20, and 30
mg/ml, respectively; and by 24.58, 39.79 (P<0.001) and 54.61%
(P<0.001) in response to treatment with MBFD at 10, 15 and 20
mg/ml, respectively. A closer inspection of the data suggested that
MBFD exhibited a stronger effect compared with BFD; the effects of
MBFD on APE1 protein expression were more apparent than BFD.
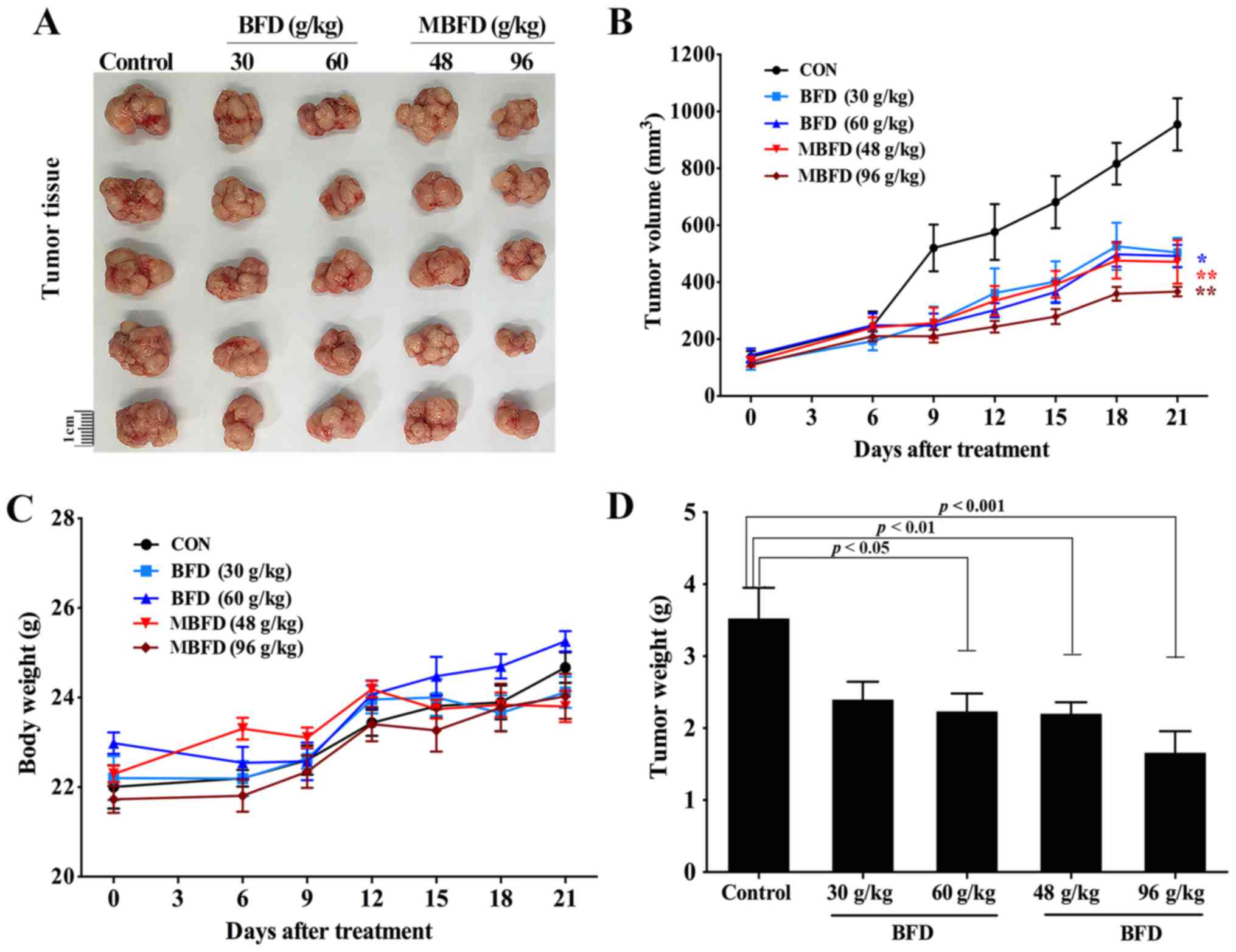
BFD and MBFD suppress tumor growth in
nude mice
BFD and MBFD inhibited tumor growth in a mouse
xenograft model (Fig. 6A). The
tumor growth inhibition rate was 31.97% following treatment with
BFD (30 g/kg), 36.60% following treatment with BFD (60 g/kg),
37.57% following treatment with MBFD (48 g/kg) and 52.92% following
treatment with MBFD (96 g/kg) (Fig.
6B). There were no significant differences in body weight among
the treatment groups at the end of the experiment (Fig. 6C). The alterations in tumor weight
were consistent with those in tumor volume (Fig. 6D).
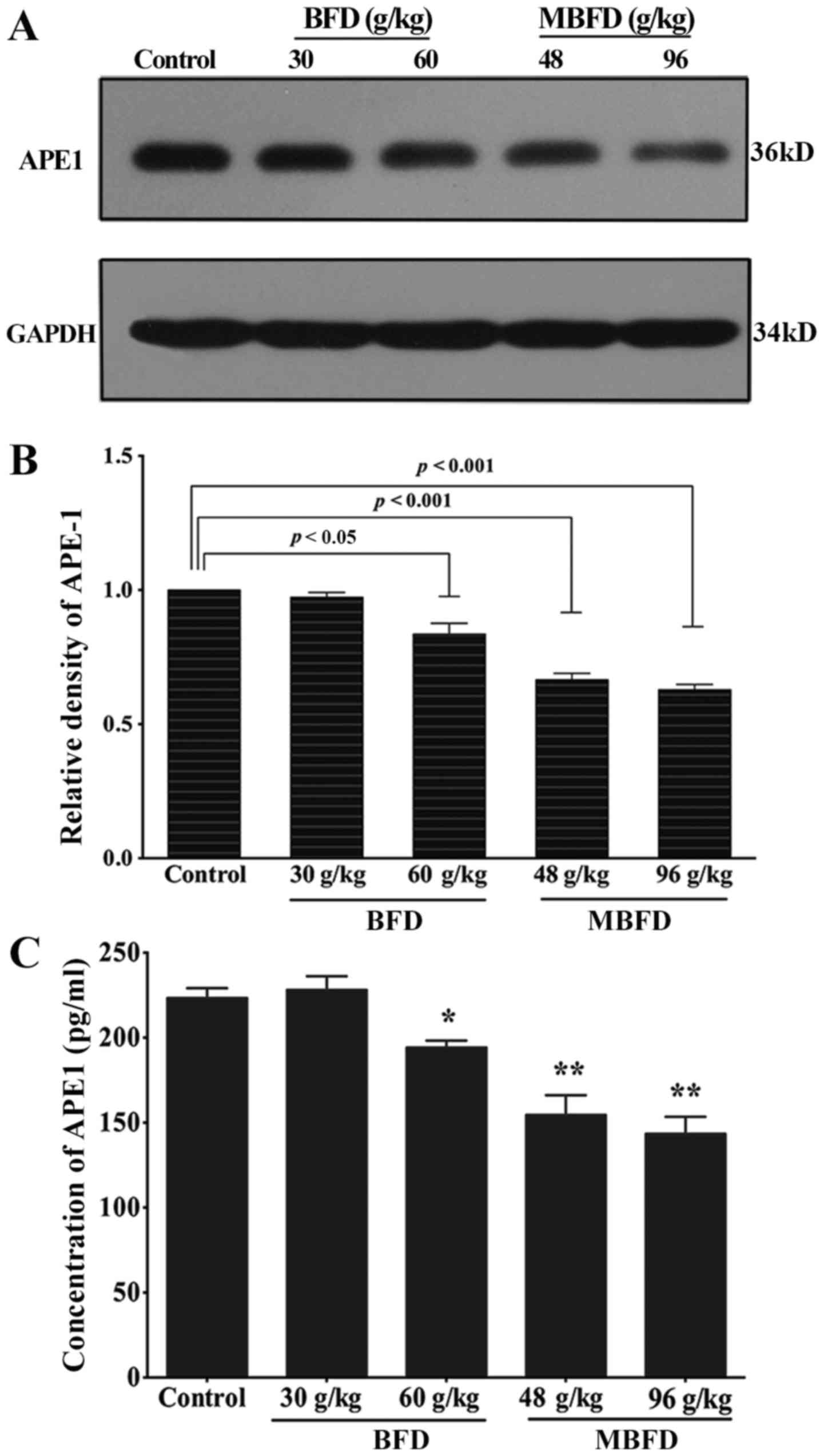
APE1 protein levels are reduced by BFD
and MBFD in mouse xenograft tumors
As shown in Fig. 7A
and B, APE1 protein expression was reduced by 16.43
(P<0.05), 33.40 (P<0.001) and 37.1% (P<0.001) in response
to treatment with BFD at 60 g/kg, MBFD at 48 g/kg and MBFD at 96
g/kg, respectively. At 30 g/kg, BFD did not affect APE1 protein
expression (P>0.05).
BFD and MBFD decrease plasma APE1
concentration
BFD and MBFD significantly decreased plasma APE1
concentrations (Fig. 7C).
Discussion
According to TCM, BFD supplements Qi, clears away
heat and nourishes the lungs, and is believed to treat symptoms
caused by insufficiency of Lung-Qi and endogenous heat-induced lung
injury. Previous studies regarding the application of BFD in lung
cancer have indicated that BFD may be beneficial in reducing lung
cancer-associated symptoms and improving the QOL of patients with
advanced lung cancer (42,43).
Hedyotic diffusa, Duchesnea and Scutellaria barbata
are always added in the application of MBFD. A previous study
regarding Milkvetch and Codonopsis demonstrated that Milkvetch may
induce tumor cell differentiation and death (44). Furthermore, according to TCM,
Hedyotic diffusa, Duchesnea and Scutellaria barbata
exhibit heat-clearing and detoxifying functions, and Scutellaria
barbata possesses antitumor properties. It is also believed
that Scutellaria barbata may serve a role strengthening
vital Qi to eliminate pathogenic factors; it has been suggested
that in advanced cancer a combination of drugs that strengthen
vital Qi and antitumor drugs may be beneficial.
Pharmacological studies have reported that
Milkvetch, which is the main component of BFD and MBFD, is able to
promote the growth of normal cells, improve immunity, accelerate
the regeneration of serum and liver proteins, and protect the liver
and kidney from damage caused by toxic substances (45,46). In addition, another active
ingredient, astragalus polysaccharide, exerts antitumor effects.
Epifriedelanol, which is contained in aster flowers, has also been
reported to exert an anticancer effect on mouse Ehrlich ascites
carcinoma. In addition, Cortex Mori Radicis exerts inhibitory
effects on lung cancer cells (45,46). In TCM, it is believed that
Hedyotic diffusa possesses strong heat-clearing and
detoxifying function (45,46),
and it is therefore widely used in the treatment of various types
of cancer. Experimental studies have indicated that Hedyotic
diffusa is able to suppress the proliferation of lung cancer
cells, arrest cells in G0/G1 phase and
promote cell apoptosis (47,48). Duchesnea and Scutellaria
barbata also exhibit pharmacological effects of detoxification.
Previous studies have indicated that the crude extract or active
ingredient of these two herbs exert inhibitory effects on the
proliferation of lung cancer cell lines and the growth of mouse
xenograft tumors; however, they do not exhibit inhibitory effects
on normal mammary epithelial cells (49–52). To the best of our knowledge, the
present study is the first to provide in vitro and in
vivo evidence to support the antitumor effects of BFD and MBFD
on NSCLC cells, and their inhibitory effects on APE1
expression.
APE1 is the rate-limiting enzyme in the BER pathway
and is the only DNA repair protein with redox regulatory activity,
which indirectly affects DNA repair via the modulation of
transcription factors (7–10). The present results revealed that
the mRNA and protein expression levels of APE1 were decreased in
H1975 and H292 lung cancer cells under BFD and MBFD treatment. In
addition, BFD and MBFD significantly inhibited the proliferation of
H1975 and H292 cells, and the results of comet assays demonstrated
that BFD and MBFD suppressed the DNA repair of these two cell
lines. Conversely, BFD and MBFD exhibited reduced activity against
normal human lung bronchial epithelial BEAS-2B cells, thus
indicating that the application of BFD and MBFD is safe for
patients with lung cancer. Furthermore, the present in vivo
experiments provided strong evidence to support these findings.
Consistent with the in vitro results, the administration of
BFD and MBFD resulted in suppression of tumor growth and a
reduction in the expression of APE1 in mouse plasma samples and
tumor tissues compared with in the control group. In addition, no
significant differences were detected in body weight between the
control and treatment groups. Notably, the appetite and temperament
of the mice was better in the treatment groups compared with in the
control group. In older to further explore the association between
BFD/MBFD and APE1, H292 and H1975 cells were transfected with a
siRNA to downregulate the expression of APE1, cells were then
treated with BFD and MBFD; the results indicated that DNA damage of
H1975 and H292 cells was significantly increased and the
proliferation of the two cell lines was significantly reduced in
response to APE1 siRNA. In addition, H1975 and H292 cells were
transfected with pReceiver-M98 plasmid containing a gene clone of
APE1; subsequently, the results of an MTT assay indicated that
proliferation of the two cell lines was promoted by APE1
overexpression; however, the effects of cell proliferation were
attenuated by BFD or MBFD treatment. It may be hypothesized that
BFD and MBFD inhibit proliferation due to an increase in DNA damage
via inhibition of APE1 expression.
Notably, MBFD exhibited a better antitumor effect
compared with BFD in vivo and in vitro. A possible
explanation may be that since MBFD contained more Hedyotic
diffusa, Duchesnea and Scutellaria barbata than BFD, the
pharmacological activities of these three herbs served a central
role in the improved antitumor effects. Clinically, patients with
cancer that received Chinese medicine were mostly unable to receive
chemotherapy or targeted therapy due to their poor health status.
The present study indicated that MBFD strengthened the antitumor
effects of BFD, which provides evidence to suggest that MBFD may be
applied to treat patients with cancer.
Although the present results revealed the antitumor
effects of BFD and MBFD, there are some limitations to the present
study. There are various DNA repair pathways, and the results of
the comet assays indicated that DNA damage was increased following
treatment with BFD or MBFD; however, the reduction in APE1
expression may be just one of the most relevant factors, and
targeted evidence is required to verify whether DNA damage was
caused by APE1 suppression. Furthermore, as aforementioned, APE1 is
a multifunctional protein with DNA repair and redox activities. The
current experimental results demonstrated that BFD and MBFD
inhibited lung cancer via the suppression of APE1; however, the
specific mechanisms by which BFD and MBFD inhibit lung cancer via
APE1 require further study. Therefore, future experiments should
focus on the effects of BFD/MBFD on the two activities of APE1,
including the expression of the associated DNA transcription
factors, phosphorylated (p)-NF-κB, p-AP-1 and HIF1-α, in
vivo and in vitro. In addition, the present study
observed that, compared with the mice in the control group, the
mice in the treatment groups exhibited better appetite and
temperament; therefore, it may be hypothesized that BFD and MBFD
contribute to improving the QOL of patients with cancer, which is
consistent with the characteristics of tonic Chinese herbs,
including Milkvetch. Research regarding the underlying mechanism
will be conducted in future studies.
In conclusion, the present study demonstrated that
BFD and MBFD exert inhibitory effects on NSCLC growth; these
anti-tumor effects may be associated with the inhibition of APE1.
These results suggested that BFD and MBFD may be considered
promising treatments for NSCLC; in particular, MBFD exhibited a
stronger antitumor effect without impacting the body weight and
activities of mice. Furthermore, the present study provided
evidence to suggest that APE1 may be a target factor for the
treatment of NSCLC with TCM.
Abbreviations:
|
NSCLC
|
non-small cell lung cancer
|
|
BFD
|
Bu-Fei decoction
|
|
APE1
|
apurinic/apyrimidinic endonuclease
1
|
|
MBFD
|
modified Bu-Fei decoction
|
Acknowledgments
Not applicable.
Notes
[1]
Funding
The present study was partially supported by Beijing
Municipal Health System Special Funds of High-level Medical
Personnel Construction (grant no. 2014-3-063) and the Key Program
Foundation of Beijing Administration of Traditional Chinese
Medicines (grant no. 2004-IV15).
[2] Availability
of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
[3] Authors’
contributions
STJ designed and implemented the experiment, and was
a major contributor in writing the manuscript. SYH auxiliary
designed the experiment. YNJ, LNP and XRH auxiliary implemented the
experiment. PPL overall planed and designed the experiment design.
All authors read and approved the final manuscript.
[4] Ethics
approval and consent to participate
The present study was approved by the Institutional
Animal Care and Use Committee of the First Hospital Affiliated to
PLA General Hospital.
[5] Consent for
publication
Not applicable.
[6] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Kastan MB: DNA damage responses:
mechanisms and roles in human disease: 2007 G.H.A. Clowes Memorial
Award Lecture. Mol Cancer Res. 6:517–524. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rodriguez-Rocha H, Garcia-Garcia A,
Panayiotidis MI and Franco R: DNA damage and autophagy. Mutat Res.
711:158–166. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Altieri F, Grillo C, Maceroni M and
Chichiarelli S: DNA damage and repair: from molecular mechanisms to
health implications. Antioxid Redox Signal. 10:891–937. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Christmann M, Tomicic MT, Roos WP and
Kaina B: Mechanisms of human DNA repair: An update. Toxicology.
193:3–34. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Roos WP, Thomas AD and Kaina B: DNA damage
and the balance between survival and death in cancer biology. Nat
Rev Cancer. 16:20–33. 2016. View Article : Google Scholar
|
|
6
|
Wilson DM III and Bohr VA: The mechanics
of base excision repair, and its relationship to aging and disease.
DNA Repair (Amst). 6:544–559. 2007. View Article : Google Scholar
|
|
7
|
Bhakat KK, Mantha AK and Mitra S:
Transcriptional regulatory functions of mammalian AP-endonuclease
(APE1/Ref-1), an essential multifunctional protein. Antioxid Redox
Signal. 11:621–638. 2009. View Article : Google Scholar
|
|
8
|
Li M and Wilson DM III: Human
apurinic/apyrimidinic endonuclease 1. Antioxid Redox Signal.
20:678–707. 2014. View Article : Google Scholar :
|
|
9
|
Wood RD, Mitchell M, Sgouros J and Lindahl
T: Human DNA repair genes. Science. 291:1284–1289. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Luo M, He H, Kelley MR and Georgiadis MM:
Redox regulation of DNA repair: implications for human health and
cancer therapeutic development. Antioxid Redox Signal.
12:1247–1269. 2010. View Article : Google Scholar :
|
|
11
|
Robertson KA, Bullock HA, Xu Y, Tritt R,
Zimmerman E, Ulbright TM, Foster RS, Einhorn LH and Kelley MR:
Altered expression of Ape1/ref-1 in germ cell tumors and
overexpression in NT2 cells confers resistance to bleomycin and
radiation. Cancer Res. 61:2220–2225. 2001.PubMed/NCBI
|
|
12
|
Wang D, Xiang DB, Yang XQ, Chen LS, Li MX,
Zhong ZY and Zhang YS: APE1 overexpression is associated with
cisplatin resistance in non-small cell lung cancer and targeted
inhibition of APE1 enhances the activity of cisplatin in A549
cells. Lung Cancer. 66:298–304. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Di Maso V, Avellini C, Crocè LS, Rosso N,
Quadrifoglio F, Cesaratto L, Codarin E, Bedogni G, Beltrami CA,
Tell G, et al: Subcellular localization of APE1/Ref-1 in human
hepatocellular carcinoma: Possible prognostic significance. Mol
Med. 13:89–96. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kakolyris S, Kaklamanis L, Engels K, Fox
SB, Taylor M, Hickson ID, Gatter KC and Harris AL: Human AP
endonuclease 1 (HAP1) protein expression in breast cancer
correlates with lymph node status and angiogenesis. Br J Cancer.
77:1169–1173. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mahjabeen I, Ali K and Zhouxand Kayani MA:
Deregulation of base excision repair gene expression and enhanced
proliferation in head and neck squamous cell carcinoma. Tumour
Biol. 35:5971–5983. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Poletto M, Di Loreto C, Marasco D, Poletto
E, Puglisi F, Damante G and Tell G: Acetylation on critical lysine
residues of Apurinic/apyrimidinic endonuclease 1 (APE1) in triple
negative breast cancers. Biochem Biophys Res Commun. 424:34–39.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Qing Y, Li Q, Ren T, Xia W, Peng Y, Liu
GL, Luo H, Yang YX, Dai XY, Zhou SF, et al: Upregulation of PD-L1
and APE1 is associated with tumorigenesis and poor prognosis of
gastric cancer. Drug Des Devel Ther. 9:901–909. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Qing Y, Wang D, Lei X, Xiang DB, Li MX, Li
ZP and Shan JL: The expression of APE1 and its correlation with
prognostic significance after 252Cf radiotherapy in cervical
cancer. Sichuan Da Xue Xue Bao Yi Xue Ban. 40:125–128. 2009.In
Chinese. PubMed/NCBI
|
|
19
|
Sheng Q, Zhang Y, Wang R, Zhang J, Chen B,
Wang J, Zhang W and Xin X: Prognostic significance of APE1
cytoplasmic localization in human epithelial ovarian cancer. Med
Oncol. 29:1265–1271. 2012. View Article : Google Scholar
|
|
20
|
Tell G, Damante G, Caldwell D and Kelley
MR: The intracellular localization of APE1/Ref-1: more than a
passive phenomenon? Antioxid Redox Signal. 7:367–384. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chantre-Justino M, Alves G, Britto C,
Cardoso A, Scherrer L, Moreira Ados S, Quirino R, Ornellas A,
Leitão A and Lage C: Impact of reduced levels of APE1 transcripts
on the survival of patients with urothelial carcinoma of the
bladder. Oncol Rep. 34:1667–1674. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang Y, Zhou S, Sandusky GE, Kelley MR
and Fishel ML: Reduced expression of DNA repair and redox signaling
protein APE1/Ref-1 impairs human pancreatic cancer cell survival,
proliferation, and cell cycle progression. Cancer Invest.
28:885–895. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dai N, Cao XJ, Li MX, Qing Y, Liao L, Lu
XF, Zhang SH, Li Z, Yang YX and Wang D: Serum APE1 autoantibodies:
A novel potential tumor marker and predictor of chemotherapeutic
efficacy in non-small cell lung cancer. PLoS One. 8:e580012013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mok TS, Wu YL, Thongprasert S, Yang CH,
Chu DT, Saijo N, Sunpaweravong P, Han B, Margono B, Ichinose Y, et
al: Gefitinib or carboplatin-paclitaxel in pulmonary
adenocarcinoma. N Engl J Med. 361:947–957. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Socinski MA, Bondarenko I, Karaseva NA,
Makhson AM, Vynnychenko I, Okamoto I, Hon JK, Hirsh V, Bhar P,
Zhang H, et al: Weekly nab-paclitaxel in combination with
carboplatin versus solvent-based paclitaxel plus carboplatin as
first-line therapy in patients with advanced non-small-cell lung
cancer: Final results of a phase III trial. J Clin Oncol.
30:2055–2062. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Solomon BJ, Mok T, Kim DW, Wu YL, Nakagawa
K, Mekhail T, Felip E, Cappuzzo F, Paolini J, Usari T, et al
PROFILE 1014 Investigators: First-line crizotinib versus
chemotherapy in ALK-positive lung cancer. N Engl J Med.
371:2167–2177. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
National Comprehensive Cancer Network
(NCCN): NCCN Clinical Practice Guidelines in Oncology (NCCN
Guidelines®). Non-Small Cell Lung Cancer. https://www.nccn.org/professionals/physician_gls/PDF/nscl.pdf.
|
|
31
|
Piao BK, Tang WX, Zhang ZQ, Lin HS, Duan
FW and Yu GQ: The observation of Feiliuping ointment treatment of
advanced lung cancer-with 339 cases of clinical analysis. J Tradit
Chin Med. 21–23. 1991.
|
|
32
|
Li PW, Zhang DZ and Hao YX: Clinical
analysis of Ping-Fei decoction treatment of 109 cases of non-small
cell lung cancer. J Tradit Chin Med. 87–88. 1995.
|
|
33
|
Yang GW, Wang XM, Han D, et al: Study on
TCM comprehensive therapy in treatment of advanced Non-Small Cell
Lung Cancer. Zhong Guo Zhong Yi Yao Xin Xi Za Zhi. 12:11–13.
2005.In Chinese.
|
|
34
|
Han Y, Wang H, Xu W, Cao B, Han L, Jia L,
Xu Y, Zhang Q, Wang X, Zhang G, et al: Chinese herbal medicine as
maintenance therapy for improving the quality of life for advanced
non-small cell lung cancer patients. Complement Ther Med. 24:81–89.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jiang Y, Liu LS, Shen LP, Han ZF, Jian H,
Liu JX, Xu L, Li HG, Tian JH and Mao ZJ: Traditional Chinese
Medicine treatment as maintenance therapy in advanced
non-small-cell lung cancer: A randomized controlled trial.
Complement Ther Med. 24:55–62. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li W, Chen C, Saud SM, Geng L, Zhang G,
Liu R and Hua B: Fei-Liu-Ping ointment inhibits lung cancer growth
and invasion by suppressing tumor inflammatory microenvironment.
BMC Complement Altern Med. 14:1532014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
He XR, Han SY, Li XH, Zheng WX, Pang LN,
Jiang ST and Li PP: Chinese medicine Bu-Fei decoction attenuates
epithelial-mesenchymal transition of non-small cell lung cancer via
inhibition of transforming growth factor β1 signaling pathway in
vitro and in vivo. J Ethnopharmacol. 204:45–57. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pang L, Han S, Jiao Y, Jiang S, He X and
Li P: Bu Fei Decoction attenuates the tumor associated macrophage
stimulated proliferation, migration, invasion and immunosuppression
of non-small cell lung cancer, partially via IL-10 and PD-L1
regulation. Int J Oncol. 51:25–38. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
40
|
Guide for the care and use of laboratory
animals. 8th. The National Academies Press; Washington, USA:
|
|
41
|
Zhou N, Han SY, Zhou F and Li PP:
Antitumor effect of Shu-Gan-Liang-Xue decoction in breast cancer is
related to the inhibition of aromatase and steroid sulfatase
expression. J Ethnopharmacol. 154:687–695. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang QJ and Xu XP: The application of
Bu-Fei decoction in lung deficiency symptom. Hubei Journal of
Traditional Chinese Medicine. 34:1999.
|
|
43
|
Ke MY, Li XF and Li RH: Bu-Fei decoction
treatment of 32 cases of advanced lung cancer. Fujian Journal of
Traditional Chinese Medicine. 21:1995.
|
|
44
|
Cheng XD, Hou CH, Zhang XJ, Xie HY, Zhou
WY, Yang L, Zhang SB and Qian RL: Effects of Huangqi (Hex) on
inducing cell differentiation and cell death in K562 and HEL cells.
Acta Biochim Biophys Sin (Shanghai). 36:211–217. 2004. View Article : Google Scholar
|
|
45
|
Shen YJ: Pharmacology of Traditional
Chinese Medicine. 1st. People’s Medical Publishing House; Beijing,
China: 2000
|
|
46
|
Gao XM: Pharmacy. China Press of
Traditional Chinese Medicine; Beijing, China: 2007
|
|
47
|
Kuo YJ, Yang JS, Lu CC, Chiang SY, Lin JG
and Chung JG: Ethanol extract of Hedyotis diffusa willd upregulates
G0/G1 phase arrest and induces apoptosis in human leukemia cells by
modulating caspase cascade signaling and altering associated genes
expression was assayed by cDNA microarray. Environ Toxicol.
30:1162–1177. 2015. View Article : Google Scholar
|
|
48
|
Lee HZ, Bau DT, Kuo CL, Tsai RY, Chen YC
and Chang YH: Clarification of the phenotypic characteristics and
Antitumor activity of Hedyotis diffusa. Am J Chin Med. 39:201–213.
2011. View Article : Google Scholar
|
|
49
|
Shoemaker M, Hamilton B, Dairkee SH, Cohen
I and Campbell MJ: In vitro anticancer activity of twelve Chinese
medicinal herbs. Phytother Res. 19:649–651. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Gong T, Wang CF, Yuan JR, Li Y, Gu JF,
Zhao BJ, Zhang L, Jia XB, Feng L and Liu SL: Inhibition of Tumor
Growth and immunomodulatory effects of flavonoids and
scutebarbatines of Scutellaria barbata D Don in Lewis-bearing
C57BL/6 mice. Evid Based Complement Alternat Med. 2015:6307602015.
View Article : Google Scholar
|
|
51
|
Shiau AL, Shen YT, Hsieh JL, Wu CL and Lee
CH: Scutellaria barbata inhibits angiogenesis through
downregulation of HIF-1 α in lung tumor. Environ Toxicol.
29:363–370. 2014. View Article : Google Scholar
|
|
52
|
Yin X, Zhou J, Jie C, Xing D and Zhang Y:
Anticancer activity and mechanism of Scutellaria barbata extract on
human lung cancer cell line A549. Life Sci. 75:2233–2244. 2004.
View Article : Google Scholar : PubMed/NCBI
|