Introduction
During an individual's lifetime, human skin is
continuously exposed to harmful environmental factors, such as
ultra violet (UV) light. Even though UV light stimulates the
synthesis of vitamin D (1), which
has beneficial effects in terms of regulating the immune system and
calcium homeostasis, excessive UV exposure can cause oxidative
damage (2,3), immune suppression (4), collagen degradation (5,6),
photoaging, and skin cancer (7–9).
UV irradiation induces the synthesis of matrix metalloproteinases
(MMPs) in the skin (5,6,10).
MMPs are a family of zinc-dependent endopeptidases that play a
major role in extracellular matrix (ECM) degradation, including
collagen fibers. More specifically, UV induces activation of
mitogen-activated protein kinase (MAPK) components, such as
extracellular signal-regulated kinase (ERK), c-Jun N-terminal
kinase (JNK), and p38 MAPK (p38), which in turn regulate activator
protein-1 (AP-1), leading to increased MMPs expression, and
decreased collagen production (10,11).
Black rice is a grain from the species Oryza
sativa L. var. japonica. It contains various
polyphenolic compounds, such as anthocyanins (12), which have antioxidative activity.
It has been reported that black rice extract (BRE) has many
pharmacological effects, including antioxidant (13) and anti-inflammatory (14–16) activity. Furthermore, black rice
anthocyanin extracts have been studied in various types of cancer,
and have been shown to suppress cancer cell invasion (17), metastasis (18), and angiogenesis (19,20). Additionally, black rice bran
inhibits tyrosinase activity in cell-free in vitro systems,
suggesting its potential as a melanogenesis inhibitor (21). Germinated black rice also enhances
hyaluronan production in HaCaT keratinocytes (22). However, it remains unknown whether
BRE prevents UV-induced ECM alteration. Therefore, the aim of the
present study was to investigate the effects of BRE in
UV-irradiated HaCaT cells, and human dermal fibroblasts (HDF), by
analyzing the expression of MMPs and collagen, and generation of
reactive oxygen species (ROS), as well as the underlying signaling
pathways involved.
Materials and methods
Reagents and antibodies
3-(4,5-dimethylthiazol-2-yl)-2,
5-diphenyltetrazolium bromide (MTT), 2′7′-dichlorofluorescein
diacetate (DCF-DA), N-Acetyl-L-cysteine (NAC) and
phosphatase inhibitor cocktail were obtained from Sigma-Aldrich
(St. Louis, MO, USA). Protease inhibitor cocktail tablets were
purchased from Roche Applied Science (Penzberg, Germany).
Anti-MMP-1 antibody (Oncogene Research Products, Boston, MA, USA)
and monoclonal anti-procollagen type I N-terminal extension peptide
(SP1.D8) antibody (Developmental Studies Hybridoma Bank, Iowa City,
IA, USA) were used. Phospho- ERK1/2, JNK, p38 MAPK, and c-Jun, and
the total ERK1/2, JNK, p38 MAPK, and c-Jun were purchased from Cell
Signaling Technology, Inc. (Beverly, MA, USA). Anti-c-Fos, β-actin
and lamin B were purchased from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA).
Preparation of BRE
BRE was prepared, and provided by the Korea Food
Research Institute (Seongnam-si, Korea). Briefly, whole grains of
black rice were extracted with 50% ethanol using a microwave
system. The ethanol solvent was then evaporated using a freeze
dryer (yield: 2.68% of dry wt). Stock solutions of BRE were
prepared by dissolving the powder in DMSO, and stored at −20°C. The
final concentration of DMSO in the medium was kept below 0.1%.
Cell line and culture conditions
Two types of cells were used in this study: HaCaT
cells, which is a spontaneously transformed human keratinocyte cell
line and primary HDF, isolated from the foreskin of young
volunteers (aged 10–19 years). The present study was approved by
the Institutional Review Board (IRB no. 1101-116-353) at Seoul
National University Hospital and conducted according to the
Declaration of Helsinki. All subjects provided written informed
consent. HaCaT (immortalized human keratinocytes) and HDF between
fifth to fifteenth passages were used for all experiments. The
HaCaT cells and HDF were maintained in Dulbecco's modified Eagle's
medium (DMEM; Welgene, Daegu, Korea) containing 10% fetal bovine
serum (FBS; Welgene) and penicillin/streptomycin (400 U/ml, 50 g/l)
at 37°C in a humidified condition with 5% CO2.
UV irradiation and BRE treatment
The cells were starved of media for 24 h, and washed
twice and replaced with PBS prior to UV irradiation. Philips TL
20W/12 RS fluorescent sun lamps with an emission spectrum between
275 and 380 nm (peak, 310–315 nm) were used as the UV source, as
previously described (23). To
block UVC (<290 nm) wavelengths, Kodacel filter (TA401/407;
Kodak, Rochester, NY, USA) was placed 2 cm in front of UV lamp.
Waldmann UV meter (model 585100, Villingen-Schwenningen, Germany)
was used to measure UV irradiance. Immediately after UV irradiation
(HaCaT cell; 50 mJ/cm2, HDF; 100 mJ/cm2), PBS
was replaced with serum-free media with or without BRE for
indicated time periods.
MTT assay
The cell viability was measured by MTT assay. The
HaCaT (2×104 cells/well) and HDF (5×104
cells/well) cells were seeded into 96-well plates and were maintain
until 80% confluency, and then cells were UV-irradiated or
non-irradiated. The cells were post-treated with various
concentrations of BRE for 48 h. After incubation, MTT solution (0.5
mg/ml in PBS) was added into culture plates and further incubated
for 4 h. The formazan crystals were dissolved in DMSO after culture
media was removed and were quantified at 570 nm using ELISA reader
(Thermo Fisher Scientific, Waltham, MA, USA).
Nuclear protein extraction
HaCaT cells were scraped from culture dishes,
suspended in ice-cold cytoplasmic extraction buffer (20 mM Tris,
1.5 mM MgCl2, 10 mM KCl, 0.2 mM EDTA, 0.5 mM DTT, 0.5% Nonidet
P-40, protease inhibitor cocktail) and incubated on ice for 10 min.
Subsequently, nuclear pellets were collected by centrifugation at
6,000 rpm for 5 min at 4°C and were re-suspended with extraction
buffer (cytoplasmic extraction buffer with 400 mM NaCl, 5%
glycerol) and incubated on ice for 30 min with shaking. The pellets
were centrifuged at 12,000 rpm for 10 min at 4°C to obtain the
nuclear fraction.
Western blot analysis
To determine the amount of MMPs and procollagen
secreted into the culture medium, equal amounts of culture medium
were separated using polyacrylamide gel electrophoresis, and then
the separated proteins were electro-transferred onto a PVDF
membrane (Amersham, Buckinghamshire, UK). The membrane was blocked
with blocking buffer, and then incubated with appropriate primary
antibodies. After incubation, the membranes were washed, then
incubated with horseradish peroxidase-conjugated secondary
antibody. Enhanced chemiluminescence detection system (GE
Healthcare) was used to visualize the protein bands. To analyze
MAPK signaling, HaCaT cells were lysed with RIPA lysis buffer
(Millipore Corp., Billerica, MA, USA) containing protease and
phosphatase inhibitors. Furthermore, to analyze AP-1 activation,
the nuclear proteins were extracted from the cells as described
above.
Reverse transcription and real-time
quantitative PCR (RT-qPCR)
RNAiso Plus (Takara Bio Inc., Shiga, Japan) was used
to isolate the total RNA from the HaCaT cells and HDF (24). Isolated total RNA (1 μg)
was used as templates for cDNA synthesis using a First Strand cDNA
Synthesis kit (MBI Fermentas, Vilnius, Lithuania) according to the
manufacturer's instructions, which was then used for real-time PCR.
The PCR cycling conditions were 50°C for 2 min and 95°C for 2 min,
followed by 40 cycles at 95°C for 15 sec and 60°C for 1 min.
Quantitation of PCR reactions was performed by 7500 Real-time PCR
System (Applied Biosystems Life Technologies, Foster City, CA, USA)
with the SYBR Premix Ex Taq II kit (Takara Bio Inc.) using suitable
primers as follows: hMMP-1 forward, 5′-AAGCGTGTGACAGTAAGCTA-3′ and
reverse, 5′-AACCGGACTTCATCTCTG-3′, hMMP-3 forward,
5′-CTCACAGACCTGACTCGGTT 3′ and reverse, 5′-CACGCCTGAAGGAAGAGATG-3′,
h type I procollagen forward, 5′-CTCGAGGTGGACACCACCCT-3′ and
reverse, 5′-CAGCTGGATGGCCACATCGG-3′ h36B4 forward,
5′-TCGACAATGGCAGCATCTAC-3′ and reverse, 5′-TGATGCAACAGTTGGGTAGC-3′.
Relative mRNA expression was analyzed using the 2−ΔΔCt
methods, and 36B4 was used as an internal control. Data are
expressed as the fold number of gene expression.
Measurement of ROS production
To quantify intracellular oxidative stress, the
cells were pretreated with NAC (2 μM) or BRE for 1 h, and
then washed with PBS. After washing, the cells were labeled with 20
mM DCF-DA at 37°C for 1 h. To induce ROS generation, the cells were
exposed to UV irradiation at a dose of 50 mJ/cm2. The
cells were then washed twice with PBS, and the relative ROS level
was determined using a Victor3 multilabel plate reader
(PerkinElmer, Inc., Waltham, MA, USA), with an excitation
wavelength of 485 nm and an emission wavelength of 535 nm. At the
same time, fluorescence images were captured under a fluorescence
microscope (DMIL; Leica Microsystems GmbH, Wetzlar, Germany) with a
20×0.75 NA objective lens.
Ultra performance liquid
chromatography-quadrupole-time of flight (UPLC-Q-TOF) mass
spectrometry (MS)
To identify black rice metabolites, metabolites from
50% ethanolic extract of black rice were analyzed by an ultra
performance liquid chromatography-quadrupole-time of flight
(UPLC-Q-TOF) MS (Waters Corp., Milford, MA, USA). The extract was
injected into an Acquity UPLC BEH C18 column (2.1×100 mm, 1.7
μm; Waters Corp.) at a column temperature of 40°C. Mobile
phase consisted of water with 0.1% formic acid (FA) and
acetonitrile with 0.1% FA at a flow rate of 0.35 ml/min for 9 min.
The eluents were ionized by electrospray ionization (ESI) with
positive or negative mode and analyzed using a Q-TOF MS. The scan
range of TOF MS data was from 50 to 1,500 m/z with a scan time of
0.2 sec. The capillary voltage was set at 3 or 2.5 kV for positive
mode and negative mode, respectively, while the sample cone voltage
was 40 V. The desolvation flow rate was 900 l/h at a temperature of
400°C and source temperature set to 100°C. Leucine-enkephalin
([M+H] = m/z 556.2771) was used as a reference for lock mass at a
frequency of 10 sec. The MS/MS spectra were obtained using
collision energy ramps from 20 to 45 eV. Metabolites were
identified by Unifi software with various LC/MS databases.
Statistical analysis
The results are presented as means ± SEM, and
statistical analyses were performed using a Student's t-test or
one-way ANOVA. P<0.05 was considered to indicate a statistically
significant difference.
Results
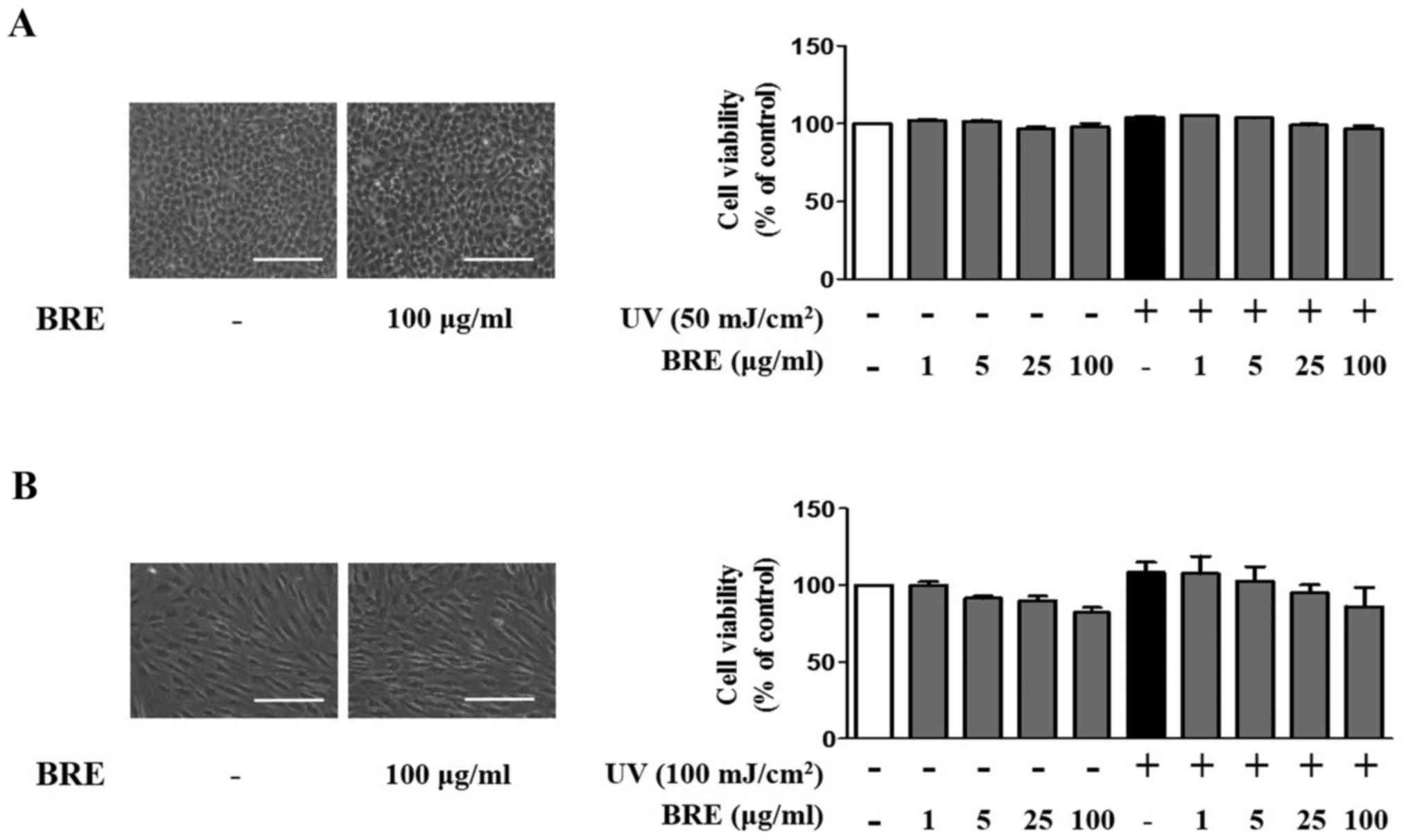
Effects of BRE on cell viability in HaCaT
and HDF
To examine the effect of BRE on skin cells, we first
tested the cytotoxicity of BRE using an MTT assay. BRE (5–100
μg/ml) did not influence the viability of HaCaT cells, with
or without UV irradiation (Fig.
1A). A slight, but not significant, reduction in the viability
of HDF was observed after treatment with 5–100 μg/ml of BRE
(Fig. 1B). Under the same
experimental conditions, 100 μg/ml of BRE did not change
cell morphology. These results indicate that BRE is not
significantly cytotoxic to HaCaT cells or HDF. Therefore, 5–100
μg/ml of BRE was used for the subsequent experiments.
BRE inhibits MMP-1 expression in HaCaT
cells
To determine the effect of BRE on MMP-1 production
in HaCaT cells, the cells were post-treated with BRE for 48 h, with
or without UV irradiation. The basal level of MMP-1 expression was
decreased to 24% in 100 μg/ml BRE treated cells compared
with non-treated cells by western blot analysis. Additionally,
MMP-1 mRNA level also showed similar result. BRE reduced basal
MMP-1 expression in a dose-dependent manner (0.5±0.2, 0.3±0.1, and
0.2±0.1-fold, respectively) (Fig.
2A). Furthermore, BRE ameliorated UV-induced MMP-1 expression
(158% of control) in a concentration-dependent manner (25, and 100
μg/ml BRE showed 92 and 36% of control, respectively). The
MMP-1 mRNA level was 1.8±0.3-fold higher in the UV-exposed cells
than in the control cells. However, when the cells were treated
with 5, 25, and 100 μg/ml BRE, the expression of MMP-1 was
markedly suppressed (1.5±0.2, 1.3±0.1, and 0.5±0.1-fold,
respectively) (Fig. 2B).
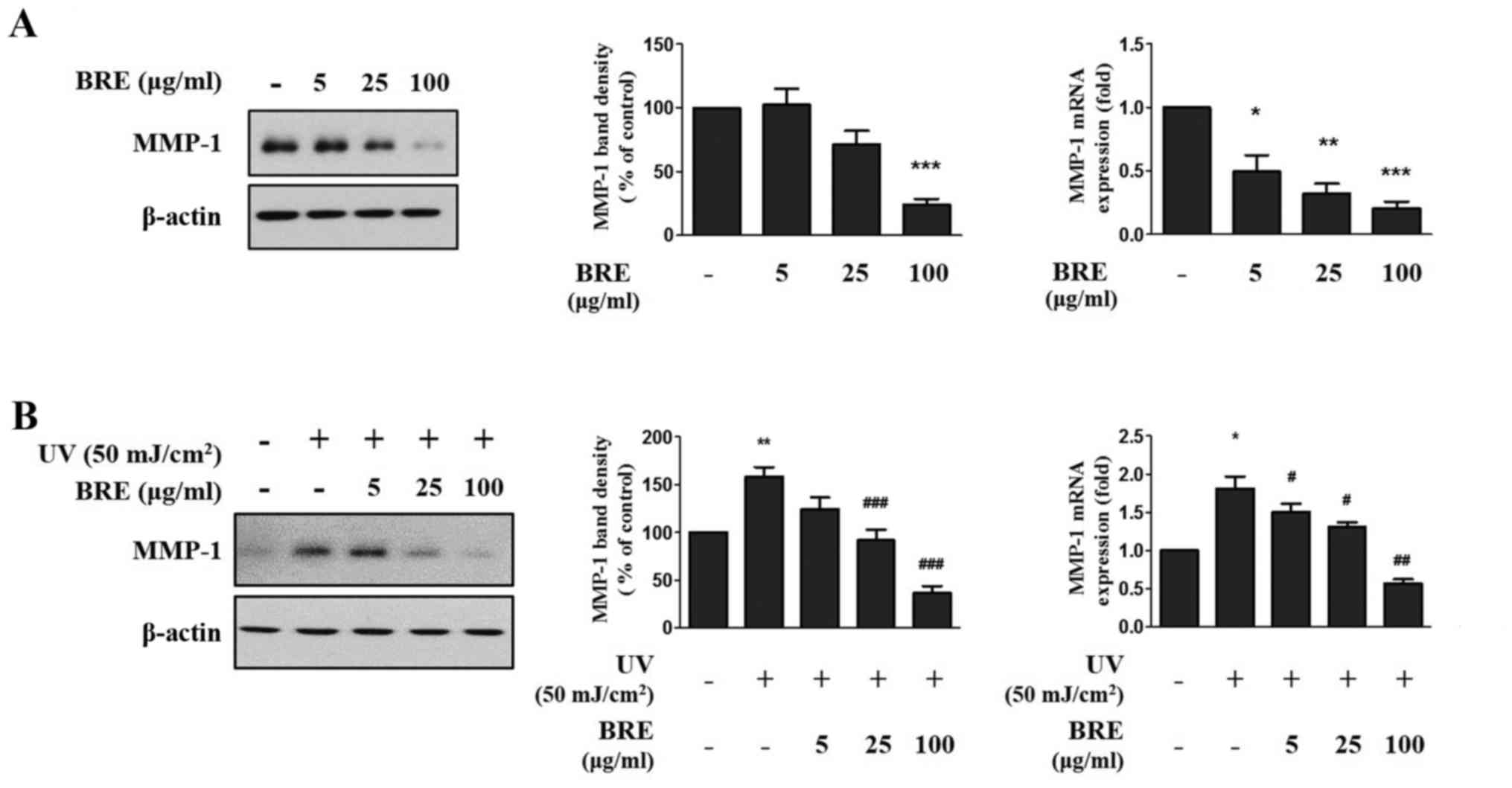 | Figure 2The inhibition effect of BRE on MMP-1
expression in HaCaT cells. HaCaT cells were post-treated with the
indicated concentration of BRE for 48 h, (A) without or (B) with UV
irradiation (50 mJ/cm2). The MMP-1 protein level in the
conditioned medium was investigated by western blot analysis and
relative band density was analyzed using ImageJ software. Actin was
used as endogenous control. The MMP-1 mRNA level was analyzed by
RT-qPCR, respectively. 36B4 mRNA was used to normalize each mRNA
expression level. The bar graphs show the mean ± SEM of three
independent experiments. *P<0.05,
**P<0.01, ***P<0.001 compared with the
non-irradiated control cells. #P<0.05,
##P<0.01, ###P<0.001 compared with the
UV-irradiated control cells. BRE, black rice extract; MMP, matrix
metalloproteinase; UV, ultraviolet; RT-qPCR, reverse
transcription-quantitative PCR. |
BRE suppresses UV-induced MMP-1 and MMP-3
expression, and decreased procollagen expression in HDF
To investigate the effect of BRE in UV-irradiated
HDF, the cells were post-treated with BRE for 48 h after UV
irradiation (100 mJ/cm2). The amount of procollagen and
MMPs (MMP-1, MMP-2, and MMP-3) secreted into the culture medium was
analyzed, and the mRNA levels were analyzed using total RNA. The
results showed that MMP-1 and MMP-3 secretion dramatically
increased in the UV-irradiated HDF compared to that in the control
cells (200 and 430% of the control, respectively), but treatment
with 5, 25, or 100 μg/ml of BRE inhibited UV-induced MMP-1
(115, 80 and 31% of the control, respectively) and MMP-3 production
(315, 320 and 155% of the control, respectively). Furthermore,
UV-induced suppression of procollagen production (46% of control)
was recovered in 100 μg/ml of BRE treated cells (80% of
control) (Fig. 3A and 3B). The MMP-2 level was unaffected, and
served as a loading control. The expression level of MMP-1 in the
UV irradiated cells was 4.4±1.0-fold higher than that in the
non-irradiated cells. However, treatment with 25 and 100
μg/ml BRE significantly reduced the MMP-1 expression level
to just 2.1±1.0 and 1.5±0.8-fold higher than the control value,
respectively. The MMP-3 expression level was also significantly
upregulated by UV irradiation (10.0±4.9-fold). However, the cells
treated with 100 μg/ml of BRE showed significantly inhibited
MMP-3 expression (2.9±2.0-fold). Next, we investigated the change
in procollagen production following UV and BRE treatment. While
type I procollagen production was suppressed (0.4±0.3-fold) in the
HDF following UV exposure, treatment with 100 μg/ml BRE
increased the type I procollagen protein and mRNA levels
(0.6±0.3-fold) in the HDF (Fig.
3C).
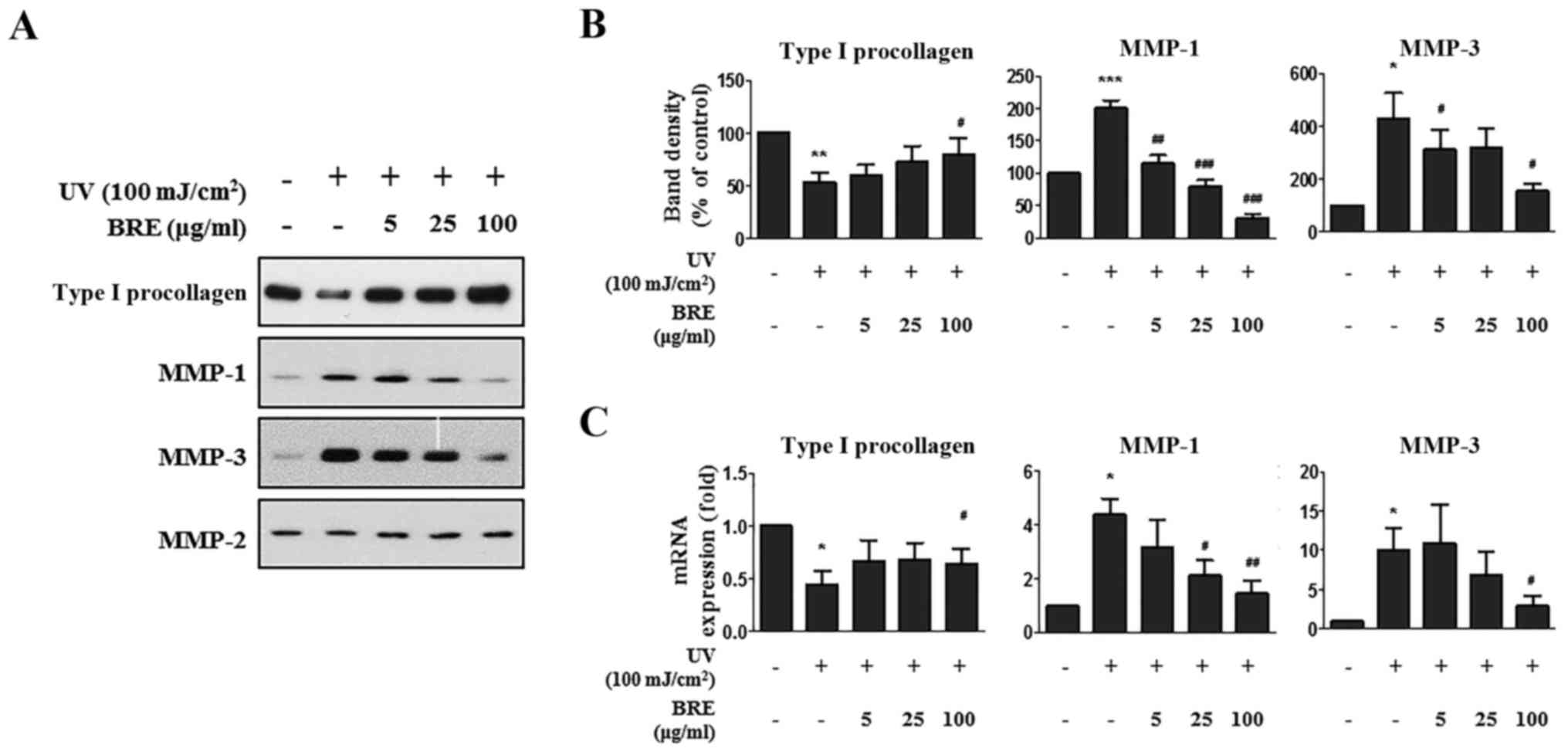 | Figure 3The effect of BRE on MMPs and
procollagen expression in HDF. HDF were exposed to 100
mJ/cm2 of UV irradiation and post-treated with BRE. The
MMPs and procollagen expression levels were then examined. (A) The
western blot shows the MMP-1, MMP-2, MMP-3, and procollagen protein
levels in the conditioned medium from the UV-irradiated HDF treated
with BRE for 48 h. (B) Relative band density was analyzed using
ImageJ software. MMP-2 was used as endogenous control. (C) The
MMP-1, MMP-3, and procollagen mRNA levels in the UV-exposed HDF,
following treatment with the indicated concentration of BRE for 48
h, were assessed using RT-qPCR. 36B4 mRNA was used to normalize
each mRNA expression level. The graphs show the mean ± SEM of three
independent experiments. *P<0.05,
**P<0.01, ***P<0.001 compared with the
non-irradiated control cells. #P<0.05,
##P<0.01, ###P<0.001 compared with the
UV-irradiated control cells. BRE, black rice extract; MMP, matrix
metalloproteinase; HDF, human dermal fibroblasts; UV, ultraviolet;
RT-qPCR, reverse transcription-quantitative PCR. |
Effect of BRE on UV-induced MAPK
signaling and AP-1 activity
To elucidate whether MAPK signaling pathways are
involved in the attenuation of UV-induced MMP-1 expression in HaCaT
cells, the cells were treated with the indicated concentrations of
BRE for 4 h before UV irradiation (50 mJ/cm2). As shown
in Fig. 4A, UV irradiation
significantly increased the phosphorylation of ERK, JNK, and p38
within 15 min. Post-treatment with BRE diminished the UV-induced
phosphorylation of JNK and p38 in a dose-dependent manner, but ERK
phosphorylation was unaffected.
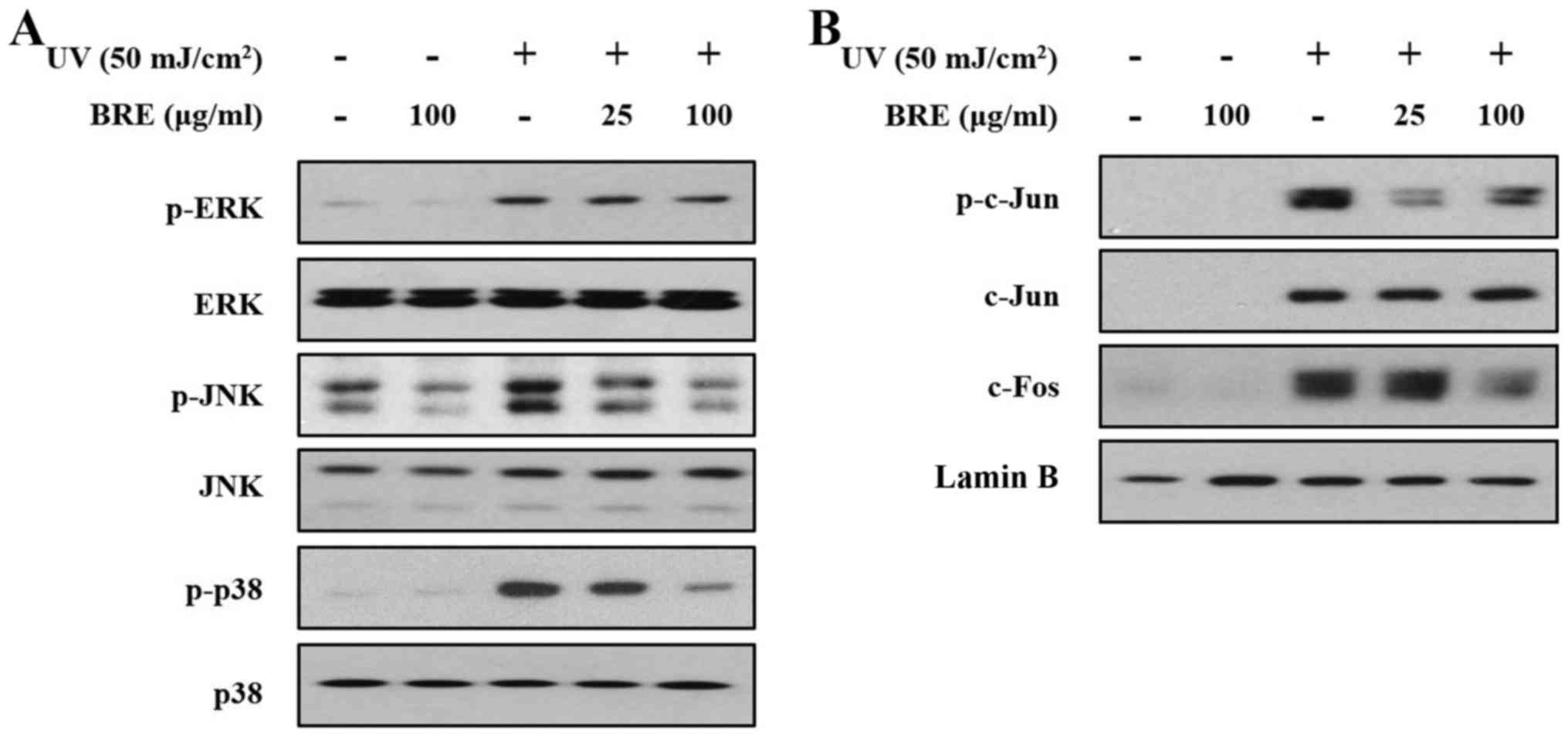 | Figure 4The effect of BRE on UV-induced MAPK
signaling, and AP-1 (c-Jun/c-Fos) activity in HaCaT cells. The
effects of BRE on UV-induced MAPK signaling, and AP-1 activity were
examined. The HaCaT cells were post-treated with the indicated
concentration of BRE for 15 min, with or without UV irradiation (50
mJ/cm2). (A) The phosphorylation status of ERK, JNK, and
p38 was then assessed using western blot analysis. (B)
UV-irradiated or non-irradiated HaCaT cells were also post-treated
with BRE for 4 h, and the nuclear levels of phospho-c-Jun, c-Jun
and c-Fos were analyzed. The blots shown are representative images
of three independent experiments. BRE, black rice extract; UV,
ultraviolet; MAPK, mitogen-activated protein kinase; AP-1,
activator protein-1; ERK, extracellular signal-regulated kinase;
JNK, c-Jun N-terminal kinase. |
AP-1 is a transcription factor composed of c-Jun and
c-Fos subunits (10,25). Next, we analyzed c-Jun
phosphorylation and c-Fos expression in nuclear extracts from the
UV-irradiated HaCaT cells. The cells were irradiated with UV, and
then incubated for 4 h with the indicated concentrations of BRE.
Although UV exposure increased c-Jun and c-Fos expression, and
c-Jun phosphorylation, BRE treatment suppressed c-Jun
phosphorylation, and c-Fos expression in the nucleus (Fig. 4B).
BRE reduces UV-induced intracellular ROS
generation
To determine whether BRE can reduce UV-induced ROS
production, HaCaT cells were pre-treated with BRE or NAC for 1 h
before UV irradiation (50 mJ/cm2). The intracellular ROS
levels measured using DCF-DA assay were 82.0±7.6% and 73.8±8.0% of
the control value, following treatment with 25 and 100 μg/ml
BRE, respectively. Furthermore, the UV-irradiated cells displayed
dramatically increased ROS generation (234.0±9.6%) when compared
with the control, whereas treatment with 25 and 100 μg/ml
BRE significantly attenuated the intracellular ROS level to
183.0±10.2% and 169.2±5.9%, respectively (Fig. 5A). Under the same experimental
conditions, a decrease in the ROS level following BRE treatment was
also observed using fluorescence microscopy (Fig. 5B). Analysis of the components in
BRE using the optimized UPLC-Q-TOF MS method showed that
polysaccharides such as isomaltose and mannotriose were detected at
0.74 min and lysophosphatidylcholine (LysoPC) components at 4.83,
5.11 and 5.22 min. The flavonoid components,
cyanidin-3-O-β-D-glycoside (C3G), known as an anthocyanin and
taxifolin-7-O-glucoside (T7G), were detected at 2.09 and 2.22 min,
respectively (Fig. 5C). These
results indicated that the photoprotective effect of BRE in skin
cells may be attributed to antioxidant effects possibly by
flavonoids.
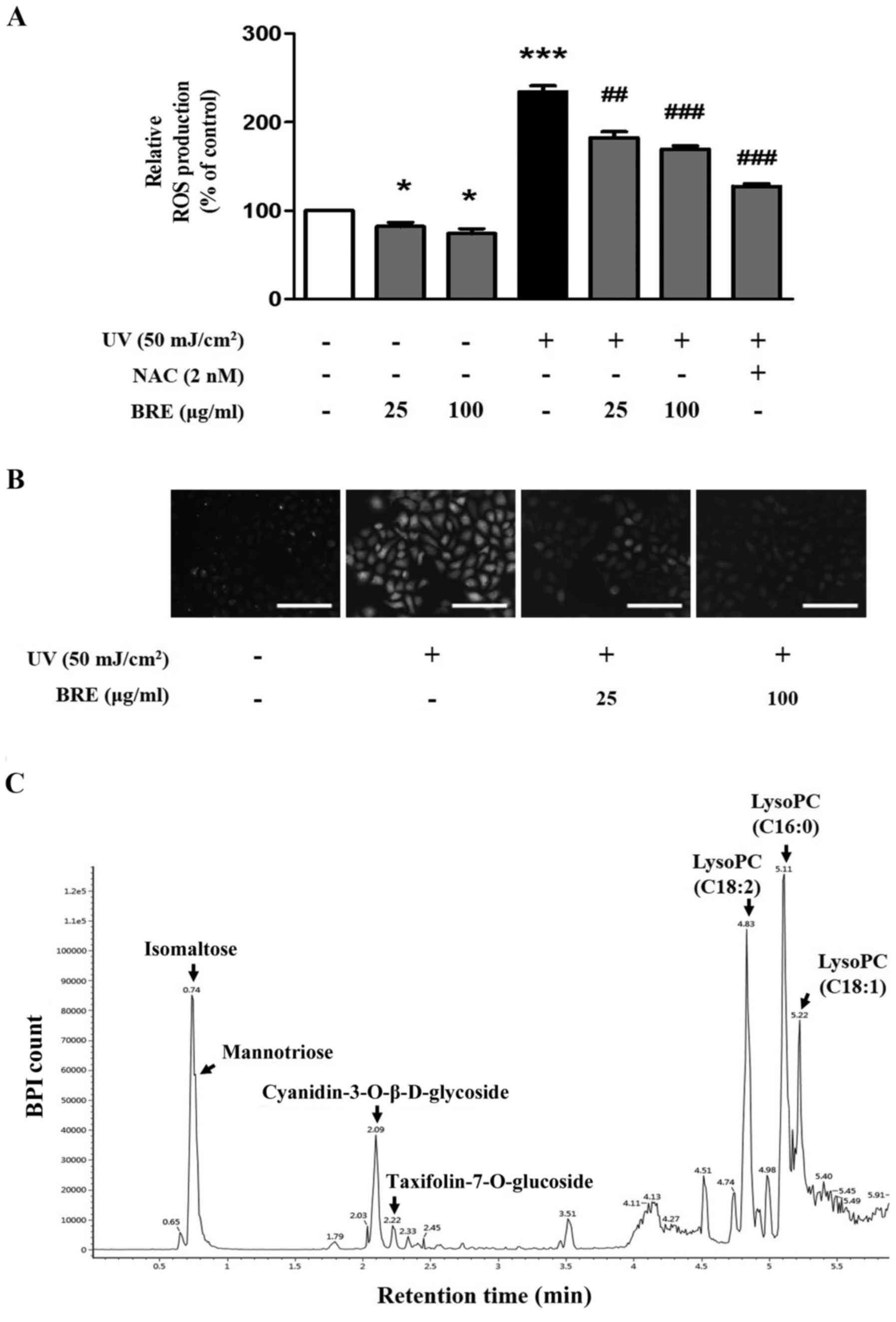 | Figure 5BRE reduced intracellular oxidative
stress in HaCaT cells. The cells were pre-treated with BRE or NAC
for 1 h before UV exposure, and the relative ROS levels were
analyzed using DCF-DA. (A) The difference in DCF fluorescence
generated from DCF-DA by intracellular ROS was measured using a
multilabel plate reader, and (B) florescence microscopy. (C) BRE
was analyzed using UPLC-Q-TOF MS profiles. The graph shows the mean
± SEM of at least three experiments. *P<0.05,
***P<0.001 compared with the non-irradiated control
cells. ##P<0.01, ###P<0.001 compared
with the UV-irradiated control cells. Scale bar indicates 200
μm. BRE, black rice extract; NAC, N-acetylcysteine;
UV, ultraviolet; ROS, reactive oxygen species, DCF-DA,
2′7′-dichlorofluorescein diacetate. |
Discussion
UV-induced DNA damage and ROS initiate various
changes in expression of genes, which are involved in inflammatory
responses that lead to pre-mature skin aging, known as 'photoaging'
(26,27). Therefore, the application of
antioxidants and/or anti-inflammatory compounds is thought to
enhance resistance to photoaging. Previous investigations have
suggested that BRE has antioxidative effects in HepG2 cells and
C57BL/6 mice, through the induction of superoxide dismutase and
catalase activity (13).
Furthermore, anthocyanins from black rice bran have been shown to
have ROS-scavenging activity in vitro (28). BRE also relieves skin inflammation
in a chemical-induced inflammatory mouse model (15). However, the precise effects and
underlying mechanisms of BRE in UV-induced responses in skin cells
are still unknown.
In the present study, we demonstrated that BRE has
potent photoprotective effects against UV-irradiated changes in
skin cells, possibly via ROS/MAPK signaling pathways. UV
irradiation induces MMP expression, which leads to the breakdown of
collagen, a dominant component of connective tissue (29,30). We demonstrated that BRE inhibits
UV-induced MMP-1 expression in HaCaT cells, and MMP-1 and MMP-3
expression in HDF.
In the skin, collagen is necessary to maintain
resiliency and strength, and decreased collagen expression is
observed in photoaged skin (31,32). In the present study, UV-exposed
HDF showed a reduced level of type I procollagen, which was
recovered by BRE treatment. Our findings indicate that BRE could
prevent or rescue UV-induced ECM alterations via a dual pathway
involving inhibition of MMP expression and increase of procollagen
expression.
Oxidative stress induced by UV irradiation is one of
the pivotal triggers of MMP-1 and MMP-3 upregulation by both the
AP-1 dependent and independent (p38 activity) pathways in HDF
(33). Thus, reducing ROS levels
could be an effective strategy to prevent photo-damage (26,27,34). In the present study, BRE
significantly reduced intracellular ROS generation in HaCaT cells.
In addition, BRE inhibited UV-induced AP-1 activity (c-Jun/c-Fos),
and MAPK (JNK/p38) signaling, possibly owing to its antioxidative
effects.
Botanical phenolic acids and flavonoids exert
antioxidant effects and these are commonly used in the prevention
of photoaging (35,36). Black rice contains abundant
flavonoids and polyphenols, including cyanidin-3-O-glycoside
chloride (C3G), peonidin-3-O-glucoside chloride, and ferulic acid
(13,37). C3G is a typical anthocyanin, which
shows anti-oxidative effects in various conditions (38–40). In addition, taxifolin has been
reported to be detected in black rice bran (41) and has antioxidant properties
(42,43). UPLC-Q-TOF MS result showed that
BRE here contains considerable amounts of C3G and T7G. Therefore,
the antioxidative and protective effect of BRE on UV-irradiated
skin cells are thought to be attributed to flavonoids, such as C3G
or T7G in BRE.
One of the most important protective mechanisms of
flavonoids is their antioxidative ability through the activation of
Nrf2-dependent genes. Under normal physiological conditions,
inactive Nrf2 is located in the cytoplasm with its inhibitor Keap1.
Under high ROS conditions, Nrf2 is released from Keap1 and moved
into the nucleus, regulating the expression of downstream target
genes via antioxidant response element (ARE) binding sites in the
promoter (44). Flavonoids are
potent inducers of the ARE/Nrf2/Keap1 signaling pathway, and the
anthocyanin is the most prominent among the flavonoids (45,46). Thus, the antioxidant activity
through Nrf2 activation by anthocyanin contained in BRE is also a
possible mechanism.
In conclusion, BRE attenuates markers of photoaging
including reduction of collagen and increases of MMPs in skin
cells. The underlying mechanism involved in these beneficial
effects could be the inhibition of ROS generation and AP-1
activation in vitro. Further study employing proteomic
analysis by mass spectroscopy is warranted to determine other
factors in addition to MMP-1 and collagen. Additionally, if
stability, safety, and efficacy are demonstrated in human clinical
studies, BRE could be used as a cosmetic ingredient for preventing
photoaging of the skin.
Acknowledgments
Not applicable.
Notes
[1]
Funding
This research was supported by a grant from the
Korea Health Technology R&D Project through the Korea Health
Industry Development Institute (KHIDI), funded by the Ministry of
Health and Welfare, Republic of Korea (grant nos. HI12C1723 and
HI14C1277).
[2] Availability
of data and material
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
[3] Authors'
contributions
MH, JSB and DHL designed the study and performed the
experiments. HSS provided the black rice extract and analyzed
UPLC-Q-TOP. JJB performed the cell viability assay and was involved
in drafting the article. JHC had full access to all the data, and
takes full responsibility for the integrity of data and the
accuracy of data analysis. All authors read and approved the final
manuscript.
[4] Ethics
approval and consent to participate
Not applicable.
[5] Consent for
publication
Not applicable.
[6] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Bikle DD: The vitamin D receptor: A tumor
suppressor in skin. Adv Exp Med Biol. 810:282–302. 2014.PubMed/NCBI
|
|
2
|
Berneburg M, Plettenberg H and Krutmann J:
Photoaging of human skin. Photodermatol Photoimmunol Photomed.
16:239–244. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ichihashi M, Ueda M, Budiyanto A, Bito T,
Oka M, Fukunaga M, Tsuru K and Horikawa T: UV-induced skin damage.
Toxicology. 189:21–39. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yamazaki S, Nishioka A, Kasuya S, Ohkura
N, Hemmi H, Kaisho T, Taguchi O, Sakaguchi S and Morita A:
Homeostasis of thymus-derived Foxp3+ regulatory T cells is
controlled by ultraviolet B exposure in the skin. J Immunol.
193:5488–5497. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Brenneisen P, Sies H and
Scharffetter-Kochanek K: Ultraviolet-B irradiation and matrix
metalloproteinases: From induction via signaling to initial events.
Ann N Y Acad Sci. 973:31–43. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fisher GJ, Wang ZQ, Datta SC, Varani J,
Kang S and Voorhees JJ: Pathophysiology of premature skin aging
induced by ultraviolet light. N Engl J Med. 337:1419–1428. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bald T, Quast T, Landsberg J, Rogava M,
Glodde N, Lopez-Ramos D, Kohlmeyer J, Riesenberg S, van den
Boorn-Konijnenberg D, Hömig-Hölzel C, et al:
Ultraviolet-radiation-induced inflammation promotes angiotropism
and metastasis in melanoma. Nature. 507:109–113. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bowden GT: Prevention of non-melanoma skin
cancer by targeting ultraviolet-B-light signalling. Nat Rev Cancer.
4:23–35. 2004. View
Article : Google Scholar
|
|
9
|
Rigel DS: Cutaneous ultraviolet exposure
and its relationship to the development of skin cancer. J Am Acad
Dermatol. 58(Suppl 2): S129–S132. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fisher GJ and Voorhees JJ: Molecular
mechanisms of photoaging and its prevention by retinoic acid:
Ultraviolet irradiation induces MAP kinase signal transduction
cascades that induce Ap-1-regulated matrix metalloproteinases that
degrade human skin in vivo. J Investig Dermatol Symp Proc. 3:61–68.
1998.PubMed/NCBI
|
|
11
|
Angel P, Szabowski A and Schorpp-Kistner
M: Function and regulation of AP-1 subunits in skin physiology and
pathology. Oncogene. 20:2413–2423. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang H, Shao Y, Bao J and Beta T:
Phenolic compounds and antioxidant properties of breeding lines
between the white and black rice. Food Chem. 172:630–639. 2015.
View Article : Google Scholar
|
|
13
|
Chiang AN, Wu HL, Yeh HI, Chu CS, Lin HC
and Lee WC: Antioxidant effects of black rice extract through the
induction of superoxide dismutase and catalase activities. Lipids.
41:797–803. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang Q, Han P, Zhang M, Xia M, Zhu H, Ma
J, Hou M, Tang Z and Ling W: Supplementation of black rice pigment
fraction improves antioxidant and anti-inflammatory status in
patients with coronary heart disease. Asia Pac J Clin Nutr.
16(Suppl 1): 295–301. 2007.PubMed/NCBI
|
|
15
|
Choi SP, Kim SP, Kang MY, Nam SH and
Friedman M: Protective effects of black rice bran against
chemically-induced inflammation of mouse skin. J Agric Food Chem.
58:10007–10015. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xia X, Ling W, Ma J, Xia M, Hou M, Wang Q,
Zhu H and Tang Z: An anthocyanin-rich extract from black rice
enhances atherosclerotic plaque stabilization in apolipoprotein
E-deficient mice. J Nutr. 136:2220–2225. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen PN, Kuo WH, Chiang CL, Chiou HL,
Hsieh YS and Chu SC: Black rice anthocyanins inhibit cancer cells
invasion via repressions of MMPs and u-PA expression. Chem Biol
Interact. 163:218–229. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fan MJ, Wang IC, Hsiao YT, Lin HY, Tang
NY, Hung TC, Quan C, Lien JC and Chung JG: Anthocyanins from black
rice (Oryza sativa L.) demonstrate antimetastatic properties by
reducing MMPs and NF-κB expressions in human oral cancer CAL 27
cells. Nutr Cancer. 67:327–338. 2015. View Article : Google Scholar
|
|
19
|
Choi SP, Kim SP, Nam SH and Friedman M:
Antitumor effects of dietary black and brown rice brans in
tumor-bearing mice: Relationship to composition. Mol Nutr Food Res.
57:390–400. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hui C, Bin Y, Xiaoping Y, Long Y, Chunye
C, Mantian M and Wenhua L: Anticancer activities of an
anthocyanin-rich extract from black rice against breast cancer
cells in vitro and in vivo. Nutr Cancer. 62:1128–1136. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Miyazawa M, Oshima T, Koshio K, Itsuzaki Y
and Anzai J: Tyrosinase inhibitor from black rice bran. J Agric
Food Chem. 51:6953–6956. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sim GS, Lee DH, Kim JH, An SK, Choe TB,
Kwon TJ, Pyo HB and Lee BC: Black rice (Oryza sativa L. var.
japonica) hydrolyzed peptides induce expression of hyaluronan
synthase 2 gene in HaCaT keratinocytes. J Microbiol Biotechnol.
17:271–279. 2007.PubMed/NCBI
|
|
23
|
Jin CL, Oh JH, Han M, Shin MK, Yao C, Park
CH, Jin ZH and Chung JH: UV irradiation-induced production of
monoglycosylated biglycan through downregulation of
xylosyltransferase 1 in cultured human dermal fibroblasts. J
Dermatol Sci. 79:20–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bae JS, Han M, Yao C and Chung JH:
Chaetocin inhibits IBMX-induced melanogenesis in B16F10 mouse
melanoma cells through activation of ERK. Chem Biol Interact.
245:66–71. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Silvers AL, Bachelor MA and Bowden GT: The
role of JNK and p38 MAPK activities in UVA-induced signaling
pathways leading to AP-1 activation and c-Fos expression.
Neoplasia. 5:319–329. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Natarajan VT, Ganju P, Ramkumar A, Grover
R and Gokhale RS: Multifaceted pathways protect human skin from UV
radiation. Nat Chem Biol. 10:542–551. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pillai S, Oresajo C and Hayward J:
Ultraviolet radiation and skin aging: Roles of reactive oxygen
species, inflammation and protease activation, and strategies for
prevention of inflammation-induced matrix degradation - a review.
Int J Cosmet Sci. 27:17–34. 2005. View Article : Google Scholar
|
|
28
|
Kaneda I, Kubo F and Sakurai H:
Antioxidative compounds in the extracts of black rice brans. J
Health Sci. 52:495–511. 2006. View Article : Google Scholar
|
|
29
|
Jabłońska-Trypuć A, Matejczyk M and
Rosochacki S: Matrix metalloproteinases (MMPs), the main
extracellular matrix (ECM) enzymes in collagen degradation, as a
target for anticancer drugs. J Enzyme Inhib Med Chem. 31:177–183.
2016. View Article : Google Scholar
|
|
30
|
Quan T, Qin Z, Xia W, Shao Y, Voorhees JJ
and Fisher GJ: Matrix-degrading metalloproteinases in photoaging. J
Investig Dermatol Symp Proc. 14:20–24. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fisher GJ, Datta S, Wang Z, Li XY, Quan T,
Chung JH, Kang S and Voorhees JJ: c-Jun-dependent inhibition of
cutaneous procollagen transcription following ultraviolet
irradiation is reversed by all-trans retinoic acid. J Clin Invest.
106:663–670. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fisher GJ, Kang S, Varani J, Bata-Csorgo
Z, Wan Y, Datta S and Voorhees JJ: Mechanisms of photoaging and
chronological skin aging. Arch Dermatol. 138:1462–1470. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Reunanen N, Li SP, Ahonen M, Foschi M, Han
J and Kähäri VM: Activation of p38 alpha MAPK enhances
collagenase-1 (matrix metalloproteinase (MMP)-1) and stromelysin-1
(MMP-3) expression by mRNA stabilization. J Biol Chem.
277:32360–32368. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shin MH, Rhie GE, Kim YK, Park CH, Cho KH,
Kim KH, Eun HC and Chung JH: H2O2
accumulation by catalase reduction changes MAP kinase signaling in
aged human skin in vivo. J Invest Dermatol. 125:221–229. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jadoon S, Karim S, Bin Asad MH, Akram MR,
Khan AK, Malik A, Chen C and Murtaza G: Anti-aging potential of
phytoextract loaded-pharmaceutical creams for human skin cell
longetivity. Oxid Med Cell Longev. 2015:7096282015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nichols JA and Katiyar SK: Skin
photoprotection by natural polyphenols: Anti-inflammatory,
antioxidant and DNA repair mechanisms. Arch Dermatol Res.
302:71–83. 2010. View Article : Google Scholar
|
|
37
|
Sumczynski D, Kotásková E, Družbíková H
and Mlček J: Determination of contents and antioxidant activity of
free and bound phenolics compounds and in vitro digestibility of
commercial black and red rice (Oryza sativa L.) varieties. Food
Chem. 211:339–346. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kim MK, Kim HA, Koh K, Kim HS, Lee YS and
Kim YH: Identification and quantification of anthocyanin pigments
in colored rice. Nutr Res Pract. 2:46–49. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Min B, McClung AM and Chen MH:
Phytochemicals and antioxidant capacities in rice brans of
different color. J Food Sci. 76:C117–C126. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang MW, Zhang RF, Zhang FX and Liu RH:
Phenolic profiles and antioxidant activity of black rice bran of
different commercially available varieties. J Agric Food Chem.
58:7580–7587. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sriseadka T, Wongpornchai S and Rayanakorn
M: Quantification of flavonoids in black rice by liquid
chromatography-negative electrospray ionization tandem mass
spectrometry. J Agric Food Chem. 60:11723–11732. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Rice-Evans CA, Miller NJ and Paganga G:
Structure-antioxidant activity relationships of flavonoids and
phenolic acids. Free Radic Biol Med. 20:933–956. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Sun X, Chen RC, Yang ZH, Sun GB, Wang M,
Ma XJ, Yang LJ and Sun XB: Taxifolin prevents diabetic
cardiomyopathy in vivo and in vitro by inhibition of oxidative
stress and cell apoptosis. Food Chem Toxicol. 63:221–232. 2014.
View Article : Google Scholar
|
|
44
|
Surh YJ, Kundu JK and Na HK: Nrf2 as a
master redox switch in turning on the cellular signaling involved
in the induction of cytoprotective genes by some chemopreventive
phytochemicals. Planta Med. 74:1526–1539. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ji LL, Sheng YC, Zheng ZY, Shi L and Wang
ZT: The involvement of p62-Keap1-Nrf2 antioxidative signaling
pathway and JNK in the protection of natural flavonoid quercetin
against hepatotoxicity. Free Radic Biol Med. 85:12–23. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hwang YP, Choi JH, Yun HJ, Han EH, Kim HG,
Kim JY, Park BH, Khanal T, Choi JM, Chung YC and Jeong HG:
Anthocyanins from purple sweet potato attenuate
dimethylnitrosamine-induced liver injury in rats by inducing
Nrf2-mediated antioxidant enzymes and reducing COX-2 and iNOS
expression. Food Chem Toxicol. 49:93–99. 2011. View Article : Google Scholar
|