Introduction
Collagen degradation is the primary cause of skin
aging. The modulation of collagen synthesis/degradation is thus a
pivotal target for anti-wrinkle agents. Glycosaminoglycans (GAGs)
are highly charged polysaccharides that are an important structural
component of the extracellular matrix (ECM) and are often used in
cosmetic products (1-3). In a previous clinical trial, it was
identified that a cream containing GAGs regulated wrinkles, skin
elasticity, dermal density and skin tightening (4). However, although GAGs serve
important roles in skin aging, their regulatory mechanism of action
has not yet been fully elucidated.
Skin wrinkles develop as a result of intrinsic and
extrinsic aging processes (5,6).
Intrinsic aging, which is characterized by a smooth, thinned
epidermis that exhibits fine wrinkles, naturally occurs over time
and is dependent on the accumulation of inflammatory mediators.
Extrinsic aging, characterized by a roughened texture, skin laxity
and mottled pigmentation with deep wrinkles, occurs due to solar
irradiation, smoking and poor nutrition (7,8).
These alterations may also be caused by ECM destruction as a dermal
fibroblast aging that occur during intrinsic and extrinsic aging
(9). Dermal fibroblasts serve an
important role in the production of ECM, including the production
of ground substance, collagen fibers and elastins (10). Therefore, the disruption of
fibroblast function affects the mechanical properties of skin
connective tissue. Inflammatory mediators, including cytokines,
serve a crucial role in stimulating skin aging (11). The expression of a gene cluster
associated with inflammation (12) and the quantity of halogenated
tyrosine produced following inflammation are increased in aging
skin (13). Specifically,
exposure to ultraviolet B promotes the production of tumor necrosis
factor (TNF)-α from dermal fibroblasts, macrophages and epidermal
keratinocytes, resulting in increased inflammation and the
degradation of ECM components via the activation of matrix
metalloproteinases (MMPs) (14-18). Sustained exposure to TNF-α induces
the expression of MMP-1, MMP-3 and MMP-9 (also known as
collagenase, stromelysin and gelatinase, respectively), causing
irreparable damage to the epidermis and dermis (19-21). In particular, MMP-1 activation in
dermal fibroblasts may cause collagen fragmentation and functional
alterations in dermal fibroblasts (22). Thus, regulating TNF-α activity may
be a novel therapeutic strategy of treating inflammatory skin
diseases and reversing skin aging.
Type I collagen, the primary component of ECM in the
skin, is synthesized and secreted by dermal fibroblasts. Type I
collagen is responsible for the strength and elasticity of skin
(23). In addition to aging, the
stimulation of various cytoplasmic signal transduction pathways,
including transforming growth factor (TGF)-β/Smad,
mitogen-activated protein kinase (MAPK) and nuclear factor (NF)-κB,
cause fibroblasts to lose their proliferative potential.
Fibroblasts then synthesize reduced levels of ECM, meaning that the
secretion of type I collagen is also reduced (24-28). Impairments in dermal fibroblast
functions during aging contribute to skin thinning, as they cause
reductions in the quantity of collagen in aged human skin (29). Previous studies have demonstrated
that various MAPKs, including extracellular signal-regulated kinase
(ERK), c-jun N-terminal kinase (JNK) and p38, are associated with
skin aging (26,30-32). The phosphorylation of ERK 1/2
mediates the inhibition of type I collagen synthesis in human skin
fibroblasts (32,33). TNF-α increases MMP-1 expression
via MAPK and activator protein-1 (AP-1) pathways in rheumatoid
arthritis synovial fibroblasts (34) and HCS-2/8 chondrocytes (35). Furthermore, it has been
demonstrated that compound K suppresses ERK activation, resulting
in reduced MMP-1 secretion and increased type I procollagen
secretion in TNF-α-stimulated human skin fibroblasts, suggesting
that compound K acts as an anti-aging agent (36). However, to the best of our
knowledge, there have been no previous studies assessing the
ability of GAGs on attenuating TNF-α-induced type I collagen
denaturation or the molecular mechanisms underlying the
anti-inflammatory effects in human dermal fibroblasts.
The present study assessed the effect of GAGs on the
collagen synthesis process in TNF-α-stimulated human dermal
fibroblasts (HDFs). The results demonstrated that the molecular
mechanism underlying the inhibitory effect of GAGs is associated
with the inhibition of ERK/AP-1 signaling. These data indicate that
GAGs may serve a critical role in the attenuation of skin
inflammation and aging.
Materials and methods
Materials
Recombinant human TNF-α (cat. no. RC214-12) was
purchased from Bio Basic, Inc. (Markham, ON, Canada). PD98059 (cat.
no. P215) was purchased from Sigma-Aldrich; Merck KGaA (Darmstadt,
Germany). Antibodies against phosphorylated (p)-ERK (cat. no.
#9101), ERK (cat. no. #9102), p-p38 (cat. no. #4511), p38 (cat. no.
#9212), JNK (cat. no. #9252), p-c-fos (cat. no. #5348), c-fos (cat.
no. #2250), p-c-jun (cat. no. #9164), c-jun (cat. no. #9165), total
caspase-3 (cat. no. #9662), cleaved caspase-3 (cat. no. #9662) and
TIMP-1 (cat. no. #8946) were all purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA). The antibody against MMP-1
(cat. no. ab137332) was purchased from Abcam (Cambridge, UK) and
the antibodies against Bax (cat. no. sc-7480), Bcl-2 (cat. no.
sc-492) and β-actin (cat. no. sc-47778) were purchased from Santa
Cruz Biotechnology, Inc. (Dallas, TX, USA). The antibody against
collagen type I (cat. no. NB600-408) was purchased from Novus
Biologicals, LLC (Littleton, CO, USA) and the antibody against
p-JNK (cat. no. 612541) was purchased from BD Transduction
Laboratories; BD Biosciences (Franklin Lakes, NJ, USA). GAG
complexes were provided by Taeyoung cosmetics (Elensilia, Seongnam,
Korea).
Cell cultures and cell viability
assay
HDFs were obtained by skin biopsy from one healthy
male donor aged 12 years old on September 2014 (Chung-Ang
University Hospital, Seoul, Seoul, Korea). The present study was
approved by the Ethical Committee of Chung-Ang University Hospital.
Written informed consent was obtained from the legal guardians of
the donor prior to enrolment. Primary explant cultures were
established in 60-cm2 culture flasks in Dulbecco's
modified Eagle's medium (DMEM; Welgene, Inc., Gyeongsan, Korea)
supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), 100 U/ml penicillin and 10
μg/ml streptomycin. HDFs were maintained at 37°C in a
humidified atmosphere of 95% air and 5% CO2. HDFs at
passages 3-8 were used for experiments. To investigate the effect
of GAGs, HDFs were pretreated with GAGs (0, 0.1, 0.5, and 1%) for
30 min and were then stimulated with TNF-α (20 ng/ml) for 24 h.
Cell viability was determined using an MTT assay following the
method described by Twentyman and Luscombe (37), with minor modifications. HDFs were
seeded at a density of 5×104 cells/well in 24-well
plates. Prior to treatment, cells were cultured for 24 h in
serum-free DMEM, which was followed by treatment with GAGs (0, 0.1,
0.5, and 1%) for 24 h. HDFs were then incubated with 5 mg/ml MTT
for 4 h prior to addition of dimethyl sulfoxide (Sigma-Aldrich;
Merck KGaA) was added to dissolve the formazan crystals. Following
MTT assays, absorbance was measured at 570 nm using a SpectraMax i3
microplate reader (Molecular Devices, Sunnyvale, CA, USA).
Enzyme-linked immunosorbent assay
(ELISA)
ELISA was performed using a procollagen type I
C-peptide ELISA assay kit (cat. no. MK101; Takara Bio, Inc., Otsu,
Japan) and an MMP-1 ELISA kit (cat. no. DY900-05; R&D Systems,
Inc., Minneapolis, MN, USA) following the manufacturer's protocol.
HDFs were pretreated with GAGs (0, 0.1, 0.5, and 1%) for 30 min and
were then stimulated with TNF-α (20 ng/ml) for 24 h. Other HDFs
were pretreated with 20 μM PD98059 (an ERK inhibitor) for 1
h and were then stimulated with TNF-α (20 ng/ml) for 24 h.
Collected supernatants (obtained from conditioned media) were used
for ELISA and relative absorbance was measured at 450 nm using the
SpectraMax i3 microplate reader.
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from HDFs using TRIzol
(Invitrogen; Thermo Fisher Scientific, Inc.). First-strand cDNA
synthesis from the total RNA template was performed using the
PrimeScript™ RT master mix (Takara Bio, Inc.). The reverse
transcription product was subsequently diluted with 200 μl
H2O. The resulting cDNA was subjected to quantitative
PCR using TOPreal™ qPCR 2X PreMIX SYBR (Enzynomics, Daejeon, Korea)
with a CFX-96 Touch™ Real-Time PCR Detection system (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The thermocycling
conditions used to amplify all genes were as follows: 10 min at
95°C, 40 cycles of 95°C for 15 sec, 60°C for 30 sec and 72°C for 30
sec. Expression data were calculated from the cycle threshold (Cq)
value using the ΔΔCq method of quantification (38). Gene expression values were
normalized to the expression of GAPDH. The sequences of the
oligonucleotide primers used for qPCR are presented in Table I.
 | Table IOligonucleotide primers used for
quantitative polymerase chain reaction. |
Table I
Oligonucleotide primers used for
quantitative polymerase chain reaction.
| Gene | Forward
(5′-3′) | Reverse
(5′-3′) |
|---|
| GAPDH C |
ACCCACTCCTCCACCTTTGAC |
GTCCACCACCCTGTTGCTGTAG |
| COL1A1 |
ATCAACCGGAGGAATTTCCGT |
CACCAGGACGACCAGGTTTTC |
| MMP-1 |
GATGTGGCTCAGTTTGTCCTCAC C |
TTGGCAAATCTGGCGTGTAAT |
Cell lysate preparation and western blot
analysis
Cell extracts were prepared as described previously
(39). Treated whole cell
extracts were lysed in radioimmunoprecipitation buffer [50 mM Tris
(pH 7.4), 150 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate]
containing a protease inhibitor cocktail (Roche Diagnostics,
Indianapolis, IN, USA). Protein concentrations were determined
using a BCA assay kit (Thermo Fisher Scientific Inc.). For western
blot analysis, cell lysates, each containing 20-30 μg
protein, were resolved using 8-12% SDS-PAGE and were then
transferred to polyvinylidene fluoride membranes. Membranes were
soaked in 5% skim milk blocking buffer for 1 h at room temperature.
Subsequently, membranes were incubated with anti-collagen I
(1:1,000), anti-MMP-1 (1:2,500), anti-TIMP-1 (1:2,500), anti-p-ERK
(1:2,500), anti-ERK (1:2,500), anti-p-p38 (1:2,500), anti-p38
(1:2,500), anti-p-JNK (1:2,500), anti-JNK (1:2,500), anti-p-c-Fos
(1:2,500), anti-c-Fos (1:2,500), anti-p-c-Jun (1:2,500), anti-c-Jun
(1:2,500), anti-Bax (1:1,000), anti-Bcl-2 (1:1,000), anti-total
caspase-3 (1:2,500), anti-cleaved caspase-3 (1:2,500) and
anti-β-actin (1:1,000; loading control) antibodies overnight at
4°C. Then, the membrane was incubated with anti-mouse (cat. no.
PI-2000, 1:10,000), anti-rabbit (cat. no. PI-1000, 1:10,000; Vector
Labs Inc., Burlingame, CA, USA) secondary antibodies conjugated to
horseradish peroxidase for 1 h at room temperature. Membranes were
developed using enhanced chemiluminescence western blot detection
reagents (GE Healthcare, Chicago, IL, USA). Immunoblots were
analyzed using ImageJ 1.44 software (National Institutes of Health,
Bethesda, MD, USA).
Statistical analyses
All quantitative data are presented as the mean ±
standard error of the mean for three independent experiments.
Statistical analyses were performed using the statistical package
for SPSS software version 10.0 (SPSS, Inc., Chicago, IL, USA).
Differences between the two groups were evaluated using a paired
t-test. For multiple comparisons, one-way analysis of variance was
used followed by Tukey's multiple comparisons test. P<0.05 was
considered to indicate a statistically significant result.
Results
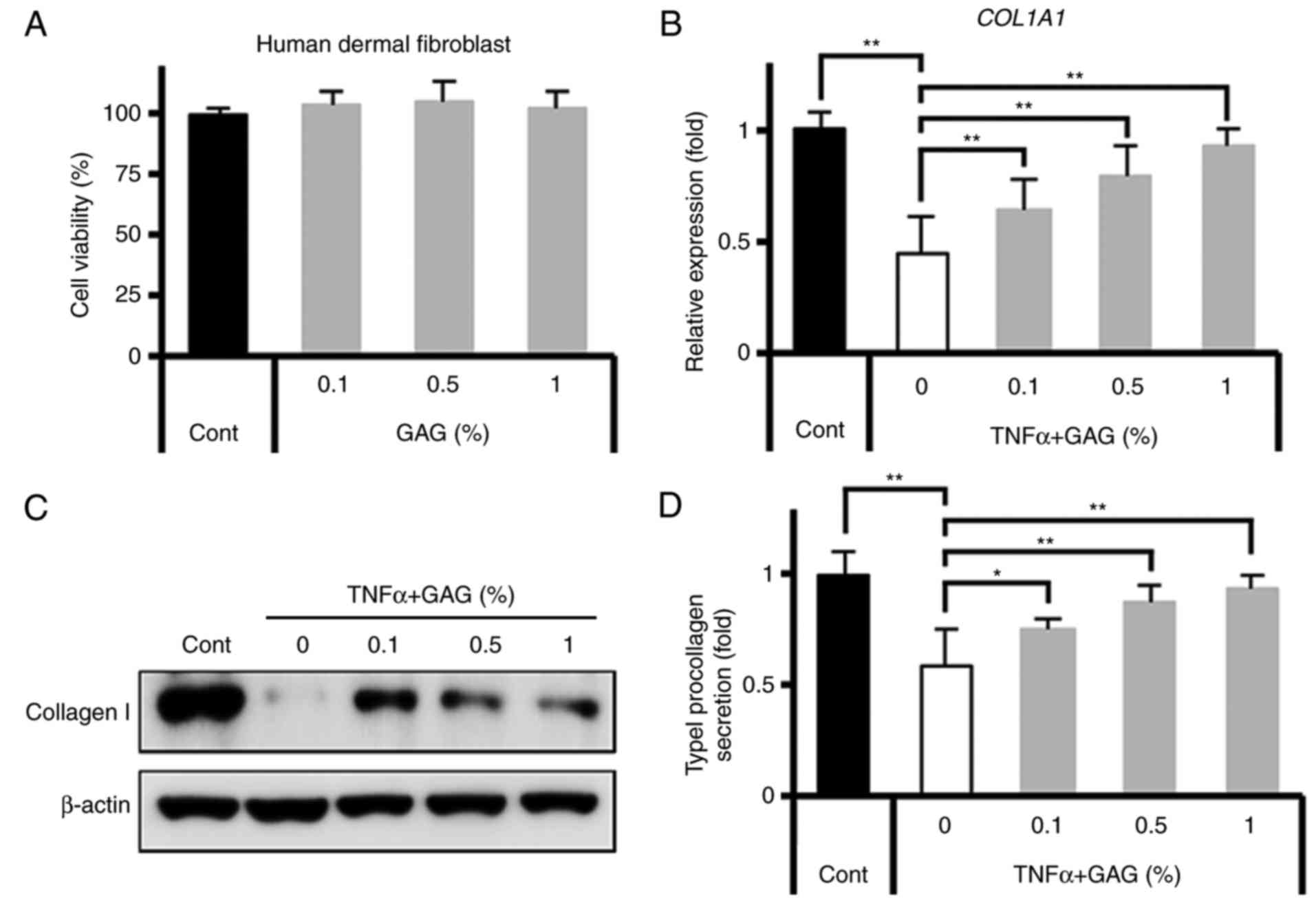
GAGs regulate t ype I collagen production
in TNF-α-stimulated human dermal fibroblasts
Firstly, to measure the cytotoxic effect of GAGs,
HDFs were serially treated for 24 h with various concentrations of
GAGs (0, 0.1, 0.5 and 1%). The results demonstrated that GAGs had
no significant effect on cell viability at any of the tested
concentrations (Fig. 1A). To
identify the protective effect of GAGs against factors that
influence skin aging, the effect of GAGs on TNF-α-induced type I
collagen production in HDFs were assessed. HDFs were pretreated
with GAGs for 30 min and were then stimulated with TNF-α (20 ng/ml)
for 24 h. The expression of type I collagen decreased significantly
following stimulation with TNF-α (Fig. 1B). However, treatment with GAGs
significantly reversed this decrease in type I collagen expression
in a dose-dependent manner (Fig. 1B
and C). Furthermore, type I collagen secretion by HDFs
following stimulation was significantly decreased; this decrease
was reversed following treatment with GAGs (Fig. 1D). These results suggest that GAGs
regulate the production of type I collagen in TNF-α-stimulated
HDFs.
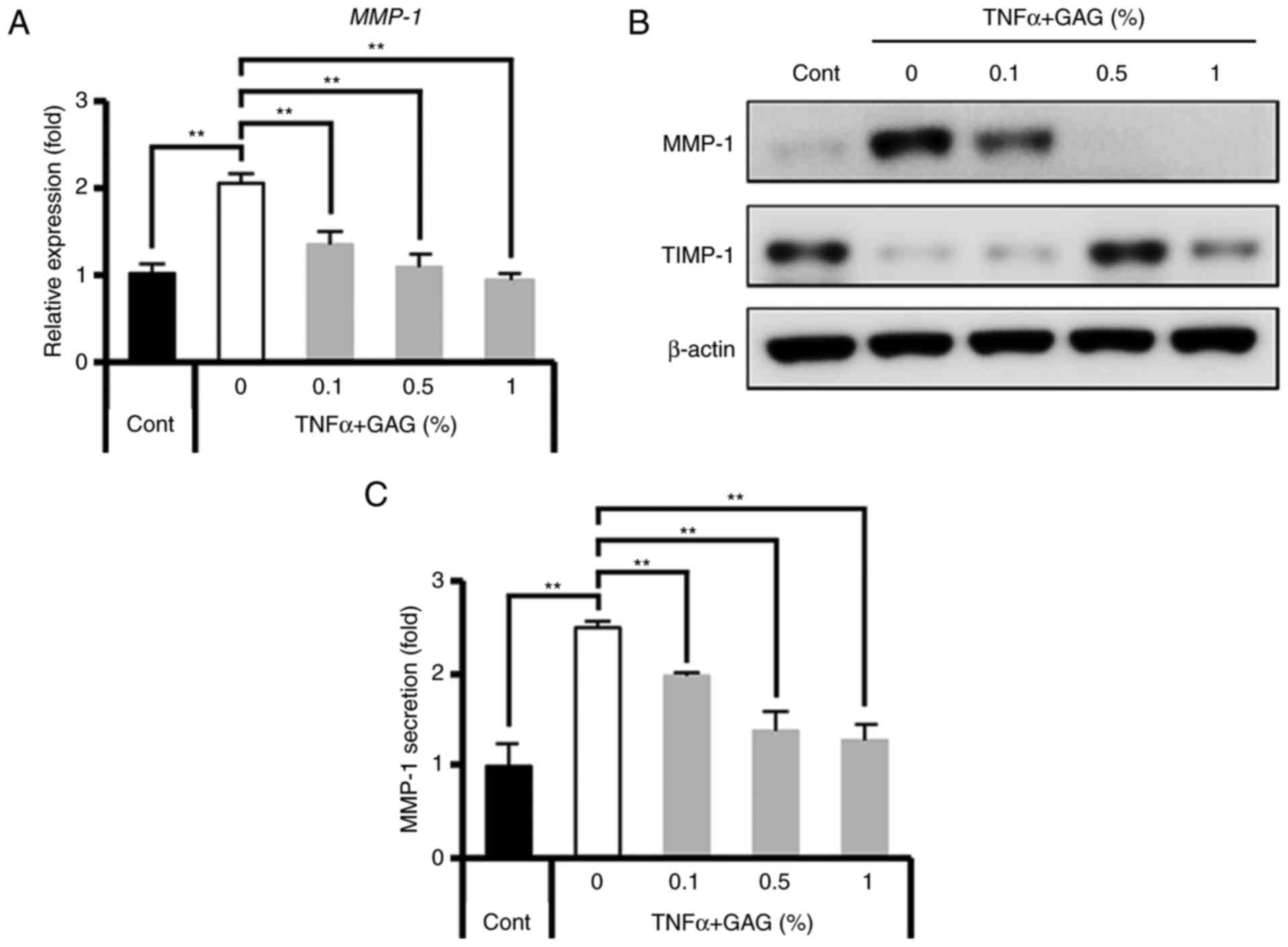
GAGs enhance type I collagen production
via MMP-1 inhibition
The results demonstrated that GAGs reversed the
reduction in type I collagen levels that occurred in
TNF-α-stimulated HDFs (Fig. 1).
The regulatory proteins of collagen degradation were further
assessed as many enzymes, including MMPs and tissue inhibitors of
metalloproteinase (TIMPs), are directly and indirectly involved in
collagen type I degradation (40-43). The results demonstrated that GAGs
inhibited MMP-1 gene activation (Fig.
2A and B) and also MMP-1 secretion (Fig. 2C) in TNF-α-stimulated HDFs in a
dose-dependent manner. However, the expression of TIMP-1, a tissue
inhibitor of metalloproteinases, decreased in TNF-α-stimulated
HDFs, but increased following GAG treatment, peaking following
treatment with 0.5% GAGs (Fig.
2B). Subsequently, the molecular mechanism by which MMP-1
production is inhibited in HDFs by GAGs following stimulation with
TNF-α was assessed.
GAGs inhibit MMP-1 expression via
suppression of the ERK signaling pathway
Previous studies have indicated that MAPK pathways,
specifically those induced by TNF-α, serve an important role in the
regulation of procollagen synthesis (26,30-32,35,44). Thus, the present study assessed
TNF-α induced MAPK signaling. The results demonstrated that TNF-α
activated the phosphorylation of ERK, JNK and p38, peaking at 30
min and then the signal gradually weakened (Fig. 3A). To determine whether GAGs
suppressed the activation of MAPK, HDFs were pretreated for 30 min
with GAGs (0, 0.1, 0.5 and 1%) and then stimulated with TNF-α for a
further 30 min. The results demonstrated that GAGs attenuated ERK
phosphorylation but did not affect the phosphorylation of JNK and
p38 (Fig. 3B). In addition, GAGs
markedly attenuated the phosphorylation of AP-1, including c-fos
and c-jun, the major transcription factors downstream of ERK that
are responsible for the expression of MMP-1 (Fig. 3C). The results confirmed that
MMP-1 activation and type I collagen regulation is dependent on ERK
signaling using the MEK/ERK inhibitor PD98059 in HDFs. Treatment
with PD98059 (20 μM) significantly attenuated the
upregulation of MMP-1 by TNF-α (Fig.
3D). Furthermore, treatment with PD98059 resulted in recovered
type I collagen secretion (Fig.
3D). These results confirmed that GAGs inhibit MMP-1 expression
via ERK inactivation, followed by an increase in type I collagen
production.
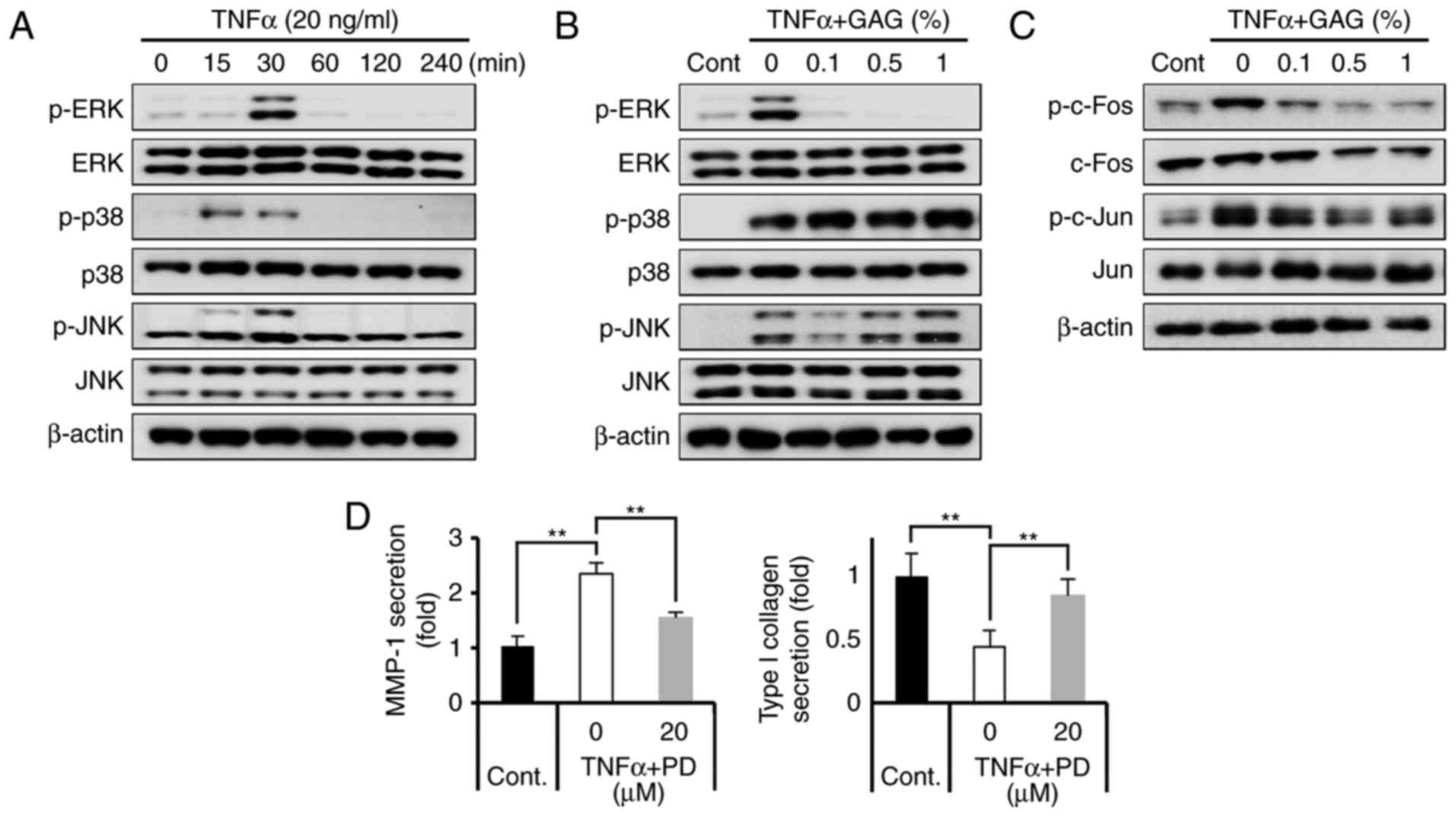 | Figure 3GAGs regulate type I collagen
degradation by inhibiting MMP-1 via the ERK/AP-1 cascade in
TNF-α-stimulated HDFs. (A) HDFs were treated with TNF-α (20 ng/ml)
for 0, 15, 30, 60, 120 and 240 min, and lysates from HDFs were
immunoblotted with anti-p-ERK, anti-ERK, anti-p-p38, anti-p38,
anti-p-JNK, anti-JNK and anti-β-actin antibodies. (B) HDFs were
pretreated with GAGs at the indicated concentrations (0, 0.1, 0.5
and 1%) for 30 min and then stimulated with TNF-α (20 ng/ml) for 30
min. Lysates from HDFs were immunoblotted with anti-p-ERK,
anti-ERK, anti-p-p38, anti-p38, anti-p-JNK, anti-JNK and
anti-β-actin antibodies. (C) Lysates from HDFs were immunoblotted
with anti-p-c-fos, anti-fos, anti-p-c-jun, anti-c-jun and
anti-β-actin antibodies. (D) HDFs were pretreated with 20 μM
PD98059 for 1 h and then stimulated with TNF-α (20 ng/ml) for 24 h.
MMP-1 secretion and type I collagen secretion were subsequently
determined using an ELISA kit. All data are presented as the mean ±
standard error of the mean for three independent experiments.
**P<0.01. GAGs, glycosaminoglycans; MMP-1, matrix
metalloproteinase-1; ERK, extracellular signal-regulated kinase;
TNF-α, tumor necrosis factor-α; HDF, human dermal fibroblast; p-,
phosphorylated; JNK, c-jun N-terminal kinase; Cont, control; PD,
PD98059. |
GAGs inhibit the TNF-α-induced-apoptosis
of HDFs
TNF-α is a pleiotropic cytokine with diverse
cellular responses. TNF-α induces apoptosis in different types of
cells, including fibroblasts (45-47). Therefore, the present study
assessed whether GAGs inhibit TNF-α-induced HDF apoptosis. HDFs
were pretreated with GAGs for 30 min and then stimulated with TNF-α
for 24 h. Cell apoptosis was then evaluated by assessing the
expression of the apoptosis-associated proteins caspase-3,
Bcl-2-associated X protein (Bax) and B-cell lymphoma 2 (Bcl-2).
Caspase-3 serves a key role in TNF-α-mediated apop-tosis (48,49). The results demonstrated that TNF-α
stimulation markedly increased cleaved caspase-3 levels (as
indicated by the bottom band on the western blot in Fig. 4A) and pro-apoptotic Bax levels.
However, treatment with GAGs markedly abolished these increases
(Fig. 4A). Additionally, GAGs
treatment did not alter levels of the anti-apoptotic protein Bcl-2,
but restored the Bax/Bcl-2 ratio following stimulation with TNF-α
in HDFs, in a dose-dependent manner (Fig. 4A and B). These results indicate
that GAGs protect HDFs against the apoptosis induced by TNF-α.
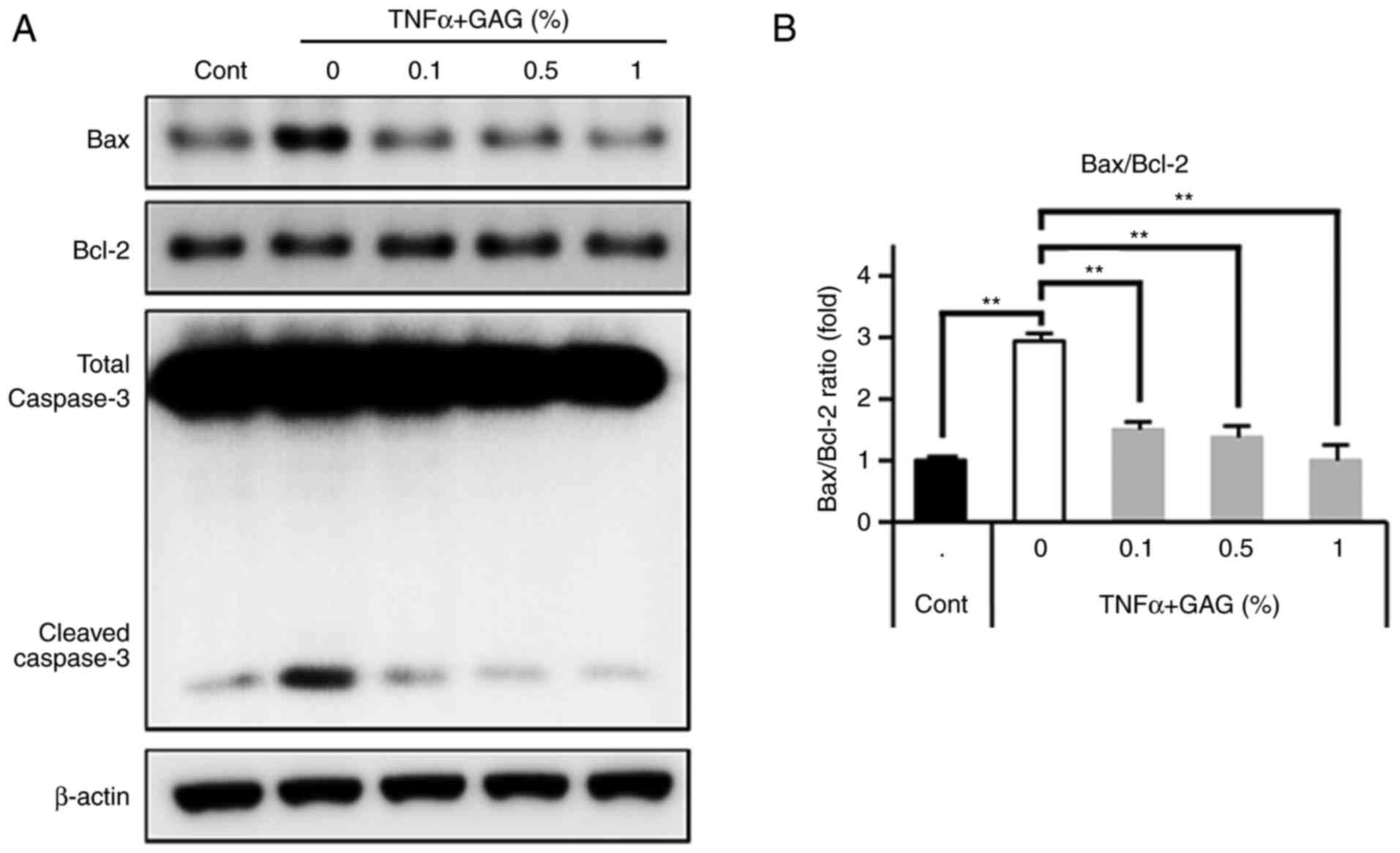 | Figure 4Anti-apoptotic effect of GAGs in
TNF-α-stimulated HDFs. HDFs were pretreated with GAGs at the
indicated concentrations (0, 0.1, 0.5 and 1%) for 30 min and then
stimulated with TNF-α (20 ng/ml) for 24 h. (A) Lysates from HDFs
were immunoblotted with anti-Bax, anti-Bcl-2, anti-caspase-3 and
anti-β-actin antibodies. (B) Western blots were analyzed
quantitatively. All data are presented as the mean ± standard error
of the mean for three independent experiments.
**P<0.01. GAGs, glycosaminoglycans; TNF-α, tumor
necrosis factor-α; HDF, human dermal fibroblast; Bax,
Bcl-2-associated X protein; Bcl-2, B-cell lymphoma 2; Cont,
control. |
Discussion
The various anti-aging benefits of GAGs have
contributed to their widespread inclusion in cosmetic and
pharmaceutical products (50,51). GAGs, including hyaluronic acid,
chondroitin sulfate, dermatan sulfate and keratan sulfate are
abundant structural components of extracellular structures and
regulate many biological processes, including cell growth,
migration, differentiation, adhesion and lipid synthesis in the
dermis and epidermis (1,52-56). The contents of GAGs change during
intrinsic and extrinsic aging (57-60). Specifically, GAGs containing
proteoglycans, including versican, decorin and biglycan, serve
important roles in the synthesis and maintenance of collagen and
elastin (61-64). It has been demonstrated that the
removal of collagen-attached GAGs affects collagen-olysis via
cathepsin K (65). However, few
studies have reported the molecular mechanisms of GAGs underlying
the anti-aging effects on the skin.
To the best of our knowledge, the present study is
the first to demonstrate that GAGs significantly inhibit
TNF-α-induced type I collagen denaturation in HDFs. It has been
demonstrated that versican, a member of the chondroitin sulfate
proteoglycan (PG) family, binds to collagens and controls fibril
formation (64). In addition,
deficiencies in decorin and biglycan, which are members of the
dermatan sulfate PG family, cause abnormal collagen fibril
formation and result in thin and fragile skin (61-63). The keratan sulfate PGs, lumican
and fibromodulin, are also able to regulate the formation of
collagen fibrils (66).
The molecular components of the ECM are remodeled by
matrix metalloproteinases released from fibroblasts (67). Collagen degradation can be induced
by MMP-1 and the expression and activity of MMP-1 may become
elevated as a result of intrinsic and extrinsic aging (68,69). The present study demonstrated that
GAGs inhibit the TNF-α-induced increase in MMP-1 expression in
HDFs. However, the results demonstrated that levels of TIMP-1, an
inhibitor of MMP-1, increased along with the concentration of GAGs
in HDFs stimulated with TNF-α. The results of the present study
therefore support the conclusion that MMPs and TIMPs are directly
and indirectly involved in the synthesis of collagen type I
(40-43).
The present study also assessed the upstream signal
transduction pathways of collagen synthesis via the regulation of
MMP-1 and TIMP-1. Previous studies have demonstrated that MAPK
signaling pathways serve pivotal roles in controlling various
cellular functions, including cell growth, MMP expression and
collagen synthesis (26,30-32). In addition, several studies have
revealed that MMP-1 expression is enhanced by activated MAPK in
dermal fibroblasts following stimulation with particular stimuli
(70,71). The present study demonstrated that
GAG treatment resulted in the suppression of ERK phosphorylation
(but not the phosphorylation of JNK or p38) in TNF-α-induced HDFs,
leading to the reduced phosphorylation of AP-1. It has also
demonstrated that tensile force induces the expression of type I
collagen and MMP-1, and activates MAPKs in periodontal ligament
fibroblasts (72). In particular,
the inhibition of ERK, but not JNK or p38 MAPK, negatively
regulates the tensile force-mediated activation of NF-κB and MMP-1
expression (72). The
TNF-α-induced expression of MMP-9 was increased via the
Ras/ERK-regulated activation of NF-κB and AP-1 in human vascular
smooth muscle cells (73). In
addition, various cytokines, including interleukin (IL)-1β, IL-6,
IL-8 and inducible nitric oxide synthase induced by MAPKs, are
involved in the inflammation associated with skin diseases
(74,75). Thus, further studies are required
to assess the anti-inflammatory effect of GAGs in skin diseases or
in conditions associated with aging.
Finally, the current study confirmed the
anti-apoptotic effects of GAGs in HDF stimulated with TNF-α. It has
established that TNF-α stimulates a variety of responses, including
inflammation and apoptosis in vitro and in vivo
(76-78). The present study demonstrated that
GAGs exert anti-apoptotic effects in HDFs stimulated with TNF-α.
Furthermore, a previous study determined that TNF-α-induced ERK
activation mediates p53 activation in apoptotic and autophagic L929
cells (79). In addition, PD98059
has been demonstrated to significantly reduced the cytotoxic effect
of TNF-α in L929 and U251 cells (80).
In conclusion, the results of the current study
indicate that GAGs serve an important role in type I collagen
production. Specifically, GAGs reduced MMP-1 expression and
elevated TIMP-1 expression by inhibiting the ERK/AP-1 cascade in
TNF-α-stimulated HDFs. In addition, a protective effect of GAGs
against cell apoptosis was observed. A previous study demonstrated
that a cream comprised of GAGs positively regulated wrinkles, skin
elasticity, dermal density and skin tightening in a clinical trial
(4). Given that the expression of
various proinflammatory cytokine/chemokines are significantly
elevated and the expression of type I collagens is decreased in
aged skin dermis, the current study suggests that GAGs may be used
as anti-inflammatory and anti-aging agents for skin.
Acknowledgments
Not applicable.
Notes
[1]
Funding
The present study was supported by Elensilia,
Taeyoung Co., Korea (grant no. 20130994).
[2] Availability
of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
[3] Authors'
contributions
BJK, YL, and JN designed the research. JN, DHB, and
SII conducted the research. BJK, YL, JN, DHB, HC, JHH, SYK, and YAN
analyzed the data. All authors read and approved the final
manuscript.
[4] Ethical
approval and consent to participate
The Ethical committee of Chung-Ang University
Hospital Institute Review Board [IRB no. C2015051 (1509)] approved
the present study. Written informed consent was obtained from the
legal guardians of the donor prior to enrolment.
[5] Consent for
publication
Not applicable.
[6] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Chajra H, Auriol D, Joly F, Pagnon A,
Rodrigues M, Allart S, Redziniak G and Lefevre F: Reactivating the
extracellular matrix synthesis of sulfated glycosaminoglycans and
proteoglycans to improve the human skin aspect and its mechanical
properties. Clin Cosmet Investig Dermatol. 9:461–472. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kim YS, Jo YY, Chang IM, Toida T, Park Y
and Linhardt RJ: A new glycosaminoglycan from the giant African
snail Achatina fulica. J Biol Chem. 271:11750–11755. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shim JY, Lee YS, Jung SH, Choi HS, Shin KH
and Kim YS: Pharmacological activities of a new glycosaminoglycan,
acharan sulfate isolated from the giant African snail Achatina
fulica. Arch Pharm Res. 25:889–894. 2002. View Article : Google Scholar
|
|
4
|
Kim BJ, No YA, Lee Y, Kim MN, Hong CK, Yoo
KH, Kim YM, Hwang JH and Kong SY: Use of cream containing mucus
secreted by snails has an anti-aging effect on skin. Korean J
Dermatol. 53:430–436. 2015.
|
|
5
|
Zeng JP, Bi B, Chen L, Yang P, Guo Y, Zhou
YQ and Liu TY: Repeated exposure of mouse dermal fibroblasts at a
sub-cytotoxic dose of UVB leads to premature senescence: A robust
model of cellular photoaging. J Dermatol Sci. 73:49–56. 2014.
View Article : Google Scholar
|
|
6
|
Gaur M, Dobke M and Lunyak VV: Mesenchymal
stem cells from adipose tissue in clinical applications for
dermatological indications and skin aging. Int J Mol Sci.
18:pii:E208. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
El-Domyati M, Attia S, Saleh F, Brown D,
Birk DE, Gasparro F, Ahmad H and Uitto J: Intrinsic aging vs.
photoaging: A comparative histopathological, immunohistochemical,
and ultrastructural study of skin. Exp Dermatol. 11:398–405. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Svobodová A, Psotová J and Walterová D:
Natural phenolics in the prevention of UV-induced skin damage. A
review. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub.
147:137–145. 2003. View Article : Google Scholar
|
|
9
|
Fisher GJ, Varani J and Voorhees JJ:
Looking older: Fibroblast collapse and therapeutic implications.
Arch Dermatol. 144:666–672. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Quan T and Fisher GJ: Role of
Age-associated alterations of the dermal extracellular matrix
microenvironment in human skin aging: A Mini-review. Gerontology.
61:427–434. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Borg M, Brincat S, Camilleri G,
Schembri-Wismayer P, Brincat M and Calleja-Agius J: The role of
cytokines in skin aging. Climacteric. 16:514–521. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chung JH, Seo AY, Chung SW, Kim MK,
Leeuwenburgh C, Yu BP and Chung HY: Molecular mechanism of PPAR in
the regulation of age-related inflammation. Ageing Res Rev.
7:126–136. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ishitsuka Y, Maniwa F, Koide C, Kato Y,
Nakamura Y, Osawa T, Tanioka M and Miyachi Y: Increased halogenated
tyrosine levels are useful markers of human skin ageing, reflecting
proteins denatured by past skin inflammation. Clin Exp Dermatol.
37:252–258. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bashir MM, Sharma MR and Werth VP: UVB and
proinflammatory cytokines synergistically activate TNF-alpha
production in keratinocytes through enhanced gene transcription. J
Invest Dermatol. 129:994–1001. 2009. View Article : Google Scholar
|
|
15
|
Agius E, Lacy KE, Vukmanovic-Stejic M,
Jagger AL, Papageorgiou AP, Hall S, Reed JR, Curnow SJ,
Fuentes-Duculan J, Buckley CD, et al: Decreased TNF-alpha synthesis
by macrophages restricts cutaneous immunosurveillance by memory
CD4+ T cells during aging. J Exp Med. 206:1929–1940.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dada LA and Sznajder JI: Mitochondrial
Ca2+ and ROS take center stage to orchestrate
TNF-α-mediated inflammatory responses. J Clin Invest.
121:1683–1685. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Han YP, Tuan TL, Wu H, Hughes M and Garner
WL: TNF-alpha stimulates activation of pro-MMP2 in human skin
through NF-(kappa)B mediated induction of MT1-MMP. J Cell Sci.
114:131–139. 2001.
|
|
18
|
Buommino E, De Filippis A, Gaudiello F,
Balato A, Balato N, Tufano MA and Ayala F: Modification of
osteopontin and MMP-9 levels in patients with psoriasis on
anti-TNF-α therapy. Arch Dermatol Res. 304:481–485. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Youn UJ, Nam KW, Kim HS, Choi G, Jeong WS,
Lee MY and Chae S: 3-Deoxysappanchalcone inhibits tumor necrosis
factor-α-induced matrix metalloproteinase-9 expression in human
keratinocytes through activated protein-1 inhibition and nuclear
factor-kappa B DNA binding activity. Biol Pharm Bull. 34:890–893.
2011. View Article : Google Scholar
|
|
20
|
Fisher GJ, Datta SC, Talwar HS, Wang ZQ,
Varani J, Kang S and Voorhees JJ: Molecular basis of sun-induced
premature skin ageing and retinoid antagonism. Nature. 379:335–339.
1996. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dasu MR, Barrow RE, Spies M and Herndon
DN: Matrix metal-loproteinase expression in cytokine stimulated
human dermal fibroblasts. Burns. 29:527–531. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xia W, Hammerberg C, Li Y, He T, Quan T,
Voorhees JJ and Fisher GJ: Expression of catalytically active
matrix metalloproteinase-1 in dermal fibroblasts induces collagen
fragmentation and functional alterations that resemble aged human
skin. Aging Cell. 12:661–671. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Di Lullo GA, Sweeney SM, Korkko J,
Ala-Kokko L and San Antonio JD: Mapping the ligand-binding sites
and disease-associated mutations on the most abundant protein in
the human, type I collagen. J Biol Chem. 277:4223–4231. 2002.
View Article : Google Scholar
|
|
24
|
Cho JW, Il KJ and Lee KS: Downregulation
of type I collagen expression in silibinin-treated human skin
fibroblasts by blocking the activation of Smad2/3-dependent
signaling pathways: Potential therapeutic use in the
chemoprevention of keloids. Int J Mol Med. 31:1148–1152. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Helenius M, Mäkeläinen L and Salminen A:
Attenuation of NF-kappaB signaling response to UVB light during
cellular senescence. Exp Cell Res. 248:194–202. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jin MH, Park SG, Hwang YL, Lee MH, Jeong
NJ, Roh SS, Lee Y, Kim CD and Lee JH: Cedrol enhances extracellular
matrix production in dermal fibroblasts in a MAPK-dependent manner.
Ann Dermatol. 24:16–21. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kim DS, Park SH and Park KC: Transforming
growth factor-beta1 decreases melanin synthesis via delayed
extracellular signal-regulated kinase activation. Int J Biochem
Cell Biol. 36:1482–1491. 2004.PubMed/NCBI
|
|
28
|
Lee DJ, Rosenfeldt H and Grinnell F:
Activation of ERK and p38 MAP kinases in human fibroblasts during
collagen matrix contraction. Exp Cell Res. 257:190–197. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Uitto J: Connective tissue biochemistry of
the aging dermis. Age-related alterations in collagen and elastin.
Dermatol Clin. 4:433–446. 1986.PubMed/NCBI
|
|
30
|
Katiyar SK and Meeran SM: Obesity
increases the risk of UV radiation-induced oxidative stress and
activation of MAPK and NF-kappaB signaling. Free Radic Biol Med.
42:299–310. 2007. View Article : Google Scholar
|
|
31
|
Muthusamy V and Piva TJ: The UV response
of the skin: A review of the MAPK, NFkappaB and TNFalpha signal
transduction pathways. Arch Dermatol Res. 302:5–17. 2010.
View Article : Google Scholar
|
|
32
|
Ghosh AK: Factors involved in the
regulation of type I collagen gene expression: Implication in
fibrosis. Exp Biol Med. 227:301–314. 2002. View Article : Google Scholar
|
|
33
|
Reunanen N, Foschi M, Han J and Kahari VM:
Activation of extracellular signal-regulated kinase 1/2 inhibits
type I collagen expression by human skin fibroblasts. J Biol Chem.
275:34634–34639. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yun HJ, Yoo WH, Han MK, Lee YR, Kim JS and
Lee SI: Epigallocatechin-3-gallate suppresses TNF-alpha-induced
production of MMP-1 and-3 in rheumatoid arthritis synovial
fibroblasts. Rheumatol Int. 29:23–29. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fushimi K, Nakashima S, You F, Takigawa M
and Shimizu K: Prostaglandin E2 downregulates TNF-alpha-induced
production of matrix metalloproteinase-1 in HCS-2/8 chondrocytes by
inhibiting Raf-1/MEK/ERK cascade through EP4 prostanoid receptor
activation. J Cell Biochem. 100:783–793. 2007. View Article : Google Scholar
|
|
36
|
Lee CS, Bae IH, Han J, Choi GY, Hwang KH,
Kim DH, Yeom MH, Park YH and Park M: Compound K inhibits MMP-1
expression through suppression of c-Src-dependent ERK activation in
TNF-α-stimulated dermal fibroblast. Exp Dermatol. 23:819–824. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Twentyman PR and Luscombe M: A study of
some variables in a tetrazolium dye (MTT) based assay for cell
growth and chemo-sensitivity. Br J Cancer. 56:279–285. 1987.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
39
|
Na J, Lee K, Na W, Shin JY, Lee MJ, Yune
TY, Lee HK, Jung HS, Kim WS and Ju BG: Histone H3K27 demethylase
JMJD3 in cooperation with NF-κB regulates keratinocyte wound
healing. J Invest Dermatol. 136:847–858. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Song KC, Chang TS, Lee H, Kim J, Park JH
and Hwang GS: Processed panax ginseng, Sun ginseng increases type I
collagen by regulating MMP-1 and TIMP-1 expression in human dermal
fibroblasts. J Ginseng Res. 36:61–67. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xia W, Quan T, Hammerberg C, Voorhees JJ
and Fisher GJ: A mouse model of skin aging: Fragmentation of dermal
collagen fibrils and reduced fibroblast spreading due to expression
of human matrix metalloproteinase-1. J Dermatol Sci. 78:79–82.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Fisher GJ, Quan T, Purohit T, Shao Y, Cho
MK, He T, Varani J, Kang S and Voorhees JJ: Collagen fragmentation
promotes oxidative stress and elevates matrix metalloproteinase-1
in fibroblasts in aged human skin. Am J Pathol. 174:101–114. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Majewska N, Zareba I, Surazynski A and
Galicka A: Methylparaben-induced decrease in collagen production
and viability of cultured human dermal fibroblasts. J Appl Toxicol.
37:1117–1124. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tsai CL, Chen WC, Lee IT, Chi PL, Cheng SE
and Yang CM: c-Src-dependent transactivation of PDGFR contributes
to TNF-α-induced MMP-9 expression and functional impairment in
osteoblasts. Bone. 60:186–197. 2014. View Article : Google Scholar
|
|
45
|
Laster SM, Wood JG and Gooding LR: Tumor
necrosis factor can induce both apoptic and necrotic forms of cell
lysis. J Immunol. 141:2629–2634. 1988.PubMed/NCBI
|
|
46
|
Fehsel K, Kolb-Bachofen V and Kolb H:
Analysis of TNF alpha-induced DNA strand breaks at the single cell
level. Am J Pathol. 139:251–254. 1991.PubMed/NCBI
|
|
47
|
Alikhani M, Alikhani Z, Raptis M and
Graves DT: TNF-alpha in vivo stimulates apoptosis in fibroblasts
through caspase-8 activation and modulates the expression of
pro-apoptotic genes. J Cell Physiol. 201:341–348. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Beyaert R, Kidd VJ, Cornelis S, Van de
Craen M, Denecker G, Lahti JM, Gururajan R, Vandenabeele P and
Fiers W: Cleavage of PITSLRE kinases by ICE/CASP-1 and CPP32/CASP-3
during apoptosis induced by tumor necrosis factor. J Biol Chem.
272:11694–11697. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Jaeschke H, Fisher MA, Lawson JA, Simmons
CA, Farhood A and Jones DA: Activation of caspase 3 (CPP32)-like
proteases is essential for TNF-alpha-induced hepatic parenchymal
cell apoptosis and neutrophil-mediated necrosis in a murine
endotoxin shock model. J Immunol. 160:3480–3486. 1998.PubMed/NCBI
|
|
50
|
Oh JH, Kim YK, Jung JY, Shin JE and Chung
JH: Changes in glycosaminoglycans and related proteoglycans in
intrinsically aged human skin in vivo. Exp Dermatol. 20:454–456.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Lee DH, Oh JH and Chung JH:
Glycosaminoglycan and proteoglycan in skin aging. J Dermatol Sci.
83:174–181. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Carrino DA, Onnerfjord P, Sandy JD,
Cs-Szabo G, Scott PG, Sorrell JM, Heinegård D and Caplan AI:
Age-related changes in the proteoglycans of human skin. Specific
cleavage of decorin to yield a major catabolic fragment in adult
skin. J Biol Chem. 278:17566–17572. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zimmermann DR, Dours-Zimmermann MT,
Schubert M and Bruckner-Tuderman L: Versican is expressed in the
proliferating zone in the epidermis and in association with the
elastic network of the dermis. J Cell Biol. 124:817–825. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Knott A, Reuschlein K, Lucius R, Stäb F,
Wenck H and Gallinat S: Deregulation of versican and elastin
binding protein in solar elastosis. Biogerontology. 10:181–190.
2009. View Article : Google Scholar
|
|
55
|
Bourguignon LY: Matrix
hyaluronan-activated CD44 signaling promotes keratinocyte
activities and improves abnormal epidermal functions. Am J Pathol.
184:1912–1919. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Anderegg U, Simon JC and Averbeck M: More
than just a filler-the role of hyaluronan for skin homeostasis. Exp
Dermatol. 23:295–303. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Oh JH, Kim YK, Jung JY, Shin JE, Kim KH,
Cho KH, Eun HC and Chung JH: Intrinsic aging- and
photoaging-dependent level changes of glycosaminoglycans and their
correlation with water content in human skin. J Dermatol Sci.
62:192–201. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Meyer LJ and Stern R: Age-dependent
changes of hyaluronan in human skin. J Invest Dermatol.
102:385–389. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Tzellos TG, Sinopidis X, Kyrgidis A,
Vahtsevanos K, Triaridis S, Printza A, Klagas I, Karakiulakis G and
Papakonstantinou E: Differential hyaluronan homeostasis and
expression of proteoglycans in juvenile and adult human skin. J
Dermatol Sci. 61:69–72. 2011. View Article : Google Scholar
|
|
60
|
Willen MD, Sorrell JM, Lekan CC, Davis BR
and Caplan AI: Patterns of glycosaminoglycan/proteoglycan
immunostaining in human skin during aging. J Invest Dermatol.
96:968–974. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Corsi A, Xu T, Chen XD, Boyde A, Liang J,
Mankani M, Sommer B, Iozzo RV, Eichstetter I, Robey PG, et al:
Phenotypic effects of biglycan deficiency are linked to collagen
fibril abnormalities, are synergized by decorin deficiency, and
mimic Ehlers-Danlos-like changes in bone and other connective
tissues. J Bone Miner Res. 17:1180–1189. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Danielson KG, Baribault H, Holmes DF,
Graham H, Kadler KE and Iozzo RV: Targeted disruption of decorin
leads to abnormal collagen fibril morphology and skin fragility. J
Cell Biol. 136:729–743. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Li Y, Xia W, Liu Y, Remmer HA, Voorhees J
and Fisher GJ: Solar ultraviolet irradiation induces decorin
degradation in human skin likely via neutrophil elastase. PLoS One.
8:e725632013. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Carrino DA, Sorrell JM and Caplan AI:
Age-related changes in the proteoglycans of human skin. Arch
Biochem Biophys. 373:91–101. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Panwar P, Butler GS, Jamroz A, Azizi P,
Overall CM and Brömme D: Aging-associated modifications of collagen
affect its degradation by matrix metalloproteinases. Matrix Biol.
Jun 17–2017.Epub ahead of print. PubMed/NCBI
|
|
66
|
Shin JE, Oh JH, Kim YK, Jung JY and Chung
JH: Transcriptional regulation of proteoglycans and
glycosaminoglycan chainsynthe-sizing glycosyltransferases by UV
irradiation in cultured human dermal fibroblasts. J Korean Med Sci.
26:417–424. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Visse R and Nagase H: Matrix
metalloproteinases and tissue inhibitors of metalloproteinases:
Structure, function, and biochemistry. Circ Res. 92:827–839. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Brenneisen P, Sies H and
Scharffetter-Kochanek K: Ultraviolet-B irradiation and matrix
metalloproteinases: From induction via signaling to initial events.
Ann N Y Acad Sci. 973:31–43. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Hwang E, Lee TH, Park SY, Yi TH and Kim
SY: Enzyme-modified Panax ginseng inhibits UVB-induced skin aging
through the regulation of procollagen type I and MMP-1 expression.
Food Funct. 5:265–274. 2014. View Article : Google Scholar
|
|
70
|
Park CH, Moon Y, Shin CM and Chung JH:
Cyclic AMP suppresses matrix metalloproteinase-1 expression through
inhibition of MAPK and GSK-3beta. J Invest Dermatol. 130:2049–2056.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Reunanen N, Westermarck J, Häkkinen L,
Holmström TH, Elo I, Eriksson JE and Kähäri VM: Enhancement of
fibroblast collagenase (matrix metalloproteinase-1) gene expression
by ceramide is mediated by extracellular signal-regulated and
stress-activated protein kinase pathways. J Biol Chem.
273:5137–5145. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Kook SH, Jang YS and Lee JC: Involvement
of JNK-AP-1 and ERK-NF-κB signaling in tension-stimulated
expression of type I collagen and MMP-1 in human periodontal
ligament fibroblasts. J Appl Physiol. 111:1575–1583. 2011.
View Article : Google Scholar
|
|
73
|
Moon SK, Cha BY and Kim CH: ERK1/2
mediates TNF-alpha-induced matrix metalloproteinase-9 expression in
human vascular smooth muscle cells via the regulation of NF-kappaB
and AP-1: Involvement of the ras dependent pathway. J Cell Physiol.
198:417–427. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Sebastiani S, Albanesi C, De PO, Puddu P,
Cavani A and Girolomoni G: The role of chemokines in allergic
contact dermatitis. Arch Dermatol Res. 293:552–559. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Mattii M, Ayala F, Balato N, Filotico R,
Lembo S, Schiattarella M, Patruno C, Marone G and Balato A: The
balance between pro- and anti-inflammatory cytokines is crucial in
human allergic contact dermatitis pathogenesis: The role of IL-1
family members. Exp Dermatol. 22:813–819. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Ruan W, Xu JM, Li SB, Yuan LQ and Dai RP:
Effects of down-regulation of microRNA-23a on TNF-α-induced
endothelial cell apoptosis through caspase-dependent pathways.
Cardiovasc Res. 93:623–632. 2011. View Article : Google Scholar
|
|
77
|
Markelic M, Velickovic K, Golic I,
Otasevic V, Stancic A, Jankovic A, Vucetic M, Buzadzic B, Korac B
and Korac A: Endothelial cell apoptosis in brown adipose tissue of
rats induced by hyperinsulinaemia: The possible role of TNF-α. Eur
J Histochem. 55:e342011. View Article : Google Scholar
|
|
78
|
Skoog T, Dichtl W, Boquist S,
Skoglund-Andersson C, Karpe F, Tang R, Bond MG, de Faire U, Nilsson
J, Eriksson P and Hamsten A: Plasma tumour necrosis factor-alpha
and early carotid atherosclerosis in healthy middle-aged men. Eur
Heart J. 23:376–383. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Cheng Y, Qiu F, Tashiro S, Onodera S and
Ikejima T: ERK and JNK mediate TNFalpha-induced p53 activation in
apoptotic and autophagic L929 cell death. Biochem Biophys Res
Commun. 376:483–488. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Harhaji L, Mijatovic S, Maksimovic-Ivanic
D, Popadic D, Isakovic A, Todorovic-Markovic B and Trajkovic V:
Aloe emodin inhibits the cytotoxic action of tumor necrosis factor.
Eur J Pharmacol. 568:248–259. 2007. View Article : Google Scholar : PubMed/NCBI
|